
Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts

 ,
,  ,
, 
Abstract
1. Introduction
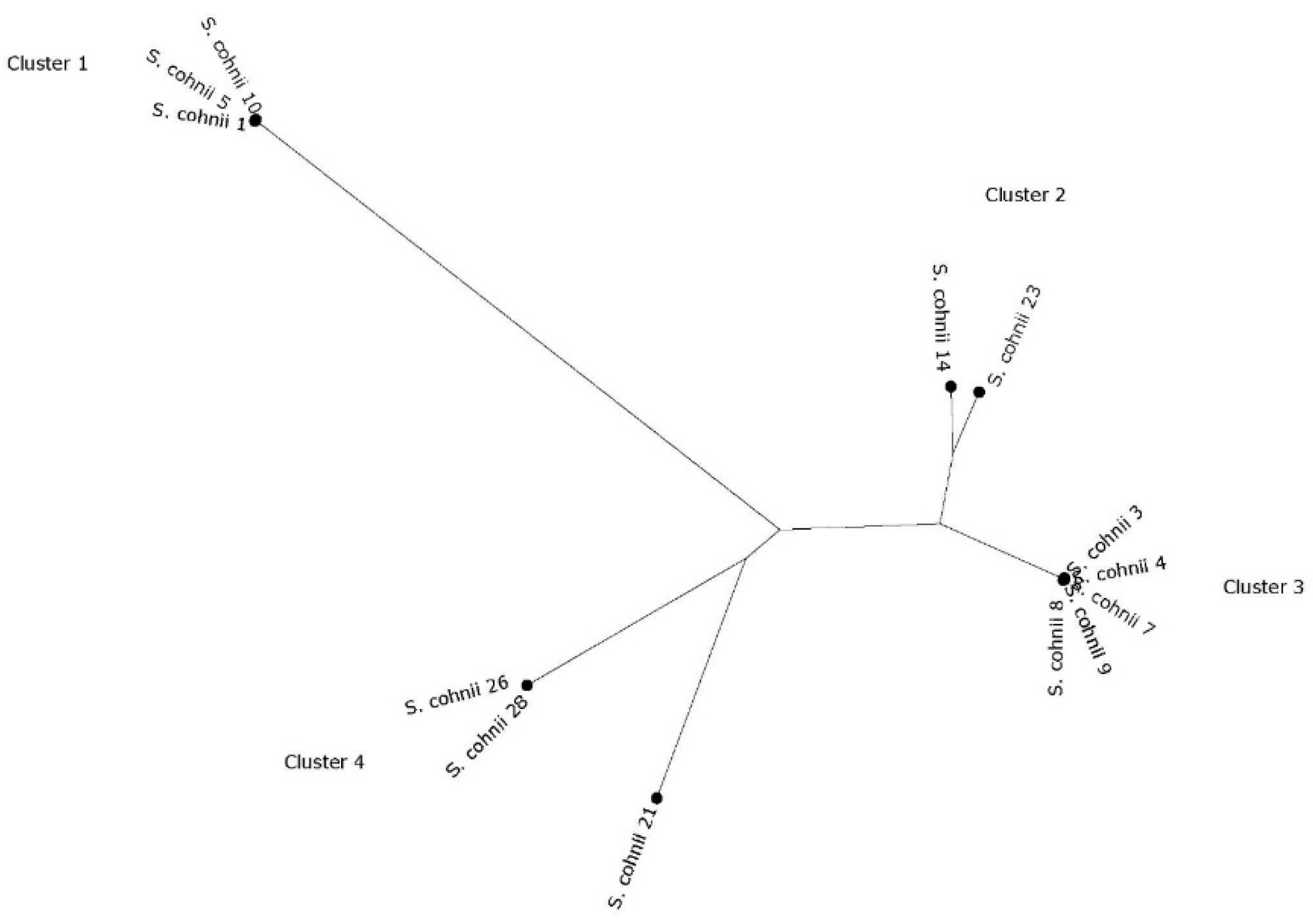
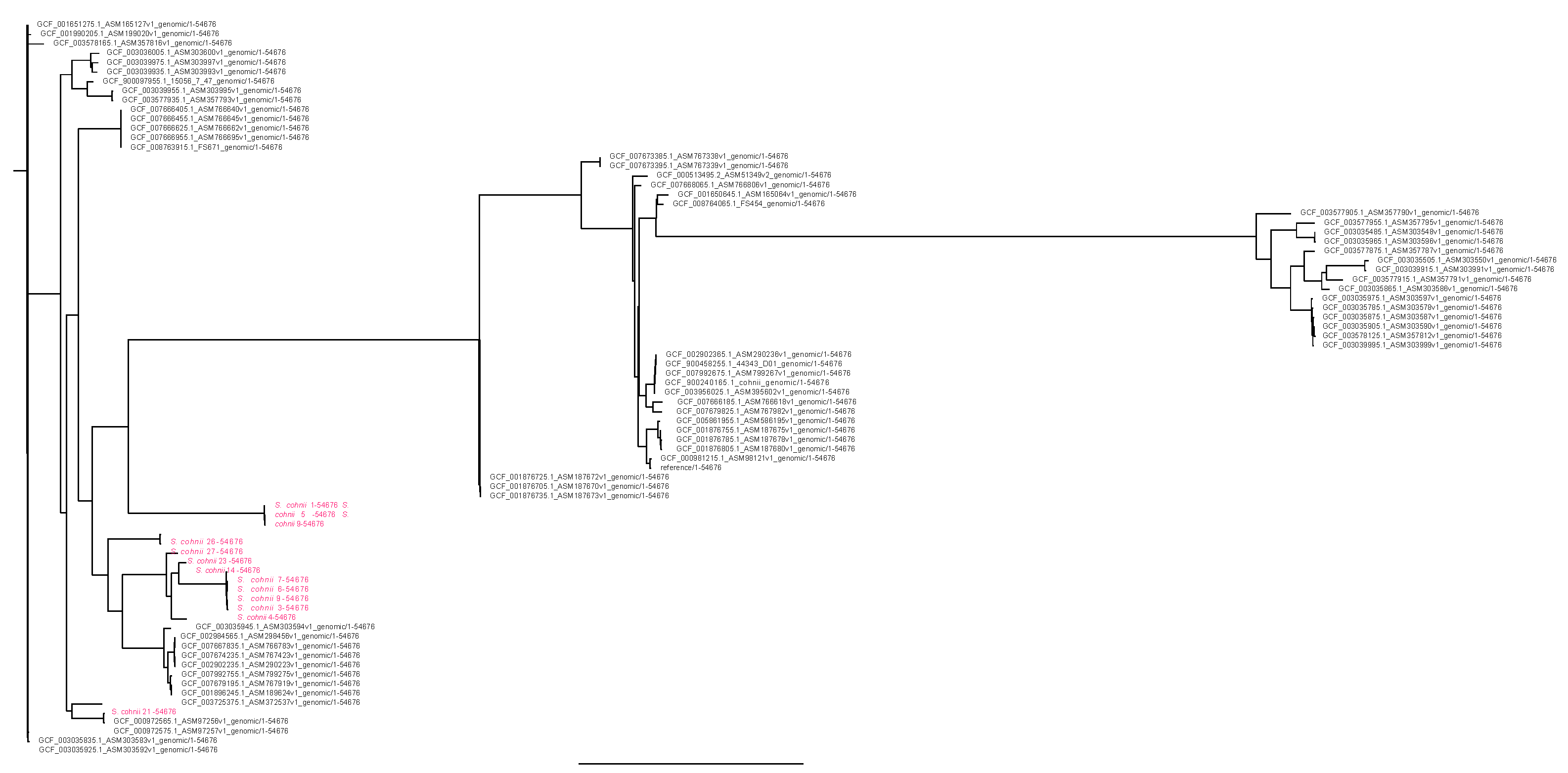
2. Results
3. Discussion
4. Materials and Methods
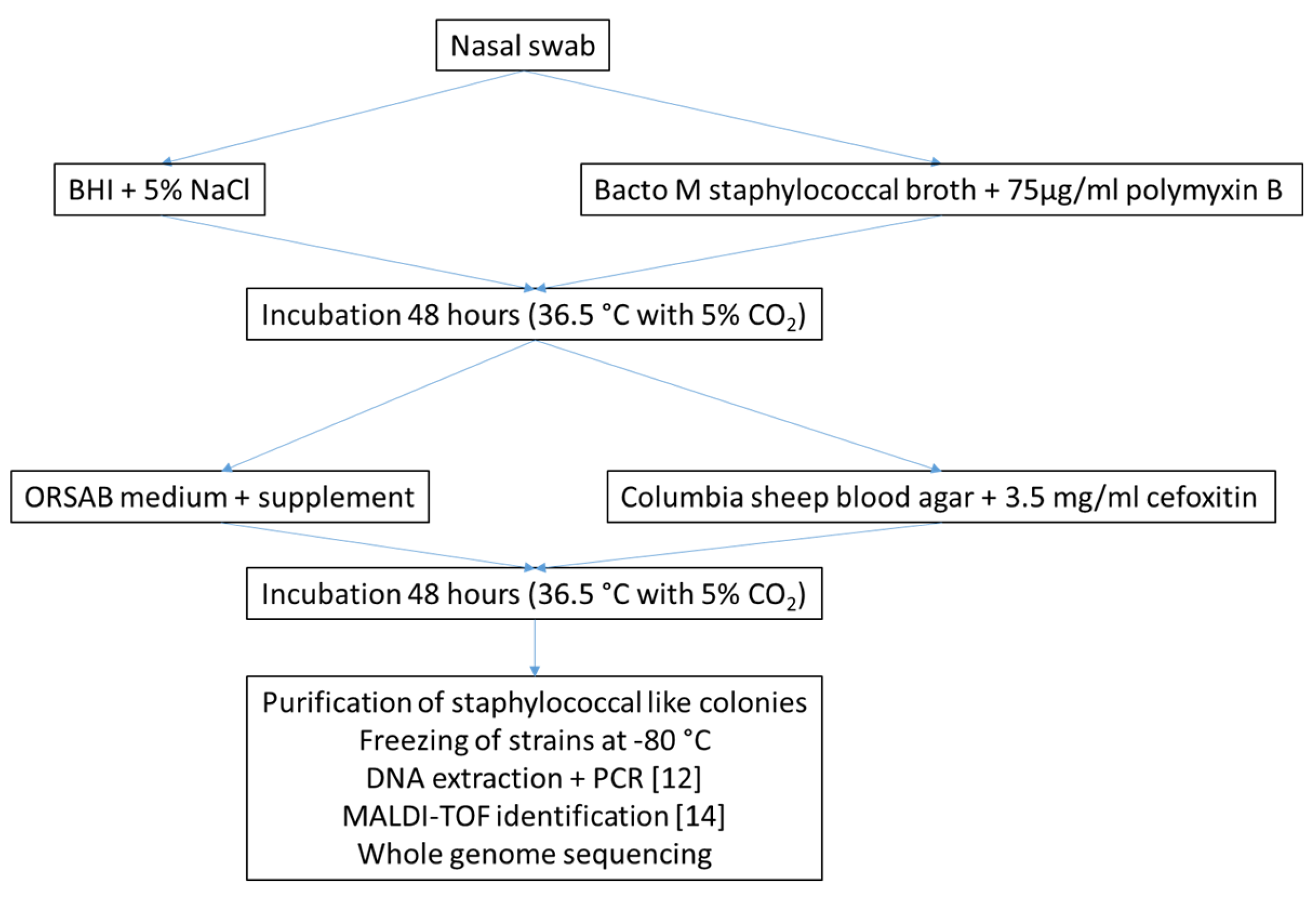
4.1. Sampling
4.2. Isolation of Methicillin-Resistant Staphylococci
4.3. Bacterial Identification
4.4. Whole-Genome Sequencing
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Butaye, P.; Argudín, M.A.; Smith, T.C. Livestock-Associated MRSA and its current evolution. Curr. Clin. Microbiol. Rep. 2016, 3, 19–31. [Google Scholar] [CrossRef]
- Roberts, M.C.; Joshi, P.R.; Greninger, A.L.; Melendez, D.; Paudel, S.; Acharya, M.; Bimali, N.K.; Koju, N.P.; No, D.; Chalise, M.; et al. The human clone ST22 SCCmec IV methicillin-resistant Staphylococcus aureus isolated from swine herds and wild primates in Nepal: Is man the common source? FEMS Microbiol. Ecol. 2018, 94, fiy052. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C.; Joshi, P.R.; Monecke, S.; Ehricht, R.; Müller, E.; Gawlik, D.; Diezel, C.; Braun, S.D.; Praudel, S.; Acharya, M.; et al. Staphylococcus aureus and Methicillin Resistant S. aureus in Nepalese Primates; Resistance to Antimicrobials, Virulence and Genetic Lineages. Antibiotics 2020, 9, 689. [Google Scholar] [CrossRef] [PubMed]
- Pires Dos Santos, T.; Damborg, P.; Moodley, A.; Guardabassi, L. Systematic review on global epidemiology of methicillin-resistant Staphylococcus pseudintermedius: Inference of population structure from multilocus sequence typing data. Front. Microbiol 2016, 7, 1599. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Vanderhaeghen, W.; Butaye, P. Diversity of antimicrobial resistance and virulence genes in methicillin-resistant non-Staphylococcus aureus staphylococci from veal calves. Res. Vet. Sci. 2015, 99, 10–16. [Google Scholar] [CrossRef]
- Vanderhaeghen, W.; Vandendriessche, S.; Crombé, F.; Nemeghaire, S.; Dispas, M.; Denis, O.; Hermans, K.; Haesebrouck, F.; Butaye, P. Characterization of methicillin-resistant non-Staphylococcus aureus staphylococci carriage isolates from different bovine populations. J. Antimicrob. Chemother. 2013, 68, 300–307. [Google Scholar] [CrossRef] [PubMed]
- Miragaia, M. Factors contributing to the evolution of mecA-mediated β-lactam resistance in Staphylococci: Update and new Insights from Whole Genome Sequencing (WGS). Front. Microbiol. 2018, 9, 2723. [Google Scholar] [CrossRef]
- Bittar, F.; Keita, M.B.; Lagier, J.C.; Peeters, M.; Delaporte, E.; Raoult, D. Gorilla gorilla gorilla gut: A potential reservoir of pathogenic bacteria as revealed using culturomics and molecular tools. Sci. Rep. 2014, 4, 7174. [Google Scholar] [CrossRef] [PubMed]
- Liang, D.; Alvarado, T.P.; Oral, D.; Vargas, J.M.; Denena, M.M.; McCulley, J.P. Ophthalmic examination of the captive western lowland gorilla (Gorilla gorilla gorilla). J. Zoo Wildl. Med. 2005, 36, 430–433. [Google Scholar] [CrossRef]
- Donato, A.C.J.; Penna, B.; Consalter, A.; Carvalho, D.D.; Lilenbaum, W.; Ferreira, A.M.R. Antibiotic resistance in Staphylococcus sp. isolated from the vaginal environment of squirrel monkeys (Saimiri spp.) bred ex situ. J. Med. Primatol. 2017, 46, 75–78. [Google Scholar] [CrossRef]
- Pantůček, R.; Sedláček, I.; Petráš, P.; Koukalová, D.; Švec, P.; Štětina, V.; Vancanneyt, M.; Chrastinová, L.; Vokurková, J.; Růžičková, V.; et al. Staphylococcus simiae sp. nov., isolated from South American squirrel monkeys. Int. J. Syst. Evol. Microbiol. 2005, 55, 1953–1958. [Google Scholar] [CrossRef]
- Carvalho, V.M.; Vanstreels, R.E.; Paula, C.D.; Kolesnikovas, C.K.; Ramos, M.C.; Coutinho, S.D.; Martins, C.S.; Pissinatti, A.; Catão-Dias, J.L. Nasal, oral and rectal microbiota of Black lion tamarins (Leontopithecus chrysopygus). Braz. J. Microbiol. 2015, 45, 1531–1539.D. [Google Scholar] [CrossRef]
- Lilenbaum, W.; Moraes, I.A.; Cardoso, V.S.; Varges, R.G.; Ferreira, A.M.; Pissinatti, A. Antibiotic resistance in Staphylococci isolated from the vaginas of captive female Leontopithecus (Callitrichidae-Primates). Am. J. Primatol. 2006, 68, 825–831. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C.; Feßler, A.T.; Monecke, S.; Ehricht, R.; No, D.; Schwarz, S. Molecular analysis of two different MRSA clones ST188 and ST2368 from primates (Macaca spp.) in a United States primate center. Front. Microbiol. 2018, 9, 2199. [Google Scholar] [CrossRef] [PubMed]
- Soge, O.O.; No, D.; Michael, K.; Dankoff, J.; Lane, J.; Vogel, K.; Smedley, J.; Roberts, M.C. Transmission of MDR MRSA between primates personnel and environment at a United States primate center. J. Antimicrob. Chemother. 2016, 71, 2798–2803. [Google Scholar] [CrossRef] [PubMed]
- Roberts, M.C.; Joshi, P.R.; Monecke, S.; Ehricht, R.; Muller, E.; Gawlik, D.; Paudel, S.; Acharya, M.; Bhattarai, S.; Pokharel, S.; et al. MRSA strains among Nepalese Rhesus macaques (Macaca mulatta) and their environment. Front. Microbiol. 2019, 10, 2505. [Google Scholar] [CrossRef]
- Heaton, C.J.; Gerbig, G.R.; Sensius, L.D.; Patel, V.; Smith, T.C. Staphylococcus aureus epidemiology in wildlife: A systematic review. Antibiotics 2020, 9, 89. [Google Scholar] [CrossRef] [PubMed]
- Guardabassi, L.; Moodley, A.; Williams, A.; Stegger, M.; Damborg, P.; Halliday-Simmonds, I.; Butaye, P. High prevalence of USA300 among clinical isolates of methicillin-resistant Staphylococcus aureus on St. Kitts and Nevis, West Indies. Front. Microbiol. 2019, 10, 1123. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Ripa, L.; Feßler, A.T.; Hanke, D.; Sanz, S.; Olarte, C.; Mama, O.M.; Eichhorn, I.; Schwarz, S.; Torres, C. Coagulase-negative staphylococci carrying cfr and PVL genes, and MRSA/MSSA-CC398 in the swine farm environment. Vet. Microbiol. 2020, 43, 108631. [Google Scholar] [CrossRef]
- Heilmann, C.; Ziebuhr, W.; Becker, K. Are coagulase-negative staphylococci virulent? Clin. Microbiol. Infect. 2019, 25, 1071–1080. [Google Scholar] [CrossRef]
- Wuytack, A.; De Visscher, A.; Piepers, S.; Boyen, F.; Haesebrouck, F.; De Vliegher, S. Non-aureus staphylococci in fecal samples of dairy cows: First report and phenotypic and genotypic characterization. J. Dairy Sci. 2019, 102, 9345–9359. [Google Scholar] [CrossRef]
- Loncaric, I.; Tichy, A.; Handler, S.; Szostak, M.P.; Tickert, M.; Diab-Elschahawi, M.; Spergser, J.; Künzel, F. Prevalence of Methicillin-Resistant Staphylococcus sp. (MRS) in different companion animals and determination of risk factors for colonization with MRS. Antibiotics 2019, 8, 36. [Google Scholar] [CrossRef]
- Bean, D.C.; Wigmore, S.M.; Wareham, D.W. Draft Genome Sequence of Staphylococcus cohnii subsp. urealyticus Isolated from a healthy dog. Genome Announc. 2017, 5, e01628-16. [Google Scholar] [PubMed]
- Argudín, M.A.; Vanderhaeghen, W.; Vandendriessche, S.; Vandecandelaere, I.; André, F.X.; Denis, O.; Coenye, T.; Butaye, P. Antimicrobial resistance and population structure of Staphylococcus epidermidis recovered from animals and humans. Vet. Microbiol. 2015, 178, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.N. The ABC of ribosome-related antibiotic resistance. mBio 2016, 7, e00598-16. [Google Scholar] [CrossRef] [PubMed]
- Sharkey, L.K.R.; Edwards, T.A.; O’Neil, A.J. ABC-F Proteins Mediate Antibiotic Resistance through Ribosomal Protection. mBio 2016, 7, e01975. [Google Scholar] [CrossRef]
- Partridge, S.R.; Kwong, S.M.; Firth, N.; Jensen, S.O. Mobile genetic elements associated with antimicrobial resistance. Clin. Microbiol. Rev. 2018, 31, e00088-17. [Google Scholar] [CrossRef]
- Vanderhaeghen, W.; Vandendriessche, S.; Crombé, F.; Dispas, M.; Denis, O.; Hermans, K.; Haesebrouck, F.; Butaye, P. Species and staphylococcal cassette chromosome mec (SCCmec) diversity among methicillin-resistant non-Staphylococcus aureus staphylococci isolated from pigs. Vet. Microbiol. 2012, 158, 123–128. [Google Scholar] [CrossRef]
- Maes, N.; Magdalena, J.; Rottiers, S.; De Gheldre, Y.; Struelens, M.J. Evaluation of a triplex PCR assay to discriminate Staphylococcus aureus from coagulase-negative Staphylococci and determine methicillin resistance from blood cultures. J. Clin. Microbiol. 2002, 40, 1514–1517. [Google Scholar] [CrossRef] [PubMed]
- Cameron, M.; Perry, J.; Middleton, J.R.; Chaffer, M.; Lewis, J.; Keefe, G.P. Evaluation of MALDI-TOF mass spectrometry and a custom reference spectra expanded database for the identification of bovine-associated coagulase-negative staphylococci. J. Dairy Sci. 2018, 101, 590–595. [Google Scholar] [CrossRef] [PubMed]
- Argudín, M.A.; Vanderhaeghen, W.; Vandendriessche, S.; Vandecandelaere, I.; Denis, O.; Coenye, T.; Butaye, P. Biofilm formation of ica operon-positive Staphylococcus epidermidis from different sources. APMIS 2015, 123, 1081–1089. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Sample | Origin | mecA | Species | Resistance Genes *** | SCCmec Elements | Plasmids |
|---|---|---|---|---|---|---|
| 1 ** | Captive | + | S. cohnii | blaZ, mph(C) | SCCmecIII(3A) | rep7a rep(pSBK203) U35036 |
| 2 ** | Captive | + | S. epidermidis | fosB, dfrG, erm(C) | SCCmecV(5C2) | rep10 repL(pDLK1) GU562624, rep39 repA(SAP110A) GQ900465 |
| 3 ** | Captive | - | S. cohnii | blaZ, tet(K) | ccrA1:4:LGA251:FR821779 subtype-Vc(5C2&5):10:AB505629 | rep7a repC(pS0385p1) AM990993 rep21 rep(pSHaeA) AP006717 |
| 4 ** | Captive | - | S. cohnii | blaZ, tet(K) | ccrA1:4:LGA251:FR821779 subtype-Vc(5C2&5):10:AB505629 | rep7a repC(pS0385p1) AM990993/ repC(Cassette) AB037671 |
| 5 ** | Captive | + | S. cohnii | blaZ, mph(C) | rep7a rep(pSBK203) U35036 | |
| 6 | Captive | - | S. pettenkoferi | none | ||
| 7 ** | Captive | - | S. cohnii | blaZ, tet(K) | ccrA1:4:LGA251:FR821779 subtype-Vc(5C2&5):10:AB505629 | rep7a repC(pS0385p1) AM990993/repC(Cassette) AB037671/ORF(pKH1) SAU38656 |
| 8 ** | Captive | - | S. cohnii | blaZ | ccrA1:4:LGA251:FR821779 subtype-Vc(5C2&5):10:AB505629 | rep21 rep(pKH21) EU350088 |
| 9 ** | Captive | - | S. cohnii | blaZ, tet(K) | ccrA1:4:LGA251:FR821779 subtype-Vc(5C2&5):10:AB505629 | rep7a repC(pS0385p1) AM990993 |
| 10 ** | Captive | + | S. cohnii | blaZ, mph(C) | none | |
| 11 ** | Captive | - | S. warneri * | none | repUS35 A28412072(pvSw2) CP003671 | |
| 12 | Wild | - | S. kloosii | none | ||
| 13 | Wild | - | S. saprophyticus | none | ||
| 14 ** | Wild | - | S. cohnii | none | ccrB3:2:LGA251:FR821779 | rep7a rep(pSBK203) U35036 rep21 rep(pKH21) EU350088 |
| 15 | Wild | - | S. xylosus | none | ||
| 16 ** | Wild | + | S. arlettae | str | rep13 ORF(pC194) V01277 | |
| 17 | Wild | - | S. aureus | none | ||
| 18 ** | Wild | + | S. arlettae | none | none | |
| 19 ** | Wild | + | S. epidermidis | fosB, dfrG | SCCmecV(5C2) | rep10 repL(pDLK1) GU562624 rep39 repA(SAP110A) GQ900465 |
| 20 | Wild | - | S. aureus | |||
| 21 ** | Wild | - | S. cohnii | none | ccrA1:4:LGA251:FR821779 subtype-Vc(5C2&5):10:AB505629 | rep21 rep(pSHaeA) AP006717 |
| 22 | Wild | - | S. xylosus | |||
| 23 ** | Wild | - | S. cohnii | blaZ, tet(K), cat(pC211) | rep7a rep(pSBK203) U35036/repD(pTZ4) NC010111/ORF(pKH1) SAU38656 | |
| 24 | Wild | - | S. cohnii | |||
| 25 ** | Wild | + | S. hominis | blaZ, msr(A) | rep7a repA(SAP105B) GQ900453 | |
| 26 ** | Captive | - | S. cohnii | none | ccrA1:3:JCSC6943:AB505628 | none |
| 27 ** | Captive | - | S. cohnii * | none | none | |
| 28 | Wild | NA | Negative | |||
| 29 | Wild | NA | Negative | |||
| 30 | Wild | NA | Negative | |||
| 31 | Wild | - | S. hominis | |||
| 32 | Wild | NA | Negative | |||
| 33 | Wild | NA | Negative | |||
| 34 | Wild | - | S. cohnii |
| Sample | Number of Contigs | Mean Contig Length | Median Contig Length | Minimum Contig Length | Maximum Contig Length | Base Count | N50 | Coverage |
|---|---|---|---|---|---|---|---|---|
| 1 | 539 | 4548.83 | 2656 | 137 | 27,277 | 2,451,819 | 9591 | 29 |
| 2 | 469 | 196.37 | 2725 | 137 | 46,151 | 2,210,175 | 9380 | 31 |
| 3 | 514 | 4917.82 | 2570 | 137 | 35,832 | 2,527,758 | 10,396 | 29 |
| 4 | 676 | 152.46 | 2033 | 137 | 30,607 | 2,474,092 | 7349 | 27 |
| 5 | 524 | 187.09 | 2386 | 137 | 54,340 | 2,352,829 | 9277 | 27 |
| 7 | 554 | 4463.89 | 2638 | 137 | 37,050 | 2,472,996 | 8527 | 25 |
| 8 | 674 | 153.83 | 2184 | 137 | 38,142 | 2,488,125 | 7397 | 24 |
| 9 | 614 | 166.17 | 2257 | 137 | 43,634 | 2,448,661 | 8110 | 24 |
| 10 | 433 | 5385.63 | 3060 | 137 | 39,146 | 2,331,979 | 12,088 | 42 |
| 11 | 397 | 248.19 | 2854 | 137 | 44,913 | 2,364,664 | 12,992 | 35 |
| 14 | 543 | 4377.91 | 2462 | 137 | 37,869 | 2,377,204 | 8798 | 38 |
| 16 | 814 | 121.20 | 1738 | 137 | 24,999 | 2,367,457 | 5316 | 48 |
| 18 | 427 | 231.74 | 2963 | 137 | 55,529 | 2,374,768 | 11,863 | 39 |
| 19 | 375 | 6025.86 | 3340 | 137 | 38,343 | 2,259,697 | 12,172 | 40 |
| 21 | 435 | 238.79 | 2540 | 137 | 52,277 | 2,493,071 | 14,825 | 32 |
| 23 | 371 | 279.92 | 3240 | 137 | 71,562 | 2,492,410 | 14,287 | 32 |
| 25 | 386 | 220.87 | 2819 | 137 | 47,447 | 2,046,024 | 10,904 | 34 |
| 26 | 362 | 277.80 | 3243 | 137 | 56,956 | 2,413,478 | 15,510 | 29 |
| 27 | 441 | 228.60 | 3284 | 137 | 55,778 | 2,419,417 | 10,757 | 31 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hoefer, A.; Boyen, F.; Beierschmitt, A.; Moodley, A.; Roberts, M.C.; Butaye, P. Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts. Antibiotics 2021, 10, 290. https://doi.org/10.3390/antibiotics10030290
Hoefer A, Boyen F, Beierschmitt A, Moodley A, Roberts MC, Butaye P. Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts. Antibiotics. 2021; 10(3):290. https://doi.org/10.3390/antibiotics10030290
Chicago/Turabian StyleHoefer, Andreas, Filip Boyen, Amy Beierschmitt, Arshnee Moodley, Marilyn C. Roberts, and Patrick Butaye. 2021. "Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts" Antibiotics 10, no. 3: 290. https://doi.org/10.3390/antibiotics10030290
APA StyleHoefer, A., Boyen, F., Beierschmitt, A., Moodley, A., Roberts, M. C., & Butaye, P. (2021). Methicillin-Resistant and Methicillin-Susceptible Staphylococcus from Vervet Monkeys (Chlorocebus sabaeus) in Saint Kitts. Antibiotics, 10(3), 290. https://doi.org/10.3390/antibiotics10030290