
Expansion of the pRAS3 Plasmid Family in Aeromonas salmonicida subsp. salmonicida and Growing Evidence of Interspecies Connections for These Plasmids


 and
and 
Abstract
1. Introduction
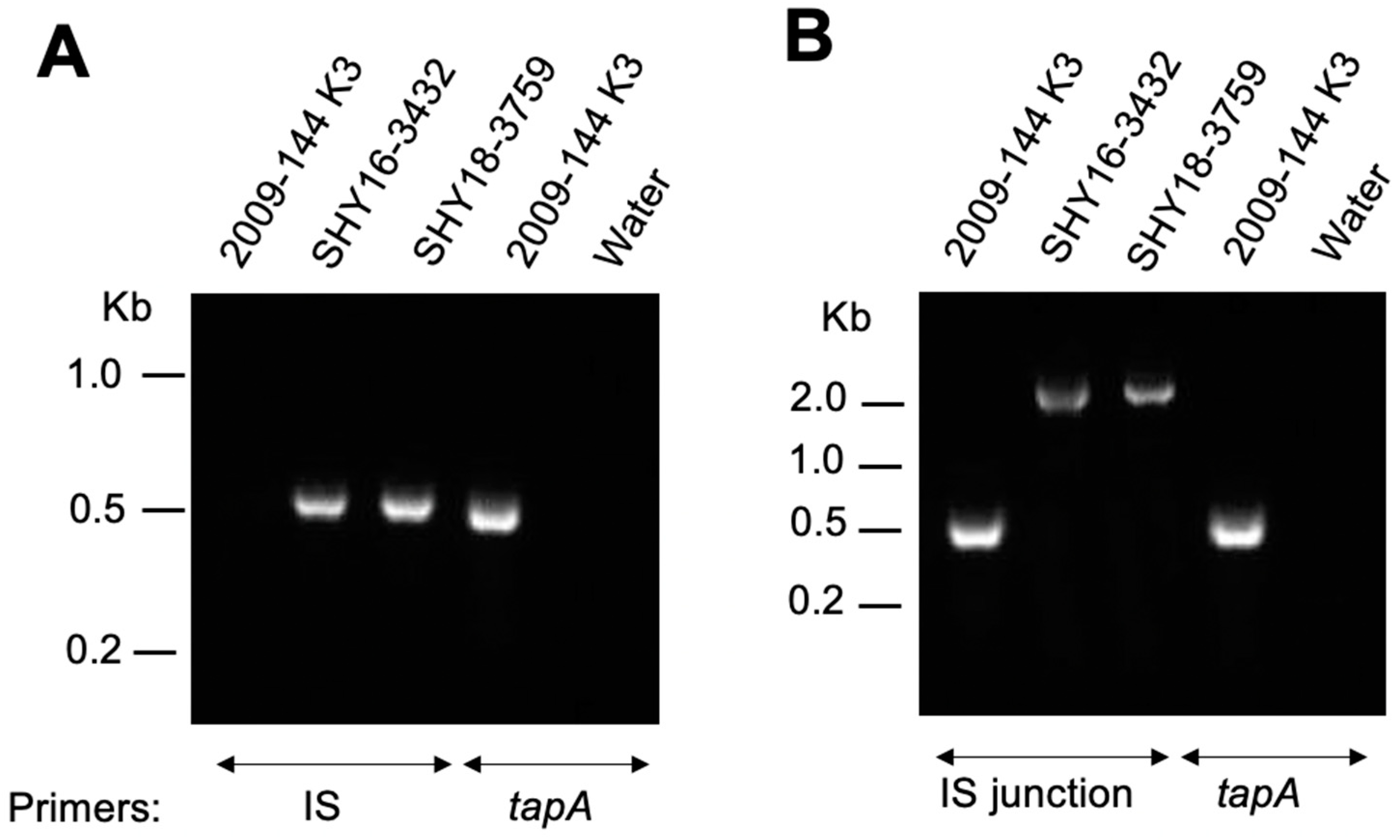
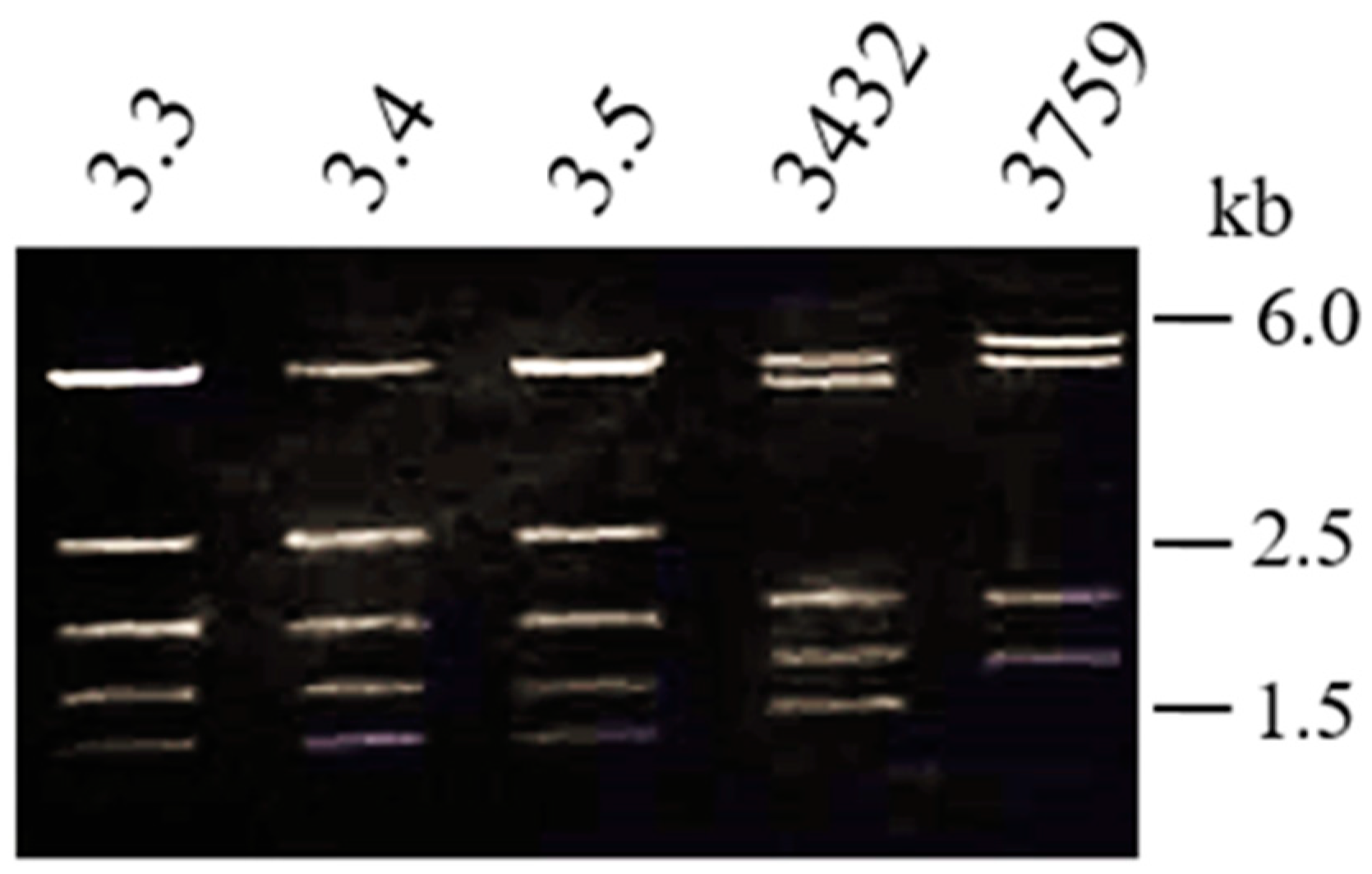
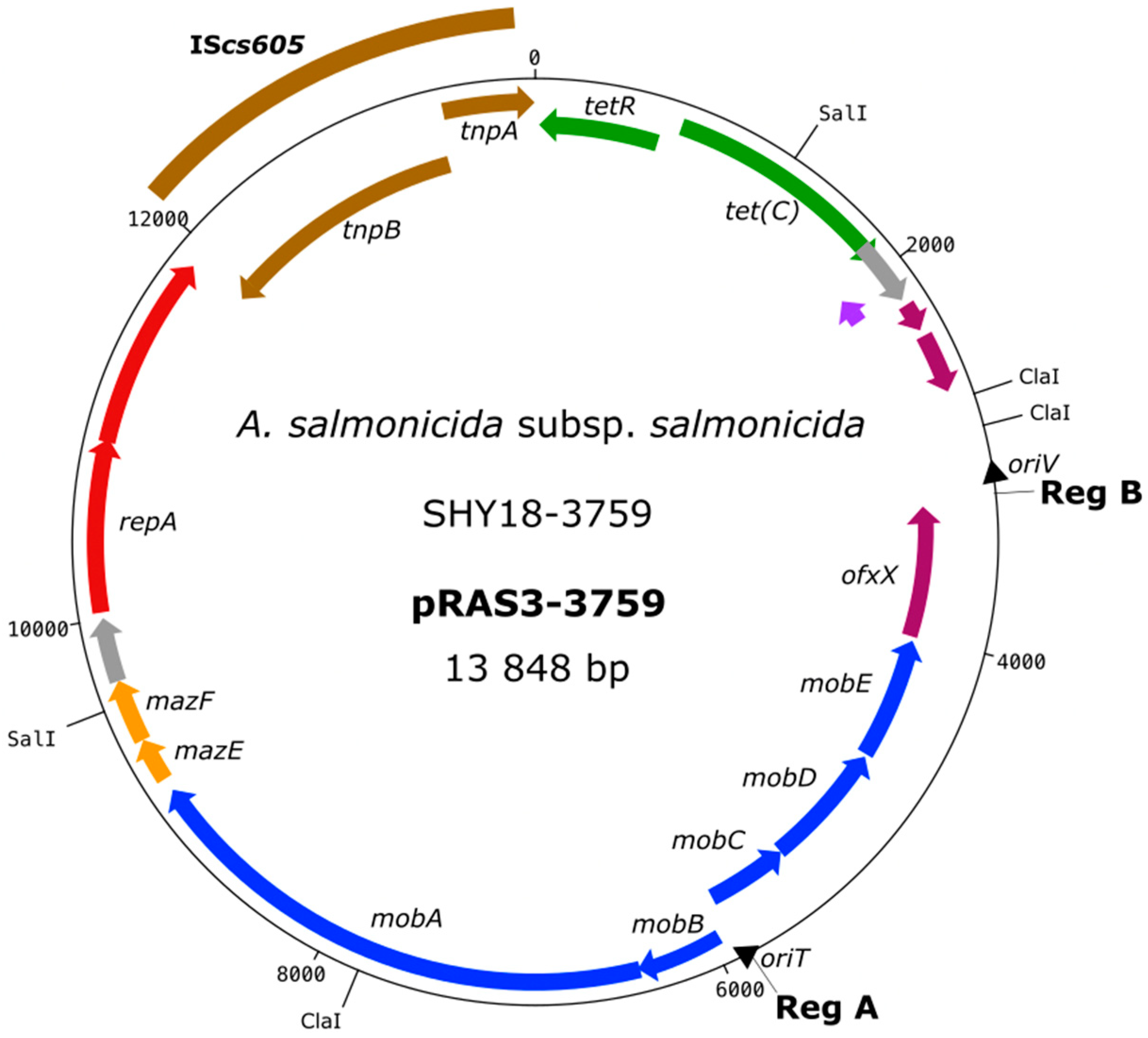
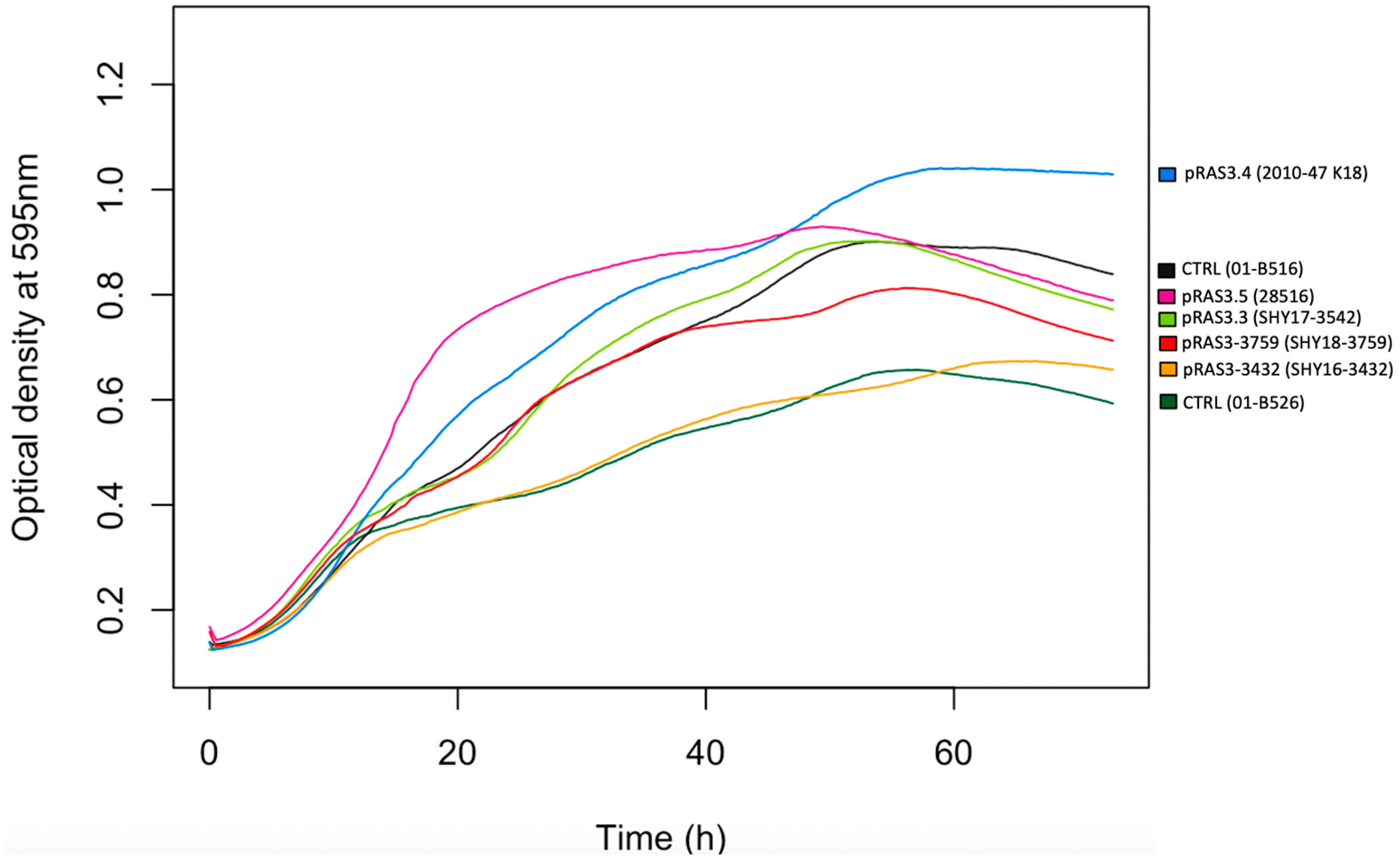
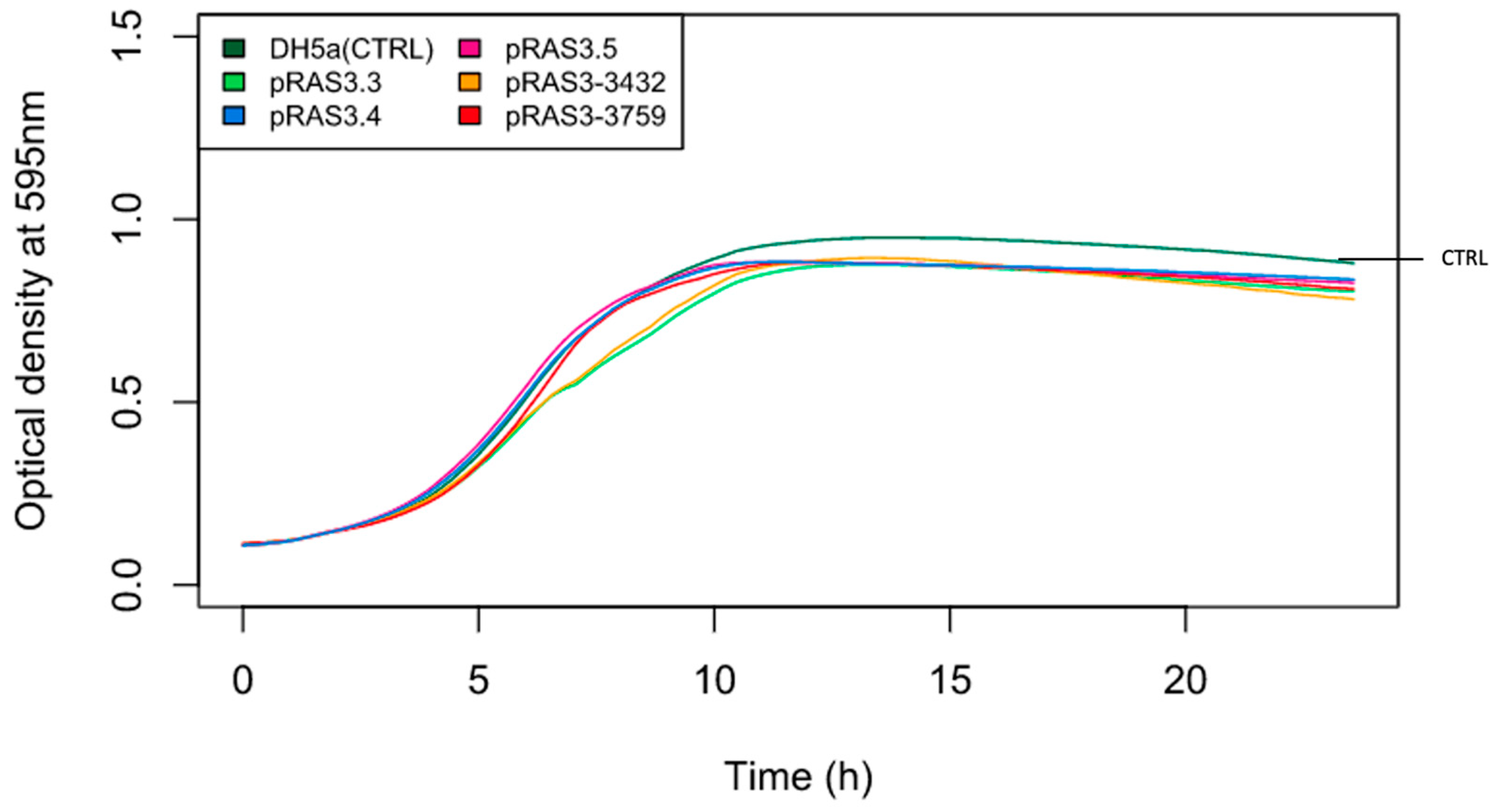
2. Results
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Growth Conditions
4.2. PCR Analyses
4.3. DNA Extraction and Sequencing
4.4. Sequence Assembly and Analyses
4.5. Copy Number of pRAS3 Variants in Their Host Strains
4.6. Plasmid Electroporation
4.7. Growth Curves
4.8. Assessment of the Minimum Inhibitory Concentration (MIC) of Tetracycline
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dallaire-Dufresne, S.; Tanaka, K.H.; Trudel, M.V.; Lafaille, A.; Charette, S.J. Virulence, genomic features, and plasticity of Aeromonas salmonicida subsp. salmonicida, the causative agent of fish furunculosis. Vet. Microbiol. 2014, 169, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Charette, S.J. Microbe profile: Aeromonas salmonicida: An opportunistic pathogen with multiple personalities. Microbiology 2021, 167, 001052. [Google Scholar] [CrossRef] [PubMed]
- Vincent, A.T.; Hosseini, N.; Charette, S.J. The Aeromonas salmonicida plasmidome: A model of modular evolution and genetic diversity. Ann. N. Y. Acad. Sci. 2021, 1488, 16–32. [Google Scholar] [CrossRef]
- Attéré, S.A.; Vincent, A.T.; Paccaud, M.; Frenette, M.; Charette, S.J. The Role for the Small Cryptic Plasmids As Moldable Vectors for Genetic Innovation in Aeromonas salmonicida subsp. salmonicida. Front. Genet. 2017, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Massicotte, M.-A.; Vincent, A.T.; Schneider, A.; Paquet, V.E.; Frenette, M.; Charette, S.J. One Aeromonas salmonicida subsp. salmonicida isolate with a pAsa5 variant bearing antibiotic resistance and a pRAS3 variant making a link with a swine pathogen. Sci. Total Environ. 2019, 690, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Loftie-Eaton, W.; Rawlings, D.E. Evolutionary competitiveness of two natural variants of the IncQ-like plasmids, pRAS3.1 and pRAS3.2. J. Bacteriol. 2010, 192, 6182–6190. [Google Scholar] [CrossRef] [PubMed]
- Aoki, T.; Takahashi, A. Tetracycline-Resistant Gene of a Non-Transferable R Plasmid from Fish-Pathogenic Bacteria Aeromonas salmonicida. Nippon Suisan Gakkaishi 1986, 52, 1913–1917. [Google Scholar] [CrossRef]
- L’Abée-Lund, T.M.; Sørum, H. A global non-conjugative Tet C plasmid, pRAS3, from Aeromonas salmonicida. Plasmid 2002, 47, 172–181. [Google Scholar] [CrossRef]
- Vincent, A.T.; Trudel, M.V.; Paquet, V.E.; Boyle, B.; Tanaka, K.H.; Dallaire-Dufresne, S.; Daher, R.K.; Frenette, M.; Derome, N.; Charette, S.J. Detection of variants of the pRAS3, pAB5S9, and pSN254 plasmids in Aeromonas salmonicida subsp. salmonicida: Multidrug resistance, interspecies exchanges, and plasmid reshaping. Antimicrob. Agents Chemother. 2014, 58, 7367–7374. [Google Scholar] [CrossRef]
- Gauthier, J.; Marquis, H.; Paquet, V.E.; Charette, S.J.; Levesque, R.C.; Derome, N. Genomic Perspectives on Aeromonas salmonicida subsp. salmonicida Strain 890054 as a Model System for Pathogenicity Studies and Mitigation of Fish Infections. Front. Mar. Sci. 2021, 8, 744052. [Google Scholar] [CrossRef]
- Bertram, R.; Neumann, B.; Schuster, C.F. Status quo of tet regulation in bacteria. Microb. Biotechnol. 2021, 15, 1101–1119. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Mascaretti, O.A. Bacteria Versus Antibacterial Agents: An Integrated Approach; ASM Press: Washington, DC, USA, 2003. [Google Scholar]
- Loftie-Eaton, W.; Rawlings, D.E. Comparative biology of two natural variants of the IncQ-2 family plasmids, pRAS3.1 and pRAS3.2. J. Bacteriol. 2009, 191, 6436–6446. [Google Scholar] [CrossRef][Green Version]
- Dugan, J.; Rockey, D.D.; Jones, L.; Andersen, A.A. Tetracycline Resistance in Chlamydia suis Mediated by Genomic Islands Inserted into the Chlamydial inv-Like Gene. Antimicrob. Agents Chemother. 2004, 48, 3989–3995. [Google Scholar] [CrossRef] [PubMed]
- Karvelis, T.; Druteika, G.; Bigelyte, G.; Budre, K.; Zedaveinyte, R.; Silanskas, A.; Kazlauskas, D.; Venclovas, Č.; Siksnys, V. Transposon-associated TnpB is a programmable RNA-guided DNA endonuclease. Nature 2021, 599, 692–696. [Google Scholar] [CrossRef]
- Darphorn, T.S.; Bel, K.; Koenders-van Sint Anneland, B.B.; Brul, S.; Ter Kuile, B.H. Antibiotic resistance plasmid composition and architecture in Escherichia coli isolates from meat. Sci. Rep. 2021, 11, 2136. [Google Scholar] [CrossRef]
- Attéré, S.A.; Vincent, A.T.; Trudel, M.V.; Chanut, R.; Charette, S.J. Diversity and Homogeneity among Small Plasmids of Aeromonas salmonicida subsp. salmonicida Linked with Geographical Origin. Front. Microbiol. 2015, 6, 1274. [Google Scholar] [CrossRef]
- Yamasaki, S.; Nikaido, E.; Nakashima, R.; Sakurai, K.; Fujiwara, D.; Fujii, I.; Nishino, K. The crystal structure of multidrug-resistance regulator RamR with multiple drugs. Nat. Commun. 2013, 4, 2078. [Google Scholar] [CrossRef]
- Hassan, H.A.; Ding, X.; Zhang, X.; Zhu, G. Fish borne Edwardsiella tarda eha involved in the bacterial biofilm formation, hemolytic activity, adhesion capability and pathogenicity. Arch. Microbiol. 2020, 202, 835–842. [Google Scholar] [CrossRef]
- Bagley, S.T. Habitat association of Klebsiella species. Infect. Control 1985, 6, 52–58. [Google Scholar] [CrossRef]
- Roberts, M.C. Update on acquired tetracycline resistance genes. FEMS Microbiol. Lett. 2005, 245, 195–203. [Google Scholar] [CrossRef] [PubMed]
- Lafaille, A. Rapport des Activités en Ichtyopathologie du 1er Janvier 2013 au 31 Décembre 2013; Service de Diagnostic en Ichtyopathologie de la Faculté de Médecine Vétérinaire de l’Université de Montréal: Saint-Hyacinthe, QC, Canada, 2014; 21p. [Google Scholar]
- Public Health Agency of Canada. Canadian Antimicrobial Resistance Surveillance System Report; Public Health Agency of Canada: Ottawa, ON, Canada, 2021.
- Gouvernement du Québec. Élevage Porcin (porc). Available online: https://www.quebec.ca/agriculture-environnement-et-ressources-naturelles/agriculture/industrie-agricole-au-quebec/productions-agricoles/elevage-porcin-porc (accessed on 28 June 2022).
- Programme Québécois D’antibiosurveillance Vétérinaire. Résultat de la Surveillance Passive de L’antibiorésistance. Available online: https://www.mapaq.gouv.qc.ca/SiteCollectionDocuments/Santeanimale/Antibioresistance/Rapportannuel2020-Surveillance_passive_antibioresistance.pdf (accessed on 28 June 2022).
- Zhao, Y.; Yang, Q.E.; Zhou, X.; Wang, F.-H.; Muurinen, J.; Virta, M.P.; Brandt, K.K.; Zhu, Y.-G. Antibiotic resistome in the livestock and aquaculture industries: Status and solutions. Crit. Rev. Environ. Sci. Technol. 2021, 51, 2159–2196. [Google Scholar] [CrossRef]
- Daher, R.K.; Filion, G.; Tan, S.G.; Dallaire-Dufresne, S.; Paquet, V.E.; Charette, S.J. Alteration of virulence factors and rearrangement of pAsa5 plasmid caused by the growth of Aeromonas salmonicida in stressful conditions. Vet. Microbiol. 2011, 152, 353–360. [Google Scholar] [CrossRef]
- Coil, D.; Jospin, G.; Darling, A.E. A5-miseq: An updated pipeline to assemble microbial genomes from Illumina MiSeq data. Bioinformatics 2015, 31, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Seemann, T. Prokka: Rapid prokaryotic genome annotation. Bioinformatics 2014, 30, 2068–2069. [Google Scholar] [CrossRef]
- Carver, T.; Harris, S.R.; Berriman, M.; Parkhill, J.; McQuillan, J.A. Artemis: An integrated platform for visualization and analysis of high-throughput sequence-based experimental data. Bioinformatics 2012, 28, 464–469. [Google Scholar] [CrossRef]
- Carver, T.; Thomson, N.; Bleasby, A.; Berriman, M.; Parkhill, J. DNAPlotter: Circular and linear interactive genome visualization. Bioinformatics 2009, 25, 119–120. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Langmead, B.; Salzberg, S.L. Fast gapped-read alignment with Bowtie 2. Nat. Methods 2012, 9, 357–359. [Google Scholar] [CrossRef]
- Li, H.; Handsaker, B.; Wysoker, A.; Fennell, T.; Ruan, J.; Homer, N.; Marth, G.; Abecasis, G.; Durbin, R.; Genome Project Data Processing Subgroup. The Sequence Alignment/Map format and SAMtools. Bioinformatics 2009, 25, 2078–2079. [Google Scholar] [CrossRef]
- García-Alcalde, F.; Okonechnikov, K.; Carbonell, J.; Cruz, L.M.; Götz, S.; Tarazona, S.; Dopazo, J.; Meyer, T.F.; Conesa, A. Qualimap: Evaluating next-generation sequencing alignment data. Bioinformatics 2012, 28, 2678–2679. [Google Scholar] [CrossRef] [PubMed]
- Dower, W.J.; Miller, J.F.; Ragsdale, C.W. High efficiency transformation of E. coli by high voltage electroporation. Nucleic Acids Res. 1988, 16, 6127–6145. [Google Scholar] [CrossRef] [PubMed]
- Ebanks, R.O.; Knickle, L.C.; Goguen, M.; Boyd, J.M.; Pinto, D.M.; Reith, M.; Ross, N.W. Expression of and secretion through the Aeromonas salmonicida type III secretion system. Microbiology 2006, 152, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Trudel, M.V.; Tanaka, K.H.; Filion, G.; Daher, R.K.; Frenette, M.; Charette, S.J. Insertion sequence AS5 (ISAs5) is involved in the genomic plasticity of Aeromonas salmonicida. Mob. Genet. Elem. 2013, 3, e25640. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain | Fish | Origin a | Tetracycline Resistance Gene-Bearing Plasmid (pRAS3 and pAsa10) | Presence of the pRAS3-3432 Related IS |
|---|---|---|---|---|
| 01-B526 b | Brook trout | Qc (E) | None | No |
| 01-B516 b | Brook trout | Qc (F) | None | No |
| 2009-157 K5 | Brook trout | NB | pRAS3 | No |
| 2010-47 K18 | Brook trout | NB | pRAS3 | No |
| 2009-195 K29 | Brook trout | NB | pRAS3 | No |
| 2009-144 K3 c | Brook trout | NB | pRAS3 | No |
| SHY16-3432 c | Brook trout | Qc (F) | pRAS3 | Yes |
| 91005 * | Atlantic salmon | NY | pRAS3 | No |
| 91009 * | Atlantic salmon | NY | pRAS3 | No |
| 96049 c * | Trout | NY | pRAS3 | No |
| 900001 * | Rainbow trout | NY | pRAS3 | No |
| 890054 * | Brook trout | NY | pRAS3 | No |
| SHY17-3542 * | Brook trout | Qc (A) | pRAS3 | No |
| SHY17-5108 * | NA | Qc (A) | pRAS3 | No |
| SHY-18-2492 * | Brook trout | Qc (C) | pRAS3 | Yes |
| SHY18-3221 * | Brook trout | Qc (A) | pRAS3 | No |
| SHY18-3759 c * | Brook trout | Qc (C) | pRAS3 | Yes |
| 28516 c * | Trout | France | pRAS3 | No |
| SHY19-3932 ** | Brook trout | Qc (G) | pAsa10 | No |
| SHY19-4656 * | Brook trout | Qc (C) | pRAS3 | Yes |
| SHY19-4655 * | Brook trout | Qc (C) | pRAS3 | Yes |
| SHY19-4654 * | Brook trout | Qc (C) | pRAS3 | Yes |
| SHY20-2575 * | Brook trout | Qc (C) | pRAS3 | Yes |
| SHY20-2715 * | Brook trout | Qc (B) | pRAS3 | No |
| SHY20-3274 * | Brook trout | Qc (C) | pRAS3 | Yes |
| SHY20-3753 * | Brook trout | Qc (C) | pRAS3 | No |
| SHY20-1481** | Brook trout | Qc (G) | pAsa10 | No |
| Plasmid | RegA | RegB |
|---|---|---|
| pRAS3.1 | CTCCTTCTCGC (CGGGGG) X4 CGGGGTGTGTCT | TGTCA (GGTTACCACCTGCGCCGGGGGT) X3 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| pRAS3.2 | CTCCTTCTCGC (CGGGGG) X3 CGGGGTGTGTCT | TGTCA (GGTTACCACCTGCGCCGGGGGT) X2 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| pRAS3.3 | CTCCTTCTCGC (CGGGGG) X3 CGGGGTGTGTCT | TGTCA (GGTTACCACCTGCGCCGGGGGT) X3 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| pRAS3.4 | CTCCTTCTCGC (CGGGGG) X3 CGGGGTGTGTCT | TGTCA (GGTTACCACCTGCGCCGGGGGT) X4 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| pRAS3.5 a | CTCCTTCTCGC (CGGGGG) X4 CGGGGTGTGTCT | TGTCA (GGTTACCACCTGCGCCGGGGGT) X5 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| pRAS3-3432 | CTCCTTCTCGC (CGGGGG) X2 GCGGGGTGTGTCT b | TGTCA (GGTTACCACCTGCGCCGGGGGT) X6 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| pRAS3-3759 a | CTCCTTCTCGC (CGGGGG) X3 CGGGGTGTGTCT | TGTCA (GGTTACCACCTGCGCCGGGGGT) X3 GGTTACCACCTGCGCCGGGGGCTTGAATT |
| Plasmid | MIC (µg/mL) for Tetracycline a |
|---|---|
| pRAS3.3 | 128 |
| pRAS3.4 | 256 |
| pRAS3.5 | 128 |
| pRAS3-3432 | 64 |
| pRAS3-3759 | 256 |
| Isolate | Type of pRAS3 a | Copy number of pRAS3 b | Number of Repeats at RegA | Number of Repeats at RegB |
|---|---|---|---|---|
| SHY16-3432 | pRAS3-3432 | 6 | 2 | 6 |
| SHY18-3759 SHY17-3542 | pRAS3-3759 pRAS3.3 | 3 4 | 3 3 | 3 3 |
| 96049 | pRAS3.4 | 10 | 3 | 4 |
| 28516 | pRAS3.5 | 4 | 4 | 5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fournier, K.C.; Paquet, V.E.; Attéré, S.A.; Farley, J.; Marquis, H.; Gantelet, H.; Ravaille, C.; Vincent, A.T.; Charette, S.J. Expansion of the pRAS3 Plasmid Family in Aeromonas salmonicida subsp. salmonicida and Growing Evidence of Interspecies Connections for These Plasmids. Antibiotics 2022, 11, 1047. https://doi.org/10.3390/antibiotics11081047
Fournier KC, Paquet VE, Attéré SA, Farley J, Marquis H, Gantelet H, Ravaille C, Vincent AT, Charette SJ. Expansion of the pRAS3 Plasmid Family in Aeromonas salmonicida subsp. salmonicida and Growing Evidence of Interspecies Connections for These Plasmids. Antibiotics. 2022; 11(8):1047. https://doi.org/10.3390/antibiotics11081047
Chicago/Turabian StyleFournier, Kim C., Valérie E. Paquet, Sabrina A. Attéré, Judith Farley, Hélène Marquis, Hubert Gantelet, Christian Ravaille, Antony T. Vincent, and Steve J. Charette. 2022. "Expansion of the pRAS3 Plasmid Family in Aeromonas salmonicida subsp. salmonicida and Growing Evidence of Interspecies Connections for These Plasmids" Antibiotics 11, no. 8: 1047. https://doi.org/10.3390/antibiotics11081047
APA StyleFournier, K. C., Paquet, V. E., Attéré, S. A., Farley, J., Marquis, H., Gantelet, H., Ravaille, C., Vincent, A. T., & Charette, S. J. (2022). Expansion of the pRAS3 Plasmid Family in Aeromonas salmonicida subsp. salmonicida and Growing Evidence of Interspecies Connections for These Plasmids. Antibiotics, 11(8), 1047. https://doi.org/10.3390/antibiotics11081047