
Virulence Determinants and Antimicrobial Profiles of Pasteurella multocida Isolated from Cattle and Humans in Egypt

Abstract
1. Introduction
2. Results
2.1. Origin, Types of Collected Samples, and P. multocida Isolation
2.2. Results of Isolation of P. multocida from Different Cases
2.3. Molecular Confirmation of Isolates and Detection of Capsular Types and Virulence Genes
2.4. The Obtained Genotypic Profiles Based on Capsular Types and Virulence Genes
2.5. Phenotypic Antimicrobial Resistance and Multiple Antimicrobial Resistance Index
2.6. The Existence of Macrolide–Lincosamide–Streptogramin B (MLSB) Methylases, Monomethyltransferase, Macrolide Efflux Pump, and Macrolide-Inactivating Phosphotransferase Genes among All the Gained Isolates
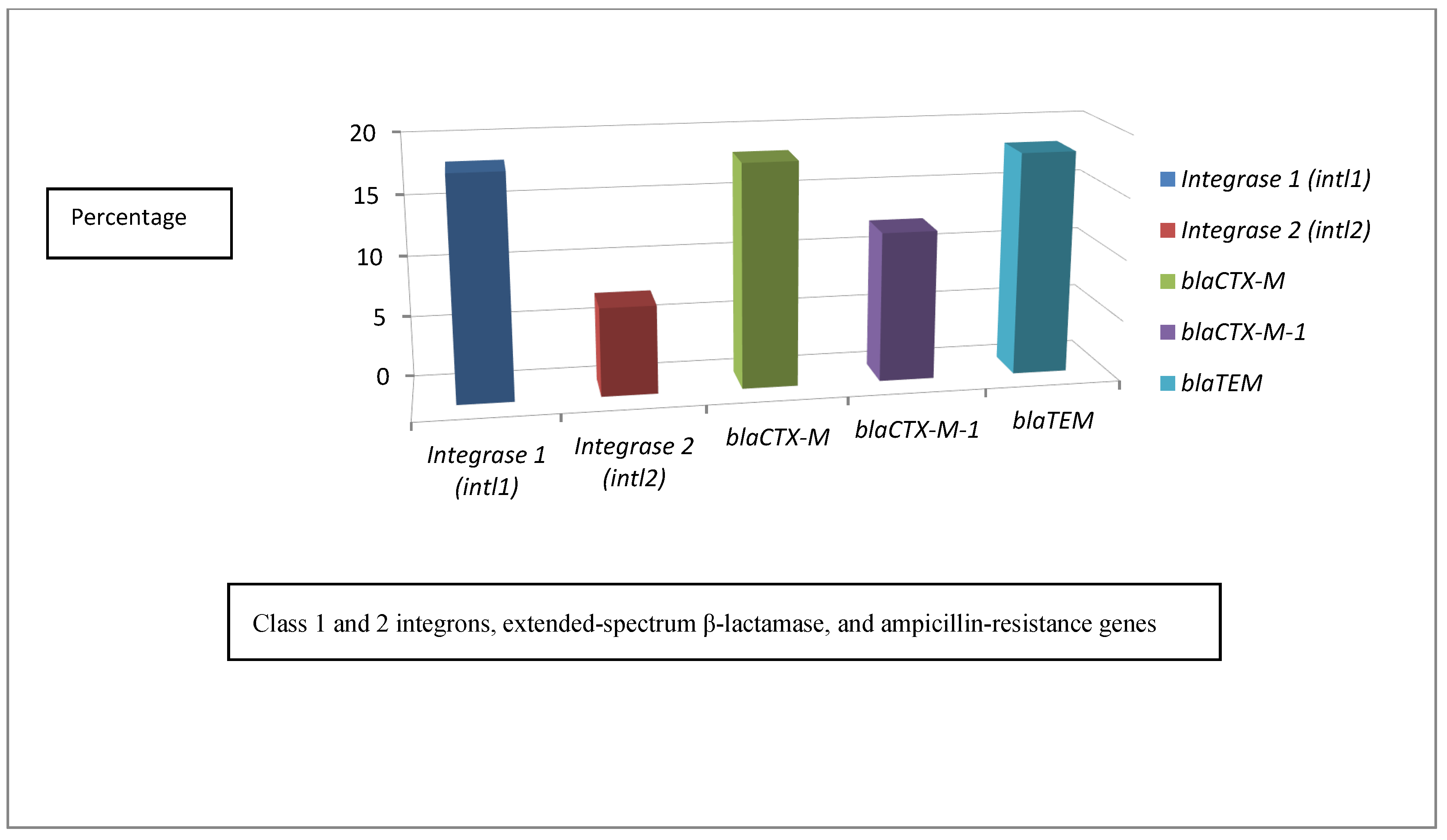
2.7. Distribution Patterns of Class 1 and 2 Integrons, Extended-Spectrum β-Lactamase, and Ampicillin-Resistance Genes
2.8. Associations between Isolation Source, Strain, and Phenotypic and Genotypic Characteristics
3. Discussion
4. Materials and Methods
4.1. Sampling, Isolation, and Identification
4.2. Phenotypic Antimicrobial Susceptibility Profiles of P. multocida Isolates
4.3. Molecular Techniques for Capsular Classification, Detection of Virulence Genes, and Antimicrobial Resistance Genes
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Zhao, G.; He, H.; Wang, H. Use of a recombinase polymerase amplification commercial kit for rapid visual detection of Pasteurella multocida. BMC Vet. Res. 2019, 15, 154. [Google Scholar] [CrossRef] [PubMed]
- Stanford, K.; Zaheer, R.; Klima, C.; McAllister, T.; Peters, D.; Niu, Y.D.; Ralston, B. Antimicrobial resistance in members of the bacterial bovine respiratory disease complex isolated from lung tissue of cattle mortalities managed with or without the use of antimicrobials. Microorganisms 2020, 8, 288. [Google Scholar] [CrossRef] [PubMed]
- Arashima, Y.; Kubo, N.; Iwasaki, Y.; Okuyama, K.; Kumasaka, K.; Tsuchiya, T.; Kawano, K.; Ootsuka, M.; Saitoh, F.; Namikawa, K. Human respiratory tract infection by Pasteurella multocida subsp. multocida presumably derived from the cat. Kansenshogaku Zasshi. 1990, 64, 1200–1204. [Google Scholar] [CrossRef][Green Version]
- Guo, Y.; McMullen, C.; Timsit, E.; Hallewell, J.; Orsel, K.; van der Meer, F.; Yan, S.; Alexander, T.W. Genetic relatedness and antimicrobial resistance in respiratory bacteria from beef calves sampled from spring processing to 40 days after feedlot entry. Vet. Microbiol. 2020, 240, 108478. [Google Scholar] [CrossRef]
- Annas, S.; Zamri-Saad, M.; Jesse, F.F.; Zunita, Z. New sites of localisation of Pasteurella multocida B:2 in buffalo surviving experimental haemorrhagic septicaemia. BMC Vet. Res. 2014, 10, 88. [Google Scholar] [CrossRef][Green Version]
- Al-Maary, K.S.; Dawoud, T.M.; Mubarak, A.S.; Hessain, A.M.; Galal, H.M.; Kabli, S.A.; Mohamed, M.I. Molecular characterization of the capsular antigens of Pasteurella multocida isolates using multiplex PCR. Saudi J. Biol. Sci. 2017, 24, 367–370. [Google Scholar] [CrossRef]
- St Michael, F.; Harper, M.; Parnas, H.; John, M.; Stupak, J.; Vinogradov, E.; Adler, B.; Boyce, J.D.; Cox, A.D. Structural and genetic basis for the serological differentiation of Pasteurella multocida Heddleston serotypes 2 and 5. J. Bacteriol. 2009, 191, 6950–6959. [Google Scholar] [CrossRef]
- Carter, G.R. A new serological type of Pasteurella multocida from central Africa. Vet. Rec. 1961, 73, 1052. [Google Scholar]
- Rimler, R.B.; Rhoades, K.R. Serogroup F. A new capsule serogroup of Pasteurella multocida. J. Clin. Microbiol. 1987, 25, 615–618. [Google Scholar] [CrossRef]
- Peng, Z.; Liang, W.; Wu, B. Molecular typing methods for Pasteurella multocida—A review. Acta Microbiol. Sin. 2016, 56, 1521–1529. [Google Scholar] [CrossRef]
- Townsend, K.M.; Boyce, J.D.; Chung, J.Y.; Frost, A.J.; Adler, B. Genetic organization of Pasteurella multocida cap Loci and development of a multiplex capsular PCR typing system. J. Clin. Microbiol. 2001, 39, 924–929. [Google Scholar] [CrossRef]
- Harper, M.; John, M.; Turni, C.; Edmunds, M.; St Michael, F.; Adler, B.; Blackall, P.J.; Cox, A.D.; Boyce, J.D. Development of a rapid multiplex PCR assay to genotype Pasteurella multocida strains by use of the lipopolysaccharide outer core biosynthesis locus. J. Clin. Microbiol. 2015, 53, 477–485. [Google Scholar] [CrossRef]
- Peng, Z.; Liang, W.; Wang, F.; Xu, Z.; Xie, Z.; Lian, Z.; Hua, L.; Zhou, R.; Chen, H.; Wu, B. Genetic and phylogenetic characteristics of Pasteurella multocida isolates from different host species. Front. Microbiol. 2018, 9, 1408. [Google Scholar] [CrossRef] [PubMed]
- Portis, E.; Lindeman, C.; Johansen, L.; Stoltman, G. A ten-year (2000–2009) study of antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex—Mannheimia haemolytica, Pasteurella multocida, and Histophilus somni—In the United States and Canada. J. Vet. Diagn. Investig. 2012, 24, 932–944. [Google Scholar] [CrossRef]
- DeDonder, K.D.; Apley, M.D. A literature review of antimicrobial resistance in pathogens associated with bovine respiratory disease. Anim. Health Res. Rev. 2015, 16, 125–134. [Google Scholar] [CrossRef] [PubMed]
- Anholt, R.M.; Klima, C.; Allan, N.; Matheson-Bird, H.; Schatz, C.; Ajilkumar, P.; Otto, S.J.G.; Peters, D.; Schmid, K.; Olson, M.; et al. Antimicrobial susceptibility of bacteria that cause bovine respiratory disease complex in Alberta Canada. Front. Vet. Sci. 2017, 4, 207. [Google Scholar] [CrossRef] [PubMed]
- Klima, C.L.; Zaheer, R.; Cook, S.R.; Booker, C.W.; Hendrick, S.; Alexander, T.W.; McAllister, T.A. Pathogens of bovine respiratory disease in north american feedlots conferring multidrug resistance via integrative conjugative elements. J. Clin. Microbiol. 2014, 52, 438–448. [Google Scholar] [CrossRef]
- Feng, Y.; Qi, W.; Wang, X.; Wang, L.; Li, X.; Luo, J.; Zhang, S.; Li, H. Genetic characterization of antimicrobial resistance in Staphylococcus aureus isolated from bovine mastitis cases in northwest China. J. Integr. Agric. 2016, 15, 60345–60347. [Google Scholar] [CrossRef]
- EFSA. Scientific Opinion on the public health risks of bacterial strains producing extendedspectrum β-lactamases and/or AmpC β-lactamases in food and food-producing animals. EFSA J. 2011, 9, 2322. [Google Scholar] [CrossRef]
- Michael, G.B.; Freitag, C.; Wendlandt, S.; Eidam, C.; Feßler, A.T.; Lopes, G.V.; Kadlec, K.; Schwarz, S. Emerging issues in antimicrobial resistance of bacteria from food-producing animals. Future Microbiol. 2015, 10, 427–443. [Google Scholar] [CrossRef]
- Bush, K.; Jacoby, G.A. Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 2010, 54, 969–976. [Google Scholar] [CrossRef]
- Ur Rahman, S.; Ali, T.; Ali, I.; Khan, N.A.; Han, B.; Gao, J. The growing genetic and functional diversity of extended spectrum beta-lactamases. BioMed Res. Int. 2018, 2018, 9519718. [Google Scholar] [CrossRef]
- Steen, J.A.; Steen, J.A.; Harrison, P.; Seemann, T.; Wilkie, I.; Harper, M.; Adler, B.; Boyce, J.D. Fis is essential for capsule production in Pasteurella multocida and regulates expression of other important virulence factors. PLOS Pathog. 2010, 6, e1000750. [Google Scholar] [CrossRef]
- Khamesipour, F.; Momtaz, H.; Azhdary Mamoreh, M. Occurrence of virulence factors and antimicrobial resistance in Pasteurella multocida strains isolated from slaughter cattle in Iran. Front. Microbiol. 2014, 5, 536. [Google Scholar] [CrossRef] [PubMed]
- Glickman, M.; Klein, R.S. Acute Epiglottitis due to Pasteurella multocida in an Adult without Animal Exposure. Emerg. Infect. Dis. 1997, 3, 408–409. [Google Scholar] [CrossRef] [PubMed]
- Moyko, A.; Ali, N.J.; Dubosh, N.M.; Wong, M.L. Pasteurella multocida Epiglottitis. Clin. Pract. Cases Emerg. Med. 2017, 1, 22–24. [Google Scholar] [CrossRef] [PubMed]
- Pandit, K.K.; Smith, J.E. Capsular hyaluronic acid in Pasteurella multocida type A and its counterpart in type D. Res. Vet. Sci. 1993, 54, 20–24. [Google Scholar] [CrossRef]
- Rimler, R.B.; Rhoades, K.R. Hyaluronidase and chondroitinase activity of Pasteurella multocida serotype B:2 involved in hemorrhagic septicaemia. Vet. Rec. 1994, 134, 67–68. [Google Scholar] [CrossRef]
- DeAngelis, P.L. Enzymological characterization of the Pasteurella multocida hyaluronic acid synthase. Biochemistry 1996, 35, 9768–9771. [Google Scholar] [CrossRef] [PubMed]
- DeAngelis, P.L.; Padgett-McCue, A.J. Identification and molecular cloning of a chondroitin synthase from Pasteurella multocida type F. J. Biol. Chem. 2000, 275, 24124–24129. [Google Scholar] [CrossRef]
- Knox, K.W.; Bain, R.V.S. The antigens of Pasteurella multocida type I. Immunology 1960, 3, 352–362. [Google Scholar]
- Boyce, J.D.; Chung, J.Y.; Adler, B. Pasteurella multocida capsule: Composition, function and genetics. J. Biotechnol. 2000, 83, 153–160. [Google Scholar] [CrossRef]
- Ataei Kachooei, S.; Ranjbar, M.M.; Ataei Kachooei, S. Evaluation of Pasteurella multocida serotype B:2 resistance to immune serum and complement system. Vet. Res. Forum 2017, 8, 179–184. [Google Scholar]
- Katoch, S.; Sharma, M.; Patil, R.D.; Kumar, S.; Verma, S. In vitro and in vivo pathogenicity studies of Pasteurella multocida strains harbouring different ompA. Vet. Res. Commun. 2014, 38, 183–191. [Google Scholar] [CrossRef] [PubMed]
- Cameron, A.; McAllister, T.A. Antimicrobial usage and resistance in beef production. J. Anim. Sci. Biotechnol. 2016, 7, 68. [Google Scholar] [CrossRef] [PubMed]
- El-Seedy, F.R.; Abed, A.H.; Hassan, H.M.; Nabih, A.M.; Khalifa, E.; Salem, S.E. Antimicrobial and immunological studies on Pasteurella multocida and Mannheimia haemolytica recovered from calves affected with respiratory manifestations. JVMR 2019, 26, 55–63. [Google Scholar]
- Koike, S.; Inoue, K.; Yoneyama, S.; Ichikawa, Y.; Tajima, K. Antimicrobial susceptibility of respiratory bacterial pathogen isolated from cattle for the past 16 years in Tochigi Prefecture. JVMA 2009, 62, 533–537. [Google Scholar] [CrossRef][Green Version]
- Anwar, M.A.H.; Rahman, S.U.; Ahmad, R. Antibiotic Sensitivity of Pasteurella multocida isolated from cattle and buffaloes. Pak. J. Biol. Sci. 2000, 3, 692–693. [Google Scholar] [CrossRef]
- Elshemey, T.M.; Abd-Elrahman, A.H. Hemorrhagic septicemia outbreak as a consequence to SAT2 FMD infection in buffalo and cattle in Alexandria province, Egypt. Life Sci. J. 2013, 10, 816–822. [Google Scholar]
- Ujvári, B.; Makrai, L.; Magyar, T. Characterisation of a multiresistant Pasteurella multocida strain isolated from cattle. Acta Vet. Hung. 2018, 66, 12–19. [Google Scholar] [CrossRef]
- Michael, G.B.; Kadlec, K.; Sweeney, M.T.; Brzuszkiewicz, E.; Liesegang, H.; Daniel, R.; Murray, R.W.; Watts, J.L.; Schwarz, S. ICEPmu1, an integrative conjugative element (ICE) of Pasteurella multocida: Analysis of the regions that comprise 12 antimicrobial resistance genes. J. Antimicrob. Chemother. 2012, 67, 84–90. [Google Scholar] [CrossRef]
- San Millan, A.; Escudero, J.A.; Gutierrez, B.; Hidalgo, L.; Garcia, N.; Llagostera, M.; Dominguez, L.; Gonzalez-Zorn, B. Multiresistance in Pasteurella multocida is mediated by coexistence of small plasmids. Antimicrob. Agents Chemother. 2009, 53, 3399–3404. [Google Scholar] [CrossRef] [PubMed]
- Moustafa, A.M.; Seemann, T.; Gladman, S.; Adler, B.; Harper, M.; Boyce, J.D.; Bennett, M.D. Comparative genomic analysis of Asian haemorrhagic septicaemia-associated strains of Pasteurella multocida identifies more than 90 haemorrhagic septicaemia-specific genes. PLoS ONE 2015, 10, e0130296. [Google Scholar] [CrossRef]
- Rose, S.; Desmolaize, B.; Jaju, P.; Wilhelm, C.; Warrass, R.; Douthwaite, S. Multiplex PCR to identify macrolide resistance determinants in Mannheimia haemolytica and Pasteurella multocida. Antimicrob. Agents Chemother. 2012, 56, 3664–3669. [Google Scholar] [CrossRef] [PubMed]
- Desmolaize, B.; Rose, S.; Warrass, R.; Douthwaite, S. A novel Erm monomethyltransferase in antibiotic-resistant isolates of Mannheimia haemolytica and Pasteurella multocida. Mol. Microbiol. 2011, 80, 184–194. [Google Scholar] [CrossRef]
- Kadlec, K.; Brenner Michael, G.; Sweeney, M.T.; Brzuszkiewicz, E.; Liesegang, H.; Daniel, R.; Watts, J.L.; Schwarz, S. Molecular basis of macrolide, triamilide, and lincosamide resistance in Pasteurella multocida from bovine respiratory disease. Antimicrob. Agents Chemother. 2011, 55, 2475–2477. [Google Scholar] [CrossRef] [PubMed]
- Desmolaize, B.; Rose, S.; Wilhelm, C.; Warrass, R.; Douthwaite, S. Combinations of macrolide resistance determinants in field isolates of Mannheimia haemolytica and Pasteurella multocida. Antimicrob. Agents Chemother. 2011, 55, 4128–4133. [Google Scholar] [CrossRef]
- Klima, C.L.; Cook, S.R.; Zaheer, R.; Laing, C.; Gannon, V.P.; Xu, Y.; Rasmussen, J.; Potter, A.; Hendrick, S.; Alexander, T.W.; et al. Comparative genomic analysis of Mannheimia haemolytica from bovine sources. PLoS ONE 2016, 11, e0149520. [Google Scholar] [CrossRef] [PubMed]
- Klima, C.L.; Holman, D.B.; Cook, S.R.; Conrad, C.C.; Ralston, B.J.; Allan, N.; Anholt, R.M.; Niu, Y.D.; Stanford, K.; Hannon, S.J.; et al. Multidrug resistance in Pasteurellaceae associated with bovine respiratory disease mortalities in north America from 2011 to 2016. Front. Microbiol. 2020, 11, 606438. [Google Scholar] [CrossRef]
- Petrocchi-Rilo, M.; Gutiérrez-Martín, C.B.; Pérez-Fernández, E.; Vilaró, A.; Fraile, L.; Martínez-Martínez, S. Antimicrobial resistance genes in porcine Pasteurella multocida are not associated with its antimicrobial susceptibility pattern. Antibiotics 2020, 9, 614. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Bao, X.; Ji, L.; Chen, L.; Liu, J.; Miao, J.; Chen, D.; Bian, H.; Li, Y.; Yu, G. Resistance integrons: Class 1, 2 and 3 integrons. Ann. Clin. Microbiol. Antimicrob. 2015, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.C.; Wang, Z.; Wang, Y.M.; Dong, W.L.; Jia, B.Y.; Gao, D.; Jiang, X.Y.; Ma, H.X. Antimicrobial susceptibility and molecular typing of Pasteurella multocida isolated from six provinces in China. Trop. Anim. Health Prod. 2019, 51, 987–992. [Google Scholar] [CrossRef]
- Kehrenberg, C.; Schwarz, S. Trimethoprim resistance in a porcine Pasteurella aerogenes isolate is based on a dfrA1 gene cassette located in a partially truncated class 2 integron. J. Antimicrob. Chemother. 2011, 66, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Michael, G.B.; Bossé, J.T.; Schwarz, S. Antimicrobial resistance in Pasteurellaceae of veterinary origin. Microbiol. Spectr. 2018, 6, 1–33. [Google Scholar] [CrossRef]
- Zhao, W.H.; Hu, Z.Q. Epidemiology and genetics of CTX-M extended-spectrum β-lactamases in Gram-negative bacteria. Crit. Rev. Microbiol. 2013, 39, 79–101. [Google Scholar] [CrossRef] [PubMed]
- Awad, N.F.S.; Abd El-Hamid, M.I. Coexistence of virulence and antibiotic resistance genes in Pasteurella multocida isolated from diseased rabbits. Zagazig Vet. J. 2019, 47, 91–102. [Google Scholar] [CrossRef]
- Elalamy, R.A.; Tartor, Y.H.; Ammar, A.M.; Eldesouky, I.E.; Esawy, A.E.I. Molecular characterization of extensively drug-resistant Pasteurella multocida isolated from apparently healthy and diseased chickens in Egypt. Pak. Vet. J. 2020, 40, 319–324. [Google Scholar] [CrossRef]
- Naas, T.; Benaoudia, F.; Lebrun, L.; Nordmann, P. Molecular identification of TEM-1 β-lactamase in a Pasteurella multocida isolate of human origin. Eur. J. Clin. Microbiol. Infect. Dis. 2001, 20, 210–213. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Singh, V.P.; Agrawal, R.K.; Singh, S. Identification of Pasteurella multocida isolates of ruminant origin using polymerase chain reaction and their antibiogram study. Trop. Anim. Health Prod. 2009, 41, 573–578. [Google Scholar] [CrossRef] [PubMed]
- Moore, M.K.; Cicnjak-Chubbs, L.; Gates, R.J. A new selective enrichment procedure for isolating Pasteurella multocida from avian and environmental samples. Avian Dis. 1994, 38, 317–324. [Google Scholar] [CrossRef]
- Kozarev, A.; Mamadudian, B.A. Biochemical properties of Pasteurella multocida strains from ruminants. Vet. Sibrika 1988, 86, 28–30. [Google Scholar]
- Ashraf, A.; Tariq, H.; Shah, S.; Nadeem, S.; Manzoor, I.; Ali, S.; Ijaz, A.; Gailani, S.; Mehboob, S. Characterization of Pasteurella multocida strains isolated from cattle and buffaloes in Karachi, Pakistan. Afr. J. Microbiol. Res. 2011, 5, 4673–4677. [Google Scholar] [CrossRef]
- Sellyei, B.; Wehmann, E.; Makrai, L.; Magyar, T. Evaluation of the Biolog system for the identification of certain closely related Pasteurella species. Diagn. Microbiol. Infect. Dis. 2011, 71, 6–11. [Google Scholar] [CrossRef] [PubMed]
- WHO (World Health Organization). Critically Important Antimicrobials for Human Medicine—5th Revision; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fourth Informational Supplement CLSI Document M100-S24; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2014. [Google Scholar]
- Furian, T.Q.; Borges, K.A.; Rocha, S.L.S.; Rodrigues, E.E.; Nascimento, V.P.; Salle, C.T.P.; Moraes, H.L.S. Detetion of virulence-associated genes of Pasteurella multocida isolated from cases of fowl cholera by multiplex-PCR. Pesq. Vet. Bras. 2013, 33, 177–182. [Google Scholar] [CrossRef][Green Version]
- Elsayed, M.S.A.E.; Roshdey, T.; Salah, A.; Tarabees, R.; Younis, G.; Eldeep, D. Phenotypic and genotype methods for identification of slime layer production, efflux pump activity, and antimicrobial resistance genes as potential causes of the antimicrobial resistance of some mastitis pathogens from farms in Menoufia, Egypt. Mol. Biol. Rep. 2019, 46, 6533–6546. [Google Scholar] [CrossRef] [PubMed]
- Raphael, E.; Wong, L.K.; Riley, L.W. Extended-spectrum beta-lactamase gene sequences in Gram-negative saprophytes on retail organic and nonorganic spinach. Appl. Environ. Microbiol. 2011, 77, 1601–1607. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin of Samples | Types of Cases | ||||
|---|---|---|---|---|---|
| Apparently Healthy | Diseased | ||||
| No. | % | No. | % | ||
| Minufiya | Cattle (85) | 40 | 40/100 (40) | 45 | 45/100 (45) |
| Human (7) | 2 | 2/15 (13.3) | 5 | 5/15 (33.3) | |
| Qalyubia | Cattle (15) | 8 | 8/100 (8) | 7 | 7/100 (7) |
| Human (8) | 3 | 3/15 (20) | 5 | 5/15 (33.3) | |
| Total collected samples | Cattle (100) | 48 | 48/100 (48) | 52 | 52/100 (52) |
| Human (15) | 5 | 5/15 (33.3) | 10 | 10/15 (66.7) | |
| Positive isolation | Cattle (50) | 6 (5 from Minufiya and 1 from Qalyubia) | 6/48 (12.5) | 44 (40 from Minufiya and 4 from Qalyubia) | 44/52 (84.6) |
| Human (5) | 0 | 0/15 (0.0) | 5 from Qalyubia | 5/15 (33.3) | |
| Name | Amplicon Size (bp) | Cycle Number | Annealing Temperature (°C) and Time (Seconds) | Primer Sequence | Purpose | Reference |
|---|---|---|---|---|---|---|
| kmt1 | 460 | 30 | 55 °C, 30 s | KMT1T7-ATCCGCTATTTACCCAGTGG KMT1SP6-GCTGTAAACGAACTCGCCAC | Confirmation of isolates using primer targeting an integral component of membrane | [11] |
| hyaD-hyaC | 1044 | 30 | 55 °C, 30 s | CAPA-TGCCAAAATCGCAGTCAG CAPA-TTGCCATCATTGTCAGTG | Molecular capsular typing | |
| bcbD | 760 | 30 | 55 °C, 30 s | CAPB-CATTTATCCAAGCTCCACC CAPB-GCCCGAGAGTTTCAATCC | ||
| dcbF | 657 | 30 | 55 °C, 30 s | CAPD-TTACAAAAGAAAGACTAGGAGCCC CAPD-CATCTACCCACTCAACCATATCAG | ||
| ecbJ | 511 | 30 | 55 °C, 30 s | CAPE-TCCGCAGAAAATTATTGACTC CAPE-GCTTGCTGCTTGATTTTGTC | ||
| fcbD | 851 | 30 | 55 °C, 30 s | CAPF-AATCGGAGAACGCAGAAATCAG CAPF-TTCCGCCGTCAATTACTCTG | ||
| sodA | 361 | 25 | 55 °C, 30 s | F:TACCAGAATTAGGCTACGC R:GAAACGGGTTGCTGCCGCT | Superoxide dismutase | [66] |
| hgbA | 419 | 25 | 55 °C, 30 s | F:TGGCGGATAGTCATCAAG R:CCAAAGAACCACTACCCA | Iron uptake | |
| hgbB | 788 | 25 | 55 °C, 30 s | F:ACCGCGTTGGAATTATGATTG R:CATTGAGTACGGCTTGACAT | Iron uptake | |
| ptfA | 488 | 25 | 55 °C, 30 s | F:TGTGGAATTCAGCATTTTAGTGTGTC R:TCATGAATTCTTATGCGCAAAATCCTGCTGG | Type IV fimbriae | |
| pfhA | 275 | 25 | 55 °C, 30 s | F:AGCTGATCAAGTGGTGAAC RTGGTACATTGGTGAATGCTG | Hemagglutinin | |
| erm(A) | 157 | 40 | 63 °C, 30 s | F:AGTCAGGCTAAATATAGCTATC R:CAAGAACAATCAATACAGAGTCTAC | Macrolide–lincosamide–streptogramin B methylases | [67] |
| erm(B) | 191 | 40 | 65 °C, 30 s | F:GGTTGCTCTTGCACACTCAAG R:CAGTTGACGATATTCTCGATTG | ||
| erm(C) | 293 | 40 | 63 °C, 30 s | F:AATCGTGGAATACGGGTTTGC R:CGTCAATTCCTG CATGTTTTAAGG | ||
| erm(F) | 424 | 40 | 65 °C, 30 s | F:TCTGGGAGGTTCCATTGTCC R:TTCAGGGACAACTTCCAG C | ||
| erm(G) | 255 | 40 | 63 °C, 30 s | F:GTGAGGTAACTCGTAATAAGCTG R:CCTCTGCCATTAACAGCAATG | ||
| erm(Q) | 154 | 35 | 68 °C, 30 s | F:CACCAACTGATATGTGGCTAG R:CTAGGCATGGGATGGAAGTC | ||
| erm(42) | 173 | 25 | 68 °C, 30 s | F:TGCACCATCTTACAAGGAGT R:CATGCCTGTCTTCAAGGTTT | MLSB monomethyltransferase | [43] |
| mph(E) | 271 | 25 | 68 °C, 30 s | F:ATGCCCAGCATATAAATCGC R:ATATGGACAAAGATAGCCCG | Macrolide efflux pump | |
| msr(E) | 395 | 25 | 68 °C, 30 s | F:TATAGCGACTTTAGCGCCAA R:GCCGTAGAATATGAGCTGAT | Macrolide-inactivating phosphotransferase | |
| intl1 | 280 | 33 | 64 °C, 30 s | F:CCTCCCGCACGATGATC R:TCCACGCATCGTCAGGC | Detect class 1 and 2 integrons | [68] |
| intl2 | 300 | 33 | 64 °C, 30 s | F:GCAAACGCAAGCATTCATTA R:ACGGATATGCGACAAAAAGG | ||
| blaCTX-M | 500 | 35 | 55 °C, 1 min | F:TTTGCGATGTGCAGTACCAGTAA R:CTCCGCTGCCGGTTTTATC | Detect extended-spectrum β-lactamase | |
| blaCTX-M-1 | 415 | 35 | 55 °C, 1 min | F:AAAAATCACTGCGCCAGTTC R:AGCTTATTCATCGCCACGTT | ||
| blaTEM | 800 | 30 | 55 °C, 1 min | F:CCGTGTCGCCCTTATTCC R:AGGCACCTATCTCAGCGA | Ampicillin-resistance gene |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Elsayed, M.S.A.E.; Eldsouky, S.M.; Roshdy, T.; Said, L.; Thabet, N.; Allam, T.; Mohammed, A.B.A.; Nasr, G.M.; Basiouny, M.S.M.; Akl, B.A.; et al. Virulence Determinants and Antimicrobial Profiles of Pasteurella multocida Isolated from Cattle and Humans in Egypt. Antibiotics 2021, 10, 480. https://doi.org/10.3390/antibiotics10050480
Elsayed MSAE, Eldsouky SM, Roshdy T, Said L, Thabet N, Allam T, Mohammed ABA, Nasr GM, Basiouny MSM, Akl BA, et al. Virulence Determinants and Antimicrobial Profiles of Pasteurella multocida Isolated from Cattle and Humans in Egypt. Antibiotics. 2021; 10(5):480. https://doi.org/10.3390/antibiotics10050480
Chicago/Turabian StyleElsayed, Mohamed Sabry Abd Elraheam, Samah Mahmoud Eldsouky, Tamer Roshdy, Lamia Said, Nahed Thabet, Tamer Allam, A. B. Abeer Mohammed, Ghada M. Nasr, Mohamed S. M. Basiouny, Behairy A. Akl, and et al. 2021. "Virulence Determinants and Antimicrobial Profiles of Pasteurella multocida Isolated from Cattle and Humans in Egypt" Antibiotics 10, no. 5: 480. https://doi.org/10.3390/antibiotics10050480
APA StyleElsayed, M. S. A. E., Eldsouky, S. M., Roshdy, T., Said, L., Thabet, N., Allam, T., Mohammed, A. B. A., Nasr, G. M., Basiouny, M. S. M., Akl, B. A., Nader, M. M., Hasan, A. S., & Salah, A. (2021). Virulence Determinants and Antimicrobial Profiles of Pasteurella multocida Isolated from Cattle and Humans in Egypt. Antibiotics, 10(5), 480. https://doi.org/10.3390/antibiotics10050480