
A New Twist: The Combination of Sulbactam/Avibactam Enhances Sulbactam Activity against Carbapenem-Resistant Acinetobacter baumannii (CRAB) Isolates



 , and
, and 
Abstract
1. Introduction
2. Results
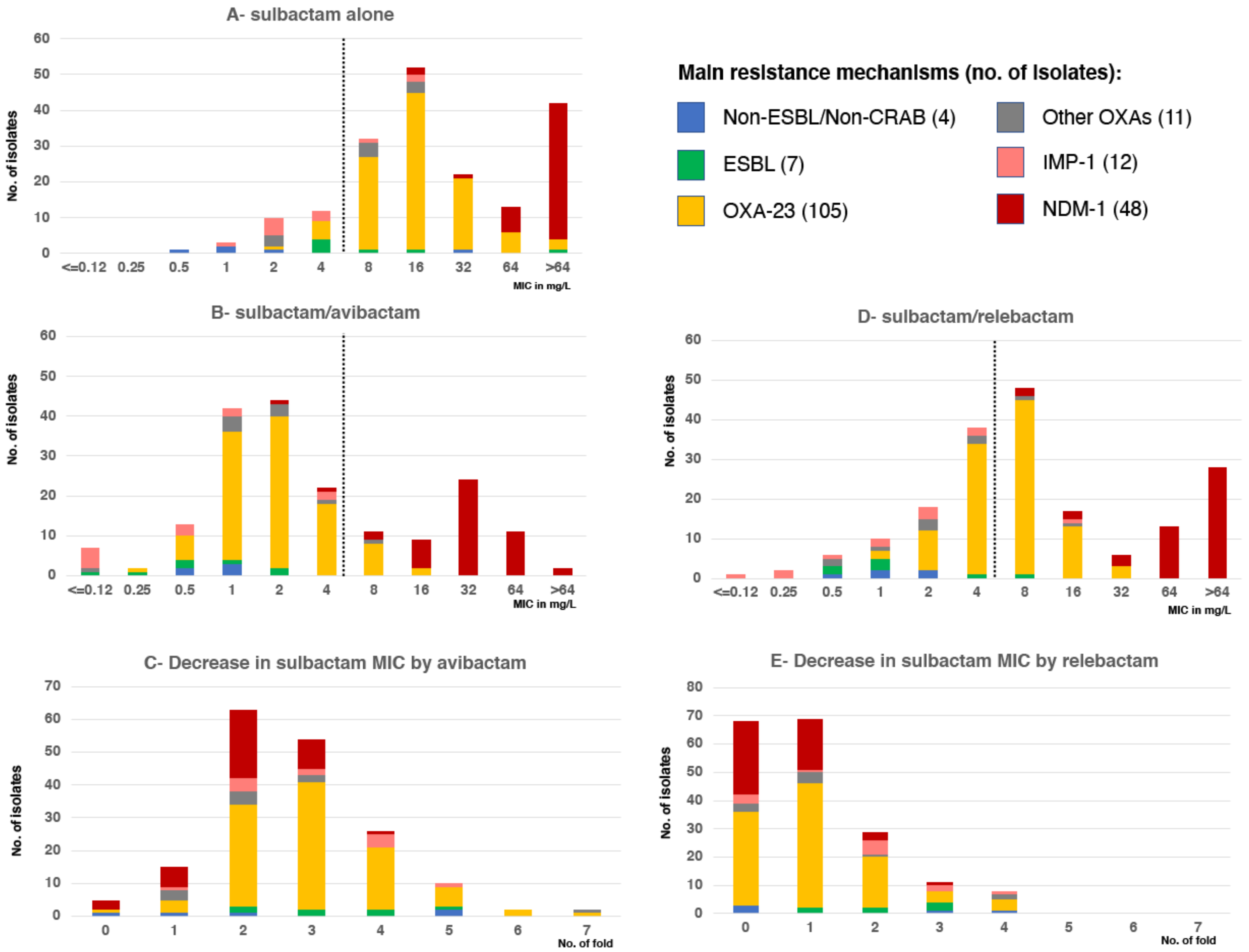
2.1. Sulbactam/Avibactam Susceptibility Testing of Acinetobacter spp. Clinical Strains
2.2. Sulbactam/Relebactam Susceptibility Testing of Acinetobacter spp. Clinical Strains
2.3. Sulbactam/Avibactam Susceptibility Testing of A. baumannii Knockout Strains
- i.
- Mutations affecting the activity of sulbactam alone: when comparing sulbactam MIC in the wild-type strains and isogenic mutants, a decrease in MIC values from 24 to 3 mg/L was observed in the AB5075ΔmrcB and AB5075ΔmreB, suggesting a potential role in sulbactam susceptibility (interaction). The mrcB and mreB genes, which code for a transglycosylase-transpeptidase and a cytoskeletal protein, respectively, were previously reported as genes involved in the formation of the peptidoglycan synthesis. Potentiation of sulbactam activity by avibactam is also lost in these mutant strains. Additionally, the A. baumannii AB5075ΔPBPG MIC of sulbactam is 1.5-fold lower than that measured for A. baumannii AB5075. PBPs are known to be sulbactam targets [36]. Previous reports demonstrated that the antibacterial activity of sulbactam was mediated through inhibition of PBP1 and PBP3.
- ii.
- Mutations that result in loss of avibactam enhancement of sulbactam activity: a 1 or 0-fold decrease in the MIC values comparing sulbactam with sulbactam/avibactam MICs was observed in seven A. baumannii knockout strains (AB5075ΔbfmR, AB5075ΔPBPG, AB5075ΔadvA, AB5075ΔadeB, AB5075ΔmrcB, AB5075ΔxerD, and AB5075ΔmreB). These results identified potential sulbactam and/or sulbactam/avibactam targets (Table 1).
- iii.
- Mutations that do not affect susceptibility to sulbactam or potentiation of sulbactam by avibactam: ≤2-fold decrease in the MIC was observed in twelve A. baumannii AB5075 knockout strains comparing sulbactam with sulbactam/avibactam MIC. We can rule out that these mutated genes are involved in the mechanisms of action (or interaction) of sulbactam and avibactam.
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains
4.2. Antimicrobial Susceptibility Assay
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Boucher, H.W.; Talbot, G.H.; Bradley, J.S.; Edwards, J.E.; Gilbert, D.; Rice, L.B.; Scheld, M.; Spellberg, B.; Bartlett, J. Bad bugs, no drugs: No ESKAPE! An update from the Infectious Diseases Society of America. Clin. Infect. Dis. 2009, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Rice, L.B. Federal funding for the study of antimicrobial resistance in nosocomial pathogens: No ESKAPE. J. Infect. Dis 2008, 197, 1079–1081. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Brown, S.; Hamouda, A.; Findlay, J.; Amyes, S.G. Eleven novel OXA-51-like enzymes from clinical isolates of Acinetobacter baumannii. Clin. Microbiol. Infect. 2007, 13, 1137–1138. [Google Scholar] [CrossRef] [PubMed]
- Evans, B.A.; Amyes, S.G. OXA beta-lactamases. Clin. Microbiol. Rev. 2014, 27, 241–263. [Google Scholar] [CrossRef]
- Bush, K.; Bradford, P.A. Epidemiology of beta-lactamase-producing pathogens. Clin. Microbiol. Rev. 2020, 33. [Google Scholar] [CrossRef]
- Heritier, C.; Dubouix, A.; Poirel, L.; Marty, N.; Nordmann, P. A nosocomial outbreak of Acinetobacter baumannii isolates expressing the carbapenem-hydrolysing oxacillinase OXA-58. J. Antimicrob. Chemother. 2005, 55, 115–118. [Google Scholar] [CrossRef]
- Jeannot, K.; Diancourt, L.; Vaux, S.; Thouverez, M.; Ribeiro, A.; Coignard, B.; Courvalin, P.; Brisse, S. Molecular epidemiology of carbapenem non-susceptible Acinetobacter baumannii in France. PLoS ONE 2014, 9, e115452. [Google Scholar] [CrossRef]
- Rodriguez, C.H.; Nastro, M.; Famiglietti, A. Carbapenemases in Acinetobacter baumannii. Review of their dissemination in Latin America. Rev. Argent. Microbiol. 2018, 50, 327–333. [Google Scholar] [CrossRef]
- Ramirez, M.S.; Bonomo, R.A.; Tolmasky, M.E. Carbapenemases: Transforming Acinetobacter baumannii into a Yet More Dangerous Menace. Biomolecules 2020, 10, 720. [Google Scholar] [CrossRef]
- Yang, J.; Chen, Y.; Jia, X.; Luo, Y.; Song, Q.; Zhao, W.; Wang, Y.; Liu, H.; Zheng, D.; Xia, Y.; et al. Dissemination and characterization of NDM-1-producing Acinetobacter pittii in an intensive care unit in China. Clin. Microbiol. Infect. 2012, 18, E506–E513. [Google Scholar] [CrossRef]
- Tran, D.N.; Tran, H.H.; Matsui, M.; Suzuki, M.; Suzuki, S.; Shibayama, K.; Pham, T.D.; Van Phuong, T.T.; Dang, D.A.; Trinh, H.S.; et al. Emergence of New Delhi metallo-beta-lactamase 1 and other carbapenemase-producing Acinetobacter calcoaceticus-baumannii complex among patients in hospitals in Ha Noi, Viet Nam. Eur. J. Clin. Microbiol. Infect. Dis. 2017, 36, 219–225. [Google Scholar] [CrossRef]
- Pillonetto, M.; Arend, L.; Vespero, E.C.; Pelisson, M.; Chagas, T.P.; Carvalho-Assef, A.P.; Asensi, M.D. First report of NDM-1-producing Acinetobacter baumannii sequence type 25 in Brazil. Antimicrob. Agents Chemother. 2014, 58, 7592–7594. [Google Scholar] [CrossRef]
- El-Mahdy, T.S.; Al-Agamy, M.H.; Al-Qahtani, A.A.; Shibl, A.M. Detection of blaOXA-23-like and blaNDM-1 in Acinetobacter baumannii from the Eastern Region, Saudi Arabia. Microb. Drug Resist. 2017, 23, 115–121. [Google Scholar] [CrossRef]
- Beigverdi, R.; Sattari-Maraji, A.; Emaneini, M.; Jabalameli, F. Status of carbapenem-resistant Acinetobacter baumannii harboring carbapenemase: First systematic review and meta-analysis from Iran. Infect. Genet. Evol. 2019, 73, 433–443. [Google Scholar] [CrossRef]
- Adams, M.D.; Pasteran, F.; Traglia, G.M.; Martinez, J.; Huang, F.; Liu, C.; Fernandez, J.S.; Lopez, C.; Gonzalez, L.J.; Albornoz, E.; et al. Distinct mechanisms of dissemination of NDM-1 metallo- beta-lactamase in Acinetobacter spp. in Argentina. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Butler, D.A.; Biagi, M.; Tan, X.; Qasmieh, S.; Bulman, Z.P.; Wenzler, E. Multidrug Resistant Acinetobacter baumannii: Resistance by Any Other Name Would Still be Hard to Treat. Curr Infect. Dis. Rep. 2019, 21, 46. [Google Scholar] [CrossRef]
- Noguchi, J.K.; Gill, M.A. Sulbactam: A beta-lactamase inhibitor. Clin. Pharm. 1988, 7, 37–51. [Google Scholar]
- Yang, Y.; Xu, Q.; Li, T.; Fu, Y.; Shi, Y.; Lan, P.; Zhao, D.; Chen, Q.; Zhou, Z.; Jiang, Y.; et al. OXA-23 Is a Prevalent Mechanism Contributing to Sulbactam Resistance in Diverse Acinetobacter baumannii Clinical Strains. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Krizova, L.; Poirel, L.; Nordmann, P.; Nemec, A. TEM-1 beta-lactamase as a source of resistance to sulbactam in clinical strains of Acinetobacter baumannii. J. Antimicrob. Chemother. 2013, 68, 2786–2791. [Google Scholar] [CrossRef]
- Chiu, C.H.; Lee, H.Y.; Tseng, L.Y.; Chen, C.L.; Chia, J.H.; Su, L.H.; Liu, S.Y. Mechanisms of resistance to ciprofloxacin, ampicillin/sulbactam and imipenem in Acinetobacter baumannii clinical isolates in Taiwan. Int. J. Antimicrob. Agents 2010, 35, 382–386. [Google Scholar] [CrossRef]
- Paterson, D.L.; Isler, B.; Stewart, A. New treatment options for multiresistant gram negatives. Curr. Opin. Infect. Dis. 2020, 33, 214–223. [Google Scholar] [CrossRef]
- McLeod, S.M.; Moussa, S.H.; Hackel, M.A.; Miller, A.A. In Vitro Activity of Sulbactam-Durlobactam against Acinetobacter baumannii-calcoaceticus Complex Isolates Collected Globally in 2016 and 2017. Antimicrob. Agents Chemother. 2020, 64. [Google Scholar] [CrossRef]
- Lomovskaya, O.; Nelson, K.; Rubio-Aparicio, D.; Tsivkovski, R.; Sun, D.; Dudley, M.N. The Impact of Intrinsic Resistance Mechanisms on Potency of QPX7728, a New Ultra-Broad-Spectrum Beta-lactamase Inhibitor of Serine and Metallo Beta-Lactamases in Enterobacteriaceae, Pseudomonas aeruginosa, and Acinetobacter baumannii. Antimicrob. Agents Chemother. 2020. [Google Scholar] [CrossRef]
- Karaiskos, I.; Galani, I.; Souli, M.; Giamarellou, H. Novel beta-lactam-beta-lactamase inhibitor combinations: Expectations for the treatment of carbapenem-resistant Gram-negative pathogens. Expert Opin. Drug Metab. Toxicol. 2019, 15, 133–149. [Google Scholar] [CrossRef]
- Barnes, M.D.; Kumar, V.; Bethel, C.R.; Moussa, S.H.; O’Donnell, J.; Rutter, J.D.; Good, C.E.; Hujer, K.M.; Hujer, A.M.; Marshall, S.H.; et al. Targeting Multidrug-Resistant Acinetobacter spp.: Sulbactam and the Diazabicyclooctenone beta-Lactamase Inhibitor ETX2514 as a Novel Therapeutic Agent. mBio 2019, 10. [Google Scholar] [CrossRef]
- Zhanel, G.G.; Lawrence, C.K.; Adam, H.; Schweizer, F.; Zelenitsky, S.; Zhanel, M.; Lagace-Wiens, P.R.S.; Walkty, A.; Denisuik, A.; Golden, A.; et al. Imipenem-Relebactam and Meropenem-Vaborbactam: Two Novel Carbapenem-beta-Lactamase Inhibitor Combinations. Drugs 2018, 78, 65–98. [Google Scholar] [CrossRef]
- Wright, H.; Bonomo, R.A.; Paterson, D.L. New agents for the treatment of infections with Gram-negative bacteria: Restoring the miracle or false dawn? Clin. Microbiol. Infect. 2017, 23, 704–712. [Google Scholar] [CrossRef]
- Durand-Reville, T.F.; Guler, S.; Comita-Prevoir, J.; Chen, B.; Bifulco, N.; Huynh, H.; Lahiri, S.; Shapiro, A.B.; McLeod, S.M.; Carter, N.M.; et al. ETX2514 is a broad-spectrum beta-lactamase inhibitor for the treatment of drug-resistant Gram-negative bacteria including Acinetobacter baumannii. Nat. Microbiol. 2017, 2, 17104. [Google Scholar] [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. beta-Lactamases and beta-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Tooke, C.L.; Hinchliffe, P.; Lang, P.A.; Mulholland, A.J.; Brem, J.; Schofield, C.J.; Spencer, J. Molecular Basis of Class A beta-Lactamase Inhibition by Relebactam. Antimicrob. Agents Chemother. 2019, 63. [Google Scholar] [CrossRef]
- Rodriguez, C.H.; Brune, A.; Nastro, M.; Vay, C.; Famiglietti, A. In vitro synergistic activity of the sulbactam/avibactam combination against extensively drug-resistant Acinetobacter baumannii. J Med. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Santimaleeworagun, W.; Wongpoowarak, P.; Chayakul, P.; Pattharachayakul, S.; Tansakul, P.; Garey, K.W. In vitro activity of colistin or sulbactam in combination with fosfomycin or imipenem against clinical isolates of carbapenem-resistant Acinetobacter baumannii producing OXA-23 carbapenemases. Southeast Asian J. Trop. Med. Public Health 2011, 42, 890–900. [Google Scholar] [PubMed]
- Pei, G.; Mao, Y.; Sun, Y. In vitro activity of minocycline alone and in combination with cefoperazone-sulbactam against carbapenem-resistant Acinetobacter baumannii. Microb. Drug Resist 2012, 18, 574–577. [Google Scholar] [CrossRef] [PubMed]
- Poulikakos, P.; Tansarli, G.S.; Falagas, M.E. Combination antibiotic treatment versus monotherapy for multidrug-resistant, extensively drug-resistant, and pandrug-resistant Acinetobacter infections: A systematic review. Eur. J. Clin. Microbiol. Infect. Dis. 2014, 33, 1675–1685. [Google Scholar] [CrossRef]
- Kalin, G.; Alp, E.; Akin, A.; Coskun, R.; Doganay, M. Comparison of colistin and colistin/sulbactam for the treatment of multidrug resistant Acinetobacter baumannii ventilator-associated pneumonia. Infection 2014, 42, 37–42. [Google Scholar] [CrossRef]
- Penwell, W.F.; Shapiro, A.B.; Giacobbe, R.A.; Gu, R.F.; Gao, N.; Thresher, J.; McLaughlin, R.E.; Huband, M.D.; DeJonge, B.L.; Ehmann, D.E.; et al. Molecular mechanisms of sulbactam antibacterial activity and resistance determinants in Acinetobacter baumannii. Antimicrob. Agents Chemother. 2015, 59, 1680–1689. [Google Scholar] [CrossRef]
- Geisinger, E.; Mortman, N.J.; Vargas-Cuebas, G.; Tai, A.K.; Isberg, R.R. A global regulatory system links virulence and antibiotic resistance to envelope homeostasis in Acinetobacter baumannii. PLoS Pathog. 2018, 14, e1007030. [Google Scholar] [CrossRef]
- Geisinger, E.; Mortman, N.J.; Dai, Y.; Cokol, M.; Syal, S.; Farinha, A.; Fisher, D.G.; Tang, A.Y.; Lazinski, D.W.; Wood, S.; et al. Antibiotic susceptibility signatures identify potential antimicrobial targets in the Acinetobacter baumannii cell envelope. Nat. Commun. 2020, 11, 4522. [Google Scholar] [CrossRef]
- Lin, D.L.; Traglia, G.M.; Baker, R.; Sherratt, D.J.; Ramirez, M.S.; Tolmasky, M.E. Functional Analysis of the Acinetobacter baumannii XerC and XerD Site-Specific Recombinases: Potential Role in Dissemination of Resistance Genes. Antibiotics 2020, 9, 405. [Google Scholar] [CrossRef]
- Jacobs, A.C.; Thompson, M.G.; Black, C.C.; Kessler, J.L.; Clark, L.P.; McQueary, C.N.; Gancz, H.Y.; Corey, B.W.; Moon, J.K.; Si, Y.; et al. AB5075, a Highly Virulent Isolate of Acinetobacter baumannii, as a Model Strain for the Evaluation of Pathogenesis and Antimicrobial Treatments. MBio 2014, 5, e01014–e01076. [Google Scholar] [CrossRef]
- Adams, M.D.; Nickel, G.C.; Bajaksouzian, S.; Lavender, H.; Murthy, A.R.; Jacobs, M.R.; Bonomo, R.A. Resistance to colistin in Acinetobacter baumannii associated with mutations in the PmrAB two-component system. Antimicrob. Agents Chemother. 2009, 53, 3628–3634. [Google Scholar] [CrossRef]
- Adams, M.D.; Wright, M.S.; Karichu, J.K.; Venepally, P.; Fouts, D.E.; Chan, A.P.; Richter, S.S.; Jacobs, M.R.; Bonomo, R.A. Rapid Replacement of Acinetobacter baumannii Strains Accompanied by Changes in Lipooligosaccharide Loci and Resistance Gene Repertoire. mBio 2019, 10. [Google Scholar] [CrossRef]
- Clinical and Laboratory Standards Institute. Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Growth Aerobically M7-A10, Ed. M7-A10; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2018. [Google Scholar]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing; M.100, twenty-Nine Informational Supplement; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2020. [Google Scholar]
- Yang, Q.; Xu, Y.; Jia, P.; Zhu, Y.; Zhang, J.; Zhang, G.; Deng, J.; Hackel, M.; Bradford, P.A.; Reinhart, H. In vitro activity of sulbactam/durlobactam against clinical isolates of Acinetobacter baumannii collected in China. J. Antimicrob. Chemother. 2020, 75, 1833–1839. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Strains | MH | MH + AVI 4 mg/L |
|---|---|---|
| AB5075 | 24 | 1.5 |
| AB5075ΔmrcB | 3 | 1.5 |
| AB5075ΔmreB | 3 | 1.5 |
| AB5075ΔPBPG | 8 | 8–12 |
| AB5075ΔbfmR | 24 | 12 |
| AB5075ΔxerD | 16 | 8 |
| AB5075ΔadvA | 16 | 16 |
| AB5075ΔadeB | 16 | 16 |
| AB5075ΔxerC | 18 | 8 |
| AB5075ΔbfmS | 16 | 6 |
| AB5075ΔadeA | 24 | 8 |
| AB5075ΔlpxB | 24 | 8 |
| AB5075ΔmreD | 24 | 8 |
| AB5075ΔdksA | 24 | 8 |
| AB5075ΔrecA | 32 | 12 |
| AB5075ΔadeK | 24 | 6 |
| AB5075ΔampC | 24 | 6 |
| AB5075ΔOXA-69 | 16 | 4 |
| AB5075ΔmreC | 16 | 4 |
| AB5075ΔmrdB | 16 | 4 |
| AB5075ΔmltB | 16 | 1.5–3 |
| AB5075ΔElsL | 24 | 4 |
| AB5075ΔdnaK | 24 | 4 |
| AB5075ΔadeR | 32 | 3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasteran, F.; Cedano, J.; Baez, M.; Albornoz, E.; Rapoport, M.; Osteria, J.; Montaña, S.; Le, C.; Ra, G.; Bonomo, R.A.; et al. A New Twist: The Combination of Sulbactam/Avibactam Enhances Sulbactam Activity against Carbapenem-Resistant Acinetobacter baumannii (CRAB) Isolates. Antibiotics 2021, 10, 577. https://doi.org/10.3390/antibiotics10050577
Pasteran F, Cedano J, Baez M, Albornoz E, Rapoport M, Osteria J, Montaña S, Le C, Ra G, Bonomo RA, et al. A New Twist: The Combination of Sulbactam/Avibactam Enhances Sulbactam Activity against Carbapenem-Resistant Acinetobacter baumannii (CRAB) Isolates. Antibiotics. 2021; 10(5):577. https://doi.org/10.3390/antibiotics10050577
Chicago/Turabian StylePasteran, Fernando, Jose Cedano, Michelle Baez, Ezequiel Albornoz, Melina Rapoport, Jose Osteria, Sabrina Montaña, Casin Le, Grace Ra, Robert A. Bonomo, and et al. 2021. "A New Twist: The Combination of Sulbactam/Avibactam Enhances Sulbactam Activity against Carbapenem-Resistant Acinetobacter baumannii (CRAB) Isolates" Antibiotics 10, no. 5: 577. https://doi.org/10.3390/antibiotics10050577
APA StylePasteran, F., Cedano, J., Baez, M., Albornoz, E., Rapoport, M., Osteria, J., Montaña, S., Le, C., Ra, G., Bonomo, R. A., Tolmasky, M. E., Adams, M., Corso, A., & Ramirez, M. S. (2021). A New Twist: The Combination of Sulbactam/Avibactam Enhances Sulbactam Activity against Carbapenem-Resistant Acinetobacter baumannii (CRAB) Isolates. Antibiotics, 10(5), 577. https://doi.org/10.3390/antibiotics10050577