
Current Status of Staphylococcal Cassette Chromosome mec (SCCmec)


Abstract
1. Introduction
2. International Working Group on the Classification of Staphylococcal Cassette Chromosome Elements (IWG-SCC)
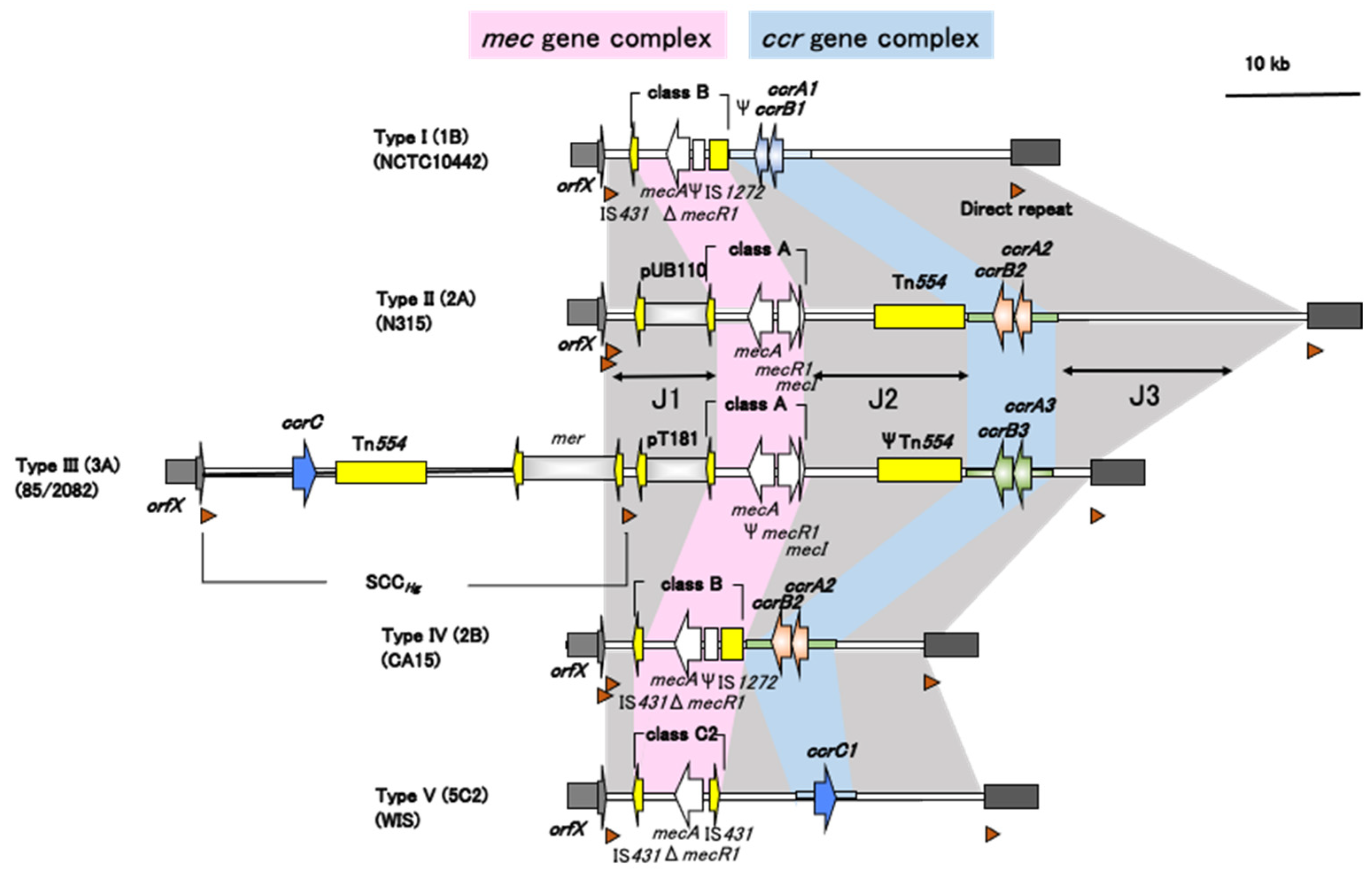
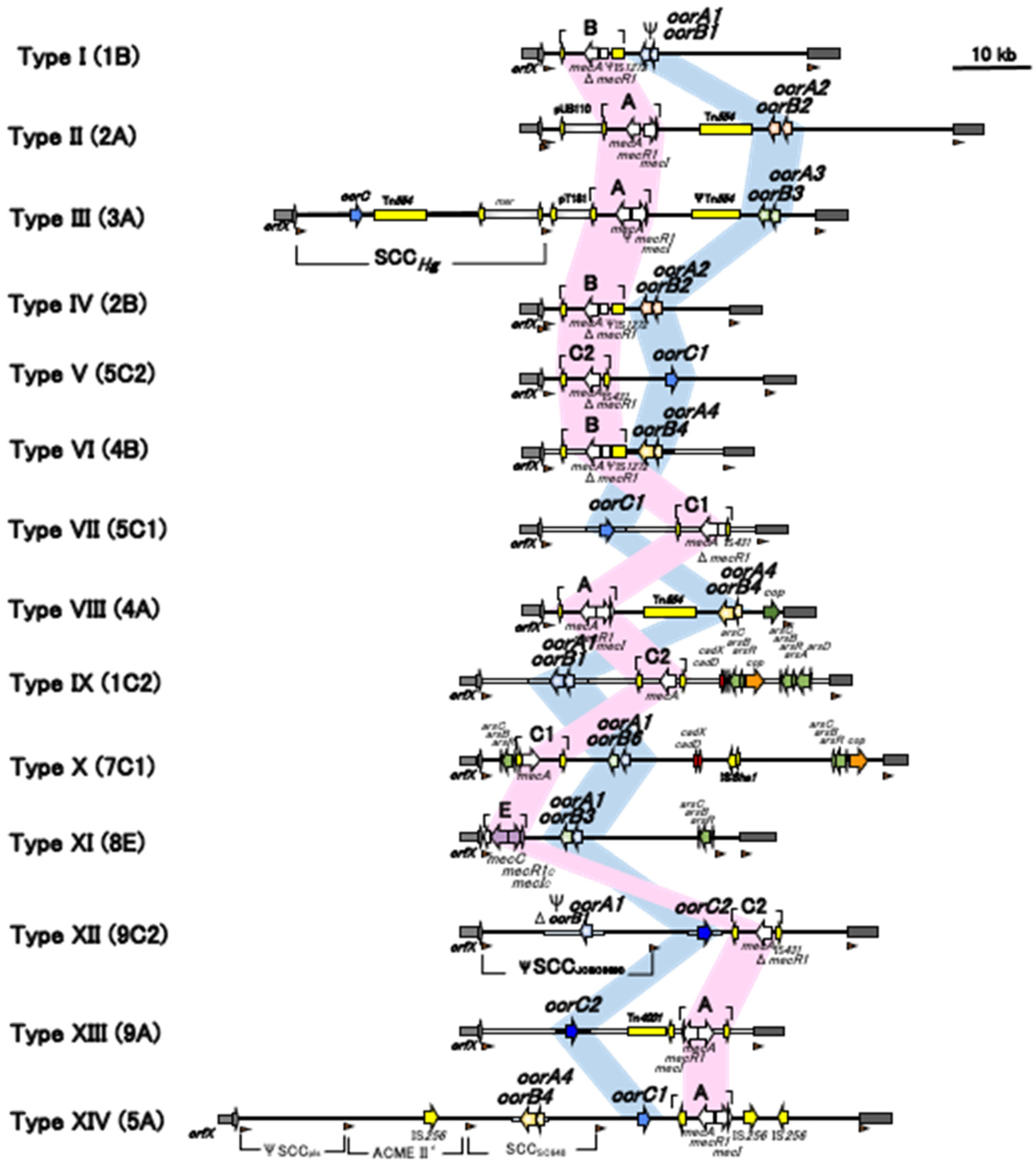
3. Structure of SCCmec
3.1. Requirement for Defining SCCmec
3.2. ccr Gene Complex
3.3. mec Gene Complex
3.4. J Regions
4. Nomenclature of SCCmec and Its Subtypes
5. Current SCCmec Types and Subtypes
6. Methods to Identify SCCmec Types and Subtypes
6.1. PCR-Based Methods
6.2. Whole-Genome Sequencing-Based Methods
7. How to Report New SCCmec and SCCmec Subtypes in the Era of Whole-Genome Sequencing: Role of IWG-SCC
8. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jevons, M.P. “Celbenin”—Resistant Staphylococci. Br. Med. J. 1961, 1, 124–125. [Google Scholar] [CrossRef]
- Centers for Disease Control and Prevention. Brief History of Resistance and Antibiotics. Available online: https://www.cdc.gov/drugresistance/about.html#anchor_1552062951754 (accessed on 14 December 2021).
- Johnson, A.P.; Pearson, A.; Duckworth, G. Surveillance and epidemiology of MRSA bacteraemia in the UK. J Antimicrob. Chemother. 2005, 56, 455–462. [Google Scholar] [CrossRef] [PubMed]
- Horner, C.; Mushtaq, S.; Allen, M.; Longshaw, C.; Reynolds, R.; Livermore, D.M. Are resistance rates among bloodstream isolates a good proxy for other infections? Analysis from the BSAC resistance surveillance programme. J. Antimicrob. Chemother. 2021, 76, 1822–1831. [Google Scholar] [CrossRef] [PubMed]
- Centers for Disease Control and Prevention (CDC). Antibiotic/ Antimicrobial Resistance (AR/AMR) U.S. Nation Action Plan. 2013. Available online: https://www.cdc.gov/drugresistance/us-activities/national-action-plan.html (accessed on 1 January 2022).
- The Japan Goverment. National Action Plan on Antimicrobial Resistance (AMR) 2016–2020. 2016. Available online: https://www.mhlw.go.jp/file/06-Seisakujouhou-10900000-Kenkoukyoku/0000138942.pdf (accessed on 1 January 2022).
- Ubukata, K.; Yamashita, N.; Konno, M. Occurrence of a beta-lactam-inducible penicillin-binding protein in methicillin-resistant staphylococci. Antimicrob. Agents Chemother. 1985, 27, 851–857. [Google Scholar] [CrossRef]
- Utsui, Y.; Yokota, T. Role of an altered penicillin-binding protein in methicillin- and cephem-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 1985, 28, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Matsuhashi, M.; Song, M.D.; Ishino, F.; Wachi, M.; Doi, M.; Inoue, M.; Ubukata, K.; Yamashita, N.; Konno, M. Molecular cloning of the gene of a penicillin-binding protein supposed to cause high resistance to beta-lactam antibiotics in Staphylococcus aureus. J. Bacteriol. 1986, 167, 975–980. [Google Scholar] [CrossRef]
- Hiramatsu, K.; Asada, K.; Suzuki, E.; Okonogi, K.; Yokota, T. Molecular cloning and nucleotide sequence determination of the regulator region of mecA gene in methicillin-resistant Staphylococcus aureus (MRSA). FEBS Lett. 1992, 298, 133–136. [Google Scholar] [CrossRef]
- Ito, T.; Katayama, Y.; Hiramatsu, K. Cloning and nucleotide sequence determination of the entire mec DNA of pre-methicillin-resistant Staphylococcus aureus N315. Antimicrob. Agents Chemother. 1999, 43, 1449–1458. [Google Scholar] [CrossRef]
- Katayama, Y.; Ito, T.; Hiramatsu, K. A new class of genetic element, staphylococcus cassette chromosome mec, encodes methicillin resistance in Staphylococcus aureus. Antimicrob. Agents Chemother. 2000, 44, 1549–1555. [Google Scholar] [CrossRef]
- Ito, T.; Katayama, Y.; Asada, K.; Mori, N.; Tsutsumimoto, K.; Tiensasitorn, C.; Hiramatsu, K. Structural comparison of three types of staphylococcal cassette chromosome mec integrated in the chromosome in methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2001, 45, 1323–1336. [Google Scholar] [CrossRef]
- Kuroda, M.; Ohta, T.; Uchiyama, I.; Baba, T.; Yuzawa, H.; Kobayashi, I.; Cui, L.; Oguchi, A.; Aoki, K.; Nagai, Y.; et al. Whole genome sequencing of meticillin-resistant Staphylococcus aureus. Lancet 2001, 357, 1225–1240. [Google Scholar] [CrossRef]
- Ito, T.; Tsubakishita, S.; Kuwahara-Arai, K.; Han, X.; Hiramatsu, K. Staphylococcal cassette chromosome (SCC): A unique gene transfer system in staphylococci. In Bacterial Integrative Mobile Genetic Elements; Roberts, A., Mullany, P., Eds.; Landes Bioscience: Austin, TX, USA, 2013; pp. 1–15. [Google Scholar]
- Harada, S.; Aoki, K.; Okamoto, K.; Kinoshita, O.; Nawata, K.; Ishii, Y.; Tateda, K.; Sasaki, M.; Saga, T.; Doi, Y.; et al. Left ventricular assist device-associated endocarditis involving multiple clones of Staphylococcus aureus with distinct antimicrobial susceptibility patterns. Int. J. Infect. Dis. 2019, 84, 44–47. [Google Scholar] [CrossRef] [PubMed]
- International Working Group on the Classification of Staphylococcal Cassette Chromosome. Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Hiramatsu, K.; Tomasz, A.; de Lencastre, H.; Perreten, V.; Holden, M.T.; Coleman, D.C.; Goering, R.; Giffard, P.M.; Skov, R.L.; et al. Guidelines for reporting novel mecA gene homologues. Antimicrob. Agents Chemother. 2012, 56, 4997–4999. [Google Scholar] [CrossRef] [PubMed]
- García-Álvarez, L.; Holden, M.T.G.; Lindsay, H.; Webb, C.R.; Brown, D.F.J.; Curran, M.D.; Walpole, E.; Brooks, K.; Pickard, D.J.; Teale, C.; et al. Meticillin-resistant Staphylococcus aureus with a novel mecA homologue in human and bovine populations in the UK and Denmark: A descriptive study. Lancet Infect. Dis. 2011, 11, 595–603. [Google Scholar] [CrossRef]
- Tsubakishita, S.; Kuwahara-Arai, K.; Sasaki, T.; Hiramatsu, K. Origin and molecular evolution of the determinant of methicillin resistance in staphylococci. Antimicrob. Agents Chemother. 2010, 54, 4352–4359. [Google Scholar] [CrossRef]
- Ma, X.X.; Ito, T.; Tiensasitorn, C.; Jamklang, M.; Chongtrakool, P.; Boyle-Vavra, S.; Daum, R.S.; Hiramatsu, K. Novel type of staphylococcal cassette chromosome mec identified in community-acquired methicillin-resistant Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2002, 46, 1147–1152. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Ma, X.X.; Takeuchi, F.; Okuma, K.; Yuzawa, H.; Hiramatsu, K. Novel type V staphylococcal cassette chromosome mec driven by a novel cassette chromosome recombinase, ccrC. Antimicrob. Agents Chemother. 2004, 48, 2637–2651. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, D.C.; Milheirico, C.; de Lencastre, H. Redefining a structural variant of staphylococcal cassette chromosome mec, SCCmec type VI. Antimicrob. Agents Chemother. 2006, 50, 3457–3459. [Google Scholar] [CrossRef] [PubMed]
- Berglund, C.; Ito, T.; Ikeda, M.; Ma, X.X.; Soderquist, B.; Hiramatsu, K. Novel type of staphylococcal cassette chromosome mec in a methicillin-resistant Staphylococcus aureus strain isolated in Sweden. Antimicrob. Agents Chemother. 2008, 52, 3512–3516. [Google Scholar] [CrossRef]
- Zhang, K.; McClure, J.A.; Elsayed, S.; Conly, J.M. Novel staphylococcal cassette chromosome mec type, tentatively designated type VIII, harboring class A mec and type 4 ccr gene complexes in a Canadian epidemic strain of methicillin-resistant Staphylococcus aureus. Antimicrob. Agents Chemother. 2009, 53, 531–540. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Li, S.; Skov, R.L.; Han, X.; Larsen, A.R.; Larsen, J.; Sorum, M.; Wulf, M.; Voss, A.; Hiramatsu, K.; Ito, T. Novel types of staphylococcal cassette chromosome mec elements identified in clonal complex 398 methicillin-resistant Staphylococcus aureus strains. Antimicrob. Agents Chemother. 2011, 55, 3046–3050. [Google Scholar] [CrossRef]
- Wu, Z.; Li, F.; Liu, D.; Xue, H.; Zhao, X. Novel type XII staphylococcal cassette chromosome mec harboring a new cassette chromosome recombinase, CcrC2. Antimicrob. Agents Chemother. 2015, 59, 7597–7601. [Google Scholar] [CrossRef]
- Baig, S.; Johannesen, T.B.; Overballe-Petersen, S.; Larsen, J.; Larsen, A.R.; Stegger, M. Novel SCCmec type XIII (9A) identified in an ST152 methicillin-resistant Staphylococcus aureus. Infect. Genet. Evol. 2018, 61, 74–76. [Google Scholar] [CrossRef] [PubMed]
- Urushibara, N.; Aung, M.S.; Kawaguchiya, M.; Kobayashi, N. Novel staphylococcal cassette chromosome mec (SCCmec) type XIV (5A) and a truncated SCCmec element in SCC composite islands carrying speG in ST5 MRSA in Japan. J. Antimicrob. Chemother. 2020, 75, 46–50. [Google Scholar] [CrossRef]
- Shore, A.; Rossney, A.S.; Keane, C.T.; Enright, M.C.; Coleman, D.C. Seven novel variants of the staphylococcal chromosomal cassette mec in methicillin-resistant Staphylococcus aureus isolates from Ireland. Antimicrob. Agents Chemother. 2005, 49, 2070–2083. [Google Scholar] [CrossRef]
- Han, X.; Ito, T.; Takeuchi, F.; Ma, X.X.; Takasu, M.; Uehara, Y.; Oliveira, D.C.; de Lencastre, H.; Hiramatsu, K. Identification of a novel variant of staphylococcal cassette chromosome mec, type II.5, and its truncated form by insertion of putative conjugative transposon Tn6012. Antimicrob. Agents Chemother. 2009, 53, 2616–2619. [Google Scholar] [CrossRef]
- Oliveira, D.C.; Milheirico, C.; Vinga, S.; de Lencastre, H. Assessment of allelic variation in the ccrAB locus in methicillin-resistant Staphylococcus aureus clones. J. Antimicrob. Chemother. 2006, 58, 23–30. [Google Scholar] [CrossRef]
- Hisata, K.; Kuwahara-Arai, K.; Yamanoto, M.; Ito, T.; Nakatomi, Y.; Cui, L.; Baba, T.; Terasawa, M.; Sotozono, C.; Kinoshita, S.; et al. Dissemination of methicillin-resistant Staphylococci among healthy Japanese children. J. Clin. Microbiol. 2005, 43, 3364–3372. [Google Scholar] [CrossRef]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [PubMed]
- Ma, X.X.; Ito, T.; Chongtrakool, P.; Hiramatsu, K. Predominance of clones carrying Panton-Valentine leukocidin genes among methicillin-resistant Staphylococcus aureus strains isolated in Japanese hospitals from 1979 to 1985. J. Clin. Microbiol. 2006, 44, 4515–4527. [Google Scholar] [CrossRef] [PubMed]
- Kwon, N.H.; Park, K.T.; Moon, J.S.; Jung, W.K.; Kim, S.H.; Kim, J.M.; Hong, S.K.; Koo, H.C.; Joo, Y.S.; Park, Y.H. Staphylococcal cassette chromosome mec (SCCmec) characterization and molecular analysis for methicillin-resistant Staphylococcus aureus and novel SCCmec subtype IVg isolated from bovine milk in Korea. J. Antimicrob. Chemother. 2005, 56, 624–632. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Milheirico, C.; Oliveira, D.C.; de Lencastre, H. Multiplex PCR strategy for subtyping the staphylococcal cassette chromosome mec type IV in methicillin-resistant Staphylococcus aureus: ‘SCCmec IV multiplex’. J. Antimicrob. Chemother. 2007, 60, 42–48. [Google Scholar] [CrossRef]
- Berglund, C.; Ito, T.; Ma, X.X.; Ikeda, M.; Watanabe, S.; Soderquist, B.; Hiramatsu, K. Genetic diversity of methicillin-resistant Staphylococcus aureus carrying type IV SCCmec in Orebro County and the western region of Sweden. J. Antimicrob. Chemother. 2009, 63, 32–41. [Google Scholar] [CrossRef] [PubMed]
- Iwao, Y.; Ishii, R.; Tomita, Y.; Shibuya, Y.; Takano, T.; Hung, W.C.; Higuchi, W.; Isobe, H.; Nishiyama, A.; Yano, M.; et al. The emerging ST8 methicillin-resistant Staphylococcus aureus clone in the community in Japan: Associated infections, genetic diversity, and comparative genomics. J. Infect. Chemother. 2012, 18, 228–240. [Google Scholar] [CrossRef]
- Hosoya, S.; Ito, T.; Misawa, S.; Yoshiike, T.; Oguri, T.; Hiramatsu, T. MRSA clones identified in outpatients dermatology clinic. J. Jpn. Assoc. Intectious Dis. 2014, 88, 840–848. (In Japanese) [Google Scholar]
- Harris, T.M.; Bowen, A.C.; Holt, D.C.; Sarovich, D.S.; Stevens, K.; Currie, B.J.; Howden, B.P.; Carapetis, J.R.; Giffard, P.M.; Tong, S.Y.C. Investigation of trimethoprim/sulfamethoxazole resistance in an emerging sequence type 5 methicillin-resistant Staphylococcus aureus clone reveals discrepant resistance reporting. Clin. Microbiol. Infect. 2018, 24, 1027–1029. [Google Scholar] [CrossRef]
- Hisata, K.; Ito, T.; Matsunaga, N.; Komatsu, M.; Jin, J.; Li, S.; Watanabe, S.; Shimizu, T.; Hiramatsu, K. Dissemination of multiple MRSA clones among community-associated methicillin-resistant Staphylococcus aureus infections from Japanese children with impetigo. J. Infect. Chemother. 2011, 17, 609–621. [Google Scholar] [CrossRef]
- Zhang, K.; McClure, J.A.; Conly, J.M. Enhanced multiplex PCR assay for typing of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. Mol. Cell Probes 2012, 26, 218–221. [Google Scholar] [CrossRef]
- Zhang, K.; McClure, J.A.; Elsayed, S.; Louie, T.; Conly, J.M. Novel multiplex PCR assay for characterization and concomitant subtyping of staphylococcal cassette chromosome mec types I to V in methicillin-resistant Staphylococcus aureus. J. Clin. Microbiol. 2005, 43, 5026–5033. [Google Scholar] [CrossRef]
- Milheirico, C.; Oliveira, D.C.; de Lencastre, H. Update to the multiplex PCR strategy for assignment of mec element types in Staphylococcus aureus. Antimicrob. Agents Chemother. 2007, 51, 3374–3377. [Google Scholar] [CrossRef] [PubMed]
- Takadama, S.; Nakaminami, H.; Takii, T.; Noguchi, N. Identification and detection of USA300 methicillin-resistant Staphylococcus aureus clones with a partial deletion in the ccrB2 gene on the type IV SCCmec element. Diagn. Microbiol. Infect. Dis. 2019, 94, 86–87. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, T.; Ono, D.; Sato, A. Staphylococcal cassette chromosome mec (SCCmec) analysis of MRSA. Methods Mol. Biol. 2020, 2069, 59–78. [Google Scholar] [CrossRef] [PubMed]
- Kaya, H.; Hasman, H.; Larsen, J.; Stegger, M.; Johannesen, T.B.; Allesoe, R.L.; Lemvigh, C.K.; Aarestrup, F.M.; Lund, O.; Larsen, A.R. SCCmecFinder, a web-based tool for typing of staphylococcal cassette chromosome mec in Staphylococcus aureus using whole-genome sequence data. mSphere 2018, 3, e00612-17. [Google Scholar] [CrossRef]
- McClure, J.A.; Conly, J.M.; Obasuyi, O.; Ward, L.; Ugarte-Torres, A.; Louie, T.; Zhang, K. A novel assay for detection of methicillin-resistant Staphylococcus aureus directly from clinical samples. Front. Microbiol. 2020, 11, 1295. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Prototype Strain | Reported mec Gene Name | New mec Gene Name | % Identity with the mecA Gene in Staphylococcus aureus N315 |
|---|---|---|---|
| Staphylococcus aureus N315 | mecA | mecA | 100 (reference gene) |
| Staphylococcal sciuri K11 | mecA (mecA1) | mecA1 | 79.1 |
| Staphylococcus vitulinus CSBO8 | mecA | mecA2 | 91 |
| Macrococcus caseolyticus JCSC5402 | mecAm | mecB | 61.6 |
| Staphylococcus aureus LGA251 | mecALGA251 | mecC | 68.7 |
| SCCmec Type | Ccr Complex Type | mec Complex Class | Representative Strain (GenBank Accession No. or NCBI Reference Sequence No.) | Source | Country | Reported Time | Reference |
|---|---|---|---|---|---|---|---|
| I (1B) | 1 (A1B1) | B | NCTC10442(AB033763) | human | United Kingdom, etc. | 2001 | [13] |
| II (2A) | 2 (A2B2) | A | N315 (D86934) | human | Japan | 2000, 2001 | [12,13] |
| III (3A) | 3 (A3B3) | A | 85/2082 (AB037671) | human | New Zealand, etc. | 2001 | [13] |
| IV (2B) | 2 (A2B2) | B | CA05(AB063172) | human | Japan | 2002 | [21] |
| V (5C2) | 5 (C1) | C2 | WIS(WBG8318) (AB121219) | human | Australia | 2004 | [22] |
| VI (4B) | 4 (A4B4) | B | HDE288 (AF411935) | human | Portugal | 2006 | [23] |
| VII (5C1) | 5 (C1) | C1 | P5747/2002(AB373032) | human | Sweden | 2008 | [24] |
| VIII (4A) | 4 (A4B4) | A | C10682 (FJ390057) | human | Canada | 2009 | [25] |
| IX (1C2) | 1(A1B1) | C2 | JCSC6943 (AB505628) | human (veterinarian) | Thailand | 2011 | [26] |
| X (7C1) | 7(A1B6) | C1 | JCSC6945 (AB505630) | human (veterinarian) | Canada | 2011 | [26] |
| XI (8E) | 8(A1B3) | E | LGA251(FR821779, WGS) | bulk milk, daily cattle, human | England, Ireland, Denmark | 2011 | [19] |
| XII (9C2) | 9(C2) | C2 | BA01611 (KR187111) | cow | China | 2015 | [27] |
| XIII (9A) | 9(C2) | A | 55-99-44 (MG674089) | human | Denmark | 2018 | [28] |
| XIV (5A) | 5 (C1) | A | SC792 (LC440647) | human | Japan | 2019 | [29] |
| SCCmec Subtype | Representative Strain (GenBank Accession No. or NCBI Reference Sequence No.) | Source | Country | Reported Time | Subtyping by | Reference |
|---|---|---|---|---|---|---|
| SCCmec I subtype | ||||||
| a | NCTC10442(AB033763) | human | United Kingdom, etc. | 2001 | (Reference) | [13] |
| b | PL72 (AB433542) | human | Poland | 2006 | J1 | [31,32] |
| SCCmec II subtype | ||||||
| a | N315 (NC_002745), Mu50 (NC_002758), MRSA252 (BX571856), JH1(NC_009632) | human | Japan | 2001 | (Reference) | [12,13] |
| b | JCSC3063(AB127982) | human | Japan | 2005 | J1 | [33] |
| c | AR13.1/3330.2(AJ810120) | human | Ireland | 2005 | J1 | [30] |
| d | RN7170 (AB261975: only J1 region) | human | United States (not clearly described) | 2006 | J1 | [34] |
| e | JCSC6833(AB435013) | human | Japan | 2009 | J1 | [31] |
| SCCmec IV subtype | ||||||
| a | CA05(AB063172) | human | Japan | 2002 | (Reference) | [21] |
| b | 8/6-3P (AB063173) | human | Japan | 2002 | J1 | [21] |
| c | 81/108 (AB096217) | human | Japan | 2004 | J1 | [35] |
| d | JCSC4469 (AB097677) | human | Japan | 2004 | J1 | [35] |
| g | M03-68 (DQ106887) | bovine milk | Korea | 2005 | J1 | [36] |
| h | HO 5096 0412(EMRSA15) (HE681097) | human | Portugal, Greece, Finland | 2007 | J1 | [37] |
| i | JCSC6668 (=CCUG41764)(AB425823) | human | Sweden | 2009 | J1 | [38] |
| j | JCSC6670 (=CCUG27050) (AB425824) | human | Sweden | 2009 | J1 | [38] |
| k | 45394F(GU122149) | human (not clearly described) | Netherlands (not clearly described) | 2010 | J1 | - |
| l | NN50 (AB633329) | human | Japan | 2012 | J1 | [39] |
| m | JCSC8843 (AB872254) | human | Japan | 2014 | J1 | [40] |
| n | No strain name found (KX385846.1) | Australia | 2016 | J3 | - | |
| o | No strain/accession number found in GenBank | human | Australia | 2018 | JI, J3 | [41] |
| SCCmec V subtype | ||||||
| a (5C2) | WIS(WBG8318) (AB121219) | human | Australia | 2004 | (Reference) | [22] |
| b (5C2&5) | TSGH17(=JCSC7190) (AB512767), PM1(AB462393), JCSC5952(AB478780) | human | Japan, Taiwan | 2011 | J1 | [42] |
| c (5C2&5) | S0385(AM990992), JCSC6944(AB505629) | human (veterinarian) | (international pig conference) | 2011 | J1 | [26] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Uehara, Y. Current Status of Staphylococcal Cassette Chromosome mec (SCCmec). Antibiotics 2022, 11, 86. https://doi.org/10.3390/antibiotics11010086
Uehara Y. Current Status of Staphylococcal Cassette Chromosome mec (SCCmec). Antibiotics. 2022; 11(1):86. https://doi.org/10.3390/antibiotics11010086
Chicago/Turabian StyleUehara, Yuki. 2022. "Current Status of Staphylococcal Cassette Chromosome mec (SCCmec)" Antibiotics 11, no. 1: 86. https://doi.org/10.3390/antibiotics11010086
APA StyleUehara, Y. (2022). Current Status of Staphylococcal Cassette Chromosome mec (SCCmec). Antibiotics, 11(1), 86. https://doi.org/10.3390/antibiotics11010086