

Development of a High-Resolution Melting Method for the Detection of Clarithromycin-Resistant Helicobacter pylori in the Gastric Microbiome

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
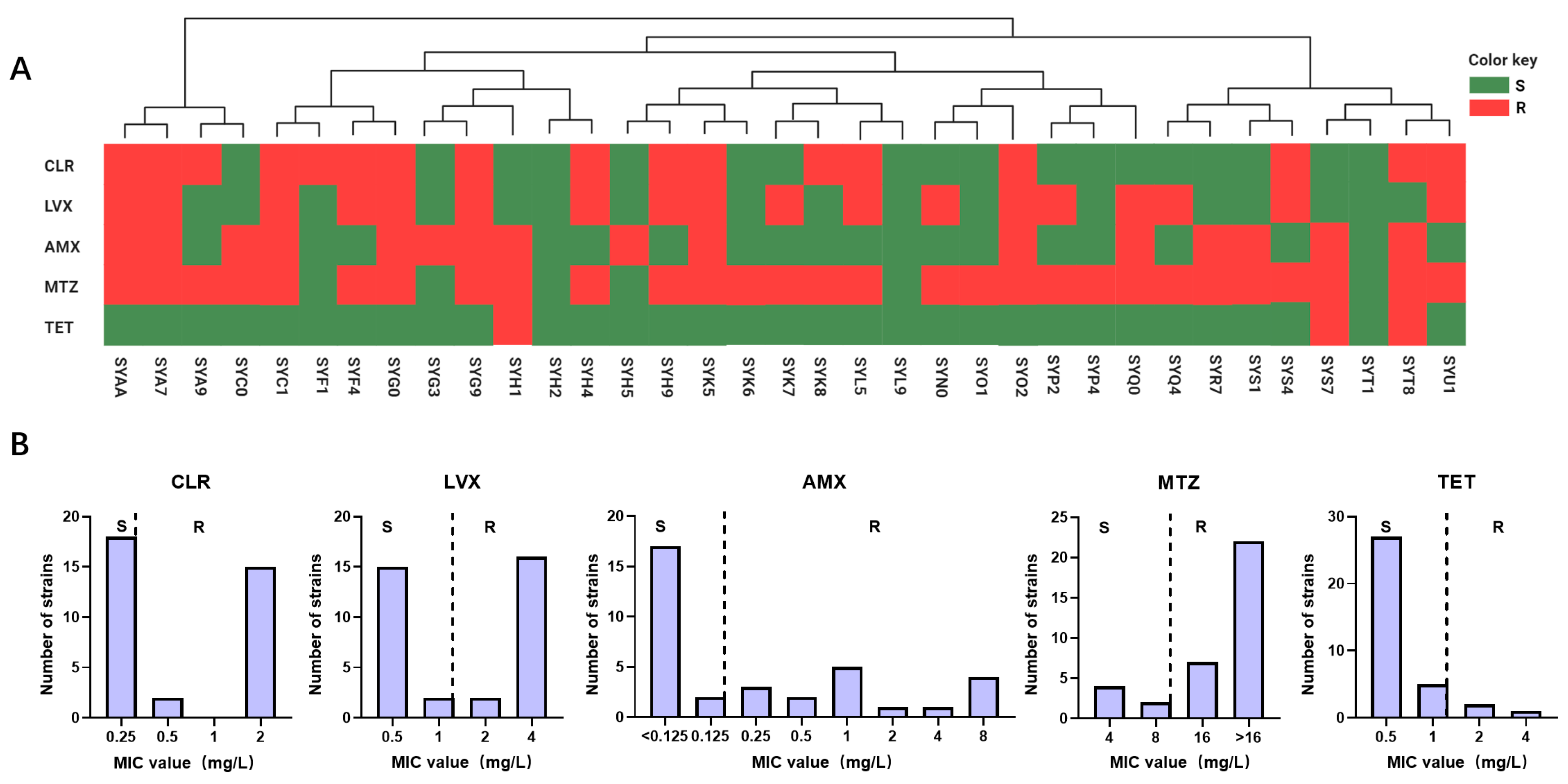
2.1. Antibiotic Resistance Profile of H. pylori
2.2. Discovery of the CLR-Resistant Mutations
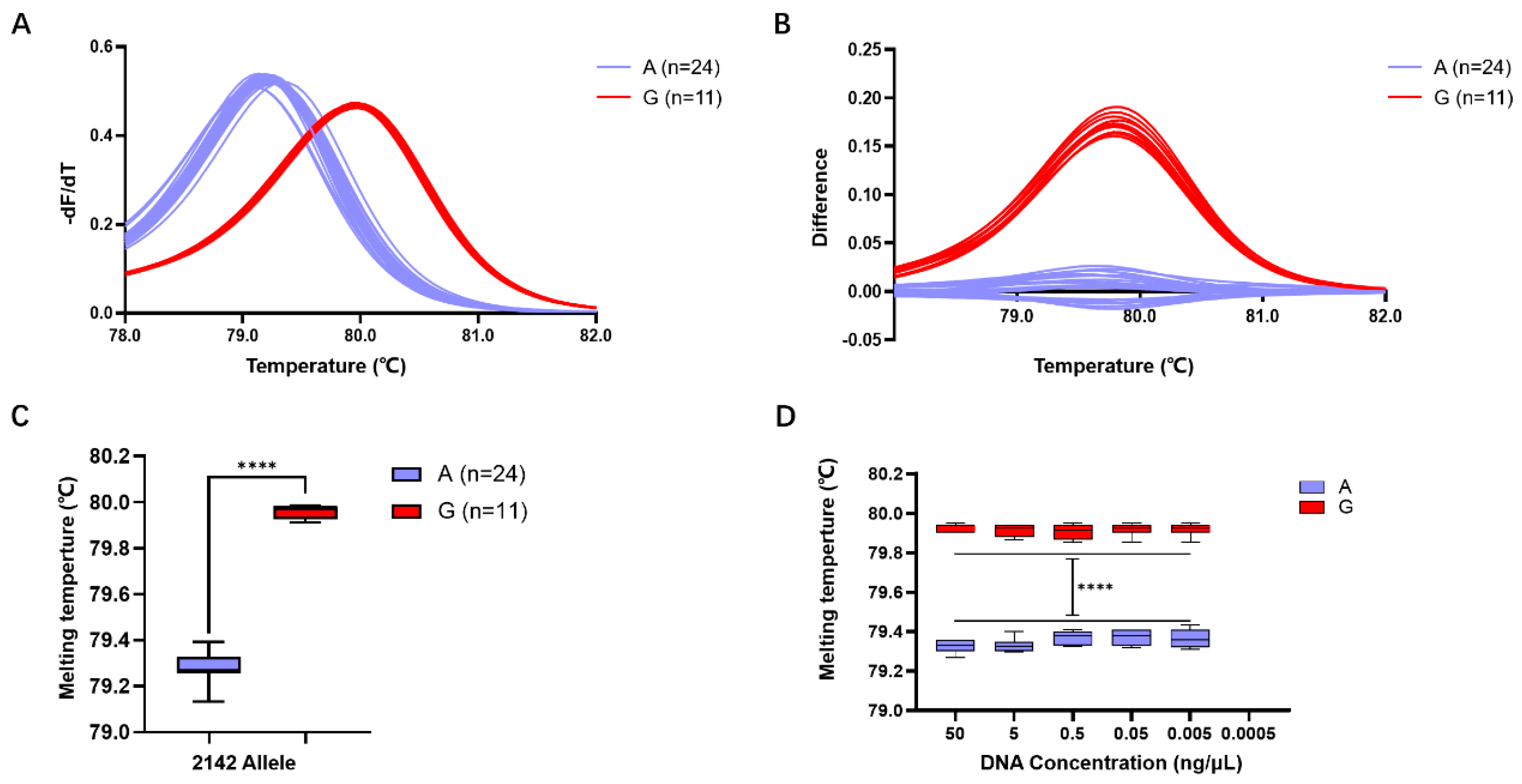
2.3. Evaluation of HRM Detection Efficiency for the A2143G Mutation
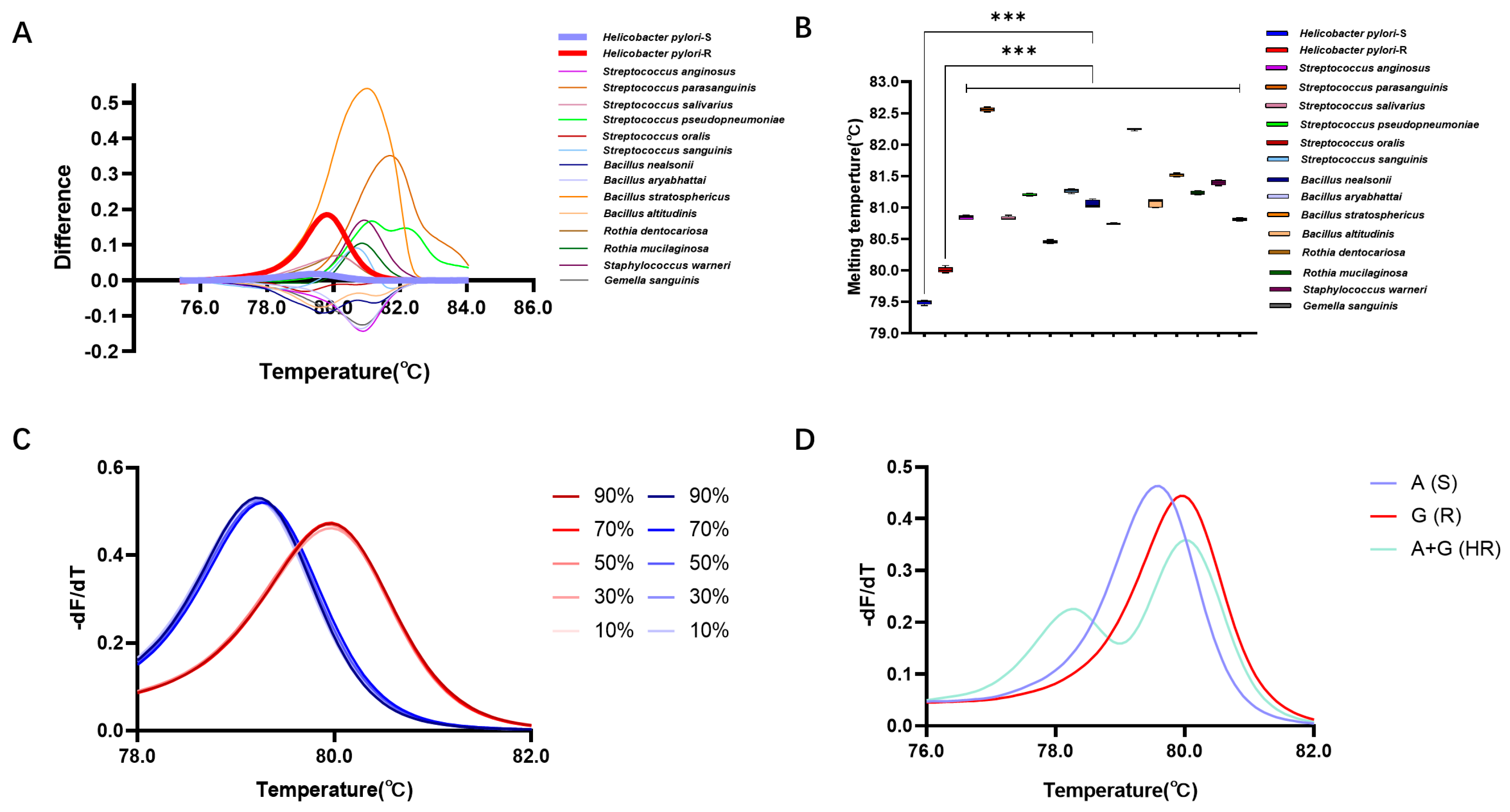
2.4. Detection Performance of the HRM System in the Gastric Microbiome
2.5. Detection Efficiency of the HRM System in the Prospective Clinical Cohort
3. Discussion
4. Materials and Methods
4.1. Study Cohorts and Sample Collection
4.2. Isolation and Identification of the Gastric Microbes
4.3. Phenotypic AST of H. Pylori
4.4. Sequencing
4.4.1. Whole-Genome Sequencing (WGS) and Genome Assembly of H. pylori
4.4.2. Gastric Microbiome Analysis
4.5. Bioinformatic Analysis of the AMR Genetic Determinants
4.6. Primer Design and HRM Analysis
4.7. Verification of the Detection Efficiency of the HRM System
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schistosomes, Liver Flukes and Helicobacter Pylori. In IARC Working Group on the Evaluation of Carcinogenic Risks to Humans; International Agency for Research on Cancer: Lyon, France, 1994; Volume 61, pp. 1–241.
- Lunn, R.M.; Mehta, S.S.; Jahnke, G.D.; Wang, A.; Wolfe, M.S.; Berridge, B.R. Cancer Hazard Evaluations for Contemporary Needs: Highlights From New National Toxicology Program Evaluations and Methodological Advancements. JNCI J. Natl. Cancer Inst. 2022, 114, 1441–1448. [Google Scholar] [CrossRef] [PubMed]
- Shah, S.C.; Camargo, M.C.; Lamm, M.; Bustamante, R.; Roumie, C.L.; Wilson, O.; Halvorson, A.E.; Greevy, R.; Liu, L.; Gupta, S.; et al. Impact of Helicobacter Pylori Infection and Treatment on Colorectal Cancer in a Large, Nationwide Cohort. J. Clin. Oncol. 2024, 42, 1881–1889. [Google Scholar] [CrossRef] [PubMed]
- Mohammadzadeh, R.; Soleimanpour, S.; Pishdadian, A.; Farsiani, H. Designing and Development of Epitope-Based Vaccines against Helicobacter Pylori. Crit. Rev. Microbiol. 2022, 48, 489–512. [Google Scholar] [CrossRef] [PubMed]
- Gebreslassie Kasahun, G.; Demoz, G.T.; Mahari Desta, D. Primary Resistance Pattern of Helicobacter Pylori to Antibiotics in Adult Population: A Systematic Review. Infect. Drug Resist. 2020, 13, 1567–1573. [Google Scholar] [CrossRef] [PubMed]
- Zagari, R.M.; Dajti, E.; Cominardi, A.; Frazzoni, L.; Fuccio, L.; Eusebi, L.H.; Vestito, A.; Lisotti, A.; Galloro, G.; Romano, M.; et al. Standard Bismuth Quadruple Therapy versus Concomitant Therapy for the First-Line Treatment of Helicobacter Pylori Infection: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Clin. Med. 2023, 12, 3258. [Google Scholar] [CrossRef]
- Savoldi, A.; Carrara, E.; Graham, D.Y.; Conti, M.; Tacconelli, E. Prevalence of Antibiotic Resistance in Helicobacter Pylori: A Systematic Review and Meta-Analysis in World Health Organization Regions. Gastroenterology 2018, 155, 1372–1382.e17. [Google Scholar] [CrossRef]
- Malfertheiner, P.; Megraud, F.; Rokkas, T.; Gisbert, J.P.; Liou, J.-M.; Schulz, C.; Gasbarrini, A.; Hunt, R.H.; Leja, M.; O’Morain, C.; et al. Management of Helicobacter Pylori Infection: The Maastricht VI/Florence Consensus Report. Gut 2022, 71, 1724–1762. [Google Scholar] [CrossRef]
- Sugano, K.; Tack, J.; Kuipers, E.J.; Graham, D.Y.; El-Omar, E.M.; Miura, S.; Haruma, K.; Asaka, M.; Uemura, N.; Malfertheiner, P. Kyoto Global Consensus Report on Helicobacter pylori Gastritis. Gut 2015, 64, 1353–1367. [Google Scholar] [CrossRef]
- Tshibangu-Kabamba, E.; Yamaoka, Y. Helicobacter Pylori Infection and Antibiotic Resistance—From Biology to Clinical Implications. Nat. Rev. Gastroenterol. Hepatol. 2021, 18, 613–629. [Google Scholar] [CrossRef]
- Binh, T.T.; Shiota, S.; Suzuki, R.; Matsuda, M.; Trang, T.T.H.; Kwon, D.H.; Iwatani, S.; Yamaoka, Y. Discovery of Novel Mutations for Clarithromycin Resistance in Helicobacter Pylori by Using Next-Generation Sequencing. J. Antimicrob. Chemother. 2014, 69, 1796–1803. [Google Scholar] [CrossRef]
- Medakina, I.; Tsapkova, L.; Polyakova, V.; Nikolaev, S.; Yanova, T.; Dekhnich, N.; Khatkov, I.; Bordin, D.; Bodunova, N. Helicobacter Pylori Antibiotic Resistance: Molecular Basis and Diagnostic Methods. Int. J. Mol. Sci. 2023, 24, 9433. [Google Scholar] [CrossRef] [PubMed]
- Kaya, A.D.; Öztürk, C.E.; Akcan, Y.; Behçet, M.; Karakoç, A.E.; Yücel, M.; Mısırlıoglu, M.; Tuncer, S. Prevalence of Helicobacter Pylori in Symptomatic Patients and Detection of Clarithromycin Resistance Using Melting Curve Analysis. Curr. Ther. Res. 2007, 68, 151–160. [Google Scholar] [CrossRef] [PubMed]
- Ohshima, C.; Takahashi, H.; Iwakawa, A.; Kuda, T.; Kimura, B. A Novel Typing Method for Listeria Monocytogenes Using High-Resolution Melting Analysis (HRMA) of Tandem Repeat Regions. Int. J. Food Microbiol. 2017, 253, 36–42. [Google Scholar] [CrossRef]
- Zhou, H.; Zhang, J.; Shao, Y.; Wang, J.; Xu, W.; Liu, Y.; Yu, S.; Ye, Q.; Pang, R.; Wu, S.; et al. Development of a High Resolution Melting Method Based on a Novel Molecular Target for Discrimination between Bacillus Cereus and Bacillus Thuringiensis. Food Res. Int. 2022, 151, 110845. [Google Scholar] [CrossRef]
- Usui, Y.; Taniyama, Y.; Endo, M.; Koyanagi, Y.N.; Kasugai, Y.; Oze, I.; Ito, H.; Imoto, I.; Tanaka, T.; Tajika, M.; et al. Helicobacter Pylori, Homologous-Recombination Genes, and Gastric Cancer. N. Engl. J. Med. 2023, 389, 379–381. [Google Scholar] [CrossRef]
- Sharafutdinov, I.; Tegtmeyer, N.; Linz, B.; Rohde, M.; Vieth, M.; Tay, A.C.-Y.; Lamichhane, B.; Tuan, V.P.; Fauzia, K.A.; Sticht, H.; et al. A Single-Nucleotide Polymorphism in Helicobacter Pylori Promotes Gastric Cancer Development. Cell Host Microbe 2023, 31, 1345–1358.e6. [Google Scholar] [CrossRef]
- Li, Y.; Huang, Z.; Shang, Y.; Xie, X.; Yang, R.; Chen, H.; Wang, Z.; Xue, L.; Pang, R.; Zhang, J.; et al. Exploration of the Molecular Mechanisms Underlying the Antibiotic Resistance of Helicobacter Pylori: A Whole-genome Sequencing-based Study in Southern China. Helicobacter 2022, 27, e12879. [Google Scholar] [CrossRef] [PubMed]
- Hooi, J.K.Y.; Lai, W.Y.; Ng, W.K.; Suen, M.M.Y.; Underwood, F.E.; Tanyingoh, D.; Malfertheiner, P.; Graham, D.Y.; Wong, V.W.S.; Wu, J.C.Y.; et al. Global Prevalence of Helicobacter Pylori Infection: Systematic Review and Meta-Analysis. Gastroenterology 2017, 153, 420–429. [Google Scholar] [CrossRef]
- Afzal, A. Molecular Diagnostic Technologies for COVID-19: Limitations and Challenges. J. Adv. Res. 2020, 26, 149–159. [Google Scholar] [CrossRef]
- Otto, C.; Chen, D. Point of Care Molecular Testing. Clin. Lab. Med. 2024, 44, 23–32. [Google Scholar] [CrossRef]
- Ren, X.; Shi, Y.; Suo, B.; Yao, X.; Lu, H.; Li, C.; Zhang, Y.; Zhou, L.; Tian, X.; Song, Z. Individualized Diagnosis and Eradication Therapy for Helicobacter Pylori Infection Based on Gene Detection of Clarithromycin Resistance in Stool Specimens: A Systematic Review and Meta-analysis. Helicobacter 2023, 28, e12958. [Google Scholar] [CrossRef]
- Eischeid, A.C. SYTO Dyes and EvaGreen Outperform SYBR Green in Real-Time PCR. BMC Res. Notes 2011, 4, 263. [Google Scholar] [CrossRef]
- Mezzasalma, V.; Ganopoulos, I.; Galimberti, A.; Cornara, L.; Ferri, E.; Labra, M. Poisonous or Non-Poisonous Plants? DNA-Based Tools and Applications for Accurate Identification. Int. J. Legal Med. 2017, 131, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Pereira, L.; Gomes, S.; Barrias, S.; Fernandes, J.R.; Martins-Lopes, P. Applying High-Resolution Melting (HRM) Technology to Olive Oil and Wine Authenticity. Food Res. Int. 2018, 103, 170–181. [Google Scholar] [CrossRef]
- Rychlik, W.; Spencer, W.J.; Rhoads, R.E. Optimization of the Annealing Temperature for DNA Amplification In Vitro. Nucleic Acids Res. 1990, 18, 6409–6412. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K. Specificity and Performance of PCR Detection Assays for Microbial Pathogens. Mol. Biotechnol. 2004, 26, 61–80. [Google Scholar] [CrossRef]
- Liu, W.; Lin, Y.; Jiang, J.; Zhang, J.; Liu, Q.; Hu, Q. Development of a 1-Step TaqMan Real-Time PCR Method for Detection of the Bovine Group A Rotavirus. Diagn. Microbiol. Infect. Dis. 2023, 107, 116081. [Google Scholar] [CrossRef] [PubMed]
- Muneeswaran, K.; Branavan, U.; De Silva, V.A.; Dayabandara, M.; Hanwella, R.; Chandrasekharan, N.V. Genotyping SNPs and Indels: A Method to Improve the Scope and Sensitivity of High-Resolution Melt (HRM) Analysis Based Applications. Clin. Chim. Acta 2024, 562, 119897. [Google Scholar] [CrossRef]
- Cai, Y.; Yuan, Y.; Lin, Q.; Chan, P. Allele-Specific Extension Allows Base-Pair Neutral Homozygotes to Be Discriminated by High-Resolution Melting of Small Amplicons. Anal. Biochem. 2010, 406, 29–33. [Google Scholar] [CrossRef]
- Aoki, A.; Adachi, H.; Mori, Y.; Ito, M.; Sato, K.; Kinoshita, M.; Kuriki, M.; Okuda, K.; Sakakibara, T.; Okamoto, Y.; et al. A Modified High-Resolution Melting-Based Assay (HRM) to Identify the SARS-CoV-2 N501Y Variant. J. Virol. Methods 2023, 314, 114678. [Google Scholar] [CrossRef]
- Nguyen, T.C.; Le, G.K.N.; Pham, D.T.H.; Pham, B.V.; Nguyen, L.T.H.; Che, T.H.; Nguyen, H.T.; Truong, D.Q.; Robert, A.; Bontems, P.; et al. Antibiotic Resistance and Heteroresistance in Helicobacter Pylori Isolates from Symptomatic Vietnamese Children: A Prospective Multicenter Study. Helicobacter 2023, 28, e13009. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Ye, Q.; Chen, M.; Zhou, B.; Xiang, X.; Wang, C.; Shang, Y.; Zhang, J.; Pang, R.; Wang, J.; et al. Mining of Novel Target Genes through Pan-Genome Analysis for Multiplex PCR Differentiation of the Major Listeria Monocytogenes Serotypes. Int. J. Food Microbiol. 2021, 339, 109026. [Google Scholar] [CrossRef] [PubMed]
- Yamaoka, Y.; Kodama, T.; Kita, M.; Imanishi, J.; Kashima, K.; Graham, D.Y. Relationship of vacA Genotypes of Helicobacter Pylori to cagA Status, Cytotoxin Production, and Clinical Outcome. Helicobacter 1998, 3, 241–253. [Google Scholar] [CrossRef] [PubMed]
- Garrity, G.M. A New Genomics-Driven Taxonomy of Bacteria and Archaea: Are We There Yet? J. Clin. Microbiol. 2016, 54, 1956–1963. [Google Scholar] [CrossRef] [PubMed]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e00213-21. [Google Scholar] [CrossRef]
- Alarcón, T.; Urruzuno, P.; Martínez, M.J.; Domingo, D.; Llorca, L.; Correa, A.; López-Brea, M. Antimicrobial Susceptibility of 6 Antimicrobial Agents in Helicobacter Pylori Clinical Isolates by Using EUCAST Breakpoints Compared with Previously Used Breakpoints. Enfermedades Infecc. Microbiol. Clínica 2017, 35, 278–282. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A Flexible Trimmer for Illumina Sequence Data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A New Genome Assembly Algorithm and Its Applications to Single-Cell Sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Fadrosh, D.W.; Ma, B.; Gajer, P.; Sengamalay, N.; Ott, S.; Brotman, R.M.; Ravel, J. An Improved Dual-Indexing Approach for Multiplexed 16S rRNA Gene Sequencing on the Illumina MiSeq Platform. Microbiome 2014, 2, 6. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid Prokaryotic Genome Annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Treangen, T.J.; Ondov, B.D.; Koren, S.; Phillippy, A.M. The Harvest Suite for Rapid Core-Genome Alignment and Visualization of Thousands of Intraspecific Microbial Genomes. Genome Biol. 2014, 15, 524. [Google Scholar] [CrossRef] [PubMed]


Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kuang, Z.; Huang, H.; Chen, L.; Shang, Y.; Huang, S.; Liu, J.; Chen, J.; Xie, X.; Chen, M.; Wu, L.; et al. Development of a High-Resolution Melting Method for the Detection of Clarithromycin-Resistant Helicobacter pylori in the Gastric Microbiome. Antibiotics 2024, 13, 975. https://doi.org/10.3390/antibiotics13100975
Kuang Z, Huang H, Chen L, Shang Y, Huang S, Liu J, Chen J, Xie X, Chen M, Wu L, et al. Development of a High-Resolution Melting Method for the Detection of Clarithromycin-Resistant Helicobacter pylori in the Gastric Microbiome. Antibiotics. 2024; 13(10):975. https://doi.org/10.3390/antibiotics13100975
Chicago/Turabian StyleKuang, Zupeng, Huishu Huang, Ling Chen, Yanyan Shang, Shixuan Huang, Jun Liu, Jianhui Chen, Xinqiang Xie, Moutong Chen, Lei Wu, and et al. 2024. "Development of a High-Resolution Melting Method for the Detection of Clarithromycin-Resistant Helicobacter pylori in the Gastric Microbiome" Antibiotics 13, no. 10: 975. https://doi.org/10.3390/antibiotics13100975
APA StyleKuang, Z., Huang, H., Chen, L., Shang, Y., Huang, S., Liu, J., Chen, J., Xie, X., Chen, M., Wu, L., Gao, H., Zhao, H., Li, Y., & Wu, Q. (2024). Development of a High-Resolution Melting Method for the Detection of Clarithromycin-Resistant Helicobacter pylori in the Gastric Microbiome. Antibiotics, 13(10), 975. https://doi.org/10.3390/antibiotics13100975