
Distribution of Extended-Spectrum-β-Lactamase-Producing Diarrheagenic Escherichia coli Clonal Complex 10 Isolates from Patients with Diarrhea in the Republic of Korea

 , and
, and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Isolation of CC10 Strains
2.2. Serotyping
2.3. Prevalence of Genomic Determinants of Antimicrobial Resistance
2.4. Transmission of Bla Gene
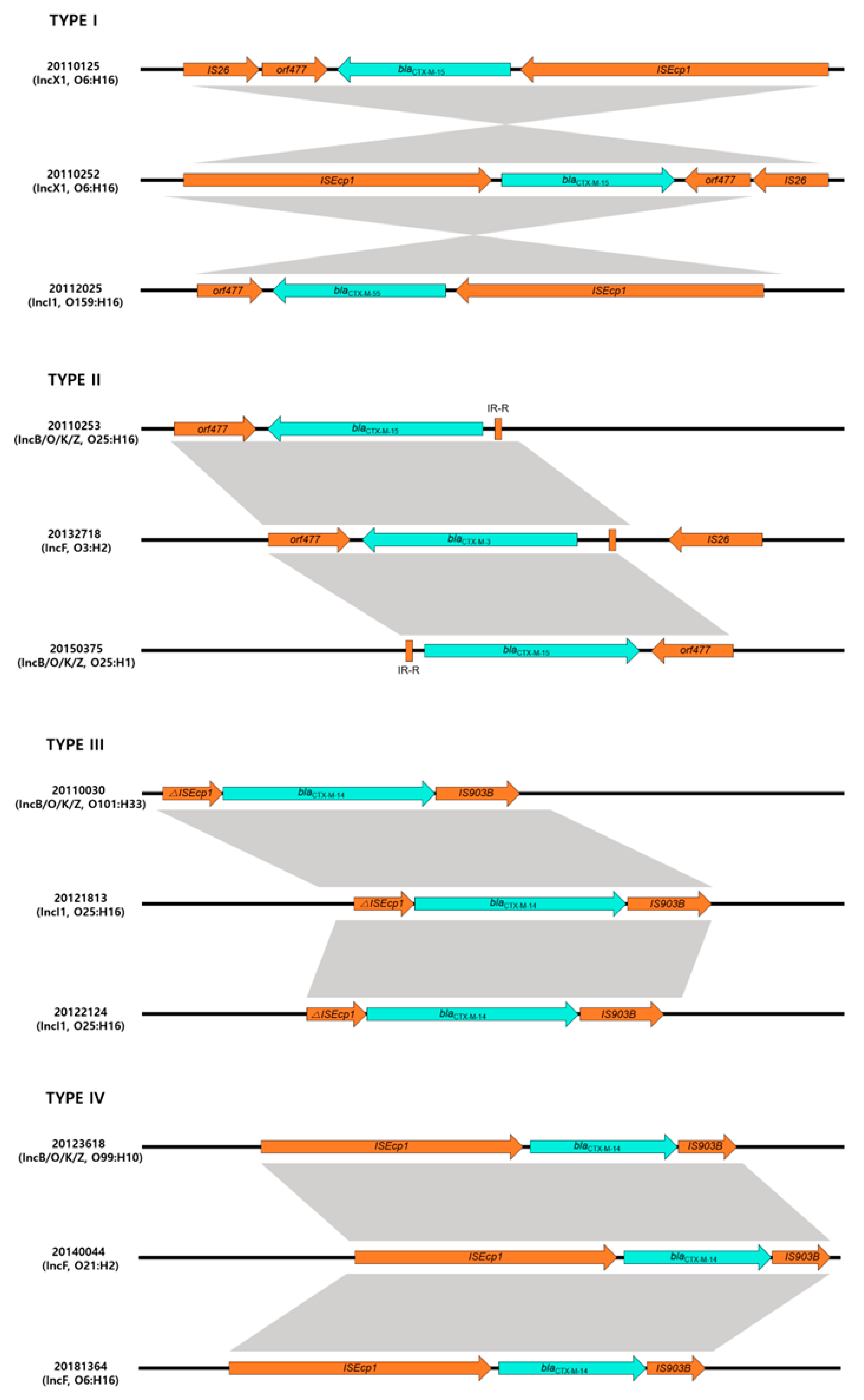
2.5. Analysis of the Regions Surrounding blaCTX-M Genes
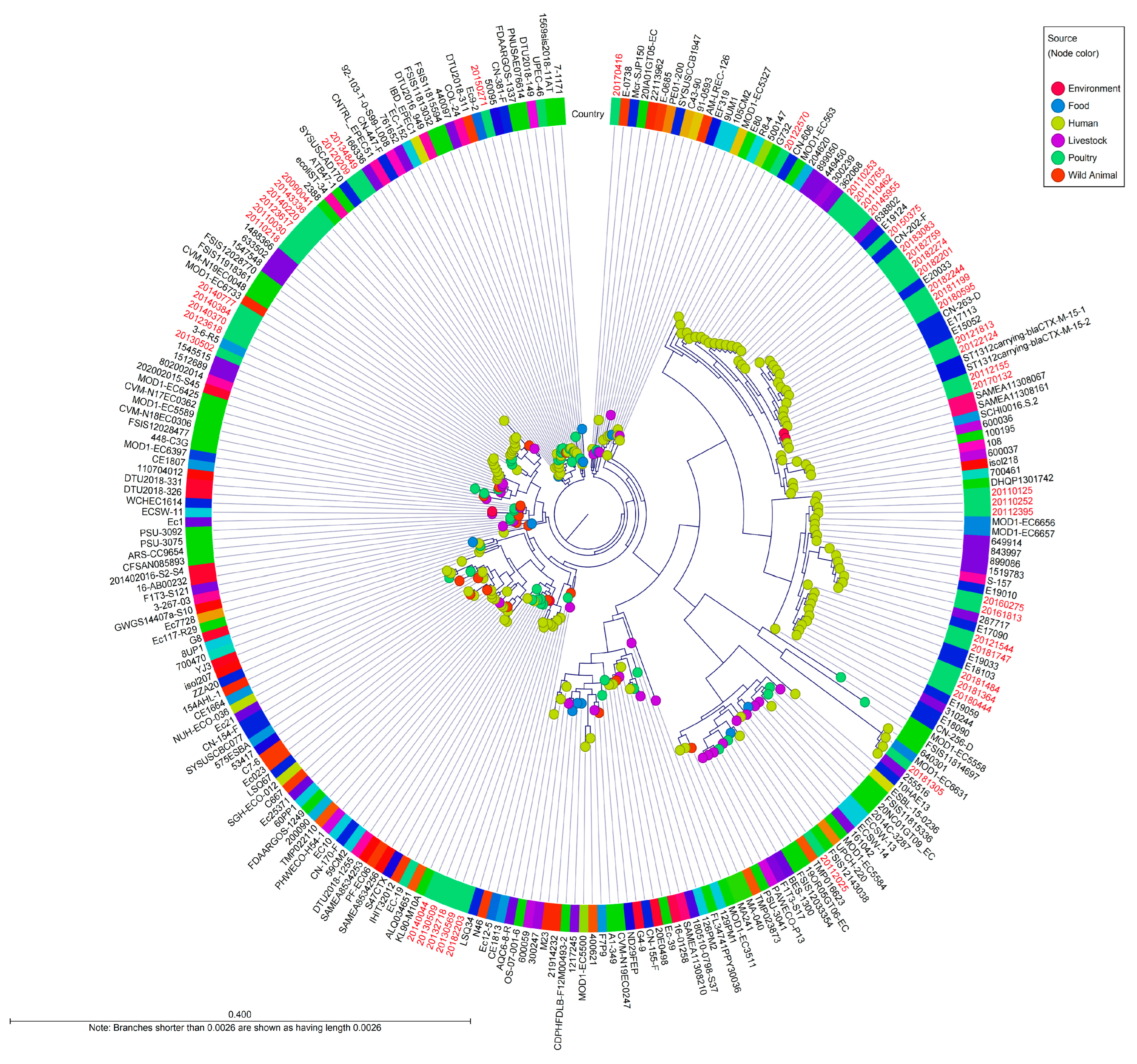
2.6. Whole-Genome SNPs-Based Phylogeny of CC10
3. Discussion
4. Materials and Methods
4.1. Isolation of CC10 Strains
4.2. Whole Genome Sequencing (WGS)
4.3. Data Analysis and Molecular Characterization
4.4. Plasmid Transfer by Bacterial Conjugation
4.5. Nucleotide Sequence Accession Numbers
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- CDC. Antibiotic Resistance Threats in The United States, 2019; U.S. Department of Health and Human Services, CDC: Atlanta, GA, USA, 2019. [CrossRef]
- Watkins, R.R.; Bonomo, R.A. Overview: Global and Local Impact of Antibiotic Resistance. Infect. Dis. Clin. N. Am. 2016, 30, 313–322. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, S.; Fatima, J.; Shakil, S.; Rizvi, S.M.D.; Kamal, M.A. Antibiotic Resistance and Extended Spectrum Beta-Lactamases: Types, Epidemiology and Treatment. Saudi J. Biol. Sci. 2015, 22, 90–101. [Google Scholar] [CrossRef] [PubMed]
- Malik, F.; Figueras, A. Continuous Rise in Cephalosporin and Fluoroquinolone Consumption in Pakistan: A 5 Year Analysis (2014–18). JAC Antimicrob. Resist. 2019, 1, dlz063. [Google Scholar] [CrossRef]
- Yu, F.; Chen, X.; Zheng, S.; Han, D.; Wang, Y.; Wang, R.; Wang, B.; Chen, Y. Prevalence and Genetic Diversity of Human Diarrheagenic Escherichia coli Isolates by Multilocus Sequence Typing. Int. J. Infect. Dis. 2018, 67, 7–13. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Y.; Guo, S.; Seow, K.L.G.; Ming, G.O.H.; Schlundt, J. Characterization of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli Isolates from Jurong Lake, Singapore with Whole-Genome-Sequencing. Int. J. Environ. Res. Public Health 2021, 18, 937. [Google Scholar] [CrossRef]
- Merino, I.; Hernández-García, M.; Turrientes, M.-C.; Pérez-Viso, B.; López-Fresneña, N.; Diaz-Agero, C.; Maechler, F.; Fankhauser-Rodriguez, C.; Kola, A.; Schrenzel, J.; et al. Emergence of ESBL-Producing Escherichia coli ST131-C1-M27 Clade Colonizing Patients in Europe. J. Antimicrob. Chemother. 2018, 73, 2973–2980. [Google Scholar] [CrossRef]
- Park, S.H.; Byun, J.-H.; Choi, S.-M.; Lee, D.-G.; Kim, S.-H.; Kwon, J.-C.; Park, C.; Choi, J.-H.; Yoo, J.-H. Molecular Epidemiology of Extended-Spectrum β-Lactamase-Producing Escherichia coli in the Community and Hospital in Korea: Emergence of ST131 Producing CTX-M-15. BMC Infect. Dis. 2012, 12, 149. [Google Scholar] [CrossRef]
- Kim, Y.A.; Kim, J.J.; Kim, H.; Lee, K. Community-Onset Extended-Spectrum-Beta-Lactamase-Producing Escherichia coli Sequence Type 131 at Two Korean Community Hospitals: The Spread of Multidrug-Resistant E. coli to the Community via Healthcare Facilities. Int. J. Infect. Dis. 2017, 54, 39–42. [Google Scholar] [CrossRef]
- Kim, S.-Y.; Park, Y.-J.; Johnson, J.R.; Yu, J.K.; Kim, Y.-K.; Kim, Y.S. Prevalence and Characteristics of Escherichia coli Sequence Type 131 and Its H30 and H30Rx Subclones: A Multicenter Study from Korea. Diagn. Microbiol. Infect. Dis. 2016, 84, 97–101. [Google Scholar] [CrossRef]
- Song, J.; Oh, S.-S.; Kim, J.; Park, S.; Shin, J. Clinically Relevant Extended-Spectrum β-Lactamase–Producing Escherichia coli Isolates from Food Animals in South Korea. Front. Microbiol. 2020, 11, 604. [Google Scholar] [CrossRef]
- Fagerström, A.; Mölling, P.; Khan, F.A.; Sundqvist, M.; Jass, J.; Söderquist, B. Comparative Distribution of Extended-Spectrum Beta-Lactamase-Producing Escherichia coli from Urine Infections and Environmental Waters. PLoS ONE 2019, 14, e0224861. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.; Li, Y.; Zuo, L.; Jiang, M.; Zhang, X.; Xie, L.; Luo, M.; She, Y.; Wang, L.; Jiang, Y.; et al. Genomic Epidemiology and Antimicrobial Susceptibility Profile of Enterotoxigenic Escherichia coli From Outpatients with Diarrhea in Shenzhen, China, 2015–2020. Front. Microbiol. 2021, 12, 732068. [Google Scholar] [CrossRef] [PubMed]
- Greig, D.R.; Jenkins, C.; Gharbia, S.; Dallman, T.J. Comparison of Single-Nucleotide Variants Identified by Illumina and Oxford Nanopore Technologies in the Context of a Potential Outbreak of Shiga Toxin-Producing Escherichia coli. GigaScience 2019, 8, giz104. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Sun, H.; Bai, X.; Fu, S.; Fan, R.; Xiong, Y. Occurrence of Multidrug-Resistant and ESBL-Producing Atypical Enteropathogenic Escherichia coli in China. Gut Pathog. 2018, 10, 8. [Google Scholar] [CrossRef] [PubMed]
- Song, J.; Oh, S.-S.; Kim, J.; Shin, J. Extended-Spectrum β-Lactamase-Producing Escherichia coli Isolated from Raw Vegetables in South Korea. Sci. Rep. 2020, 10, 19721. [Google Scholar] [CrossRef] [PubMed]
- Bartoloni, A.; Pallecchi, L.; Riccobono, E.; Mantella, A.; Magnelli, D.; Di Maggio, T.; Villagran, A.L.; Lara, Y.; Saavedra, C.; Strohmeyer, M.; et al. Relentless Increase of Resistance to Fluoroquinolones and Expanded-Spectrum Cephalosporins in Escherichia coli: 20 Years of Surveillance in Resource-Limited Settings from Latin America. Clin. Microbiol. Infect. 2013, 19, 356–361. [Google Scholar] [CrossRef]
- Lee, S.; An, J.U.; Guk, J.H.; Song, H.; Yi, S.; Kim, W.H.; Cho, S. Prevalence, Characteristics and Clonal Distribution of Extended-Spectrum β-Lactamase- and AmpC β-Lactamase-Producing Escherichia coli Following the Swine Production Stages, and Potential Risks to Humans. Front. Microbiol. 2021, 12, 710747. [Google Scholar] [CrossRef]
- Usein, C.-R.; Papagheorghe, R.; Oprea, M.; Condei, M.; Strãuţ, M. Molecular Characterization of Bacteremic Escherichia coli Isolates in Romania. Folia Microbiol. 2016, 61, 221–226. [Google Scholar] [CrossRef]
- Giufrè, M.; Graziani, C.; Accogli, M.; Luzzi, I.; Busani, L.; Cerquetti, M. Escherichia coli of Human and Avian Origin: Detection of Clonal Groups Associated with Fluoroquinolone and Multidrug Resistance in Italy. J. Antimicrob. Chemother. 2012, 67, 860–867. [Google Scholar] [CrossRef]
- Ramos, S.; Silva, N.; Dias, D.; Sousa, M.; Capelo-Martinez, J.L.; Brito, F.; Caniça, M.; Igrejas, G.; Poeta, P. Clonal Diversity of ESBL-Producing Escherichia coli in Pigs at Slaughter Level in Portugal. Foodborne Pathog. Dis. 2013, 10, 74–79. [Google Scholar] [CrossRef]
- Okeke, I.N.; Wallace-Gadsden, F.; Simons, H.R.; Matthews, N.; Labar, A.S.; Hwang, J.; Wain, J. Multi-Locus Sequence Typing of Enteroaggregative Escherichia coli Isolates from Nigerian Children Uncovers Multiple Lineages. PLoS ONE 2010, 5, e14093. [Google Scholar] [CrossRef] [PubMed]
- Oteo, J.; Pérez-Vázquez, M.; Campos, J. Extended-Spectrum Beta-Lactamase Producing Escherichia coli: Changing Epidemiology and Clinical Impact. Curr. Opin. Infect. Dis. 2010, 23, 320–326. [Google Scholar] [CrossRef] [PubMed]
- Camacho, C.; Coulouris, G.; Avagyan, V.; Ma, N.; Papadopoulos, J.; Bealer, K.; Madden, T.L. BLAST+: Architecture and Applications. BMC Bioinform. 2009, 10, 421. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Kim, J.; Jeon, S.E.; Kim, S.J.; Kim, N.O.; Hong, S.; Kang, Y.H.; Han, S.; Chung, G.T. Complete Nucleotide Sequence of the IncI1 Plasmid PSH4469 Encoding CTX-M-15 Extended-Spectrum β-Lactamase in a Clinical Isolate of Shigella sonnei from an Outbreak in the Republic of Korea. Int. J. Antimicrob. Agents 2014, 44, 533–537. [Google Scholar] [CrossRef] [PubMed]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Park, J.; Shin, E.; Han, J.; Kang, W.; Yoo, J.; Yoo, J.-S.; Roh, D.-H.; Kim, J. Distribution of Extended-Spectrum-β-Lactamase-Producing Diarrheagenic Escherichia coli Clonal Complex 10 Isolates from Patients with Diarrhea in the Republic of Korea. Antibiotics 2023, 12, 1614. https://doi.org/10.3390/antibiotics12111614
Park J, Shin E, Han J, Kang W, Yoo J, Yoo J-S, Roh D-H, Kim J. Distribution of Extended-Spectrum-β-Lactamase-Producing Diarrheagenic Escherichia coli Clonal Complex 10 Isolates from Patients with Diarrhea in the Republic of Korea. Antibiotics. 2023; 12(11):1614. https://doi.org/10.3390/antibiotics12111614
Chicago/Turabian StylePark, Jungsun, Eunkyung Shin, Joohyun Han, Wooju Kang, Jaeil Yoo, Jung-Sik Yoo, Dong-Hyun Roh, and Junyoung Kim. 2023. "Distribution of Extended-Spectrum-β-Lactamase-Producing Diarrheagenic Escherichia coli Clonal Complex 10 Isolates from Patients with Diarrhea in the Republic of Korea" Antibiotics 12, no. 11: 1614. https://doi.org/10.3390/antibiotics12111614
APA StylePark, J., Shin, E., Han, J., Kang, W., Yoo, J., Yoo, J.-S., Roh, D.-H., & Kim, J. (2023). Distribution of Extended-Spectrum-β-Lactamase-Producing Diarrheagenic Escherichia coli Clonal Complex 10 Isolates from Patients with Diarrhea in the Republic of Korea. Antibiotics, 12(11), 1614. https://doi.org/10.3390/antibiotics12111614