
Distribution of β-Lactamase Genes in Clinical Isolates from California Central Valley Hospital Deviates from the United States Nationwide Trends

Abstract
1. Introduction
2. Results
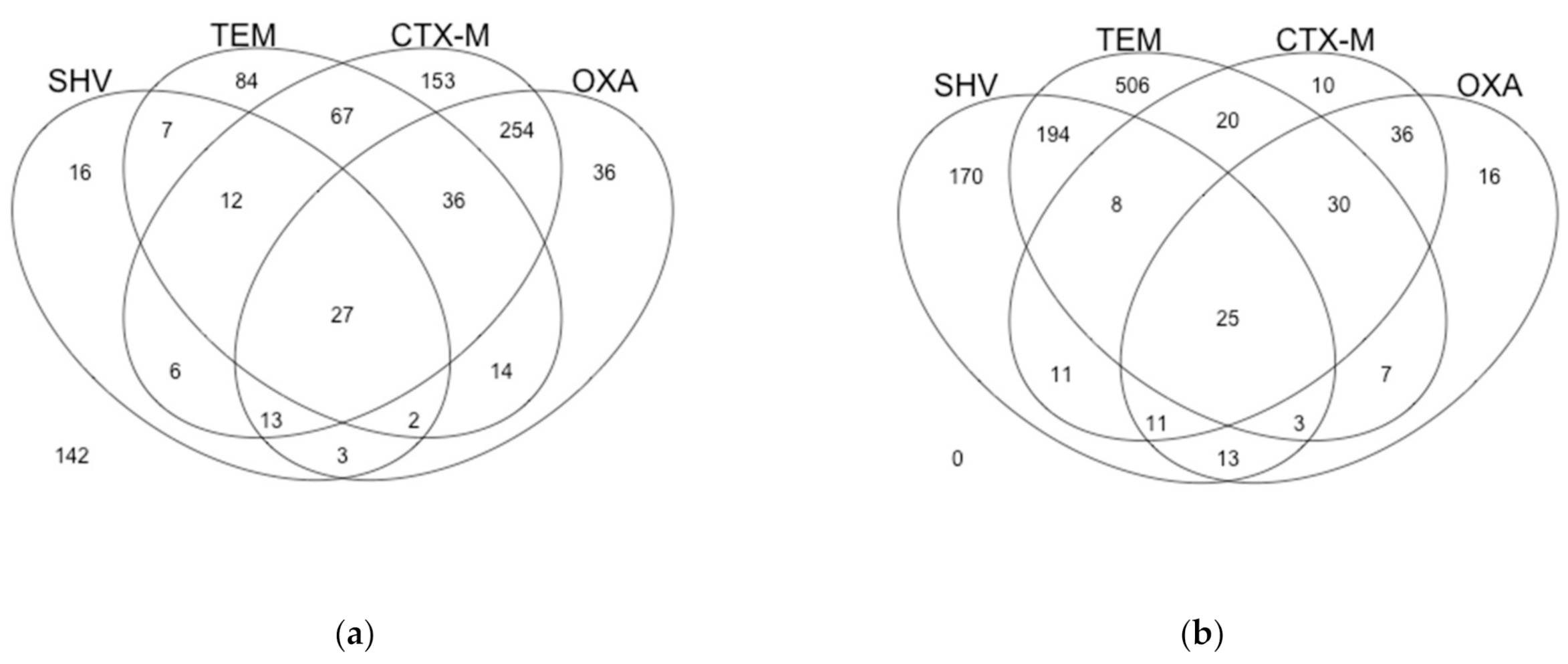
2.1. Regional Gene Frequencies
2.2. U.S. Database Gene Frequencies
2.3. Comparison of DHMMC and U.S. Populations
3. Discussion
4. Materials and Methods
4.1. Hospital Isolates
4.2. Molecular Methods
4.3. U.S. Database
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wilke, M.S.; Lovering, A.L.; Strynadka, N.C. Beta-lactam antibiotic resistance: A current structural perspective. Curr. Opin. Microbiol. 2005, 8, 525–533. [Google Scholar] [CrossRef]
- Pandey, N.; Cascella, M. Beta lactam antibiotics. In StatPearls; StatPearls Publishing: Treasure Island, FL, USA, 2020; Available online: https://www.ncbi.nlm.nih.gov/books/NBK545311/#_NBK545311_pubdet_ (accessed on 28 June 2020).
- Hall, B.G.; Acar, H.; Nandipati, A.; Barlow, M. Growth rates made easy. Mol. Biol Evol. 2014, 1, 232–238. [Google Scholar] [CrossRef]
- Schwaber, M.J.; Schwartz, D.; Giladi, M.; Chmelnitsky, I.; Leavitt, A.; Carmeli, Y. Influx of extended-spectrum b-lactamase—producing enterobacteriaceae into the hospital. Clin. Infect. Dis. 2006, 42, 925–934. [Google Scholar] [CrossRef]
- Datta, N.; Kontomichalou, P. Penicillinase synthesis controlled by infectious r factors in enterobacteriaceae. Nature 1965, 208, 239–241. [Google Scholar] [CrossRef]
- Liakopoulos, A.; Mevius, D.; Ceccarelli, D. A review of shv extended-spectrum β-lactamases: Neglected yet ubiquitous. Front. Microbiol. 2016, 7, 1374. [Google Scholar] [CrossRef]
- Pitton, J.S. Mechanisms of bacterial resistance to antibiotics. Ergeb. Physiol. 1972, 65, 15–93. [Google Scholar] [CrossRef]
- Doi, Y.; Iovleva, A.; Bonomo, R.A. The ecology of extended-spectrum β-lactamases (esbls) in the developed world. J. Travel Med. 2017, 2, S44–S51. [Google Scholar] [CrossRef] [PubMed]
- Cantón, R.; González-Alba, J.M.; Galán, J.C. Ctx-m enzymes: Origin and diffusion. Front. Microbiol. 2012. [Google Scholar] [CrossRef] [PubMed]
- Richmond, M.H.; Sykes, R.B. The β-lactamases of gram-negative bacteria and their possible physiological role. Adv. Microb. Physiol. 1973, 9, 31–88. [Google Scholar] [CrossRef]
- Poirel, L.; Naas, T.; Nordmann, P. Diversity, epidemiology, and genetics of class d β-lactamases. Antimicrob. Agents Chemother. 2010, 54, 24–38. [Google Scholar] [CrossRef] [PubMed]
- Critchley, I.A.; Cotroneo, N.; Pucci, M.J.; Mendes, R. The burden of antimicrobial resistance among urinary tract isolates of escherichia coli in the united states in 2017. PLoS ONE 2019, 14, e0220265. [Google Scholar] [CrossRef]
- Tamma, P.D.; Sharara, S.L.; Pana, Z.D.; Amoah, J.; Fisher, S.L.; Tekle, T.; Doi, Y.; Simner, P.J. Molecular epidemiology of ceftriaxone-nonsusceptible enterobacterales isolates in an academic medical center in the united states. Open Forum Infect. Dis. 2019. [Google Scholar] [CrossRef]
- Mendes, R.E.; Jones, R.N.; Woosley, L.N.; Cattoir, V.; Castanheira, M. Application of next-generation sequencing for characterization of surveillance and clinical trial isolates: Analysis of the distribution of β-lactamase resistance genes and lineage background in the united states. Open Forum Infect. Dis. 2019, 6, S69–S78. [Google Scholar] [CrossRef]
- Markovska, R.; Schneider, I.; Keuleyan, E.; Bauernfeind, A. Extended-spectrum β-lactamase (esbl) ctx-m-15-producing E.coli and Klebsiella pneumoniae in Sofia, Bulgaria. Clin. Microbiol Infect. 2004, 10, 752–755. [Google Scholar] [CrossRef]
- Sugumar, M.; Kumar, K.M.; Manoharan, A.; Anbarasu, A.; Ramaiah, S. Detection of oxa-1 β-lactamase gene of Klebsiella pneumoniae from blood stream infections (bsi) by conventional pcr and in-silico analysis to understand the mechanism of oxa mediated resistance. PLoS ONE 2014, 9, e91800. [Google Scholar] [CrossRef]
- Bajpai, T.; Pandey, M.; Varma, M.; Bhatambare, G.S. Prevalence of tem, shv, and ctx-m beta-lactamase genes in the urinary isolates of a tertiary care hospital. Avicenna J. Med. 2017, 7, 12–16. [Google Scholar] [CrossRef]
- Castanheira, M.; Farrell, S.E.; Deshpande, L.M.; Mendes, R.E.; Jones, R.N. Prevalence of β-lactamase-encoding genes among Enterobacteriaceae bacteremia isolates collected in 26 U.S. Hospitals: Report from the SENTRY Antimicrobial Surveillance Program (2010). Antimicrob. Agents Chemother. 2013, 57, 3012–3020. [Google Scholar] [CrossRef]
- MacLean, R.C.; Hall, A.R.; Perron, G.G.; Buckling, A. The population genetics of antibiotic resistance: Integrating molecular mechanisms and treatment contexts. Nat. Rev. Genet. 2010, 11, 405–414. [Google Scholar] [CrossRef]
- Perron, G.G.; Gonzalez, A.; Buckling, A. Source-sink dynamics shape the evolution of antibiotic resistance and its pleiotropic fitness cost. Proc. Biol. Sci. 2007, 274, 2351–2356. [Google Scholar] [CrossRef]
- Norén, K.; Carmichael, L.; Fuglei, E.; Eide, N.E.; Hersteinsson, P.; Angerbjörn, A. Pulses of movement across the sea ice: Population connectivity and temporal genetic structure in the arctic fox. Oecologia 2011, 166, 973–984. [Google Scholar] [CrossRef]
- Santiago, F.; Doscher, E.; Kim, J.; Camps, M.; Meza, J.; Sindi, S.; Barlow, M. Growth rate assays reveal fitness consequences of β-lactamases. PLoS ONE 2020, 15, e0228240. [Google Scholar] [CrossRef] [PubMed]
- Wagner, C.H. Simpson’s paradox in real life. Am. Stat. 1982, 36, 46–48. [Google Scholar] [CrossRef]
- Woerther, P.L.; Angebault, C.; Lescat, M.; Ruppé, E.; Skurnik, D.; Assiya, E.M.; Clermont, O.; Jacquier, H.; Costa, A.D.; Renard, M.; et al. Emergence and dissemination of extended-spectrum β-lactamase–producing E.coli in the community: Lessons from the study of a remote and controlled population. J. Infect. Dis. 2010, 202, 515–523. [Google Scholar] [CrossRef]
- Kalp, M.; Bethel, C.R.; Bonomo, R.A.; Carey, P.R. Why the extended-spectrum β-lactamases shv-2 and shv-5 are “hypersusceptible” to mechanism-based inhibitors. Biochemistry 2009, 48, 9912–9920. [Google Scholar] [CrossRef]
- Dallenne, C., da Costa; Decré, D.; Favier, C.; Arlet, G. Development of a set of multiplex pcr assays for the detection of genes encoding important β-lactamases in enterobacteriaceae. J. Antimicrob. Chemother. 2010, 65, 490–495. [Google Scholar] [CrossRef]
- O’Leary, N.A.; Wright, M.W.; Brister, J.R.; Ciufo, S.; Haddad, D.; McVeigh, R.; Rajput, B.; Robbertse, B.; Smith-White, B.; Ako-Adjei, D.; et al. Reference sequence (refseq) database at ncbi: Current status, taxonomic expansion, and functional annotation. Nucleic Acids Res. 2016, 44, D733–D745. [Google Scholar] [CrossRef]
- The NCBI Pathogen Detection Project; National Library of Medicine (US), National Center for Biotechnology Information: Bethesda, MD, USA, 2021. Available online: </named-content> https://www.ncbi.nlm.nih.gov/pathogens/isolates/ (accessed on 18 October 2018).
- Alcock, B.P.; Raphenya, A.R.; Lau, T.T.Y.; Tsang, K.K.; Bouchard, M.; Edalatmand, A.; Huynh, W.; Nguyen, A.L.V.; Cheng, A.A.; Liu, S.; et al. Card 2020: Antibiotic resistome surveillance with the comprehensive antibiotic resistance database. Nucleic Acids Res. 2020, 48, D517–D525. [Google Scholar] [CrossRef]
- Chihara, L.M.; Hesterberg, T.C. Mathematical Statistics with Resampling and R, 2nd ed.; John Wiley & Sons: Hoboken, NJ, USA, 2018; ISBN 978-1-119-41653-1. [Google Scholar]
- Griffith, A.J.F.; Miller, J.H.; Suzuki, D.T. Making Recombinant DNA in An. Introduction to Genetic Analysis, 7th ed.; W.H. Freeman: New York, NY, USA, 2000; ISBN 978-0716737711. [Google Scholar]
- Dunlap, W.P.; Brody, C.J.; Greer, T. Canonical correlation and chi-square: Relationships and interpretation. J. Gen. Psychol. 2000, 127, 341–353. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the false discovery rate: A practical and powerful approach to multiple testing. J. R Stat. Soc. Series B Stat. Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Statistics and Machine Learning Toolbox™; Version r2020a; The MathWorks Inc.: Natick, MA, USA, 2020.


{kind=link}
| DHMMC | blaSHV | blaTEM | blaCTX-M | blaOXA |
|---|---|---|---|---|
| 2013 (n = 106) | 9.4 (4.6, 16.7) | 37.7 (28.5, 47.7) | 52.8 (42.9, 62.6) | 52.8 (42.9, 62.6) |
| 2014 (n = 88) | 8.0 (3.3, 15.7) | 29.5 (20.3, 40.2) | 75.0 (64.6, 83.6) | 54.5 (43.6, 65.2) |
| 2015 (n = 255) | 7.5 (4.5, 11.4) | 29.8 (24.3, 35.8) | 71.4 (65.4, 76.8) | 49.8 (43.5, 56.1) |
| 2016 (n = 207) | 12.6 (8.4, 17.9) | 24.6 (18.9, 31.1) | 62.8 (55.8, 69.4) | 35.7 (29.2, 42.7) |
| 2017 (n = 126) | 9.5 (5.0, 16.0) | 23.8 (16.7, 32.2) | 61.1 (52.0, 69.7) | 36.5 (28.1, 45.6) |
| 2018 (n = 90) | 13.3 (7.1, 22.1) | 28.9 (19.8, 39.4) | 63.3 (52.5, 73.2) | 37.8 (27.8, 48.6) |
| Species | Markers | Chi-Square p-Value | PC | PC p-Value |
|---|---|---|---|---|
| All (n = 872) | blaSHV:blaTEM | 3.75 × 10−9 * | 0.20 | 2.74 × 10−9 * |
| blaSHV:blaCTX-M | 6.37 × 10−1 | 0.02 | 6.37 × 10−1 | |
| blaSHV:blaOXA | 1.08 × 10−1 | 0.05 | 1.08 × 10−1 | |
| blaTEM:blaCTX-M | 1.49 × 10−3 * | −0.11 | 1.46 × 10−3 * | |
| blaTEM:blaOXA | 3.00 × 10−6 * | −0.16 | 2.68 × 10−6 * | |
| blaCTX-M:blaOXA | 8.59 × 10−30 * | 0.38 | 5.24 × 10−32 * | |
| E. coli (n = 787) | blaSHV:blaTEM | 5.22 × 10−1 | 0.02 | 5.23 × 10−1 |
| blaSHV:blaCTX-M | 2.58 × 10−1 | −0.04 | 2.59 × 10−1 | |
| blaSHV:blaOXA | 4.89 × 10−1 | −0.02 | 4.89 × 10−1 | |
| blaTEM:blaCTX-M | 1.24 × 10−6 * | −0.17 | 1.07 × 10−6 * | |
| blaTEM:blaOXA | 1.40 × 10−9 * | −0.22 | 9.50 × 10−10 * | |
| blaCTX-M:blaOXA | 9.22 × 10−26 * | 0.37 | 1.53 × 10−27 * | |
| K. pneumoniae (n = 85) | blaSHV:blaTEM | 3.36 × 10−4 * | 0.39 | 2.34 × 10−4 * |
| blaSHV:blaCTX-M | 1.97 × 10−5 * | 0.46 | 8.15 × 10−6 * | |
| blaSHV:blaOXA | 6.84 × 10−3 * | 0.29 | 6.44 × 10−3 * | |
| blaTEM:blaCTX-M | 2.58 × 10−5 * | 0.46 | 1.13 × 10−5 * | |
| blaTEM:blaOXA | 3.84 × 10−2 | 0.22 | 3.88 × 10−2 | |
| blaCTX-M:blaOXA | 5.69 × 10−6 * | 0.49 | 1.72 × 10−6 * |
| Nationwide U.S. | blaSHV | blaTEM | blaCTX-M | blaOXA |
|---|---|---|---|---|
| 2013 (n = 6) | 16.7 (0.4, 64.1) | 83.3 (35.9, 99.6) | 0.0 (0.0, 45.9) | 0.0 (0.0, 45.9) |
| 2014 (n = 179) | 4.5 (1.9, 8.6) | 82.7 (76.3, 87.9) | 14.0 (9.2, 19.9) | 16.8 (11.6, 23.1) |
| 2015 (n = 268) | 66.8 (60.8, 72.4) | 74.6 (69.0, 79.7) | 8.2 (5.2, 12.2) | 9.7 (6.4, 13.9) |
| 2016 (n = 190) | 19.5 (14.1, 25.8) | 84.7 (78.8, 89.5) | 16.8 (11.8, 22.9) | 13.2 (8.7, 18.8) |
| 2017 (n = 251) | 50.6 (44.2, 56.9) | 66.9 (60.7, 72.7) | 13.9 (9.9, 18.9) | 11.6 (7.9, 16.2) |
| 2018 (n = 166) | 50.0 (42.2, 57.8) | 66.9 (59.2, 74.0) | 22.3 (16.2, 29.4) | 18.7 (13.1, 25.4) |
| Species | Markers | Chi-Square p-Value | PC | PC p-Value |
|---|---|---|---|---|
| All (n = 1060) | blaSHV:blaTEM | 7.03 × 10−43 * | −0.42 | 6.29 × 10−47 * |
| blaSHV:blaCTX-M | 2.13 × 10−1 | −0.04 | 2.14 × 10−1 | |
| blaSHV:blaOXA | 2.81 × 10−1 | −0.03 | 2.81 × 10−1 | |
| blaTEM:blaCTX-M | 1.31 × 10−9 * | −0.19 | 9.80 × 10−10 * | |
| blaTEM:blaOXA | 3.31 × 10−17 * | −0.26 | 1.02 × 10−17 * | |
| blaCTX-M:blaOXA | 1.01 × 10−99 * | 0.65 | 7.63 × 10−129 * | |
| E. coli (n = 559) | blaSHV:blaTEM | 2.03 × 10−6 * | −0.20 | 1.68 × 10−6 * |
| blaSHV:blaCTX-M | 1.48 × 10−1 | −0.06 | 1.49 × 10−1 | |
| blaSHV:blaOXA | 1.52 × 10−1 | −0.06 | 1.52 × 10−1 | |
| blaTEM:blaCTX-M | 6.72 × 10−35 * | −0.52 | 2.90 × 10−40 * | |
| blaTEM:blaOXA | 1.85 × 10−64 * | −0.72 | 2.38 × 10−89 * | |
| blaCTX-M:blaOXA | 1.83 × 10−53 * | 0.65 | 1.05 × 10−68 * | |
| K. pneumoniae (n = 501) | blaSHV:blaTEM | 6.81 × 10−10 * | −0.28 | 3.45 × 10−10 * |
| blaSHV:blaCTX-M | 4.69 × 10−5 * | −0.18 | 4.23 × 10−5 * | |
| blaSHV:blaOXA | 7.37 × 10−3 * | −0.12 | 7.30 × 10−3 * | |
| blaTEM:blaCTX-M | 4.73 × 10−1 | 0.03 | 4.74 × 10−1 | |
| blaTEM:blaOXA | 5.68 × 10−1 | 0.03 | 5.69 × 10−1 | |
| blaCTX-M:blaOXA | 4.37 × 10−48 * | 0.65 | 1.04 × 10−61 * |
| Species | Marker | FM | FN | FM—FN | p-Value | 95% CI |
|---|---|---|---|---|---|---|
| All | blaSHV | 9.9 | 41.0 | −31.2 | 2.79 × 10−53 * | (−35.2, −27.2) |
| blaTEM | 28.6 | 74.8 | −46.3 | 1.34 × 10−91 * | (−50.7, −41.8) | |
| blaCTX-M | 65.1 | 14.2 | 50.9 | 2.40 × 10−117 * | (46.6, 55.2) | |
| blaOXA | 44.2 | 13.3 | 30.8 | 6.58 × 10−52 * | (26.9, 34.8) | |
| E. coli | blaSHV | 2.8 | 2.5 | 0.3 | 7.44 × 10−1 | (−1.5, 2.0) |
| blaTEM | 25.9 | 89.1 | −63.2 | 1.14 × 10−115 | (−68.6, −57.8) | |
| blaCTX-M | 65.8 | 12.7 | 53.1 | 1.75 × 10−83 | (47.7, 58.5) | |
| blaOXA | 43.6 | 12.5 | 31.1 | 4.16 × 10−34 | (26.1, 36.1) | |
| K. pneumoniae | blaSHV | 75.3 | 84.0 | −8.7 | 4.86 × 10−2 | (−17.4, −0.1) |
| blaTEM | 52.9 | 58.9 | −5.9 | 3.05 × 10−1 | (−17.3, 5.4) | |
| blaCTX-M | 58.8 | 16.0 | 42.9 | 1.46 × 10−18 | (33.3, 52.4) | |
| blaOXA | 49.4 | 14.2 | 35.2 | 2.65 × 10−14 | (26.2, 44.3) |
| Gene | Primer Sequence (5′ to 3′) | Product Size (bp) |
|---|---|---|
| blaSHV | Forward GCCTGTGTATTATCTCCCTGTTAG Reverse TCCCGGCGATTTGCTGATTCC | 813 |
| blaTEM | Forward TGACGCCGGGCAAGAGCA Reverse AAGGGCCGAGCGCAGAAGTG | 424 |
| blaOXA | Forward AGCGCCAGTGCATCAACAG Reverse GCAAAACCCAAACAACAGAAA | 300 |
| blaCTX-M | Forward CGGCCGCGGTGCTGAAGAA Reverse GCTGCCGGTTTTATCCCCCACAA | 482 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guzman-Cole, C.; Santiago, F.; Garsevanyan, S.; Sindi, S.; Barlow, M. Distribution of β-Lactamase Genes in Clinical Isolates from California Central Valley Hospital Deviates from the United States Nationwide Trends. Antibiotics 2021, 10, 498. https://doi.org/10.3390/antibiotics10050498
Guzman-Cole C, Santiago F, Garsevanyan S, Sindi S, Barlow M. Distribution of β-Lactamase Genes in Clinical Isolates from California Central Valley Hospital Deviates from the United States Nationwide Trends. Antibiotics. 2021; 10(5):498. https://doi.org/10.3390/antibiotics10050498
Chicago/Turabian StyleGuzman-Cole, Candace, Fabian Santiago, Sona Garsevanyan, Suzanne Sindi, and Miriam Barlow. 2021. "Distribution of β-Lactamase Genes in Clinical Isolates from California Central Valley Hospital Deviates from the United States Nationwide Trends" Antibiotics 10, no. 5: 498. https://doi.org/10.3390/antibiotics10050498
APA StyleGuzman-Cole, C., Santiago, F., Garsevanyan, S., Sindi, S., & Barlow, M. (2021). Distribution of β-Lactamase Genes in Clinical Isolates from California Central Valley Hospital Deviates from the United States Nationwide Trends. Antibiotics, 10(5), 498. https://doi.org/10.3390/antibiotics10050498