
Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity—A Pharmaco-Toxicological Screening

 ,
,  ,
,  , ,
, , 
Abstract
1. Introduction
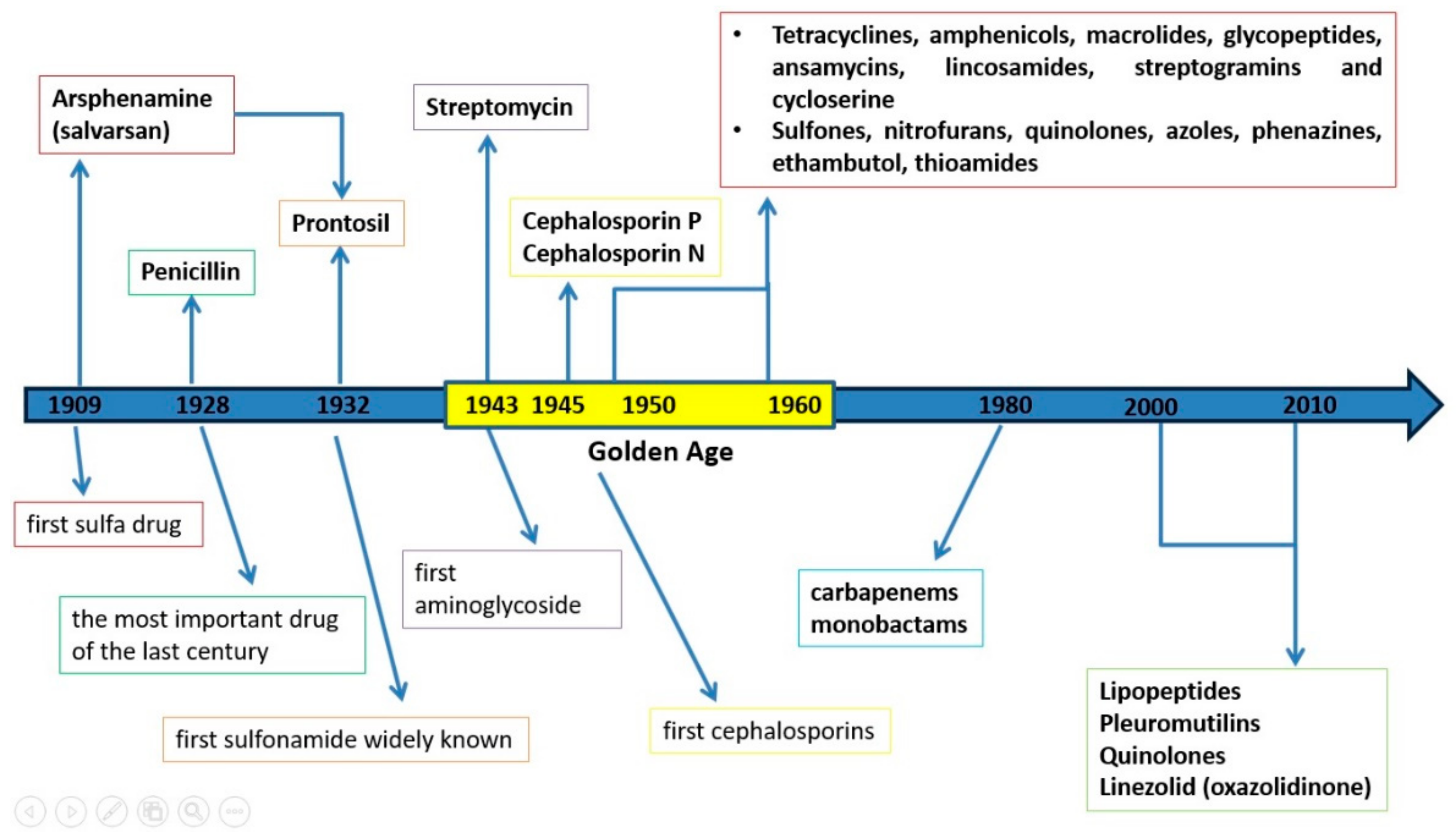
2. History of Antibiotics
3. Classification of Antibiotics
3.1. Antibiotics’ Classification by the Nature of the Source
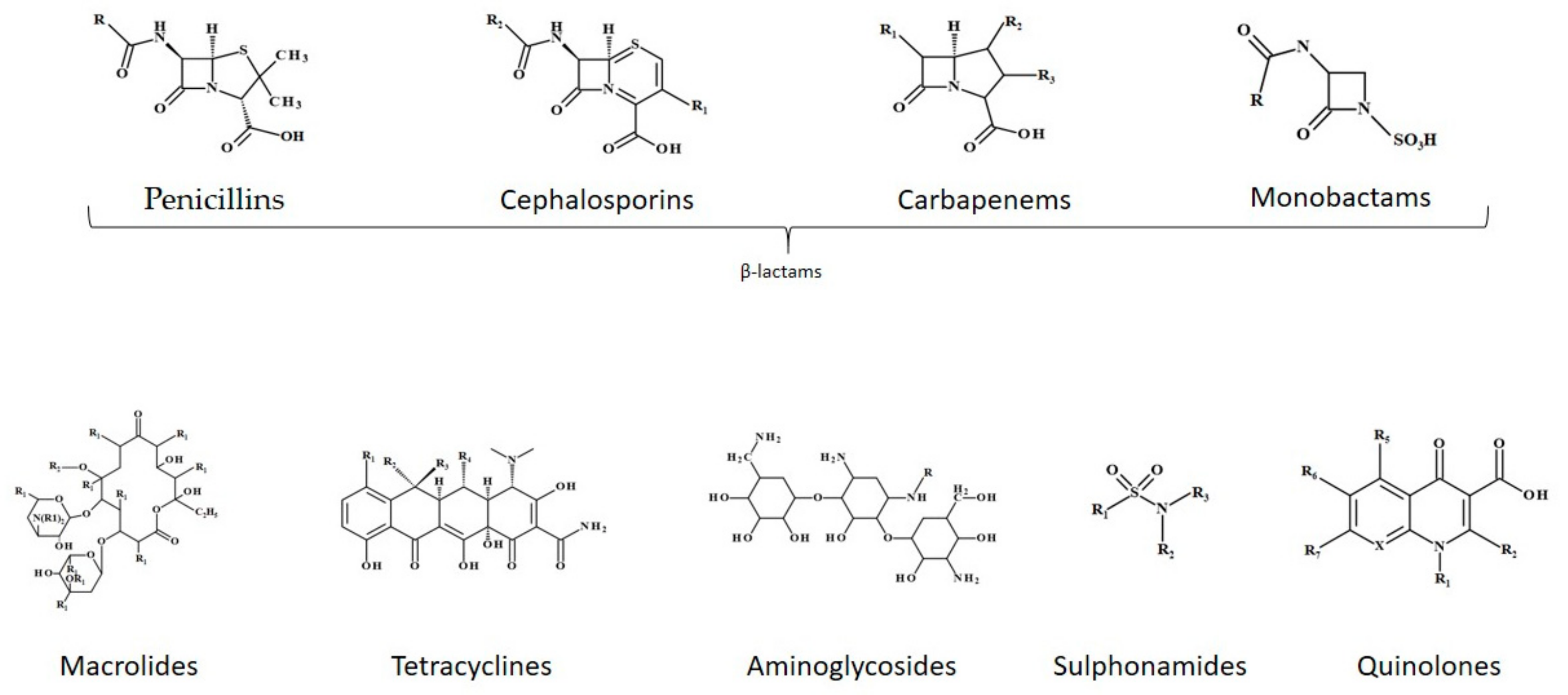
3.2. Antibiotics’ Classification Based on Chemical Structure
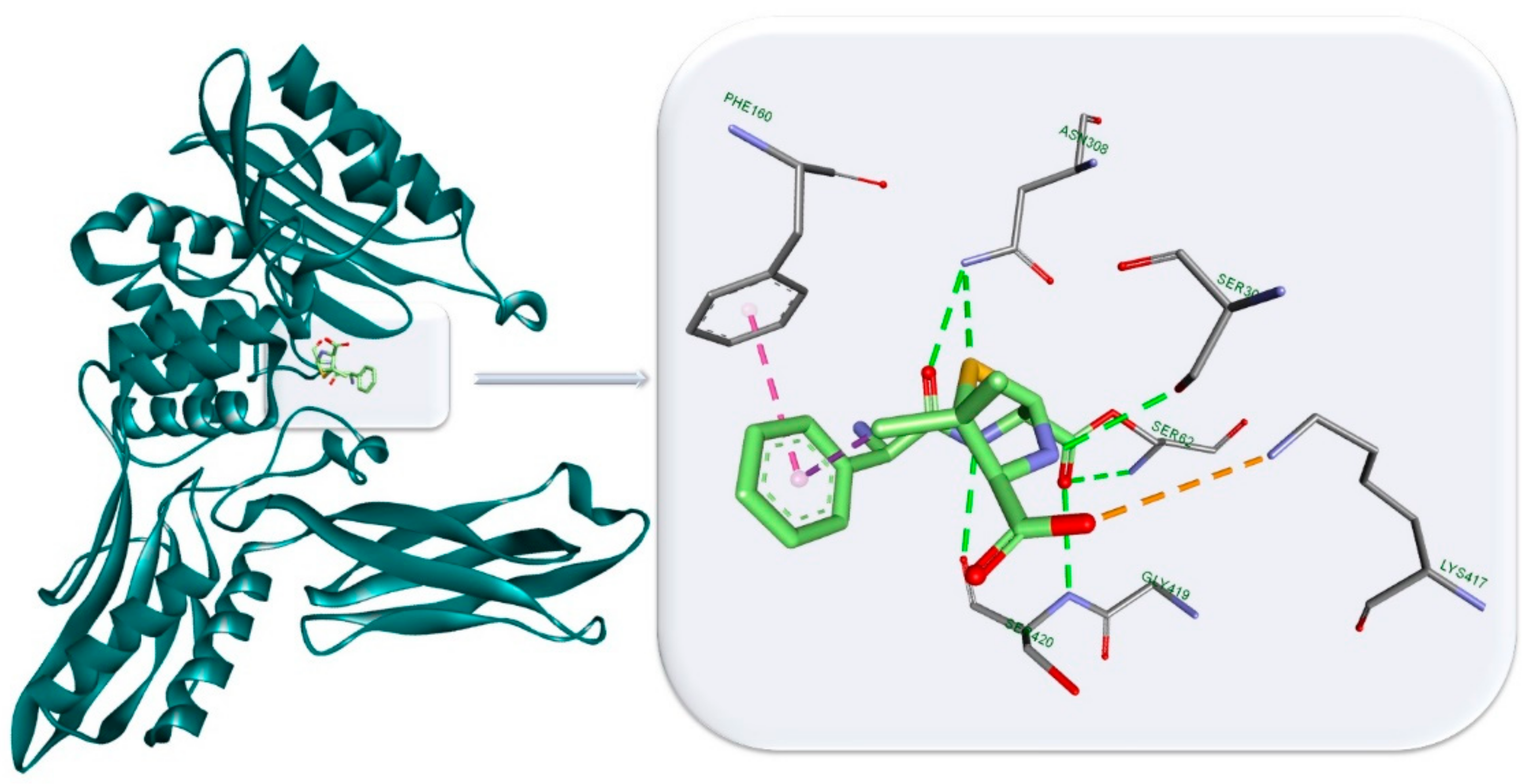
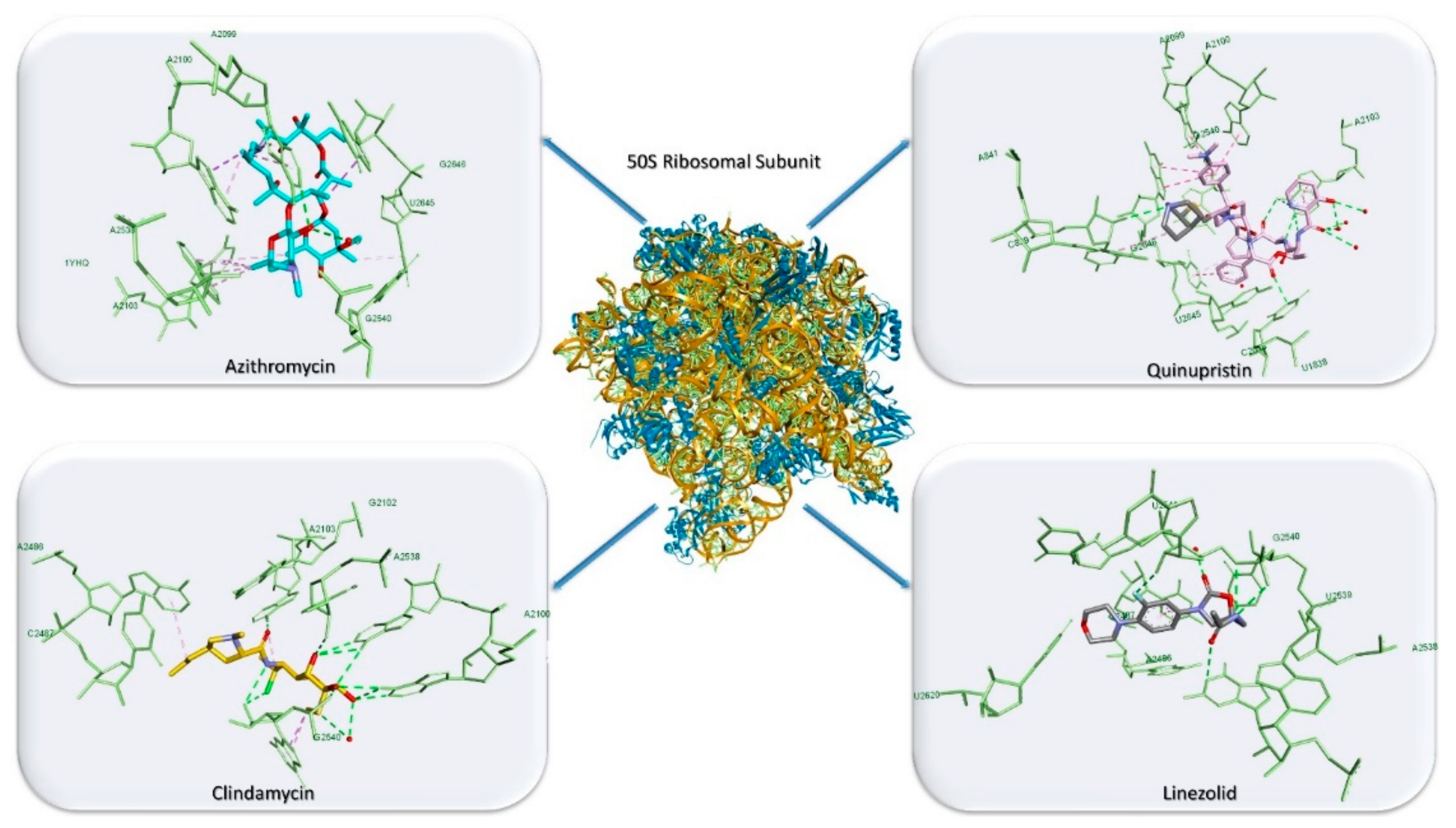
3.3. Antibiotics’ Classification by the Mechanism of Action
3.4. Antibiotics’ Classification by the Type of Pharmacological Effects
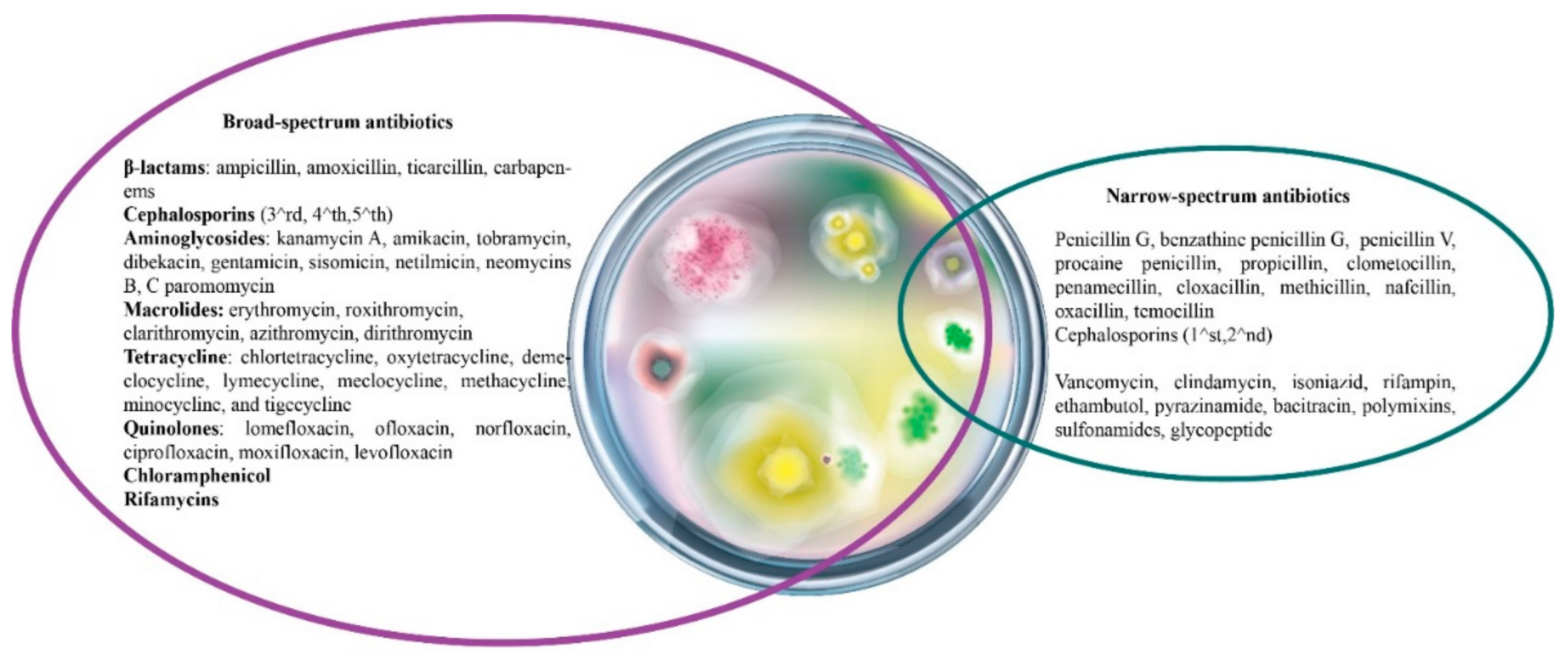
3.5. Antibiotics’ Classification Based on the Spectrum of Activity
4. Antibiotics’ Toxicity
4.1. Hypersensitivity Adverse Effects
4.2. Hematologic Adverse Effects
4.3. Neurologic Adverse Effects
4.4. Gastrointestinal Adverse Effects
4.5. Renal Adverse Effects
4.6. Cardiac Adverse Effects
4.7. Pulmonary Adverse Effects
4.8. Hepatic Adverse Effects
4.9. Accurate Selection of Relevant Criteria to Reduce the Toxicity of Antibiotics
4.10. Bacterial Resistance to Antibiotics
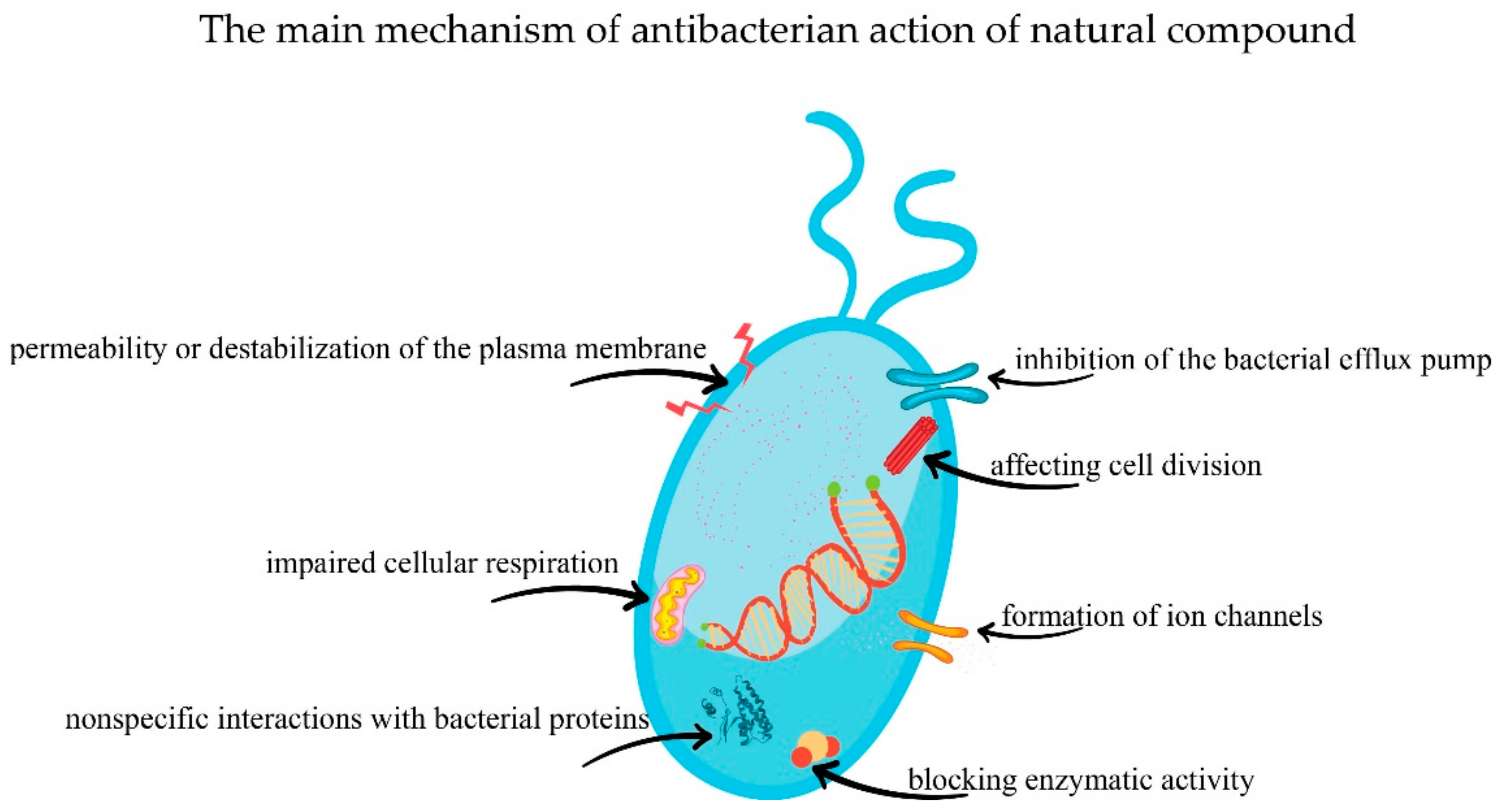
5. Natural Compounds with Antibacterial Effect
5.1. Phenols and Polyphenols
5.2. Terpenoids and Essential Oils
5.3. Alkaloids
5.4. Lectins and Polypeptide
5.5. Other Phytocompounds with Antibacterial Effect
6. Antibiotics and Microbiota
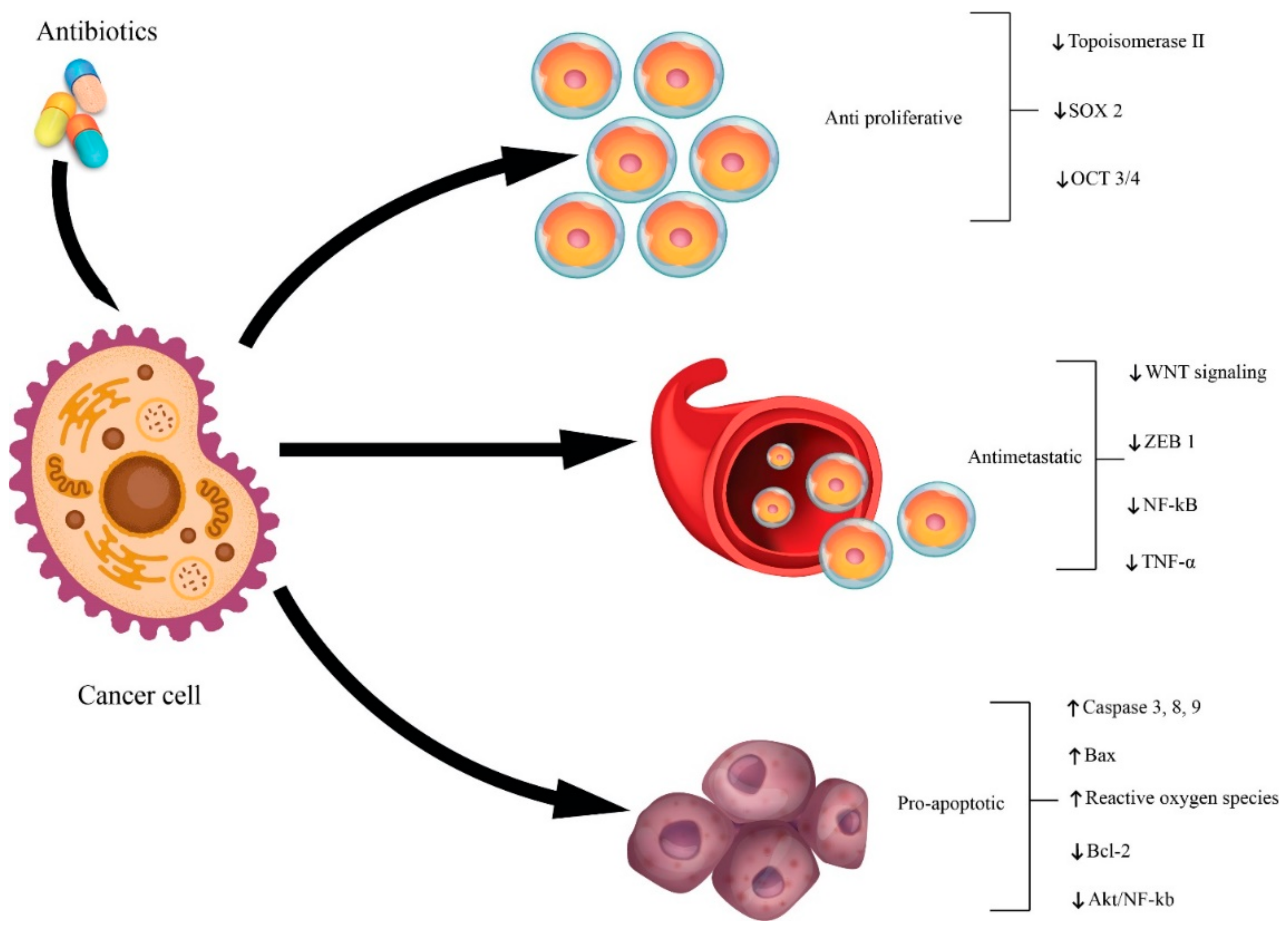
7. Antibiotics and Cancer
7.1. Antibiotics as Cancer Therapy
7.2. Antibiotics as Cancer Promoters
7.3. Potential Pro-Tumoral Mechanisms of Action Related to Antibiotics
8. Conclusions and Future Perspective
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Porter, J.R. Antony van Leeuwenhoek: Tercentenary of his discovery of bacteria. Bacteriol. Rev. 1976, 40, 260–269. [Google Scholar] [CrossRef]
- Ullah, H.; Ali, S. Classification of Anti-Bacterial Agents and Their Functions. Antibact. Agents 2017. [Google Scholar] [CrossRef]
- Mohr, K.I. History of Antibiotics Research. Curr. Top. Microbiol. Immunol. 2016, 398, 237–272. [Google Scholar] [CrossRef] [PubMed]
- Haas, L.F. Papyrus of Ebers and Smith. J. Neurol. Neurosurg. Psychiatry 1999, 67, 578. [Google Scholar] [CrossRef]
- Gelpi, A.; Gilbertson, A.; Tucker, J.D. Magic bullet: Paul Ehrlich, Salvarsan and the birth of venereology. Sex. Transm. Infect. 2015, 91, 68–69. [Google Scholar] [CrossRef] [PubMed]
- Otten, H. Domagk and the development of the sulphonamides. J. Antimicrob. Chemother. 1986, 17, 689–690. [Google Scholar] [CrossRef] [PubMed]
- Iyer, H.V. History revisited-prontosil red. J. Emerg. Med. 2008, 35, 209–210. [Google Scholar] [CrossRef]
- Hare, R. New light on the history of penicillin. Med. Hist. 1982, 26, 1–24. [Google Scholar] [CrossRef]
- Hutchings, M.; Truman, A.W.; Wilkinson, B. Antibiotics: Past, present and future. Curr. Opin. Microbiol. 2019, 51, 72–80. [Google Scholar] [CrossRef]
- Zaffiri, L.; Gardner, J.; Toledo-Pereyra, L.H. History of antibiotics. From Salvarsan to Cephalosporins. J. Investig. Surg. 2012, 25, 67–77. [Google Scholar] [CrossRef]
- Waksman, S.A.; Schatz, A.; Reynolds, D.M. production of antibiotic substances by actinomycetes. Ann. N. Y. Acad. Sci. 2010, 1213, 112–124. [Google Scholar] [CrossRef] [PubMed]
- Darville, T.; Yamauchi, T. The Cephalosporin antibiotics. Pediatr. Rev. 1994, 15, 54–62. [Google Scholar] [CrossRef]
- Vardanyan, R.; Hruby, V. Synthesis of Best-Seller Drugs; Academic Press: Cambridge, MA, USA, 2016; ISBN 978-0-12-411492-0. [Google Scholar]
- Butler, M.S.; Cooper, M.A. Antibiotics in the clinical pipeline in 2011. J. Antibiot. 2011, 64, 413–425. [Google Scholar] [CrossRef]
- Merck & Co. Overview of Antibiotics—Infections; Merck & Co.: Kenilworth, NJ, USA, 2020. [Google Scholar]
- Spížek, J.; Sigler, K.; Řezanka, T.; Demain, A. Biogenesis of antibiotics—Viewing its history and glimpses of the future. Folia Microbiol. 2016, 61, 347–358. [Google Scholar] [CrossRef] [PubMed]
- Adzitey, F. Antibiotic classes and antibiotic susceptibility of bacterial isolates from selected poultry; A mini review. Worlds Vet. J. 2015, 6, 36. [Google Scholar] [CrossRef]
- Oloke, J.K. Activity pattern of natural and synthetic antibacterial agents among hospital isolates. Microbios 2000, 102, 175–181. [Google Scholar] [PubMed]
- Ebimieowei, E.; Ibemologi, A. Antibiotics: Classification and mechanisms of action with emphasis on molecular perspectives. Int. J. Appl. Microbiol. Biotechnol. Res. 2016, 4, 90–101. [Google Scholar]
- James, C.W.; Gurk-Turner, C. Cross-reactivity of beta-lactam antibiotics. Baylor Univ. Med. Cent. Proc. 2001, 14, 106–107. [Google Scholar] [CrossRef]
- Hamilton-Miller, J.M.T. β-Lactams: Variations on a chemical theme, with some surprising biological results. J. Antimicrob. Chemother. 1999, 44, 729–734. [Google Scholar] [CrossRef]
- Chopra, I.; Roberts, M. Tetracycline Antibiotics: Mode of action, applications, molecular biology, and epidemiology of bacterial resistance. Microbiol. Mol. Biol. Rev. 2001, 65, 232–260. [Google Scholar] [CrossRef]
- Retsema, J.; Fu, W. Macrolides: Structures and microbial targets. Int. J. Antimicrob. Agents 2001, 18, 3–10. [Google Scholar] [CrossRef]
- Zhu, Z.J.; Krasnykh, O.; Pan, D.; Petukhova, V.; Yu, G.; Liu, Y.; Liu, H.; Hong, S.; Wang, Y.; Wan, B.; et al. Structure-activity relationships of macrolides against mycobacterium tuberculosis. Tuberculosis 2008, 88, S49–S63. [Google Scholar] [CrossRef]
- Heeb, S.; Fletcher, M.P.; Chhabra, S.R.; Diggle, S.P.; Williams, P.; Cámara, M. Quinolones: From antibiotics to autoinducers. FEMS Microbiol. Rev. 2011, 35, 247–274. [Google Scholar] [CrossRef] [PubMed]
- Kocsis, B.; Domokos, J.; Szabo, D. Chemical structure and pharmacokinetics of novel quinolone agents represented by avarofloxacin, delafloxacin, finafloxacin, zabofloxacin and nemonoxacin. Ann. Clin. Microbiol. Antimicrob. 2016, 15, 34. [Google Scholar] [CrossRef]
- Percival, K.M. Antibiotic classification and indication review for the infusion nurse. J. Infus. Nurs. 2017, 40, 55–63. [Google Scholar] [CrossRef]
- Cho, H.; Uehara, T.; Bernhardt, T.G. Beta-lactam antibiotics induce a lethal malfunctioning of the bacterial cell wall synthesis machinery. Cell 2014, 159, 1300–1311. [Google Scholar] [CrossRef]
- Kishida, H.; Unzai, S.; Roper, D.I.; Lloyd, A.; Park, S.-Y.; Tame, J.R.H. Crystal structure of penicillin binding protein 4 (dacB) from Escherichia coli, both in the native form and covalently linked to various antibiotics. Biochemistry 2006, 45, 783–792. [Google Scholar] [CrossRef]
- Kohanski, M.A.; Dwyer, D.J.; Collins, J.J. How antibiotics kill bacteria: From targets to networks. Nat. Rev. Genet. 2010, 8, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Tu, D.; Blaha, G.; Moore, P.B.; Steitz, T.A. Structures of MLSBK antibiotics bound to mutated large ribosomal subunits provide a structural explanation for resistance. Cell 2005, 121, 257–270. [Google Scholar] [CrossRef]
- Ippolito, J.A.; Kanyo, Z.F.; Wang, D.; Franceschi, F.J.; Moore, P.B.; Steitz, T.A.; Duffy, E.M. Crystal structure of the oxazolidinone antibiotic linezolid bound to the 50s ribosomal subunit. J. Med. Chem. 2008, 51, 3353–3356. [Google Scholar] [CrossRef]
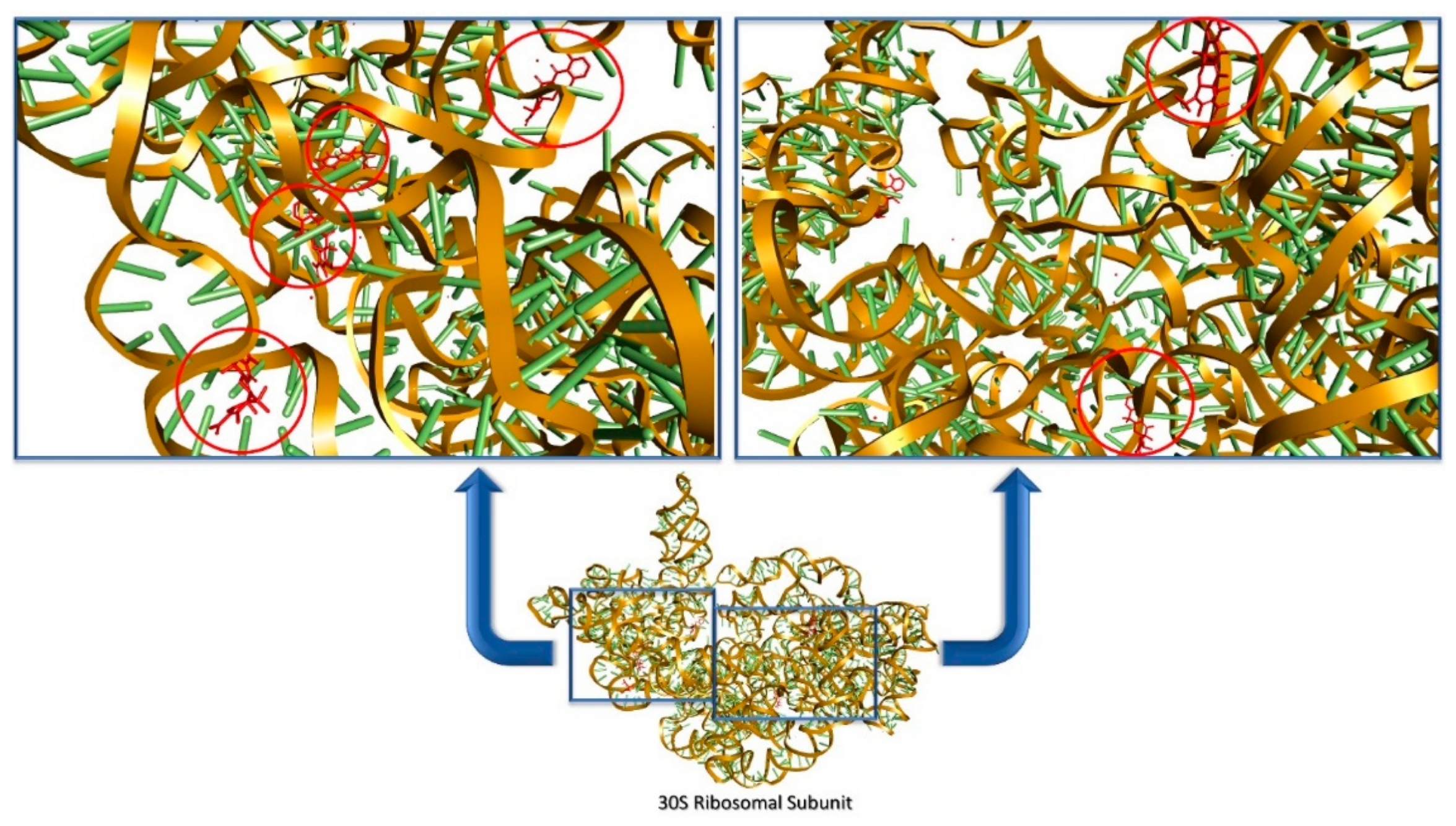
- Mehta, R.; Champney, W.S. 30S ribosomal subunit assembly is a target for inhibition by aminoglycosides in Escherichia coli. Antimicrob. Agents Chemother. 2002, 46, 1546–1549. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Atkinson, G.C.; Thakor, N.S.; Allas, Ü.; Lu, C.-C.; Chan, K.-Y.; Tenson, T.; Schulten, K.; Wilson, K.S.; Hauryliuk, V.; et al. Mechanism of tetracycline resistance by ribosomal protection protein Tet(O). Nat. Commun. 2013, 4, 1–8. [Google Scholar] [CrossRef]
- Pioletti, M.; Schlünzen, F.; Harms, J.; Zarivach, R.; Glühmann, M.; Avila, H.; Bashan, A.; Bartels, H.; Auerbach, T.; Jacobi, C.; et al. Crystal structures of complexes of the small ribosomal subunit with tetracycline, edeine and IF3. EMBO J. 2001, 20, 1829–1839. [Google Scholar] [CrossRef]
- Polikanov, Y.S.; Aleksashin, N.A.; Beckert, B.; Wilson, D.N. The mechanisms of action of ribosome-targeting peptide antibiotics. Front. Mol. Biosci. 2018, 5, 48. [Google Scholar] [CrossRef]
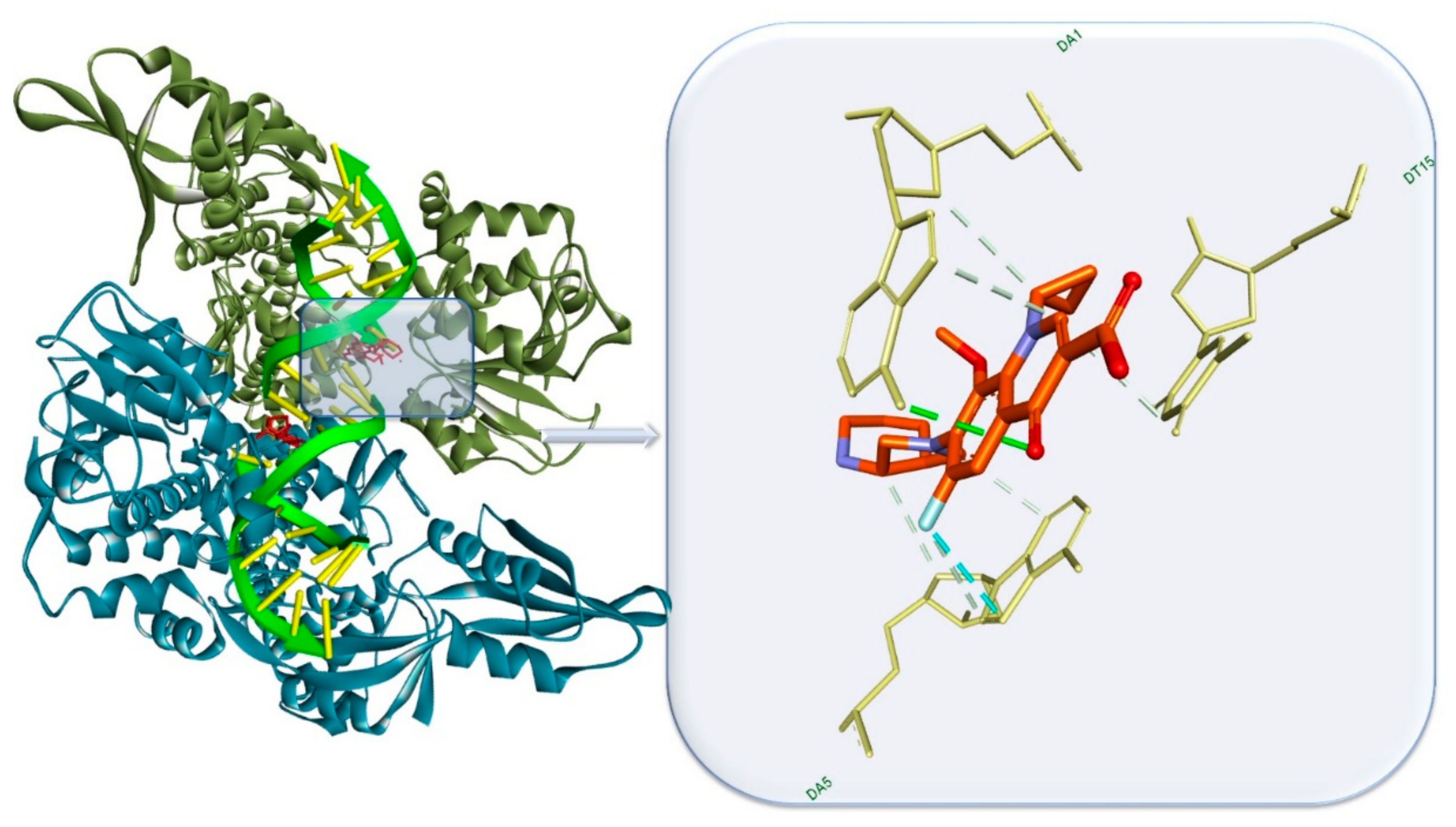
- Ma, C.; Yang, X.; Lewis, P.J. Bacterial transcription as a target for antibacterial drug development. Microbiol. Mol. Biol. Rev. 2016, 80, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Hooper, D.C.; Jacoby, G.A. Topoisomerase Inhibitors: Fluoroquinolone mechanisms of action and resistance. Cold Spring Harb. Perspect. Med. 2016, 6, a025320. [Google Scholar] [CrossRef]
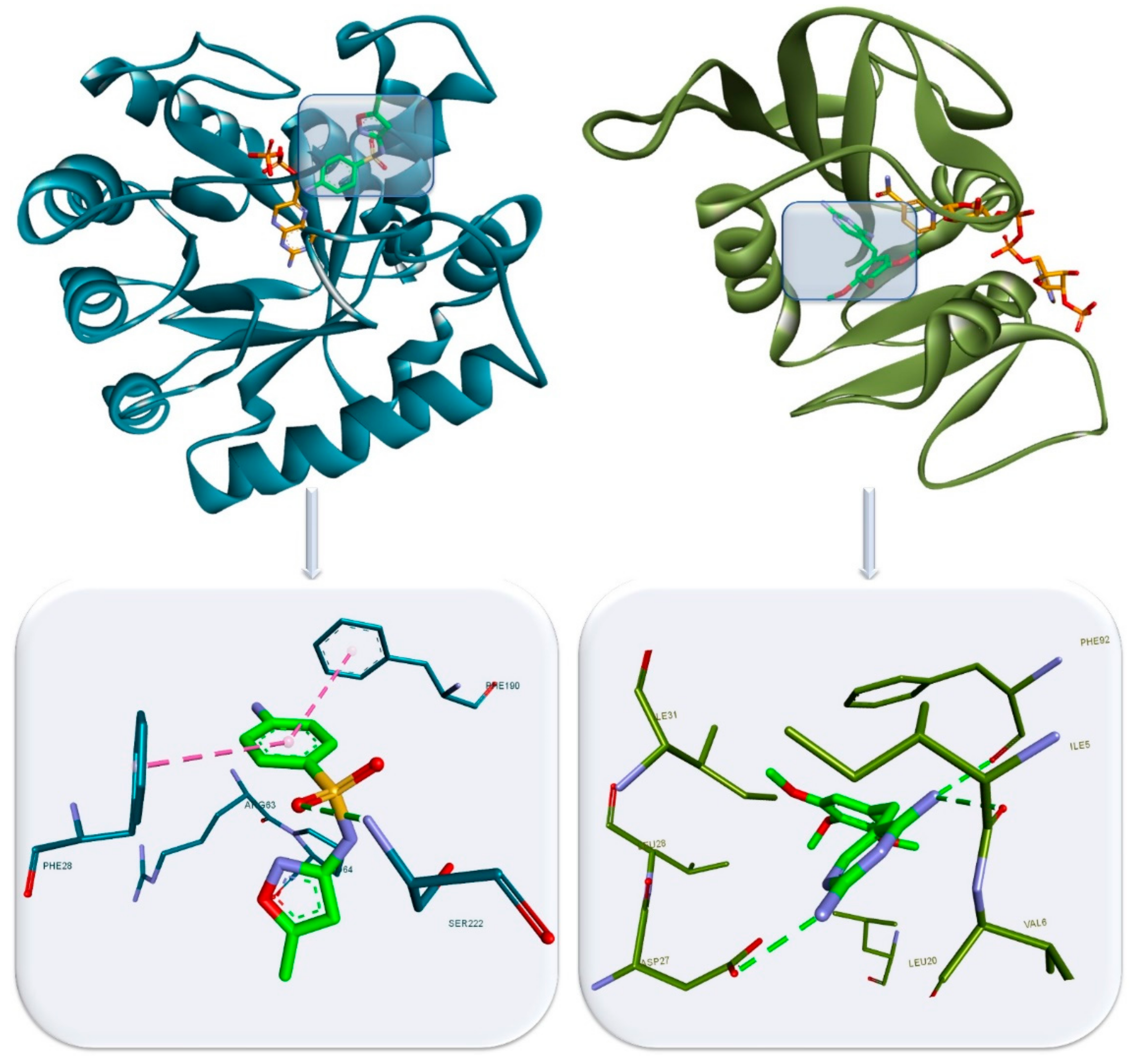
- Wróbel, A.; Maliszewski, D.; Baradyn, M.; Drozdowska, D. Trimethoprim: An old antibacterial drug as a template to search for new targets. synthesis, biological activity and molecular modeling study of novel trimethoprim analogs. Molecules 2019, 25, 116. [Google Scholar] [CrossRef]
- Heaslet, H.; Harris, M.; Fahnoe, K.; Sarver, R.; Putz, H.; Chang, J.; Subramanyam, C.; Barreiro, G.; Miller, J.R. Structural comparison of chromosomal and exogenous dihydrofolate reductase from Staphylococcus aureusin complex with the potent inhibitor trimethoprim. Proteins 2009, 76, 706–717. [Google Scholar] [CrossRef]
- Yun, M.-K.; Wu, Y.; Li, Z.; Zhao, Y.; Waddell, M.B.; Ferreira, A.M.; Lee, R.E.; Bashford, D.; White, S.W. Catalysis and sulfa drug resistance in dihydropteroate synthase. Science 2012, 335, 1110–1114. [Google Scholar] [CrossRef] [PubMed]
- Loree, J.; Lappin, S.L. Bacteriostatic Antibiotics; StatPearls Publishing: Treasure Island, FL, USA, 2020. [Google Scholar]
- Acar, J. Broad- and narrow-spectrum antibiotics: An unhelpful categorization. Clin. Microbiol. Infect. 1997, 3, 395–396. [Google Scholar] [CrossRef]
- Roberts, M.C. Antibiotic toxicity, interactions and resistance development. Periodontology 2000 2002, 28, 280–297. [Google Scholar] [CrossRef]
- Rolain, J.; Baquero, F. The refusal of the society to accept antibiotic toxicity: Missing opportunities for therapy of severe infections. Clin. Microbiol. Infect. 2016, 22, 423–427. [Google Scholar] [CrossRef]
- Souissi, S.B.; Abed, M.; Elhiki, L.; Fortemps, P.; Pirlot, M. Reducing the toxicity risk in antibiotic prescriptions by combining ontologies with a multiple criteria decision model. AMIA Annu. Symp. Proc. 2017, 2017, 1625–1634. [Google Scholar] [PubMed]
- Rouveix, B. Antibiotic safety assessment. Int. J. Antimicrob. Agents 2003, 21, 215–221. [Google Scholar] [CrossRef]
- Everts, S. Antibiotic side effects explained. Chem. Eng. News Arch. 2013, 91, 9. [Google Scholar] [CrossRef]
- Johnson, D.H.; Cunha, B.A. Drug Fever. Infect. Dis. Clin. N. Am. 1996, 10, 85–91. [Google Scholar] [CrossRef]
- Patel, R.A.; Gallagher, J.C. Drug Fever. Pharmacotherapy 2010, 30, 57–69. [Google Scholar] [CrossRef]
- Carr, A.; Swanson, C.; Penny, R.; Cooper, D.A. Clinical and laboratory markers of hypersensitivity to trimethoprim-sulfamethoxazole in patients with Pneumocystis carinii pneumonia and AIDS. J. Infect. Dis. 1993, 167, 180–185. [Google Scholar] [CrossRef]
- Albin, S.; Agarwal, S. Prevalence and characteristics of reported penicillin allergy in an urban outpatient adult population. Allergy Asthma Proc. 2014, 35, 489–494. [Google Scholar] [CrossRef] [PubMed]
- Nayak, S.; Acharjya, B. Adverse cutaneous drug reaction. Indian J. Dermatol. 2008, 53, 2–8. [Google Scholar] [CrossRef]
- Vassileva, S.G.; Mateev, G.; Parish, L.C. Antimicrobial photosensitive reactions. Arch. Intern. Med. 1998, 158, 1993–2000. [Google Scholar] [CrossRef]
- Mandell, L.; Tillotson, G. Safety of fluoroquinolones: An update. Can. J. Infect. Dis. 2002, 13, 54–61. [Google Scholar] [CrossRef] [PubMed]
- Cunha, B.; Hamid, N.; Krol, V.; Eisenstein, L. Safety of meropenem in patients reporting penicillin allergy: Lack of allergic cross reactions. J. Chemother. 2008, 20, 233–237. [Google Scholar] [CrossRef]
- Patriarca, G.; Schiavino, D.; Lombardo, C.; Altomonte, G.; Decinti, M.; Buonomo, A.; Nucera, E. Tolerability of aztreonam in patients with IgE-mediated hypersensitivity to Beta-Lactams. Int. J. Immunopathol. Pharmacol. 2008, 21, 375–379. [Google Scholar] [CrossRef]
- Maker, J.H.; Stroup, C.M.; Huang, V.; James, S.F. Antibiotic hypersensitivity mechanisms. Pharmacy 2019, 7, 122. [Google Scholar] [CrossRef]
- Legendre, D.P.; Muzny, C.A.; Marshall, G.D.; Swiatlo, E. Antibiotic hypersensitivity reactions and approaches to desensitization. Clin. Infect. Dis. 2013, 58, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Wright, A.J. The penicillins. Mayo Clin. Proc. 1999, 74, 290–307. [Google Scholar] [CrossRef]
- Shukla, P.; Bansode, F.W.; Singh, R.K. Chloramphenicol toxicity: A review. J. Med. Med. Sci. 2011, 2, 1313–1316. [Google Scholar]
- Bang, N.U.; Kammer, R.B.; Lilly, E. Hematologic complications associated with β-Lactam antibiotics [with discussion]. Kammer Source: Reviews of Infectious Diseases: A clinical perspective of antibiotic therapy. Aminogly 2016, 5, S380–S393. [Google Scholar]
- Lang, R.; Lishner, M.; Ravid, M. Adverse reactions to prolonged treatment with high doses of carbenicillin and ureidopenicillins. Rev. Infect. Dis. 1990, 13, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Babiak, L.M.; Rybak, M.J. Hematological effects associated with Beta-Lactam use. Drug Intell. Clin. Pharm. 1986, 20, 833–836. [Google Scholar] [CrossRef]
- Grill, M.F.; Maganti, R.K. Neurotoxic effects associated with antibiotic use: Management considerations. Br. J. Clin. Pharmacol. 2011, 72, 381–393. [Google Scholar] [CrossRef]
- Chow, K.M.; Hui, A.C.; Szeto, C.C. Neurotoxicity induced by beta-lactam antibiotics: From bench to bedside. Eur. J. Clin. Microbiol. Infect. Dis. 2005, 24, 649–653. [Google Scholar] [CrossRef]
- Grill, M.F.; Maganti, R. Cephalosporin-induced neurotoxicity: Clinical manifestations, potential pathogenic mechanisms, and the role of electroencephalographic monitoring. Ann. Pharmacother. 2008, 42, 1843–1850. [Google Scholar] [CrossRef]
- Alavi, S.A.N.; Rezaei, N.J.; Bazzazi, A.M. Neurotoxicity of the antibiotics: A comprehensive study. Neurol. India 2018, 66, 1732–1740. [Google Scholar] [CrossRef]
- Owens, J.R.C.; Ambrose, P.G. Antimicrobial safety: Focus on fluoroquinolones. Clin. Infect. Dis. 2005, 41, S144–S157. [Google Scholar] [CrossRef]
- Kushner, J.M.; Peckman, H.J.; Snyder, C.R. Seizures associated with fluoroquinolones. Ann. Pharmacother. 2001, 35, 1194–1198. [Google Scholar] [CrossRef]
- Schmuck, G.; Schürmann, A.; Schlüter, G. Determination of the excitatory potencies of fluoroquinolones in the central nervous system by an in vitro model. Antimicrob. Agents Chemother. 1998, 42, 1831–1836. [Google Scholar] [CrossRef] [PubMed]
- Stahlmann, R. Clinical toxicological aspects of fluoroquinolones. Toxicol. Lett. 2002, 127, 269–277. [Google Scholar] [CrossRef]
- Watanabe, I.; Hodges, G.R.; Dworzack, D.L.; Kepes, J.J.; Duensing, G.F. Neurotoxicity of intrathecal gentamicin: A case report and experimental study. Ann. Neurol. 1978, 4, 564–572. [Google Scholar] [CrossRef] [PubMed]
- Di Poggio, M.B.; Anfosso, S.; Audenino, D.; Primavera, A. Clarithromycin-induced neurotoxicity in adults. J. Clin. Neurosci. 2011, 18, 313–318. [Google Scholar] [CrossRef]
- Mellinkoff, S.M. Gastrointestinal complications of antibiotic therapy. Am. J. Gastroenterol. 1954, 22, 295–300. [Google Scholar]
- Wood, M. The tolerance and toxicity of clarithromycin. J. Hosp. Infect. 1991, 19, 39–46. [Google Scholar] [CrossRef]
- Surawicz, C.M. Antibiotic-associated diarrhea and pseudomembranous colitis: Are they less common with poorly absorbed antimicrobials? Chemotherapy 2005, 51, 81–89. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.G. Antibiotic-associated diarrhea. Clin. Infect. Dis. 1992, 15, 573–581. [Google Scholar] [CrossRef]
- Morales-Alvarez, M.C. Nephrotoxicity of antimicrobials and antibiotics. Adv. Chronic Kidney Dis. 2020, 27, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.S.; Haikal, A.; Hammoud, K.A.; Yu, A.S. Vancomycin and the risk of AKI: A systematic review and meta-analysis. Clin. J. Am. Soc. Nephrol. 2016, 11, 2132–2140. [Google Scholar] [CrossRef]
- Muriithi, A.K.; Nasr, S.H.; Leung, N. Utility of urine eosinophils in the diagnosis of acute interstitial nephritis. Clin. J. Am. Soc. Nephrol. 2013, 8, 1857–1862. [Google Scholar] [CrossRef]
- Cotner, S.E.; Rutter, W.C.; Burgess, D.R.; Wallace, K.L.; Martin, C.A.; Burgess, D.S. Influence of β-lactam infusion strategy on acute kidney injury. Antimicrob. Agents Chemother. 2017, 61, e00871-17. [Google Scholar] [CrossRef]
- Guo, D.; Cai, Y.; Chai, D.; Liang, B.; Bai, N.; Wang, R. The cardiotoxicity of macrolides: A systematic review. Pharmazie 2010, 65, 631–640. [Google Scholar] [CrossRef]
- Lu, Z.K.; Yuan, J.; Li, M.; Sutton, S.S.; A Rao, G.; Jacob, S.; Bennett, C.L. Cardiac risks associated with antibiotics: Azithromycin and levofloxacin. Expert Opin. Drug Saf. 2015, 14, 295–303. [Google Scholar] [CrossRef]
- Adams, H.R.; Parker, J.L.; Durrett, L.R. Cardiac toxicities of antibiotics. Environ. Health Perspect. 1978, 26, 217–223. [Google Scholar] [CrossRef]
- Lazarczyk, D.A.; Goldstein, N.A.; Gordon, S.C. Trovafloxacin hepatotoxicity. Am. J. Gastroenterol. 2000, 95, 2585. [Google Scholar] [CrossRef]
- Westphal, J.F.; Vetter, D.; Brogard, J.M. Hepatic side-effects of antibiotics. J. Antimicrob. Chemother. 1994, 33, 387–401. [Google Scholar] [CrossRef] [PubMed]
- Slama, T.G.; Amin, A.; Brunton, S.A.; File, T.M.; Milkovich, G.; Rodvold, K.A.; Sahm, D.F.; Varon, J.; Weiland, D. A clinician’s guide to the appropriate and accurate use of antibiotics: The Council for Appropriate and Rational Antibiotic Therapy (CARAT) criteria. Am. J. Med. 2005, 118, 1–6. [Google Scholar] [CrossRef]
- Soumerai, S.B. Principles of educational outreach (‘academic detailing’) to improve clinical decision making. J. Am. Med. Assoc. 1990, 263, 549–556. [Google Scholar] [CrossRef]
- File, T.M.; Hadley, J.A. Rational use of antibiotics to treat respiratory tract infections. Am. J. Manag. Care 2002, 8, 713–727. [Google Scholar]
- Nicolau, D. Clinical and economic implications of antimicrobial resistance for the management of community-acquired respiratory tract infections. J. Antimicrob. Chemother. 2002, 50, 61–70. [Google Scholar] [CrossRef][Green Version]
- Aminov, R.I. A brief history of the antibiotic era: Lessons learned and challenges for the future. Front. Microbiol. 2010, 1, 134. [Google Scholar] [CrossRef] [PubMed]
- Levy, S.B.; Marshall, B. Antibacterial resistance worldwide: Causes, challenges and responses. Nat. Med. 2004, 10, S122–S129. [Google Scholar] [CrossRef]
- Waglechner, N.; Wright, G.D. Antibiotic resistance: It’s bad, but why isn’t it worse? BMC Biol. 2017, 15, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Chopra, R.; Alderborn, G.; Podczeck, F.; Newton, J. The influence of pellet shape and surface properties on the drug release from uncoated and coated pellets. Int. J. Pharm. 2002, 239, 171–178. [Google Scholar] [CrossRef]
- Davies, J.; Davies, D. Origins and evolution of antibiotic resistance. Microbiol. Mol. Biol. Rev. 2010, 74, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Bin Zaman, S.; Hussain, M.A.; Nye, R.; Mehta, V.; Mamun, K.T.; Hossain, N. A review on antibiotic resistance: Alarm bells are ringing. Cureus 2017, 9, e1403. [Google Scholar] [CrossRef] [PubMed]
- Aslam, B.; Wang, W.; Arshad, M.I.; Khurshid, M.; Muzammil, S.; Rasool, M.H.; Nisar, M.A.; Alvi, R.F.; Aslam, M.A.; Qamar, M.U.; et al. Antibiotic resistance: A rundown of a global crisis. Infect. Drug Resist. 2018, 11, 1645–1658. [Google Scholar] [CrossRef]
- Mahon, C.R.; Lehman, D.C.; Manuselis, G. Textbook of Diagnostic Microbiology E-Book; Elsevier Health Sciences: Philadelphia, PA, USA, 2018; ISBN 0323482120. [Google Scholar]
- Gupta, P.D.; Birdi, T.J. Development of botanicals to combat antibiotic resistance. J. Ayurveda Integr. Med. 2017, 8, 266–275. [Google Scholar] [CrossRef]
- WHO. GlobalDALYmethodsJapan_2011_WHO; World Health Organization: Geneva, Switzerland, 2014. [Google Scholar]
- Högberg, L.D.; Heddini, A.; Cars, O. The global need for effective antibiotics: Challenges and recent advances. Trends Pharmacol. Sci. 2010, 31, 509–515. [Google Scholar] [CrossRef]
- Niu, G.; Li, W. Next-generation drug discovery to combat antimicrobial resistance. Trends Biochem. Sci. 2019, 44, 961–972. [Google Scholar] [CrossRef]
- Takó, M.; Kerekes, E.B.; Zambrano, C.; Kotogán, A.; Papp, T.; Krisch, J.; Vágvölgyi, C. Plant phenolics and phenolic-enriched extracts as antimicrobial agents against food-contaminating microorganisms. Antioxidants 2020, 9, 165. [Google Scholar] [CrossRef]
- Guimarães, A.C.; Meireles, L.M.; Lemos, M.F.; Guimarães, M.C.C.; Endringer, D.C.; Fronza, M.; Scherer, R. Antibacterial activity of terpenes and terpenoids present in essential oils. Molecules 2019, 24, 2471. [Google Scholar] [CrossRef] [PubMed]
- Cushnie, T.T.; Cushnie, B.; Lamb, A.J. Alkaloids: An overview of their antibacterial, antibiotic-enhancing and antivirulence activities. Int. J. Antimicrob. Agents 2014, 44, 377–386. [Google Scholar] [CrossRef]
- Coelho, L.B.B.; Silva, P.M.D.S.; De Oliveira, W.F.; De Moura, M.; Pontual, E.V.; Gomes, F.S.; Paiva, P.G.; Napoleão, T.; Correia, M.D.S. Lectins as antimicrobial agents. J. Appl. Microbiol. 2018, 125, 1238–1252. [Google Scholar] [CrossRef] [PubMed]
- Anyiam, I.V. Antibacterial properties of chewing stick on bacteria isolated from the mouth of individuals in Amaigbo, Imo State. Int. J. Basic Sci. Technol. 2018, 2, 71–75. [Google Scholar]
- Savoia, D. Plant-derived antimicrobial compounds: Alternatives to antibiotics. Future Microbiol. 2012, 7, 979–990. [Google Scholar] [CrossRef]
- Del Rio, D.; Rodriguez-Mateos, A.; Spencer, J.P.; Tognolini, M.; Borges, G.; Crozier, A. Dietary (poly)phenolics in human health: Structures, bioavailability, and evidence of protective effects against chronic diseases. Antiox. Redox Signal. 2013, 18, 1818–1892. [Google Scholar] [CrossRef] [PubMed]
- Górniak, I.; Bartoszewski, R.; Króliczewski, J. Comprehensive review of antimicrobial activities of plant flavonoids. Phytochem. Rev. 2019, 18, 241–272. [Google Scholar] [CrossRef]
- Shahzad, M.; Millhouse, E.; Culshaw, S.; Edwards, C.A.; Ramage, G.; Combet, E. Selected dietary (poly)phenols inhibit periodontal pathogen growth and biofilm formation. Food Funct. 2014, 6, 719–729. [Google Scholar] [CrossRef]
- Taguri, T.; Tanaka, T.; Kouno, I. Antibacterial spectrum of plant polyphenols and extracts depending upon hydroxyphenyl structure. Biol. Pharm. Bull. 2006, 29, 2226–2235. [Google Scholar] [CrossRef]
- Mason, T.L. Inactivation of red beet β-glucan synthase by native and oxidized phenolic compounds. Phytochemistry 1987, 26, 2197–2202. [Google Scholar] [CrossRef]
- Lim, A.; Subhan, N.; Jazayeri, J.A.; John, G.; Vanniasinkam, T.; Obied, H.K. Plant phenols as antibiotic boosters: In vitro interaction of olive leaf phenols with ampicillin. Phytother. Res. 2016, 30, 503–509. [Google Scholar] [CrossRef]
- Sarjit, A.; Wang, Y.; Dykes, G.A. Antimicrobial activity of gallic acid against thermophilic Campylobacter is strain specific and associated with a loss of calcium ions. Food Microbiol. 2015, 46, 227–233. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.-K.; Moo, C.-L.; Song, A.A.-L.; Chong, C.-M.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.E.; Lai, K.-S. Terpene derivatives as a potential agent against antimicrobial resistance (AMR) pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef]
- Griffin, S.G.; Wyllie, S.G.; Markham, J.L.; Leach, D.N. The role of structure and molecular properties of terpenoids in de-termining their antimicrobial activity. Flavour Fragr. J. 1999, 14, 322–332. [Google Scholar] [CrossRef]
- Lorenzi, V.; Muselli, A.; Bernardini, A.F.; Berti, L.; Pagès, J.-M.; Amaral, L.; Bolla, J.-M. Geraniol restores antibiotic activities against multidrug-resistant isolates from gram-negative species. Antimicrob. Agents Chemother. 2009, 53, 2209–2211. [Google Scholar] [CrossRef] [PubMed]
- Bhatti, H.N.; Khan, S.S.; Khan, A.; Rani, M.; Ahmad, V.U.; Choudhary, M.I. Biotransformation of monoterpenoids and their antimicrobial activities. Phytomedicine 2014, 21, 1597–1626. [Google Scholar] [CrossRef] [PubMed]
- Koo, H.; Pearson, S.K.; Scott-Anne, K.; Abranches, J.; Cury, J.A.; Rosalen, P.L.; Park, Y.K.; Marquis, R.E.; Bowen, W.H. Effects of apigenin and tt-farnesol on glucosyltransferase activity, biofilm viability and caries development in rats. Oral Microbiol. Immunol. 2002, 17, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Gomes, F.I.A.; Teixeira, P.; Azeredo, J.; Oliveira, R. Effect of farnesol on planktonic and biofilm cells of Staphylococcus epidermidis. Curr. Microbiol. 2009, 59, 118–122. [Google Scholar] [CrossRef]
- Masako, K.; Yusuke, K.; Hideyuki, I.; Atsuko, M.; Yoshiki, M.; Kayoko, M.; Makoto, K. A novel method to control the balance of skin microflora. Part 2. A study to assess the effect of a cream containing farnesol and xylitol on atopic dry skin. J. Dermatol. Sci. 2005, 38, 207–213. [Google Scholar] [CrossRef]
- Castelo-Branco, D.; Riello, G.B.; Vasconcelos, D.C.; Guedes, G.M.M.; Serpa, R.C.; Bandeira, T.J.P.G.; Monteiro, A.J.; Cordeiro, R.A.; Rocha, M.F.G.; Sidrim, J.J.C.; et al. Farnesol increases the susceptibility of Burkholderia pseudomallei biofilm to antimicrobials used to treat melioidosis. J. Appl. Microbiol. 2016, 120, 600–606. [Google Scholar] [CrossRef]
- Zacchino, S.A.; Butassi, E.; Cordisco, E.; Svetaz, L.A. Hybrid combinations containing natural products and antimicrobial drugs that interfere with bacterial and fungal biofilms. Phytomedicine 2017, 37, 14–26. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.K.; Tiwari, N.; Gupta, P.; Verma, S.; Pal, A.; Srivastava, S.K.; Darokar, M.P. A clerodane diterpene from Polyalthia longifolia as a modifying agent of the resistance of methicillin resistant Staphylococcus aureus. Phytomedicine 2016, 23, 654–661. [Google Scholar] [CrossRef]
- Walencka, E.; Rozalska, S.; Wysokinska, H.; Rozalski, M.; Kuzma, L.; Rozalska, B. Salvipisone and aethiopinone from Salvia sclarea hairy roots modulate staphylococcal antibiotic resistance and express anti-biofilm activity. Planta Med. 2007, 73, 545–551. [Google Scholar] [CrossRef]
- Barbieri, R.; Coppo, E.; Marchese, A.; Daglia, M.; Sobarzo-Sánchez, E.; Nabavi, S.F. Phytochemicals for human disease: An update on plant-derived compounds antibacterial activity. Microbiol. Res. 2017, 196, 44–68. [Google Scholar] [CrossRef]
- Jiménez-Arellanes, A.; Luna-Herrera, J.; Cornejo-Garrido, J.; López-García, S.; Castro-Mussot, M.E.; Meckes-Fischer, M.; Mata-Espinosa, D.; Marquina, B.; Torres, J.; Hernández-Pando, R. Ursolic and oleanolic acids as antimicrobial and immunomodulatory compounds for tuberculosis treatment. BMC Complement. Altern. Med. 2013, 13, 258. [Google Scholar] [CrossRef]
- Chung, P.Y.; Chung, L.Y.; Navaratnam, P. Potential targets by pentacyclic triterpenoids from Callicarpa farinosa against methicillin-resistant and sensitive Staphylococcus aureus. Fitoterapia 2014, 94, 48–54. [Google Scholar] [CrossRef] [PubMed]
- Raut, J.S.; Karuppayil, S.M. A status review on the medicinal properties of essential oils. Ind. Crops Prod. 2014, 62, 250–264. [Google Scholar] [CrossRef]
- Sandri, I.; Zacaria, J.; Fracaro, F.; Delamare, A.; Echeverrigaray, S. Antimicrobial activity of the essential oils of Brazilian species of the genus Cunila against foodborne pathogens and spoiling bacteria. Food Chem. 2007, 103, 823–828. [Google Scholar] [CrossRef]
- Oussalah, M.; Caillet, S.; Lacroix, M. Mechanism of action of Spanish oregano, Chinese cinnamon, and savory essential oils against cell membranes and walls of Escherichia coli O157:H7 and Listeria monocytogenes. J. Food Prot. 2006, 69, 1046–1055. [Google Scholar] [CrossRef] [PubMed]
- Saad, N.Y.; Muller, C.D.; Lobstein, A. Major bioactivities and mechanism of action of essential oils and their components. Flavour Fragr. J. 2013, 28, 269–279. [Google Scholar] [CrossRef]
- Casu, L.; Cottiglia, F.; Leonti, M.; De Logu, A.; Agus, E.; Tse-Dinh, Y.-C.; Lombardo, V.; Sissi, C. Ungeremine effectively targets mammalian as well as bacterial type I and type II topoisomerases. Bioorg. Med. Chem. Lett. 2011, 21, 7041–7044. [Google Scholar] [CrossRef]
- Tominaga, K.; Higuchi, K.; Hamasaki, N.; Hamaguchi, M.; Takashima, T.; Tanigawa, T.; Watanabe, T.; Fujiwara, Y.; Tezuka, Y.; Nagaoka, T.; et al. In vivo action of novel alkyl methyl quinolone alkaloids against Helicobacter pylori. J. Antimicrob. Chemother. 2002, 50, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Alhanout, K.; Malesinki, S.; Vidal, N.; Peyrot, V.; Rolain, J.M.; Brunel, J.M. New insights into the antibacterial mechanism of action of squalamine. J. Antimicrob. Chemother. 2010, 65, 1688–1693. [Google Scholar] [CrossRef]
- Hraiech, S.; Brégeon, F.; Brunel, J.-M.; Rolain, J.-M.; Lepidi, H.; Andrieu, V.; Raoult, D.; Papazian, L.; Roch, A. Antibacterial efficacy of inhaled squalamine in a rat model of chronic Pseudomonas aeruginosa pneumonia. J. Antimicrob. Chemother. 2012, 67, 2452–2458. [Google Scholar] [CrossRef]
- Hung, D.T.; Shakhnovich, E.A.; Pierson, E.; Mekalanos, J.J. Small-molecule inhibitor of vibrio cholerae virulence and intestinal colonization. Science 2005, 310, 670–674. [Google Scholar] [CrossRef]
- Chatterjee, A.; Ratner, D.M.; Ryan, C.M.; Johnson, P.J.; O’Keefe, B.R.; Secor, W.E.; Anderson, D.J.; Robbins, P.W.; Samuelson, J. Anti-retroviral lectins have modest effects on adherence of Trichomonas vaginalis to epithelial cells in vitro and on recovery of Tritrichomonas foetus in a mouse vaginal model. PLoS ONE 2015, 10, e0135340. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, B.; Martins, M.B.; Karamanska, R.; Russell, D.A.; Field, R.A. Bacterial detection using carbohydrate-functionalised CdS Quantum dots: A model study exploiting E. coli recognition of mannosides. Tetrahedron Lett. 2009, 50, 886–889. [Google Scholar] [CrossRef]
- Iordache, F.; Ionita, M.; Mitrea, L.I.; Fafaneata, C.; Pop, A. Antimicrobial and antiparasitic activity of lectins. Curr. Pharm. Biotechnol. 2015, 16, 152–161. [Google Scholar] [CrossRef] [PubMed]
- Nair, S.S.; Madembil, N.C.; Nair, P.; Raman, S.; Veerabadrappa, S.B. Comparative analysis of the antibacterial activity of some phytolectins. Int. Curr. Pharm. J. 2013, 2, 18–22. [Google Scholar] [CrossRef]
- Petnual, P.; Sangvanich, P.; Karnchanatat, A. A lectin from the rhizomes of turmeric (Curcuma longa L.) and its antifungal, antibacterial, and α-glucosidase inhibitory activities. Food Sci. Biotechnol. 2010, 19, 907–916. [Google Scholar] [CrossRef]
- Nawrot, R.; Barylski, J.; Nowicki, G.; Broniarczyk, J.; Buchwald, W.; Goździcka-Józefiak, A. Plant antimicrobial peptides. Folia Microbiol. 2014, 59, 181–196. [Google Scholar] [CrossRef]
- Miyazaki, Y.; Aoki, M.; Yano, Y.; Matsuzaki, K. Interaction of antimicrobial peptide magainin 2 with gangliosides as a target for human cell binding. Biochemistry 2012, 51, 10229–10235. [Google Scholar] [CrossRef]
- Mander, L.; Liu, H.-W. Comprehensive Natural Products II: Chemistry and Biology; Elsevier: Amsterdam, The Netherlands, 2010; Volume 1, ISBN 0080453821. [Google Scholar]
- Franco, O.L.; Murad, A.M.; Leite, J.R.; Mendes, P.A.M.; Prates, M.V.; Bloch, C. Identification of a cowpea γ-thionin with bactericidal activity. FEBS J. 2006, 273, 3489–3497. [Google Scholar] [CrossRef]
- Zhang, Y.; Lewis, K. Fabatins: New antimicrobial plant peptides. FEMS Microbiol. Lett. 2006, 149, 59–64. [Google Scholar] [CrossRef]
- Dogra, S.; Biswas, R.; Biswas, K. Plant antimicrobial peptides: A novel approach against drug resistant microorganisms. Int. J. Pharm. Sci. Res. 2018, 9, 1–15. [Google Scholar] [CrossRef]
- Kwon, D.H.; Lu, C.-D. Polyamines increase antibiotic susceptibility in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 2006, 50, 1623–1627. [Google Scholar] [CrossRef] [PubMed]
- Romeo, L.; Iori, R.; Rollin, P.; Bramanti, P.; Mazzon, E. Isothiocyanates: An overview of their antimicrobial activity against human infections. Molecules 2018, 23, 624. [Google Scholar] [CrossRef] [PubMed]
- Benmerache, A.; Magid, A.A.; Berrehal, D.; Kabouche, A.; Voutquenne-Nazabadioko, L.; Messaili, S.; Abedini, A.; Harakat, D.; Kabouche, Z. Chemical composition, antibacterial, antioxidant and tyrosinase inhibitory activities of glycosides from aerial parts of Eryngium tricuspidatum L. Phytochem. Lett. 2016, 18, 23–28. [Google Scholar] [CrossRef]
- Kulikova, V.V.; Chernukha, M.Y.; Morozova, E.A.; Revtovich, S.V.; Rodionov, A.N.; Koval, V.S.; Avetisyan, L.R.; Kuliastova, D.G.; Shaginyan, I.A.; Demidkina, T.V. Antibacterial effect of thiosulfinates on multiresistant strains of bacteria isolated from patients with cystic fibrosis. Acta Naturae 2018, 10, 77–80. [Google Scholar] [CrossRef]
- Lu, C.-D.; Itoh, Y.; Nakada, Y.; Jiang, Y. Functional analysis and regulation of the divergent spuABCDEFGH-spuI Operons for polyamine uptake and utilization in Pseudomonas aeruginosa PAO1. J. Bacteriol. 2002, 184, 3765–3773. [Google Scholar] [CrossRef]
- Fahey, J.W.; Haristoy, X.; Dolan, P.M.; Kensler, T.W.; Scholtus, I.; Stephenson, K.K.; Talalay, P.; Lozniewski, A. Sulforaphane inhibits extracellular, intracellular, and antibiotic-resistant strains of Helicobacter pylori and prevents benzo[a]pyrene-induced stomach tumors. Proc. Natl. Acad. Sci. USA 2002, 99, 7610–7615. [Google Scholar] [CrossRef]
- Haristoy, X.; Angioi-Duprez, K.; Duprez, A.; Lozniewski, A. Efficacy of sulforaphane in eradicating Helicobacter pylori in human gastric xenografts implanted in nude mice. Antimicrob. Agents Chemother. 2003, 47, 3982–3984. [Google Scholar] [CrossRef]
- Soulef, K.; Yahia, A. Anti-bacterial effects of glycosides extract of Glycyrrhiza glabra L. from the region of Djamâa (South-East of Algeria). J. Med. Herbs Ethnomed. 1970, 3, 22–25. [Google Scholar] [CrossRef][Green Version]
- Leontiev, R.; Hohaus, N.; Jacob, C.; Gruhlke, M.C.H.; Slusarenko, A.J. A comparison of the antibacterial and antifungal activities of thiosulfinate analogues of allicin. Sci. Rep. 2018, 8, 1–19. [Google Scholar] [CrossRef]
- Romulo, A.; Zuhud, E.A.M.; Rondevaldova, J.; Kokoska, L. Screening of in vitro antimicrobial activity of plants used in traditional Indonesian medicine. Pharm. Biol. 2018, 56, 287–293. [Google Scholar] [CrossRef]
- Purkayastha, S.; Dahiya, P. Phytochemical screening and antimicrobial activity of some medicinal plants against multi-drug resistant bacteria from clinical isolates. Indian J. Pharm. Sci. 2012, 74, 443–450. [Google Scholar] [CrossRef]
- Khan, U.A.; Rahman, H.; Niaz, Z.; Qasim, M.; Khan, J.; Tayyaba; Rehman, B. Antibacterial activity of some medicinal plants against selected human pathogenic bacteria. Eur. J. Microbiol. Immunol. 2013, 3, 272–274. [Google Scholar] [CrossRef] [PubMed]
- Girondi, C.M.; De Oliveira, A.B.; Prado, J.A.; Koga-Ito, C.Y.; Borges, A.C.; Delbem, A.C.B.; Pereira, D.F.A.; Salvador, M.J.; Brighenti, F.L. Screening of plants with antimicrobial activity against enterobacteria, Pseudomonas spp. and Staphylococcus spp. Future Microbiol. 2017, 12, 671–681. [Google Scholar] [CrossRef] [PubMed]
- Kakar, S.A.; Tareen, R.B.; Azam Kakar, M.; Jabeen, H.; Kakar, S.U.R.; Al-Kahraman, Y.M.S.A.; Shafee, M. Screening of an-tibacterial activity of four medicinal plants of Balochistan-Pakistan. Pakistan J. Bot. 2012, 44, 245–250. [Google Scholar]
- Nascimento, G.G.F.; Locatelli, J.; Freitas, P.C.; Silva, G.L. Antibacterial activity of plant extracts and phytochemicals on antibiotic-resistant bacteria. Braz. J. Microbiol. 2000, 31, 247–256. [Google Scholar] [CrossRef]
- Bahri-Sahloul, R.; Fredj, R.B.; Boughalleb, N.; Shriaa, J.; Saguem, S.; Hilbert, J.-L.; Trotin, F.; Ammar, S.; Bouzid, S.; Harzallah-Skhiri, F. Phenolic composition and antioxidant and antimicrobial activities of extracts obtained from Crataegus azarolus L. var. Aronia (Willd.) Batt. Ovaries Calli. J. Bot. 2014, 2014, 1–11. [Google Scholar] [CrossRef]
- Brighenti, F.L.; Salvador, M.; Delbem, A.C.B.; Delbem, Á.; Oliveira, M.; Soares, C.; Freitas, L.; Koga-Ito, C.Y. Systematic screening of plant extracts from the Brazilian pantanal with antimicrobial activity against bacteria with cariogenic relevance. Caries Res. 2014, 48, 353–360. [Google Scholar] [CrossRef]
- Luiz, J.; Júnior, R.; Stuker, C.Z.; Peruzzo, G.M.; Peres, M.T.L.P. Essential oil of Croton Urucurana Baillon (Euphorbiaceae) Stem bark. J. Braz. Chem. Soc. 2007, 18, 879–885. [Google Scholar]
- Al-Daihan, S.; Al-Faham, M.; Al-Shawi, N.; Almayman, R.; Brnawi, A.; Zargar, S.; Bhat, R.S. Antibacterial activity and phytochemical screening of some medicinal plants commonly used in Saudi Arabia against selected pathogenic microorganisms. J. King Saud Univ. Sci. 2013, 25, 115–120. [Google Scholar] [CrossRef]
- Waheed, K.; Muhammad, S.; Shomaila, A.; Muhammad, Z.; Izhar, U.; Ullah, S. Antimicrobial activity and phytochemical screening of Euphorbia helioscopia. Planta Daninha 2020, 38, 38. [Google Scholar] [CrossRef]
- Özkan, G.; Sagdiç, O.; Baydar, N.G.; Kurumahmutoglu, Z. Antibacterial activities and total phenolic contents of grape pomace extracts. J. Sci. Food Agric. 2004, 84, 1807–1811. [Google Scholar] [CrossRef]
- Sommer, F.; Bäckhed, F. The gut microbiota—Masters of host development and physiology. Nat. Rev. Microbiol. 2013, 11, 227–238. [Google Scholar] [CrossRef] [PubMed]
- Rajilić-Stojanović, M.; De Vos, W.M. The first 1000 cultured species of the human gastrointestinal microbiota. FEMS Microbiol. Rev. 2014, 38, 996–1047. [Google Scholar] [CrossRef]
- Kim, S.; Covington, A.; Pamer, E.G. The intestinal microbiota: Antibiotics, colonization resistance, and enteric pathogens. Immunol. Rev. 2017, 279, 90–105. [Google Scholar] [CrossRef]
- Popa, G.-L.; Gheorghe, S.-A.; Preda, M.; Popa, M.I. The intestinal microbiota reconfigures the boundaries of knowledge. Infect. 2017, 1. [Google Scholar] [CrossRef]
- Bermon, S.; Petriz, B.; Kajeniene, A.; Prestes, J.; Castell, L.; Franco, O.L. The microbiota: An exercise immunology perspective. Exerc. Immunol Rev. 2015, 21, 70–79. [Google Scholar]
- Vila, A.V.; Collij, V.; Sanna, S.; Sinha, T.; Imhann, F.; Bourgonje, A.R.; Mujagic, Z.; Jonkers, D.M.A.E.; Masclee, A.A.M.; Fu, J.; et al. Impact of commonly used drugs on the composition and metabolic function of the gut microbiota. Nat. Commun. 2020, 11, 1–11. [Google Scholar] [CrossRef]
- Zimmermann, P.; Curtis, N. The effect of antibiotics on the composition of the intestinal microbiota—A systematic review. J. Infect. 2019, 79, 471–489. [Google Scholar] [CrossRef] [PubMed]
- Ianiro, G.; Tilg, H.; Gasbarrini, A. Antibiotics as deep modulators of gut microbiota: Between good and evil. Gut 2016, 65, 1906–1915. [Google Scholar] [CrossRef]
- Jernberg, C.; Löfmark, S.; Edlund, C.; Jansson, J.K. Long-term impacts of antibiotic exposure on the human intestinal microbiota. Microbiology 2010, 156, 3216–3223. [Google Scholar] [CrossRef] [PubMed]
- Morgun, A.; Dzutsev, A.; Dong, X.; Greer, R.L.; Sexton, D.J.; Ravel, J.; Schuster, M.; Hsiao, W.; Matzinger, P.; Shulzhenko, N. Uncovering effects of antibiotics on the host and microbiota using transkingdom gene networks. Gut 2015, 64, 1732–1743. [Google Scholar] [CrossRef] [PubMed]
- Slimings, C.; Riley, T.V. Antibiotics and hospital-acquired Clostridium difficile infection: Update of systematic review and meta-analysis. J. Antimicrob. Chemother. 2014, 69, 881–891. [Google Scholar] [CrossRef]
- Jernberg, C.; Löfmark, S.; Edlund, C.; Jansson, J.K. Long-term ecological impacts of antibiotic administration on the human intestinal microbiota. ISME J. 2007, 1, 56–66. [Google Scholar] [CrossRef]
- Rashid, M.-U.; Zaura, E.; Buijs, M.J.; Keijser, B.J.F.; Crielaard, W.; Nord, C.E.; Weintraub, A. Determining the long-term effect of antibiotic administration on the human normal intestinal microbiota using culture and pyrosequencing methods. Clin. Infect. Dis. 2015, 60, S77–S84. [Google Scholar] [CrossRef]
- Jakobsson, H.E.; Jernberg, C.; Andersson, A.F.; Sjölund-Karlsson, M.; Jansson, J.K.; Engstrand, L. Short-term antibiotic treatment has differing long-term impacts on the human throat and gut microbiome. PLoS ONE 2010, 5, e9836. [Google Scholar] [CrossRef]
- Korpela, K.; Salonen, A.; Virta, L.J.; Kekkonen, R.A.; Forslund, K.; Bork, P.; De Vos, W.M. Intestinal microbiome is related to lifetime antibiotic use in Finnish pre-school children. Nat. Commun. 2016, 7, 10410. [Google Scholar] [CrossRef]
- Arboleya, S.; Sánchez, B.; Solís, G.; Fernández, N.; Suárez, M.; Hernández-Barranco, A.M.; Milani, C.; Margolles, A.; Reyes-Gavilán, C.G.D.L.; Ventura, M.; et al. Impact of prematurity and perinatal antibiotics on the developing intestinal microbiota: A functional inference study. Int. J. Mol. Sci. 2016, 17, 649. [Google Scholar] [CrossRef]
- Knecht, H.; Neulinger, S.C.; Heinsen, F.A.; Knecht, C.; Schilhabel, A.; Schmitz, R.A.; Zimmermann, A.; Dos Santos, V.M.; Ferrer, M.; Rosenstiel, P.C.; et al. Effects of β-Lactam antibiotics and fluoroquinolones on human gut microbiota in relation to Clostridium difficile associated diarrhea. PLoS ONE 2014, 9, e89417. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, Y.; Nakaya, R.; Chida, T.; Hashimoto, S. The effect of levofloxacin, an optically-active isomer of ofloxacin, on fecal microflora in human volunteers. Jpn. J. Antibiot. 1992, 45, 241–252. [Google Scholar]
- Hwang, I.; Park, Y.J.; Kim, Y.; Na Kim, Y.; Ka, S.; Lee, H.Y.; Seong, J.K.; Seok, Y.; Kim, J.B. Alteration of gut microbiota by vancomycin and bacitracin improves insulin resistance via glucagon-like peptide 1 in diet-induced obesity. FASEB J. 2015, 29, 2397–2411. [Google Scholar] [CrossRef] [PubMed]
- Vrieze, A.; Out, C.; Fuentes, S.; Jonker, L.; Reuling, I.; Kootte, R.S.; van Nood, E.; Holleman, F.; Knaapen, M.; Romijn, J.A.; et al. Impact of oral vancomycin on gut microbiota, bile acid metabolism, and insulin sensitivity. J. Hepatol. 2014, 60, 824–831. [Google Scholar] [CrossRef] [PubMed]
- Stewardson, A.; Gaïa, N.; François, P.; Malhotra-Kumar, S.; Delémont, C.; de Tejada, B.M.; Schrenzel, J.; Harbarth, S.; Lazarevic, V. Collateral damage from oral ciprofloxacin versus nitrofurantoin in outpatients with urinary tract infections: A culture-free analysis of gut microbiota. Clin. Microbiol. Infect. 2015, 21, 344-e1. [Google Scholar] [CrossRef] [PubMed]
- Vervoort, J.; Xavier, B.B.; Stewardson, A.; Coenen, S.; Godycki-Cwirko, M.; Adriaenssens, N.; Kowalczyk, A.; Lammens, C.; Harbarth, S.; Goossens, H.; et al. Metagenomic analysis of the impact of nitrofurantoin treatment on the human faecal microbiota. J. Antimicrob. Chemother. 2015, 70, 1989–1992. [Google Scholar] [CrossRef] [PubMed]
- Soldi, S.; Vasileiadis, S.; Uggeri, F.; Campanale, M.C.; Morelli, L.; Calanni, F.; Grimaldi, M.; Gasbarrini, A.; Fogli, M.V. Modulation of the gut microbiota composition by rifaximin in non-constipated irritable bowel syndrome patients: A molecular approach. Clin. Exp. Gastroenterol. 2015, 8, 309–325. [Google Scholar] [CrossRef]
- Ponziani, F.R.; Scaldaferri, F.; Petito, V.; Paroni Sterbini, F.; Pecere, S.; Lopetuso, L.R.; Palladini, A.; Gerardi, V.; Masucci, L.; Pompili, M.; et al. The role of antibiotics in gut microbiota modulation: The eubiotic effects of rifaximin. Dig. Dis. 2016, 34, 269–278. [Google Scholar] [CrossRef]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global cancer statistics 2020: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J. Clin. 2021. [Google Scholar] [CrossRef]
- Rajaraman, R.; Guernsey, D.L.; Rajaraman, M.M.; Rajaraman, S.R. Stem cells, senescence, neosis and self-renewal in cancer. Cancer Cell Int. 2006, 6, 25. [Google Scholar] [CrossRef]
- Li, L.T.; Jiang, G.; Chen, Q.; Zheng, J.N. Ki67 is a promising molecular target in the diagnosis of cancer (Review). Mol. Med. Rep. 2015, 11, 1566–1572. [Google Scholar] [CrossRef]
- Klein, E.Y.; Levin, S.A.; Laxminarayan, R. Reply to Abat et al.: Improved policies necessary to ensure an effective future for antibiotics. Proc. Natl. Acad. Sci. USA 2018, 115, E8111–E8112. [Google Scholar] [CrossRef]
- Xia, D.; Yang, X.; Liu, W.; Shen, F.; Pan, J.; Lin, Y.; Du, N.; Sun, Y.; Xi, X. Over-expression of CHAF1A in epithelial ovarian cancer can promote cell proliferation and inhibit cell apoptosis. Biochem. Biophys. Res. Commun. 2017, 486, 191–197. [Google Scholar] [CrossRef]
- Hernández, D.L. Letter to the editor: Use of antibiotics, gut microbiota, and risk of type 2 diabetes: Epigenetics regulation. J. Clin. Endocrinol. Metab. 2016, 101, L62–L63. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Shang, Q.; Li, W.; Guo, W.; Stojadinovic, A.; Mannion, C.; Man, Y.-G.; Chen, T. Antibiotics for cancer treatment: A double-edged sword. J. Cancer 2020, 11, 5135–5149. [Google Scholar] [CrossRef] [PubMed]
- Saeidnia, S. New Approaches to Natural Anticancer Drugs; Springer Briefs in Pharmaceutical Science & Drug Development; Springer International Publishing: Cham, Switzerland, 2015; ISBN 978-3-319-14026-1. [Google Scholar]
- Cheng, M.; Rizwan, A.; Jiang, L.; Bhujwalla, Z.M.; Glunde, K. Molecular effects of doxorubicin on choline metabolism in breast cancer. Neoplasia 2017, 19, 617–627. [Google Scholar] [CrossRef] [PubMed]
- Naci, D.; Berrazouane, S.; Barabé, F.; Aoudjit, F. Cell adhesion to collagen promotes leukemia resistance to doxorubicin by reducing DNA damage through the inhibition of Rac1 activation. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef]
- Wang, Y.; Fu, M.; Liu, J.; Yang, Y.; Yu, Y.; Li, J.; Pan, W.; Fan, L.; Li, G.; Li, X.; et al. Inhibition of tumor metastasis by targeted daunorubicin and dioscin codelivery liposomes modified with PFV for the treatment of non-small-cell lung cancer. Int. J. Nanomed. 2019, 14, 4071–4090. [Google Scholar] [CrossRef] [PubMed]
- Al-Aamri, H.M.; Ku, H.; Irving, H.R.; Tucci, J.; Meehan-Andrews, T.; Bradley, C. Time dependent response of daunorubicin on cytotoxicity, cell cycle and DNA repair in acute lymphoblastic Leukaemia. BMC Cancer 2019, 19, 1–12. [Google Scholar] [CrossRef]
- Aarts, B.M.; Klompenhouwer, E.G.; Dresen, R.C.; Deroose, C.M.A.L.O.; Beets-Tan, R.G.H.; Punie, K.; Neven, P.; Wildiers, H.; Maleux, G. Sequential intra-arterial infusion of 90Y-resin microspheres and mitomycin C in chemo refractory liver metastatic breast cancer patients: A single centre pilot study. Radiol. Oncol. 2020, 54, 33–39. [Google Scholar] [CrossRef]
- Zalipsky, S.; Saad, M.; Kiwan, R.; Ber, E.; Yu, N.; Minko, T. Antitumor activity of new liposomal prodrug of mitomycin C in multidrug resistant solid tumor: Insights of the mechanism of action. J. Drug Target. 2007, 15, 518–530. [Google Scholar] [CrossRef]
- Khasraw, M.; Bell, R.; Dang, C. Epirubicin: Is it like doxorubicin in breast cancer? A clinical review. Breast 2012, 21, 142–149. [Google Scholar] [CrossRef]
- Rao, S.; Starling, N.; Cunningham, D.; Sumpter, K.; Gilligan, D.; Ruhstaller, T.; Valladares-Ayerbes, M.; Wilke, H.; Archer, C.; Kurek, R.; et al. Matuzumab plus epirubicin, cisplatin and capecitabine (ECX) compared with epirubicin, cisplatin and capecitabine alone as first-line treatment in patients with advanced oesophago-gastric cancer: A randomised, multicentre open-label phase II study. Ann. Oncol. 2010, 21, 2213–2219. [Google Scholar] [CrossRef]
- Lopez, M.; Perno, C.-F.; Papaldo, P.; Di Lauro, L.; Ganzina, F.; Barduagni, A. Phase II study of epirubicin in advanced malignant melanoma. Investig. New Drugs 1984, 2, 315–317. [Google Scholar] [CrossRef]
- Jalalian, S.H.; Taghdisi, S.M.; Hamedani, N.S.; Kalat, S.A.M.; Lavaee, P.; ZandKarimi, M.; Ghows, N.; Jaafari, M.R.; Naghibi, S.; Danesh, N.M.; et al. Epirubicin loaded super paramagnetic iron oxide nanoparticle-aptamer bioconjugate for combined colon cancer therapy and imaging in vivo. Eur. J. Pharm. Sci. 2013, 50, 191–197. [Google Scholar] [CrossRef]
- Kristensen, G.B.; Vergote, I.; Stuart, G.; Del Campo, J.M.; Kaern, J.; Lopez, A.B.; Eisenhauer, E.; Aavall-Lundquist, E.; Ridderheim, M.; Havsteen, H.; et al. First-line treatment of ovarian cancer FIGO stages IIb-IV with paclitaxel/epirubicin/carboplatin versus paclitaxel/carboplatin. Int. J. Gynecol. Cancer 2003, 13, 172–177. [Google Scholar] [CrossRef]
- Waters, J.S.; Norman, A.; Cunningham, D.; Scarffe, J.H.; Webb, A.; Harper, P.; Joffe, J.K.; MacKean, M.; Mansi, J.; Leahy, M.; et al. Long-term survival after epirubicin, cisplatin and fluorouracil for gastric cancer: Results of a randomized trial. Br. J. Cancer 1999, 80, 269–272. [Google Scholar] [CrossRef]
- Tartakoff, S.S.; Finan, J.M.; Curtis, E.J.; Anchukaitis, H.M.; Couture, D.J.; Glazier, S. Investigations into the DNA-binding mode of doxorubicinone. Org. Biomol. Chem. 2019, 17, 1992–1998. [Google Scholar] [CrossRef]
- Burgy, O.; Wettstein, G.; Bellaye, P.S.; Decologne, N.; Racoeur, C.; Goirand, F.; Beltramo, G.; Hernandez, J.-F.; Kenani, A.; Camus, P.; et al. Deglycosylated bleomycin has the antitumor activity of bleomycin without pulmonary toxicity. Sci. Transl. Med. 2016, 8, 326ra20. [Google Scholar] [CrossRef]
- Barlow, J.J.; Piver, M.S.; Chuang, J.T.; Cortes, E.P.; Onuma, T.; Holland, J.F. Adriamycin and bleomycin, alone and in combination, in gynecologic cancers. Cancer 1973, 32, 735–743. [Google Scholar] [CrossRef]
- Beberok, A.; Wrześniok, D.; Minecka, A.; Rok, J.; Delijewski, M.; Rzepka, Z.; Respondek, M.; Buszman, E. Ciprofloxacin-mediated induction of S-phase cell cycle arrest and apoptosis in COLO829 melanoma cells. Pharmacol. Rep. 2018, 70, 6–13. [Google Scholar] [CrossRef] [PubMed]
- Beberok, A.; Wrześniok, D.; Rok, J.; Rzepka, Z.; Respondek, M.; Buszman, E. Ciprofloxacin triggers the apoptosis of human triple-negative breast cancer MDA-MB-231 cells via the p53/Bax/Bcl-2 signaling pathway. Int. J. Oncol. 2018, 52, 1727–1737. [Google Scholar] [CrossRef]
- Van Hazel, G.A.; Rubin, J.; Moertel, C.G. Treatment of metastatic carcinoid tumor with dactinomycin or dacarbazine. Cancer Treat. Rep. 1983, 67, 583–585. [Google Scholar]
- Bowman, R.V.; Manning, L.S.; Davis, M.R.; Robinson, B.W.S.; Whitaker, D. Interaction between dactinomycin and tumor necrosis factor in mesothelioma. Cachexia without oncoiysis. Cancer 1991, 67, 2495–2500. [Google Scholar] [CrossRef]
- Bloomfield, C.D.; Brunning, R.D.; Theologides, A.; Kennedy, B.J. Daunorubicin-prednisone remission induction with hy-droxyurea maintenance in acute non-lymphocytic Leukemia. Cancer 1973, 31, 931–938. [Google Scholar] [CrossRef]
- Chen, T.-C.; Hsu, Y.-L.; Tsai, Y.-C.; Chang, Y.-W.; Kuo, P.-L.; Chen, Y.-H. Gemifloxacin inhibits migration and invasion and induces mesenchymal–epithelial transition in human breast adenocarcinoma cells. J. Mol. Med. 2013, 92, 53–64. [Google Scholar] [CrossRef] [PubMed]
- Casas, E.; Kim, J.; Bendesky, A.; Ohno-Machado, L.; Wolfe, C.J.; Yang, J. Snail2 is an essential mediator of Twist1-induced epithelial mesenchymal transition and metastasis. Cancer Res. 2011, 71, 245–254. [Google Scholar] [CrossRef]
- Tomasz, M.; Palom, Y. The mitomycin bioreductive antitumor agents: Cross-linking and alkylation of DNA as the molecular basis of their activity. Pharmacol. Ther. 1997, 76, 73–87. [Google Scholar] [CrossRef]
- Volpe, A.; Racioppi, M.; D’Agostino, D.; Cappa, E.; Filianoti, A.; Bassi, P.F. Mitomycin C for the treatment of bladder cancer. Minerva Urol. Nefrol. 2010, 62, 133–144. [Google Scholar] [PubMed]
- Thürlimann, B.; Waldburger, R.; Senn, H.J.; Thiébaud, D. Original article: Plicamycin and pamidronate in symptomatic tumor-related hypercalcemia: A prospective randomized crossover trial. Ann. Oncol. 1992, 3, 619–623. [Google Scholar] [CrossRef]
- Xiao, Z.; Sperl, B.; Ullrich, A.; Knyazev, P. Metformin and salinomycin as the best combination for the eradication of NSCLC monolayer cells and their alveospheres (cancer stem cells) irrespective of EGFR, KRAS, EML4/ALK and LKB1 status. Oncotarget 2014, 5, 12877–12890. [Google Scholar] [CrossRef]
- Zhou, J.; Li, P.; Xue, X.; He, S.; Kuang, Y.; Zhao, H.; Chen, S.; Zhi, Q.; Guo, X. Salinomycin induces apoptosis in cisplatin-resistant colorectal cancer cells by accumulation of reactive oxygen species. Toxicol. Lett. 2013, 222, 139–145. [Google Scholar] [CrossRef]
- Sancheztillo, E.; Fanlo, L.; Siles, L.; Montesmoreno, S.; Moros, A.; Chivablanch, G.; Estruch, R.; Martinez, A.; Colomer, D.; Győrffy, B.; et al. The EMT activator ZEB1 promotes tumor growth and determines differential response to chemotherapy in mantle cell lymphoma. Cell Death Differ. 2014, 21, 247–257. [Google Scholar] [CrossRef]
- Rinninella, E.; Raoul, P.; Cintoni, M.; Franceschi, F.; Miggiano, G.A.D.; Gasbarrini, A.; Mele, M.C. What is the healthy gut microbiota composition? a changing ecosystem across age, environment, diet, and diseases. Microorganisms 2019, 7, 14. [Google Scholar] [CrossRef]
- Wang, T.; Cai, G.; Qiu, Y.; Fei, N.; Zhang, M.; Pang, X.; Jia, W.; Cai, S.; Zhao, L. Structural segregation of gut microbiota between colorectal cancer patients and healthy volunteers. ISME J. 2011, 6, 320–329. [Google Scholar] [CrossRef]
- Becattini, S.; Taur, Y.; Pamer, E.G. Antibiotic-induced changes in the intestinal microbiota and disease. Trends Mol. Med. 2016, 22, 458–478. [Google Scholar] [CrossRef]
- Blaser, M.J. Antibiotic use and its consequences for the normal microbiome. Science 2016, 352, 544–545. [Google Scholar] [CrossRef]
- Petrelli, F.; Ghidini, M.; Ghidini, A.; Perego, G.; Cabiddu, M.; Khakoo, S.; Oggionni, E.; Abeni, C.; Hahne, J.C.; Tomasello, G.; et al. Use of antibiotics and risk of cancer: A systematic review and meta-analysis of observational studies. Cancers 2019, 11, 1174. [Google Scholar] [CrossRef] [PubMed]
- Mao, Q.; Jiang, F.; Yin, R.; Wang, J.; Xia, W.; Dong, G.; Ma, W.; Yang, Y.; Xu, L.; Hu, J. Interplay between the lung microbiome and lung cancer. Cancer Lett. 2018, 415, 40–48. [Google Scholar] [CrossRef]
- Whiteside, S.A.; Razvi, H.; Dave, S.; Reid, G.; Burton, J.P. The microbiome of the urinary tract—A role beyond infection. Nat. Rev. Urol. 2015, 12, 81–90. [Google Scholar] [CrossRef]
- Yamamoto, M.L.; Schiestl, R.H. Intestinal microbiome and lymphoma development. Cancer J. 2014, 20, 190–194. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Class | Target Site | Representatives | |
|---|---|---|---|
| β-lactams | Penicillins | Cell wall synthesis | Penicillin G and Penicillin V, Methicillin, Oxacillin, Cloxacillin, Dicloxacillin, Nafcillin, Ampicillin, Amoxicillin, Carbenicillin, Ticarcillin, Mezlocillin, Piperacillin, Azlocillin, Temocillin |
| Cephalosporins | Cell wall synthesis | 1st generation: Cephalothin, Cephapirin, Cephradine, Cephaloridine, Cefazolin 2nd generation: Cefamandole, Cefuroxime, Cephalexin, Cefprozil, Cefaclor, Loracarbef, Cefoxitin, Cefmetazole 3rd generation: Cefotaxime, Ceftizoxime, Ceftriaxone, Cefoperazone, Ceftazidime, Cefixime, Cefpodoxime, Ceftibuten, Cefdinir 4th generation: Cefpirome, Cefepime 5th generation: Ceftaroline, Ceftobiprole | |
| Carbapenems | Cell wall synthesis | Imipenem, Meropenem, Doripenem | |
| Monobactams | Cell wall synthesis | Aztreonam | |
| Macrolides | Protein synthesis inhibitors—Inhibit 50 s subunit | Erythromycin, Azithromycin, Clarithromycin | |
| Tetracyclines | Protein synthesis inhibitors—Inhibit 30 s subunit | Tetracycline, Chlortetracycline, Oxytetracycline, Demeclocycline, Minocycline, Methacycline, Doxycycline, Tigecycline | |
| Aminoglycosides | Protein synthesis inhibitors—Inhibit 30 s subunit | Streptomycin, Neomycin, Kanamycin, Paromomycin, Gentamicin, Tobramycin, Amikacin, Netilmicin, Spectinomycin, Sisomicin, Isepamicin | |
| Sulfonamides | Folic acid synthesis inhibitors | Prontosil, Sulfonamide, Sulfanilamid, Para-Aminobenzoic Acid, Sulfadiazine, Sulfisoxazole, Sulfamethoxazole, Sulfathalidine | |
| Quinolones | DNA synthesis inhibitors | Nalidixic Acid, Ciprofloxacin, Norfloxacin, Pefloxacin, Enoxacin, Ofloxacin, Levofloxacin, Sparfloxacin, Lomefloxacin, Fleroxacin | |
| Izoniazid | Mycolic acid synthesis inhibitors | ||
| Ansamycin | RNA synthesis inhibitors | Rifampin | |
| Polymycins | Cytoplasmic membrane structure | ||
| Daptomycin | Cytoplasmic membrane structure | ||
| Scientific Name (Common Name) | Chemical Constituents | Spectrum of Action | References |
|---|---|---|---|
| Aleurites moluccanus (Candlenut) | 3-acetyl aleuritolic acid, moluccanin | C. albicans, S. aureus, E. faecalis | [161] |
| Aloe barbadensis (Aloe vera) | Reducing sugar, tannins, anthraquinone, glycoside, saponins, flavonoids | S. aureus MRSA., K. pneumonia | [162] |
| Azadirachta indica (Neem) | Reducing sugar, tannins, glycoside, saponins, flavonoids, steroids, terpenoids | S. aureus, E. coli | [162] |
| Bergenia ciliata | Tannins, terpenoids, alkaloids, flavonoids | S. aureus, B. subtilis, P. vulgaris, P. aeruginosa, E. coli | [163] |
| Buchenavia Tomentosa (Merindiba) | Ellagic acid, gallic acid, myricetin-3-O-rhamnopyranoside, quinic acid | E. coli, P. aeruginosa, S. aureus, S. epidemidis | [164] |
| Bryophyllum pinnatum (Bryophyllum) | reducing sugar, tannins, glycoside, saponins, flavonoids, steroids, terpenoids | E. coli, S. aureus, P. mirabilis | [162] |
| Cannabis sativa | α-Pinene, myrcene, linalool, limonene, trans-β-ocimene, α-terpinolene, trans-caryophyllene, α-humulene, caryophyllene | B. cereus, E. coli, B. subtilis, S. typhi | [165] |
| Caryophyllus aromaticus (Clove) | Essential oils (eugenol), tannins and flavonoids | S. aureus, S. choleraesuis, P. aeruginosa, C. albicans, K. pneumoniae, K. pneumonia, Shigella spp., Proteus spp., P. aeruginosa | [166] |
| Crataegus azarolus var. aronia calli (Hawthorn) | Pro-anthocyanidins, flavonoid, (−)-epicatechin, procyanidin B2, chlorogenic acid and hyperoside | Staphylococcus aureus, S. epidermidis, M. luteus | [167] |
| Croton doctoris | Malic acid, scopoletin, ferulic acid, ent-18-hydroxy-trachyloban-3-one, crotonin, cajucarin B | A. naeslundii , L.acidophilus, S. gordonii, S. mutans, S. sanguinis, S. sobrinus, S. mitis | [168] |
| Croton urucurana (Baillon) | Borneol, bornyl acetate, 1-isopropyl-7-methyl-4-methylene-1,3,4,5,6,8-hexahydro-2H-naphthalen-4a-ol, sesquicineole g-gurjunene epoxide | S. aureus, S. epidermidis, P. aeruginosa, E. coli, S. setubal, K. pneumoniae, S. cerevisiae, C. neoformans | [169] |
| Curcuma longa (Turmeric) | Carbohydrates, saponins, flavonoids, tannins | S. pyogenes, S. aureus, P. aeruginosa | [170] |
| Commiphora molmol (Myrrh) | Carbohydrates, alkaloids, saponins | S. pyogenes, S. aureus, P. aeruginosa | [170] |
| Cymbopogon citratus (Lemongrass) | Tannins, glycoside, saponins, flavonoids, terpenoids | S. aureus MRSA., E. coli, K. pneumoniae | [162] |
| Dodonaea viscosa (Hopbush) | Terpenes, phenols, flavonoids, saponins | B. cereus, E.coli, S. typhi | [165] |
| Euphorbia helioscopia (Sun spurge) | Alkaloids, tannins, flavonoids, steroids | E. coli, B. subtilis, T. harzianum, S. typhi, K. pneumonia, S. aureus, B. subtilis | [171] |
| Jasminum officinale (Jasmine) | Tannins, terpenoids, alkaloids, flavonoids | S. aureus, B. subtilis, P. aeruginosa, E.coli | [163] |
| Melissa offficinalis (Lemon-balm) | Essential oils (containing citral and citronellal monoterpenes), flavonoids and rosmarinic, caffeic and chlorogenic acids | S. aureus, S. choleraesuis, K. pneumoniae | [166] |
| Ocimun basilicum (Basil) | Essential oils (linalool, estragol and eugenol), tannins and flavonoids | P. aeruginosa | [166] |
| Ocimum sanctum (Tulsi) | Tannins, glycoside, saponins, flavonoids, steroids, terpenoids | S. aureus MRSA., E. coli | [166] |
| Orthosiphon aristatus (Blume) | Diterpenoids, polymethoxylated flavonoids, caffeic acid derivatives | C. albicans, S. aureus, E. faecalis, P. aeruginosa | [161] |
| Origanum vulgare (Oregano) | Reducing sugar, tannins, glycoside, saponins | S. aureus MRSA., Bacillus amyloliquefaciens, B. brevis, B. subtilis, S. aureus, C. xerosis, E. coli, K. pneumoniae, P. vulgaris, M. smegmatis | [162] |
| Pimpinella anisum (Anise) | Carbohydrates, saponins, flavonoids | E.coli, P. aeruginosa | [170] |
| Psidium guajava (Guava) | comarins, essential oils, flavonoids, triterpenes and ellagitannins | S. aureus, C. albicans | [166] |
| Punica granatum (Pomegranate) | Ellagitannins and alkaloids | P. aeruginosa, B. subtilis | [166] |
| Rosmarinus officinalis (Rosemary) | Flavonoids, phenolic acids (caffeic, chorogenic and rosmarinic) and essential oils (camphor and cineole) and diterpenes | B. subtilis, C. albicans, S. aureus MRSA., E. coli, P. mirabilis, K. pneumoniae, P. aeruginosa | [162,166] |
| Santalum album (Indian sandalwood) | Tannins, terpenoids, alkaloids, flavonoids | S. aureus, B. subtilis, P. vulgaris, E. coli | [163] |
| Solanum nigrum (Black nightshade) | alkaloids, cardiac glycosides, flavonoids, saponins, tannins, volatile oils | B. cereus, E. coli, P. aeruginosa, S. typhi | [165] |
| Syzygyum joabolanum (Jambolan) | Flavonoids and tannins | S. aureus, C. albicans, Proteus spp., K. pneumonia, Proteus spp., P. aeruginosa, E. aerogenes, S. aureus | [166] |
| Thymus vulgaris (Thyme) | Essential oils (mainly thymol and carvacrol), flavonoids, tannins and triterpenes | S. aureus, C. albicans, Proteus spp., P. aeruginosa, E. coli, S. aureus MRSA., P. mirabilis, K. pneumoniae | [162,166] |
| Vitis vinifera (Grape) | Gallic acid | E. coli, S.s aureus | [172] |
| Woodfordia floribunda (Fire flame) | Diethyl phthalate, thymol, woodfordin C | C. albicans, S. aureus, E. faecalis | [161] |
| Zingiber officinale (Ginger) | Carbohydrates, alkaloids, steroids, saponins | S. pyogenes, S. aureus, P. aeruginosa | [170] |
| Antibiotic Representative | Effects on Intestinal Microbiota | Ref. |
|---|---|---|
| Clindamycin | ↓ Bacteroides diversity | [184] |
| ↓ Gram-positive aerobic and anaerobic bacteria | [185] | |
| Clarithromycin | ↓ Actinobacteria | [186,187] |
| ↓ Firmicutes ↑ Proteobacteria ↑ Bacteroidetes | [187] | |
| Erythromycin | ↓ Actinobacteria ↓ Firmicutes ↑ Proteobacteria | [188] |
| Penicillin Ampicillin | ↓ Actinobacteria ↓ Firmicutes ↑ Proteobacteria | [188,189] |
| ↑ Bacteroidetes | [189] | |
| Cephalosporins | ↓ Firmicutes ↑ Bacteroidetes ↑ Proteobacteria | [189] |
| Fluoroquinolones Ciprofloxacin, Levofloxacin | ↑ Proteobacteria ↑ Bacteroidetes | [189] |
| ↓Gram-negative facultative anaerobes ↓Gram-positive anaerobes ↓ Bifidobacteria | [190] | |
| Vancomycin | ↓Total bacterial diversity ↓ Firmicutes ↓ Bacteroidetes | [191,192] |
| ↑ Proteobacteria | [192] | |
| Bacitracin | ↓ Firmicutes ↓ Bacteroidetes ↓Total bacterial diversity | [191] |
| Nitrofurantoin | ↑ Actinobacteria ↑ Bifidobacteria ↑ Fecalibacterium genus | [193,194] |
| Rifaximin | ↑ Bacterial diversity ↑ Firmicutes/Bacteroidetes ratio | [195] |
| ↑ abundance of Lactobacilli | [196] |
| Antibiotics | Type of Cancer | Mechanism of Action | Ref |
|---|---|---|---|
| Adriamycin | Malignant lymphoma, breast cancer, lung cancer | - Inhibition of the activity of topoisomerase I and II causing the breakdown of DNA with two strands - Upregulation of p53 - Alteration of the Bcl-2/Bax pathway (changing the Bcl-2/Bax ratio) | [217] |
| Bleomycin | Lymphomas, germ cell tumors, melanomas, gynecologic cancers | - DNA binding - Production of free radicals (hydroxyl radicals) | [218,219] |
| Ciprofloxacin | Bladder cancer, colorectal cancer, hepatocellular carcinoma, melanoma, breast cancer | - Inhibition of the activity of topoisomerase II - Impairment of cell cycle at the G2 / M checkpoint | [220,221] |
| Dactinomycin | Nephroblastoma, chorionic epithelial carcinoma, rhabdomyosarcoma and neuroblastoma | - Specific interaction with deoxyguanine in DNA - Formation of complexes with DNA - Inhibition of the mRNA synthesis | [222,223] |
| Daunorubicin | Acute myelogenous leukemia, lymphocytic leukemia | - Chimerism between basic DNA pairs - Tight DNA binding - Obstruction of the spatial structure of DNA | [224] |
| Epirubicin | Breast cancer, malignant lymphoma, soft tissue sarcoma, gastric cancer, malignant melanoma, colon cancer, lung cancer, ovarian cancer | - Direct incorporation between DNA nucleotide pairs - Interfering with the transcription process and inhibiting mRNA formation - Topoisomerase II inhibition | [216] |
| Gemifloxacin | Breast cancer | - Inhibition of bacterial DNA gyrase and topoisomerase IV - Suppression of activity of NF-kB - Inhibition of the migration and invasion induced by cancer necrosis factor (TNF-α). | [225,226] |
| Mitomycin | Bladder cancer | - Formation of double cross-links or intrachain with DNA - Inhibition of DNA replication and synthesis - Increase in oxygen radicals | [227,228] |
| Plicamycin | Testicular embryonal cancer, glioma, lymphoma | - DNA binding - Inhibition of RNA synthesis - Acting on cell proliferation at all stages | [229] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pancu, D.F.; Scurtu, A.; Macasoi, I.G.; Marti, D.; Mioc, M.; Soica, C.; Coricovac, D.; Horhat, D.; Poenaru, M.; Dehelean, C. Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity—A Pharmaco-Toxicological Screening. Antibiotics 2021, 10, 401. https://doi.org/10.3390/antibiotics10040401
Pancu DF, Scurtu A, Macasoi IG, Marti D, Mioc M, Soica C, Coricovac D, Horhat D, Poenaru M, Dehelean C. Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity—A Pharmaco-Toxicological Screening. Antibiotics. 2021; 10(4):401. https://doi.org/10.3390/antibiotics10040401
Chicago/Turabian StylePancu, Daniel Florin, Alexandra Scurtu, Ioana Gabriela Macasoi, Daniela Marti, Marius Mioc, Codruta Soica, Dorina Coricovac, Delia Horhat, Marioara Poenaru, and Cristina Dehelean. 2021. "Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity—A Pharmaco-Toxicological Screening" Antibiotics 10, no. 4: 401. https://doi.org/10.3390/antibiotics10040401
APA StylePancu, D. F., Scurtu, A., Macasoi, I. G., Marti, D., Mioc, M., Soica, C., Coricovac, D., Horhat, D., Poenaru, M., & Dehelean, C. (2021). Antibiotics: Conventional Therapy and Natural Compounds with Antibacterial Activity—A Pharmaco-Toxicological Screening. Antibiotics, 10(4), 401. https://doi.org/10.3390/antibiotics10040401