
Characterization of Diarreaghenic Escherichia coli Strains Isolated from Healthy Donors, including a Triple Hybrid Strain


 , , , ,
, , , ,  and
and
Abstract
1. Introduction
2. Results
2.1. Strains Identification
2.2. Phylogenetic Group Determination
2.3. E. coli Isolates from Healthy Humans, Harbor Several Resistant Traits
2.4. High Prevalence of aEPEC Pathotype between E. coli Strains Isolated from Healthy Donors
2.5. Adherence Patterns of E. coli Diarrheagenic Pathotypes Identified
2.6. Virulence Genes Are Differentially Spread between E. coli Strains
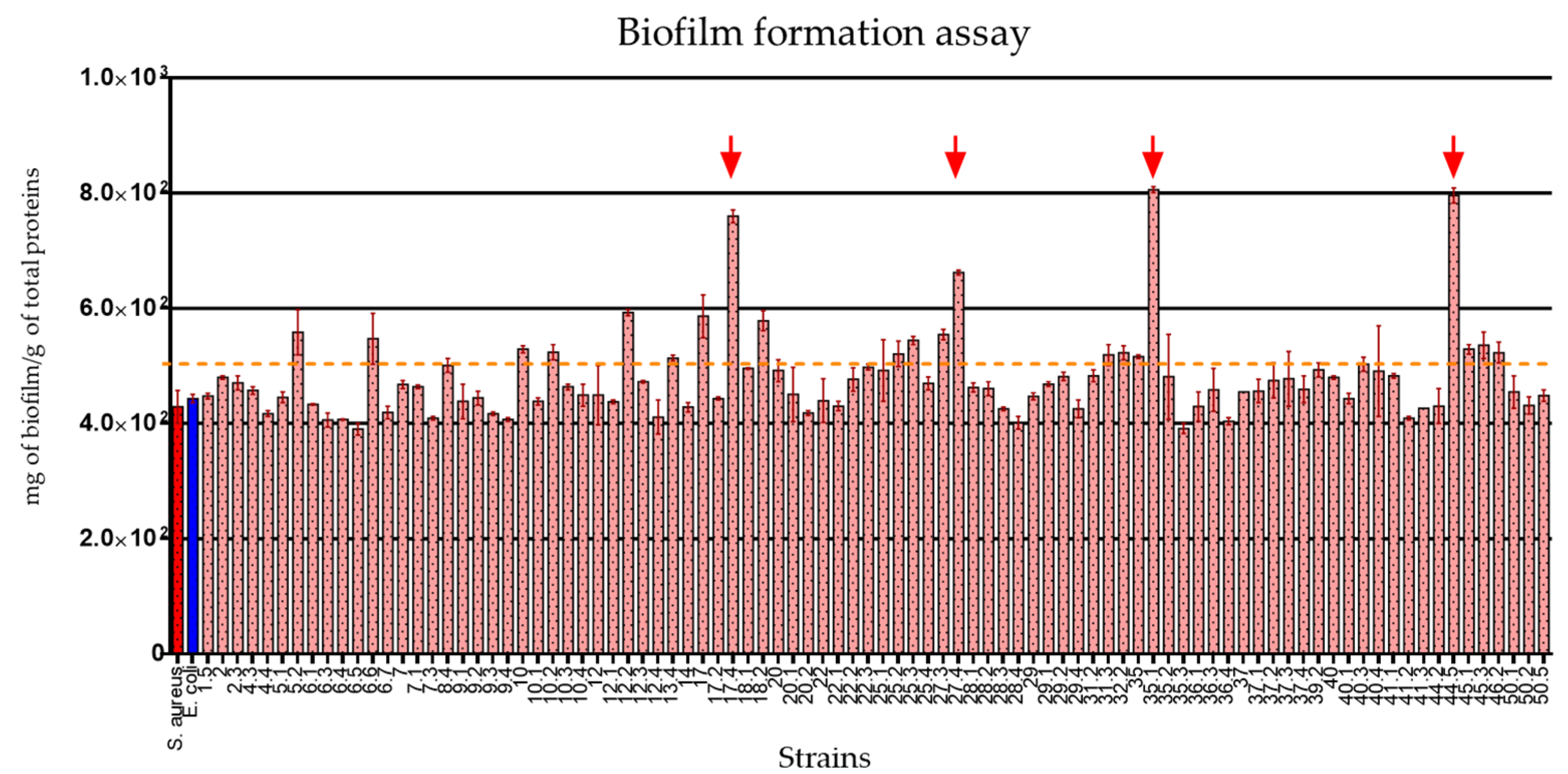
2.7. The E. coli Strains Isolated from Healthy Donors Showed Important Biofilm Production Traits
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Sampling
5.2. Isolation and Biochemical Characterization of E. coli
5.3. Genomic DNA Extraction
5.4. Molecular Identification of E. coli
5.5. Phylogenetic Group Determination
5.6. Antibiotic Agar Dilution and ESBL Genes Detection
5.7. Determination of Minimum Inhibitory Concentration (MIC)
5.8. Determination of Diarrheagenic Pathotypes
5.9. Adherence Assay
5.10. Virotyping
5.11. Biofilm Formation Assays
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Torres, A.G. Escherichia Coli in the Americas; Springer International Publishing: Cham, Switzerland, 2016; pp. 125–147. ISBN 9783319450926. [Google Scholar]
- Kaper, J.B.; Nataro, J.P.; Mobley, H.L.T. Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2004, 2, 123–140. [Google Scholar] [CrossRef] [PubMed]
- McLellan, L.K.; Hunstad, D.A. Urinary Tract Infection: Pathogenesis and Outlook. Trends Mol. Med. 2016, 22, 946–957. [Google Scholar] [CrossRef] [PubMed]
- Saldaña, Z.; Erdem, A.L.; Schüller, S.; Okeke, I.N.; Lucas, M.; Sivananthan, A.; Phillips, A.D.; Kaper, J.B.; Puente, J.L.; Girón, J.A. The Escherichia Coli Common Pilus and the Bundle-Forming Pilus Act in Concert during the Formation of Localized Adherence by Enteropathogenic E. coli. J. Bacteriol. 2009, 191, 3451–3461. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Schiller, P.; Knödler, M.; Berger, P.; Greune, L.; Fruth, A.; Mellmann, A.; Dersch, P.; Berger, M.; Dobrindt, U. The Superior Adherence Phenotype of E. coli O104:H4 Is Directly Mediated by the Aggregative Adherence Fimbriae Type I. Virulence 2021, 12, 346–359. [Google Scholar] [CrossRef] [PubMed]
- Croxen, M.A.; Finlay, B.B. Molecular Mechanisms of Escherichia coli Pathogenicity. Nat. Rev. Microbiol. 2010, 8, 26–38. [Google Scholar] [CrossRef]
- Liceaga, S.E.C.; Sandoval, G.C.; García, B.A.H.; Macías, A.C.; Pedroza, J.F.R.; García, D.A.C.; Lemuz, Z.R.F.; Escamilla, A.L.L.; Valadez, J.G. Manual de Procedimientos Estandarizados para la Vigilancia Epidemiológica de Enfermedad Diarréica Aguda; Dirección General de Epidemiología, Secretaria de Salud México: Acapulco de Juárez, Mexico, 2022. [Google Scholar]
- Alonso, C.A.; Cortés-Cortés, G.; Maamar, E.; Massó, M.; Rocha-Gracia, R.D.C.; Torres, C.; Centrón, D.; Quiroga, M.P. Molecular Diversity and Conjugal Transferability of Class 2 Integrons among Escherichia coli Isolates from Food, Animal and Human Sources. Int. J. Antimicrob. Agents 2018, 51, 905–911. [Google Scholar] [CrossRef]
- Cortés-Cortés, G.; Lozano-Zarain, P.; Torres, C.; Alonso, C.A.; Ríos-Torres, A.M.; Castañeda, M.; López-Pliego, L.; Navarro, A.; del Carmen Rocha-Gracia, R. Extended-Spectrum β-Lactamase-Producing Escherichia Coli Isolated from Healthy Humans in Mexico, Including Subclone ST131-B2-O25:H4-H30-Rx. J. Glob. Antimicrob. Resist. 2017, 9, 130–134. [Google Scholar] [CrossRef]
- Akeda, Y.; Motooka, D.; Yamamoto, N.; Okada, K.; Nakamura, S.; Sakamoto, N.; Hagiya, H.; Zin, K.N.; Nishi, I.; Aye, M.M.; et al. Genetic Characterization of BlaNDM-Harboring Plasmids in Carbapenem-Resistant Escherichia Coli from Myanmar. PLoS ONE 2017, 12, e0184720. [Google Scholar] [CrossRef]
- Tijet, N.; Faccone, D.; Rapoport, M.; Seah, C.; Pasterán, F.; Ceriana, P.; Albornoz, E.; Corso, A.; Petroni, A.; Melano, R.G. Molecular Characteristics of Mcr-1-Carrying Plasmids and New Mcr-1 Variant Recovered from Polyclonal Clinical Escherichia coli from Argentina and Canada. PLoS ONE 2017, 12, e0180347. [Google Scholar] [CrossRef]
- Karim, A.; Poirel, L.; Nagarajan, S.; Nordmann, P. Plasmid-Mediated Extended-Spectrum β-Lactamase (CTX-M-3 like) from India and Gene Association with Insertion Sequence ISEcp1. FEMS Microbiol. Lett. 2001, 201, 237–241. [Google Scholar] [CrossRef]
- Sukumaran, D.P.; Hatha, A.A.M. Antibiotic Resistance and Virulence Genes of Extraintestinal Pathogenic Escherichia coli from Tropical Estuary, South India. J. Infect. Dev. Ctries. 2015, 9, 496–504. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kadlec, K.; Schwarz, S. Analysis and Distribution of Class 1 and Class 2 Integrons and Associated Gene Cassettes among Escherichia coli Isolates from Swine, Horses, Cats and Dogs Collected in the BfT-GermVet Monitoring Study. J. Antimicrob. Chemother. 2008, 62, 469–473. [Google Scholar] [CrossRef] [PubMed]
- Coque, T.M.; Baquero, F.; Canton, R. Increasing Prevalence of ESBL-Producing Enterobacteriaceae in Europe. Eurosurveillance 2008, 13, 19044. [Google Scholar] [CrossRef] [PubMed]
- Kolenda, R.; Burdukiewicz, M.; Schierack, P. A Systematic Review and Meta-Analysis of the Epidemiology of Pathogenic Escherichia coli of Calves and the Role of Calves as Reservoirs for Human Pathogenic E. coli. Front. Cell. Infect. Microbiol. 2015, 5, 23. [Google Scholar] [CrossRef] [PubMed]
- Chaudhuri, R.R.; Henderson, I.R. The Evolution of the Escherichia coli Phylogeny. Infect. Genet. Evol. 2012, 12, 214–226. [Google Scholar] [CrossRef]
- Loayza, F.; Graham, J.P.; Trueba, G. Factors Obscuring the Role of E. coli from Domestic Animals in the Global Antimicrobial Resistance Crisis: An Evidence-Based Review. Int. J. Environ. Res. Public Health 2020, 17, 3061. [Google Scholar] [CrossRef]
- Clermont, O.; Christenson, J.K.; Denamur, E.; Gordon, D.M. The Clermont Escherichia Coli Phylo-Typing Method Revisited: Improvement of Specificity and Detection of New Phylo-Groups. Environ. Microbiol. Rep. 2013, 5, 58–65. [Google Scholar] [CrossRef]
- Dale, A.P.; Woodford, N. Extra-Intestinal Pathogenic Escherichia Coli (ExPEC): Disease, Carriage and Clones. J. Infect. 2015, 71, 615–626. [Google Scholar] [CrossRef]
- Luna-Guevara, J.J.; Arenas-Hernandez, M.M.P.; de la Peña, C.M.; Silva, J.L.; Luna-Guevara, M.L. The Role of Pathogenic E. coli in Fresh Vegetables: Behavior, Contamination Factors, and Preventive Measures. Int. J. Microbiol. 2019, 2019, 2894328. [Google Scholar] [CrossRef]
- da Cruz, C.B.N.; de Souza, M.C.S.; Serra, P.T.; Santos, I.; Balieiro, A.; Pieri, F.; Nogueira, P.A.; Orlandi, P.P. Virulence Factors Associated with Pediatric Shigellosis in Brazilian Amazon. BioMed Res. Int. 2014, 2014, 539697. [Google Scholar] [CrossRef]
- Barrios-Villa, E.; Cortés-Cortés, G.; Lozano-Zaraín, P.; Arenas-Hernández, M.M.D.L.P.; De La Peña, C.F.M.; Martínez-Laguna, Y.; Torres, C.; Rocha-Gracia, R.D.C. Adherent/invasive Escherichia coli (AIEC) isolates from asymptomatic people: New E. coli ST131 O25:H4/H30-Rx virotypes. Ann. Clin. Microbiol. Antimicrob. 2018, 17, 42. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Jimenez, D.; Garcia-Meniño, I.; Herrera, A.; García, V.; Lopez-Beceiro, A.M.; Alonso, M.P.; Blanco, J.; Mora, A. Genomic Characterization of Escherichia coli Isolates Belonging to a New Hybrid aEPEC/ExPEC Pathotype O153:H10-A-ST10 eae-beta1 Occurred in Meat, Poultry, Wildlife and Human Diarrheagenic Samples. Antibiotics 2020, 9, 192. [Google Scholar] [CrossRef] [PubMed]
- Elwell, C.; Chao, K.; Patel, K.; Dreyfus, L. Escherichia coli CdtB Mediates Cytolethal Distending Toxin Cell Cycle Arrest. Infect. Immun. 2001, 69, 3418–3422. [Google Scholar] [CrossRef] [PubMed]
- Livermore, D.M.; Canton, R.; Gniadkowski, M.; Nordmann, P.; Rossolini, G.M.; Arlet, G.; Ayala, J.; Coque, T.M.; Kern-Zdanowicz, I.; Luzzaro, F.; et al. CTX-M: Changing the face of ESBLs in Europe. J. Antimicrob. Chemother. 2006, 59, 165–174. [Google Scholar] [CrossRef]
- Horton, R.A.; Randall, L.P.; Snary, E.L.; Cockrem, H.; Lotz, S.; Wearing, H.; Duncan, D.; Rabie, A.; McLaren, I.; Watson, E.; et al. Fecal Carriage and Shedding Density of CTX-M Extended-Spectrum β-Lactamase-Producing Escherichia coli in Cattle, Chickens, and Pigs: Implications for Environmental Contamination and Food Production. Appl. Environ. Microbiol. 2011, 77, 3715–3719. [Google Scholar] [CrossRef]
- Papa-Ezdra, R.; Cordeiro, N.F.; DiPilato, V.; Chiarelli, A.; Pallecchi, L.; Garcia-Fulgueiras, V.; Vignoli, R. Description of novel resistance islands harbouring blaCTX-M-2 in IncC type 2 plasmids. J. Glob. Antimicrob. Resist. 2021, 26, 37–41. [Google Scholar] [CrossRef]
- Zhang, Z.; Chang, J.; Xu, X.; Zhou, M.; Shi, C.; Liu, Y.; Shi, X. Dissemination of IncFII plasmids carrying fos A3 and bla CTX-M-55 in clinical isolates of Salmonella enteritidis. Zoonoses Public Health 2021, 68, 760–768. [Google Scholar] [CrossRef]
- Locke, R.K.; Greig, D.R.; Jenkins, C.; Dallman, T.J.; Cowley, L.A. Acquisition and loss of CTX-M plasmids in Shigella species associated with MSM transmission in the UK. Microb. Genom. 2021, 7, 000644. [Google Scholar] [CrossRef]
- Otto, M. Staphylococcal Biofilms. Microbiol. Spectr. 2018, 6, 207–228. [Google Scholar] [CrossRef]
- Venkatesan, N.; Perumal, G.; Doble, M. Bacterial resistance in biofilm-associated bacteria. Futur. Microbiol. 2015, 10, 1743–1750. [Google Scholar] [CrossRef]
- Bardoloi, V.; Babu, K.V.Y. Comparative study of isolates from community-acquired and catheter-associated urinary tract infections with reference to biofilm-producing property, antibiotic sensitivity and multi-drug resistance. J. Med. Microbiol. 2017, 66, 927–936. [Google Scholar] [CrossRef]
- Yan, J.; Bassler, B.L. Surviving as a Community: Antibiotic Tolerance and Persistence in Bacterial Biofilms. Cell Host Microbe 2019, 26, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Barrios-Villa, E.; Cortés-Cortés, G.; Zarain, P.L.; Romero-Romero, S.; Flores, N.L.; Estepa, V.; Somalo, S.; Torres, C.; Rocha-Gracia, R.D.C. Characterization of Extended-Spectrum and CMY-2 ß-Lactamases, and Associated Virulence Genes in Escherichia coli from Food of Animal Origin in México. Br. Food J. 2018, 120, 1457–1473. [Google Scholar] [CrossRef]
- O’Toole, G.A. Microtiter Dish Biofilm Formation Assay. JoVE 2011, 47, e2437. [Google Scholar] [CrossRef] [PubMed]
- Clermont, O.; Bonacorsi, S.; Bingen, E. Rapid and Simple Determination of the Escherichia coli Phylogenetic Group. Appl. Environ. Microbiol. 2000, 66, 4555–4558. [Google Scholar] [CrossRef] [PubMed]
- Walker, D.I.; McQuillan, J.; Taiwo, M.; Parks, R.; Stenton, C.A.; Morgan, H.; Mowlem, M.C.; Lees, D.N. A Highly Specific Escherichia Coli QPCR and Its Comparison with Existing Methods for Environmental Waters. Water Res. 2017, 126, 101–110. [Google Scholar] [CrossRef] [PubMed]
- León, E.A.C. Estudio Molecular de La Resistencia y Virulencia de Cepas de Esherichia coli Productoras de β-Lactamasas de Espectro Extendido Aisladas de Vegetales Crudos. Master’s Thesis, BUAP, Puebla, Mexico, 2019. [Google Scholar]
- Aranda, K.R.S.; Fabbricotti, S.H.; Fagundes-Neto, U.; Scaletsky, I.C.A. Single Multiplex Assay to Identify Simultaneously Enteropathogenic, Enteroaggregative, Enterotoxigenic, Enteroinvasive and Shiga Toxin-Producing Escherichia Coli Strains in Brazilian Children. FEMS Microbiol. Lett. 2007, 267, 145–150. [Google Scholar] [CrossRef]
- Alikhani, M.Y.; Bouzari, S.; Farhang, E.; Hosseini, S.M. Prevalence and Antimicrobial Resistance of Shiga Toxin-Producing Escherichia Coli and Enteropathogenic Escherichia coli Isolated from Patients with Acute Diarrhea. Iran. J. Microbiol. 2018, 10, 151–157. [Google Scholar]
- Franke, J.; Franke, S.; Schmidt, H.; Schwarzkopf, A.; Wieler, L.H.; Baljer, G.; Beutin, L.; Karchl, H.; Referenzlabor, N. E. coli Nucleotide Sequence Analysis of Enteropathogenic Escherichia coli (EPEC) Adherence Factor Probe and Development of PCR for Rapid Detection of EPEC Harboring Virulence Plasmids. J. Clin. Microbiol. 1994, 32, 2460–2463. [Google Scholar] [CrossRef]
- Fratamico, P.M.; Sackitey, S.K.; Wiedmann, M.; Deng, M.Y. Detection of Escherichia coli O157:H7 by Multiplex PCR. J. Clin. Microbiol. 1995, 33, 2188–2191. [Google Scholar] [CrossRef]
- Rodríguez-Angeles, M.G. Principales Características y Diagnóstico de Los Grupos Patógenos de Escherichia coli. Salud Public. M. 2002, 44, 464–475. [Google Scholar] [CrossRef]
- Aranda, K.R.S.; Fagundes-Neto, U.; Scaletsky, I.C.A. Evaluation of Multiplex PCRs for Diagnosis of Infection With. J. Clin. Microbiol. 2004, 42, 5849–5853. [Google Scholar] [CrossRef] [PubMed]
- Hornes, E.; Wasteson, Y.; Olsvik, Ø. Detection of Escherichia coli Heat-Stable Enterotoxin Genes in Pig Stool Specimens by an Immobilized, Colorimetric, Nested Polymerase Chain Reaction. J. Clin. Microbiol. 1991, 29, 2375–2379. [Google Scholar] [CrossRef] [PubMed]
- Mansan-Almeida, R.; Pereira, A.L.; Giugliano, L.G. Diffusely Adherent Escherichia Coli Strains Isolated from Children and Adults Constitute Two Different Populations. BMC Microbiol. 2013, 13, 22. [Google Scholar] [CrossRef]
- Schmidt, H.; Knop, C.; Franke, S.; Aleksic, S.; Karch, H. Development of PCR for Screening of Enteroaggregative. Microbiology 1995, 33, 701–705. [Google Scholar]
- Carlini, F.; Maroccia, Z.; Fiorentini, C.; Travaglione, S.; Fabbri, A. Effects of the Escherichia coli Bacterial Toxin Cytotoxic Necrotizing Factor 1 on Different Human and Animal Cells: A Systematic Review. Int. J. Mol. Sci. 2021, 22, 12610. [Google Scholar] [CrossRef]
- Garza-Gonzaález, E.; Bocanegra-Ibarias, P.; Bobadilla-del-Valle, M.; Ponce-de-León-Garduño, L.A.; Esteban-Kenel, V.; Silva-Sánchez, J.; Garza-Ramos, U.; Barrios-Camacho, H.; Lopez-Jacome, L.E.; Colin-Castro, C.A.; et al. The blaSHV-5 gene is encoded in a compound transposon duplicated in tandem in Enterobacter cloacae. Clin. Microbiol. Infect 2009, 15, 878–880. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Strain | Phylogroup | Pathotype | Resistotype | ESBL Genes | Minimum Inhibitory Concentration (µg/mL) |
|---|---|---|---|---|---|
| Ec-1.5 | A | NA | TE, DCI, FOX, AMP | --- | --- |
| Ec-2 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-2.3 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-4.3 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-4.4 | B2 | NA | DCI, AMP | --- | --- |
| Ec-5.1 | C | NA | DCI, AMP | --- | --- |
| Ec-5.2 | A | NA | DCI, AMP | --- | --- |
| Ec-6.1 | C | NA | DCI, FOX, CRO, AMP | --- | --- |
| Ec-6.3 | C | NA | DCI, AMP | --- | --- |
| Ec-6.4 | Unknown | NA | DCI, AMP | --- | --- |
| Ec-6.5 | B2 | NA | TE, C, DCI, FOX, CRO, AMP | --- | --- |
| Ec-6.6 | Unknown | NA | TE, C, DCI, AMP | --- | --- |
| Ec-6.7 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-7 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-7.1 | D | NA | DCI, AMP | --- | --- |
| Ec-7.3 | B2 | NA | FOS, DCI, AMP | --- | --- |
| Ec-8.4 | B1 | aEPEC | DCI, AMP | --- | --- |
| Ec-9.1 | A | NA | FOS, DCI, FOX, CRO, AMP | CTX-M1, CTX-M2 | 256 (FOX, CTX, CRO) |
| Ec-9.2 | B2 | NA | FOS, DCI, FOX, CRO, AMP | CTX-M1, CTX-M3 | 256 (FOX, CTX, CRO) |
| Ec-9.3 | A | aEPEC | DCI, FOX, CRO, AMP | CTX-M1 | 256 (FOX, CTX, CRO) |
| Ec-9.4 | A | NA | DCI, FOX, CRO, AMP | CTX-M1 | 256 (FOX, CTX, CRO) |
| Ec-10 | C | aEPEC | DCI, AMP | --- | --- |
| Ec-10.1 | B2 | aEPEC | FOS, DCI, AMP | --- | --- |
| Ec-10.2 | A | aEPEC | DCI, AMP | --- | --- |
| Ec-10.3 | B2 | NA | DCI, AMP | --- | --- |
| Ec-10.4 | B2 | aEPEC | DCI, AMP | --- | --- |
| Ec-12 | A | NA | DCI, AMP | CTX-M1 | 128 (FOX, CTX, CRO) |
| Ec-12.1 | B2 | NA | DCI, FOX, CRO, AMP | CTX-M1 | 128 (FOX, CTX), 256 (CRO) |
| Ec-12.2 | B2 | NA | DCI, FOX, CRO, AMP | CTX-M1 | 64 (FOX, CTX), 256 (CRO) |
| Ec-12.3 | B2 | NA | DCI, FOX, CRO, AMP | --- | --- |
| Ec-12.4 | A | NA | DCI, CRO, AMP | --- | --- |
| Ec-13.4 | C | NA | DCI, FOX, AMP | --- | --- |
| Ec-14 | F | aEPEC | DCI, AMP | --- | --- |
| Ec-17 | B2 | aEPEC | SENSITIVE | --- | --- |
| Ec-17.2 | B2 | ETEC | TE, DCI, AMP | --- | --- |
| Ec-17.4 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-18.1 | C | NA | TE, DCI, AMP | --- | --- |
| Ec-18.2 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-20 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-20.1 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-20.2 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-22 | F | NA | DCI, AMP | --- | --- |
| Ec-22.1 | B2 | NA | DCI, AMP | --- | --- |
| Ec-22.2 | B2 | NA | DCI | --- | --- |
| Ec-22.3 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-25.1 | F | NA | DCI, AMP | --- | --- |
| Ec-25.2 | A | aEPEC/ETEC | DCI, AMP | --- | --- |
| Ec-25.3 | B2 | aEPEC | DCI, AMP | --- | --- |
| Ec-25.4 | A | aEPEC | DCI, AMP | --- | --- |
| Ec-27.3 | A | NA | TE, DCI, AMP | --- | --- |
| Ec-27.4 | A | aEPEC | TE, FOS, DCI, AMP | --- | --- |
| Ec-28.1 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-28.2 | Unknown | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-28.3 | Unknown | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-28.4 | A | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-29 | A | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-29.1 | A | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-29.2 | C | aEPEC | TE, DCI, FOX, AMP | --- | --- |
| Ec-29.4 | C | aEPEC | TE, DCI, DOX, AMP | --- | --- |
| Ec-31.2 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-31.3 | A | ETEC | TE, DCI, AMP | --- | --- |
| Ec-32.2 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-35 | B2 | aEPEC | TE, FOS, DCI, AMP | --- | --- |
| Ec-35.1 | Unknown | aEPEC | TE, C, FOS, DCI, AMP | --- | --- |
| Ec-35.2 | C | aEPEC | TE, C, FOS, DCI, AMP | --- | --- |
| Ec-35.3 | C | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-36.1 | B2 | aEPEC/DAEC | FOS, DCI, AMP | --- | --- |
| Ec-6.3 | B2 | aEPEC/ETEC/DAEC | DCI | --- | --- |
| Ec-36.4 | B2 | aEPEC/ETEC | DCI | --- | --- |
| Ec-37 | B2 | aEPEC | TE, C, DCI, AMP | --- | --- |
| Ec-37.1 | F | aEPEC | TE, C, DCI, AMP | --- | --- |
| Ec-37.2 | B2 | aEPEC | DCI, AMP | --- | --- |
| Ec-37.3 | B2 | NA | TE, C, DCI, AMP | --- | --- |
| Ec-37.4 | F | ETEC | TE, C, DCI, AMP | --- | --- |
| Ec-39.2 | D | NA | C, DCI, AMP | --- | --- |
| Ec-40 | B2 | NA | DCI, AMP | --- | --- |
| Ec-40.1 | C | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-40.3 | F | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-40.4 | B2 | aEPEC | TE, DCI, AMP | --- | --- |
| Ec-41.1 | B1 | NA | DCI, AMP | --- | --- |
| Ec-41.2 | B1 | NA | C, DCI, AMP | --- | --- |
| Ec-41.3 | B1 | aEPEC | DCI, AMP | --- | --- |
| Ec-44.2 | B1 | aEPEC | TE, C, DCI, AMP | --- | --- |
| Ec-44.5 | B1 | NA | TE, DCI, AMP | --- | --- |
| Ec-45.1 | B1 | NA | DCI, AMP | --- | --- |
| Ec-45.3 | B1 | NA | DCI, AMP | --- | --- |
| Ec-46.2 | F | NA | DCI, AMP | --- | --- |
| Ec-50.1 | B2 | NA | TE, DCI, AMP | --- | --- |
| Ec-50.2 | B2 | NA | DCI, FOX, CRO, AMP | --- | --- |
| Ec-50.5 | B2 | NA | TE, DCI, AMP | --- | --- |
| Virotype | afa/draBC | afa Operon | iroN | satA | ibeA | papGII | cnfI | hlyA | papGIII | cdtB | neuC-KI |
|---|---|---|---|---|---|---|---|---|---|---|---|
| A a | + | + | − | ± | − | − | − | − | − | − | − |
| B a | − | − | + | ± | − | − | − | − | − | − | − |
| C a | − | − | − | + | − | − | − | − | − | − | − |
| D a | ± | ± | ± | ± | + | − | ± | ± | ± | ± | ± |
| E a | − | − | − | + | − | + | + | + | − | − | − |
| F a | − | − | + | − | − | + | + | − | − | − | − |
| G a | − | − | − | + | − | + | − | − | − | − | − |
| H * | − | − | ± | ± | − | + | ± | − | ± | ± | ± |
| I * | − | − | − | + | + | − | − | ± | − | + | + |
| J * | − | − | − | + | − | − | − | − | + | + | − |
| 0 | − | − | − | − | − | − | − | − | − | − | − |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Méndez-Moreno, E.; Caporal-Hernandez, L.; Mendez-Pfeiffer, P.A.; Enciso-Martinez, Y.; De la Rosa López, R.; Valencia, D.; Arenas-Hernández, M.M.P.; Ballesteros-Monrreal, M.G.; Barrios-Villa, E. Characterization of Diarreaghenic Escherichia coli Strains Isolated from Healthy Donors, including a Triple Hybrid Strain. Antibiotics 2022, 11, 833. https://doi.org/10.3390/antibiotics11070833
Méndez-Moreno E, Caporal-Hernandez L, Mendez-Pfeiffer PA, Enciso-Martinez Y, De la Rosa López R, Valencia D, Arenas-Hernández MMP, Ballesteros-Monrreal MG, Barrios-Villa E. Characterization of Diarreaghenic Escherichia coli Strains Isolated from Healthy Donors, including a Triple Hybrid Strain. Antibiotics. 2022; 11(7):833. https://doi.org/10.3390/antibiotics11070833
Chicago/Turabian StyleMéndez-Moreno, Evelyn, Liliana Caporal-Hernandez, Pablo A. Mendez-Pfeiffer, Yessica Enciso-Martinez, Rafael De la Rosa López, Dora Valencia, Margarita M. P. Arenas-Hernández, Manuel G. Ballesteros-Monrreal, and Edwin Barrios-Villa. 2022. "Characterization of Diarreaghenic Escherichia coli Strains Isolated from Healthy Donors, including a Triple Hybrid Strain" Antibiotics 11, no. 7: 833. https://doi.org/10.3390/antibiotics11070833
APA StyleMéndez-Moreno, E., Caporal-Hernandez, L., Mendez-Pfeiffer, P. A., Enciso-Martinez, Y., De la Rosa López, R., Valencia, D., Arenas-Hernández, M. M. P., Ballesteros-Monrreal, M. G., & Barrios-Villa, E. (2022). Characterization of Diarreaghenic Escherichia coli Strains Isolated from Healthy Donors, including a Triple Hybrid Strain. Antibiotics, 11(7), 833. https://doi.org/10.3390/antibiotics11070833