
A Survey of Spontaneous Antibiotic-Resistant Mutants of the Halophilic, Thermophilic Bacterium Rhodothermus marinus


 and
and 
Abstract
1. Introduction
2. Results
2.1. Spectrum of Antibiotic Sensitivity
2.2. Selection of Spontaneous Mutants
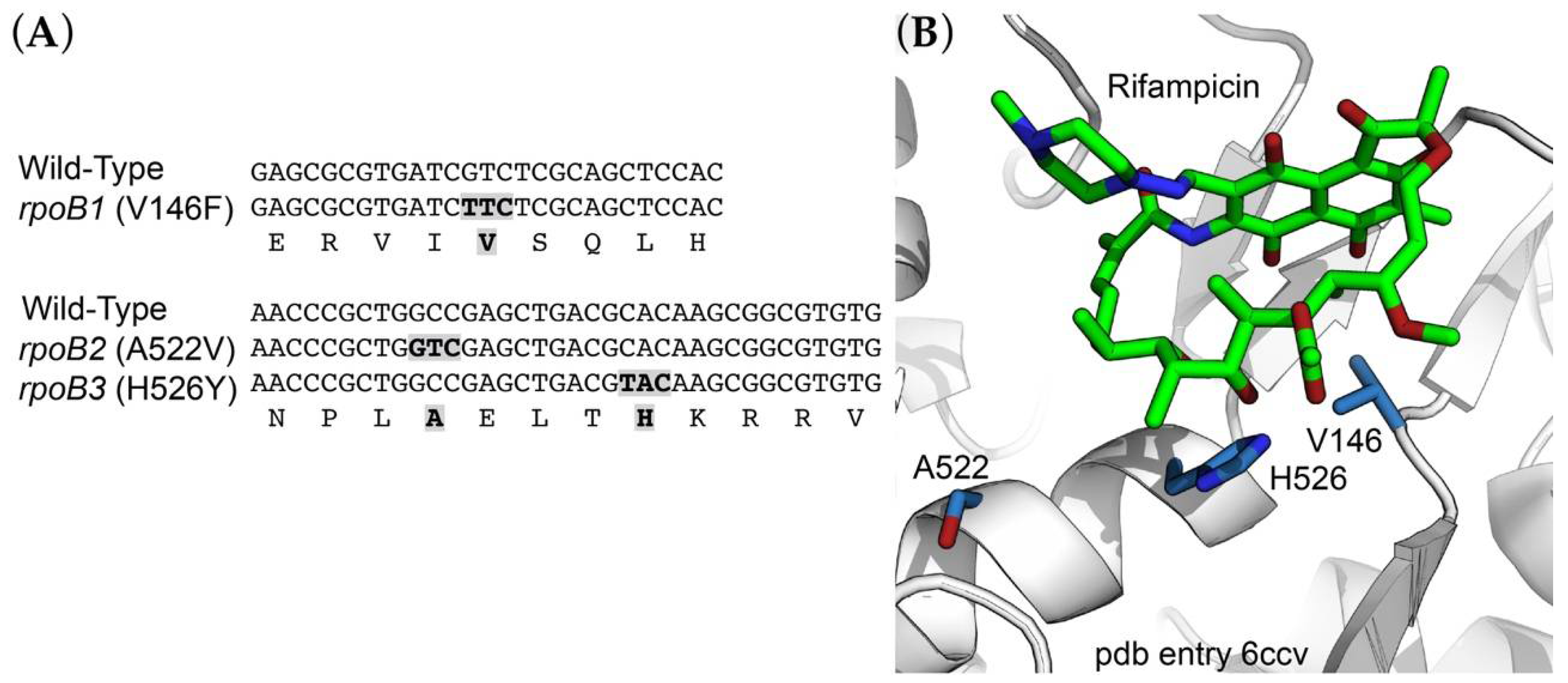
2.3. Rifampicin-Resistance (RifR) Mutations in the RNA Polymerase β Subunit
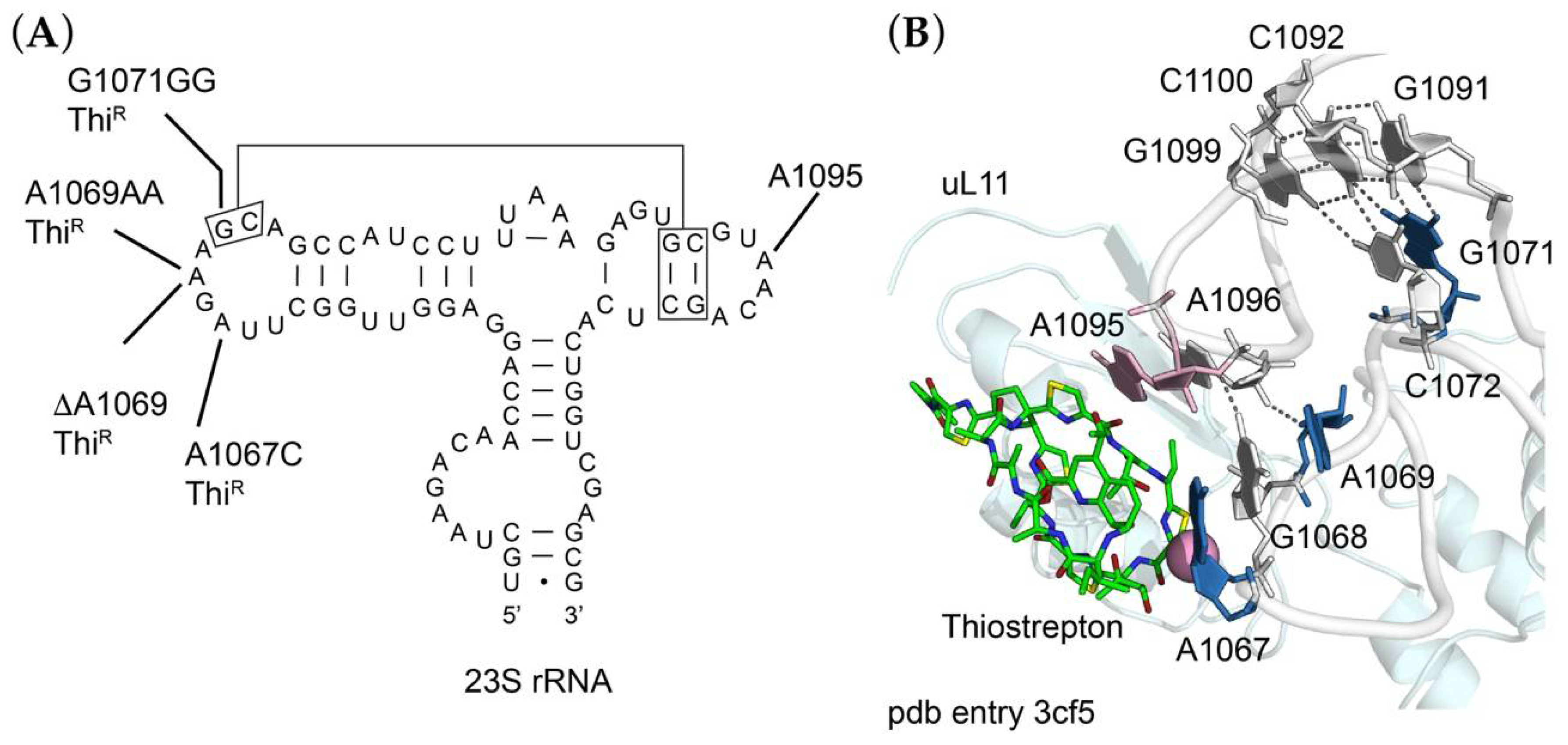
2.4. Thiostrepton-Resistance Mutations in the 50S Ribosomal Subunit
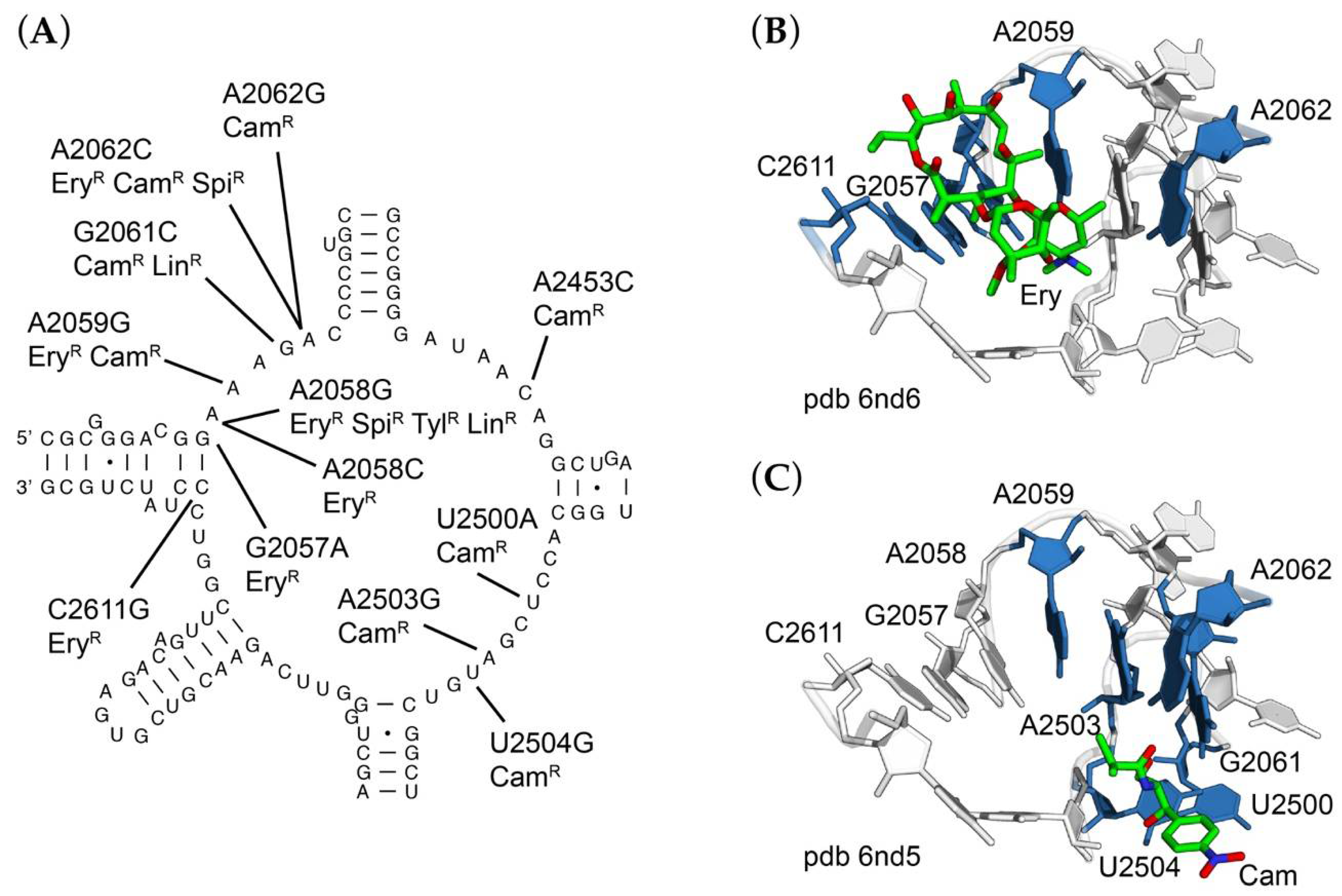
2.5. Base Substitutions of 23S rRNA Residues in and around the Ribosomal Peptidyltransferase Active Site
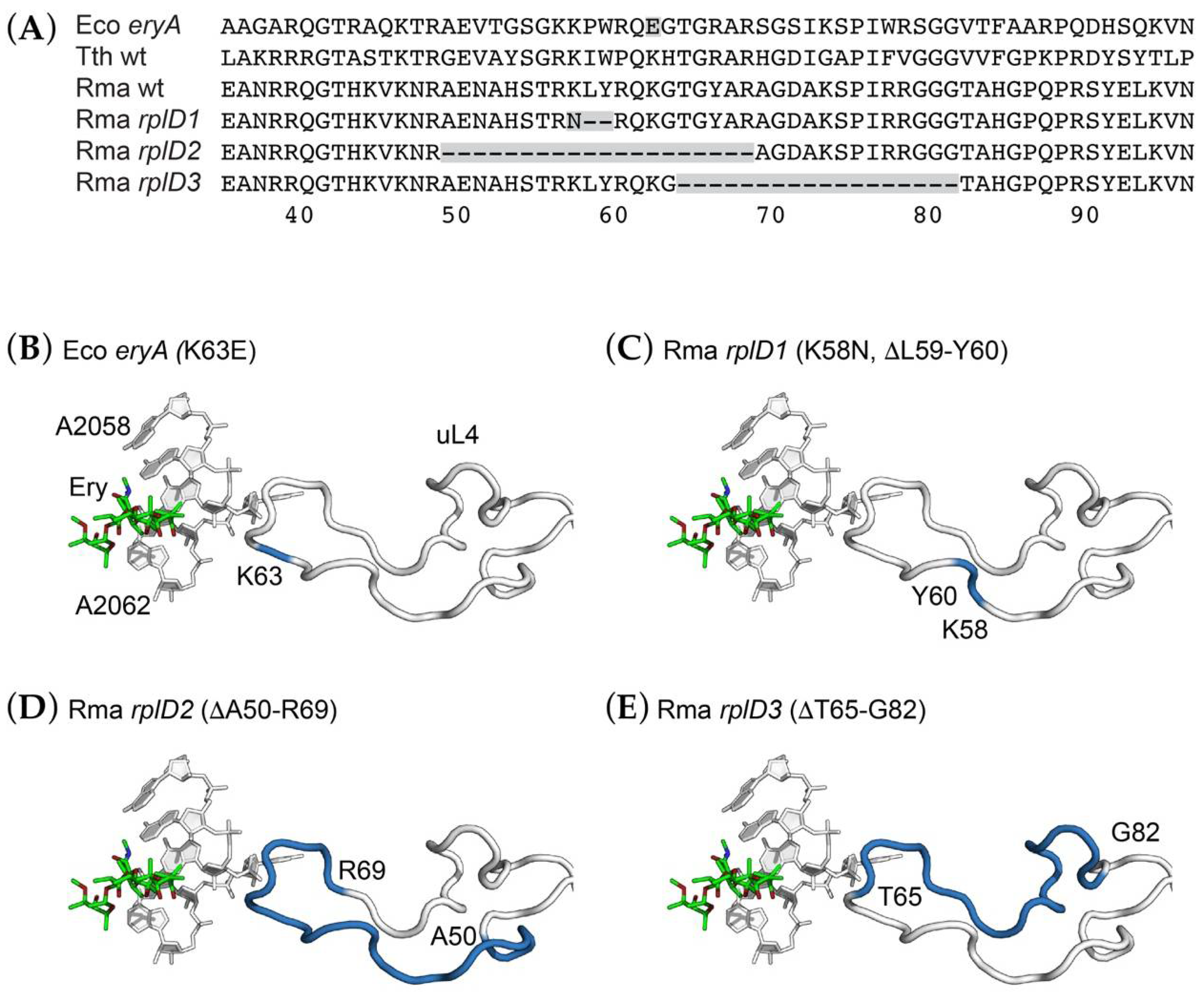
2.6. Deletions in a Conserved Loop of Ribosomal Protein uL4 Conferring Erythromycin Resistance
3. Discussion
4. Materials and Methods
4.1. Strains and Growth Conditions, Assessment of Antibiotic Sensitivity, and Isolation of Mutants
4.2. Identification of Mutations
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ma, C.; Yang, X.; Lewis, P.J. Bacterial transcription as a target for antibacterial drug development. Microbiol. Mol. Biol. Rev. 2016, 80, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Murakami, K.S. Structural biology of bacterial RNA polymerase. Biomolecules 2015, 5, 848–864. [Google Scholar] [CrossRef] [PubMed]
- Wilson, D.N. Ribosome-targeting antibiotics and mechanisms of bacterial resistance. Nat. Rev. Microbiol. 2014, 12, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Alfredsson, G.A.; Kristjansson, J.K.; Hjörleifdottir, S.; Stetter, K.O. Rhodothermus marinus, gen. nov., sp. nov., a thermophilic, halophilic bacterium from submarine hot springs in Iceland. J. Gen. Microbiol. 1988, 134, 299–306. [Google Scholar] [CrossRef]
- Bjornsdottir, S.H.; Blondal, T.; Hreggvidsson, G.O.; Eggertsson, G.; Petursdottir, S.; Hjorleifsdottir, S.; Thorbjarnardottir, S.H.; Kristjansson, J.K. Rhodothermus marinus: Physiology and molecular biology. Extremophiles 2006, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Antón, J.; Oren, A.; Benlloch, S.; Rodríguez-Valera, F.; Amann, R.; Rosselló-Mora, R. Salinibacter ruber gen. nov., sp. nov., a novel, extremely halophilic member of the Bacteria from saltern crystallizer ponds. Int. J. Syst. Evol. Microbiol. 2002, 52, 485–491. [Google Scholar] [CrossRef] [PubMed]
- Mongodin, E.F.; Nelson, K.E.; Daugherty, S.; Deboy, R.T.; Wister, J.; Khouri, H.; Weidman, J.; Walsh, D.A.; Papke, R.T.; Sanchez Perez, G.; et al. The genome of Salinibacter ruber: Convergence and gene exchange among hyperhalophilic bacteria and archaea. Proc. Natl. Acad. Sci. USA 2005, 102, 18147–18152. [Google Scholar] [CrossRef]
- Vaisman, N.; Oren, A. Salisaeta longa gen. nov., sp. nov., a red, halophilic member of the Bacteroidetes. Int. J. Syst. Evol. Microbiol. 2009, 59, 2571–2574. [Google Scholar] [CrossRef]
- Park, S.; Yoshizawa, S.; Kogure, K.; Yokota, A. Rubricoccus marinus gen. nov., sp. nov., of the family ‘Rhodothermaceae’, isolated from seawater. Int. J. Syst. Evol. Microbiol. 2011, 61, 2069–2072. [Google Scholar] [CrossRef]
- Park, S.; Song, J.; Yoshizawa, S.; Choi, A.; Cho, J.C.; Kogure, K. Rubrivirga marina gen. nov., sp. nov., a member of the family Rhodothermaceae isolated from deep seawater. Int. J. Syst. Evol. Microbiol. 2013, 63, 2229–2233. [Google Scholar] [CrossRef]
- Xia, J.; Zhou, Y.X.; Zhao, L.H.; Chen, G.J.; Du, Z.J. Longimonas halophila gen. nov., sp. nov., isolated from a marine solar saltern. Int. J. Syst. Evol. Microbiol. 2015, 65, 2272–2276. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Dunlap, C.A.; Flor-Weiler, L.; Rooney, A.P.; Chen, G.J.; Du, Z.J. Longibacter salinarum gen. nov., sp. nov., isolated from a marine solar saltern. Int. J. Syst. Evol. Microbiol. 2016, 66, 3287–3292. [Google Scholar] [CrossRef]
- Kristjansson, J.K.; Hreggvidsson, G.O.; Alfredsson, G.A. Isolation of halotolerant Thermus spp. from submarine hot springs in Iceland. Appl. Environ. Microbiol. 1986, 52, 1313–1316. [Google Scholar] [CrossRef] [PubMed]
- Sousa, J.S.; Calisto, F.; Langer, J.D.; Mills, D.J.; Refojo, P.N.; Teixeira, M.; Kühlbrandt, W.; Vonck, J.; Pereira, M.M. Structural basis for energy transduction by respiratory alternative complex III. Nat. Commun. 2018, 9, 1728. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, R.; Sekirnik, R.; Brissett, N.C.; Krojer, T.; Ho, C.H.; Ng, S.S.; Clifton, I.J.; Ge, W.; Kershaw, N.J.; Fox, G.C.; et al. Ribosomal oxygenases are structurally conserved from prokaryotes to humans. Nature 2014, 510, 422–426. [Google Scholar] [CrossRef] [PubMed]
- Andrésson, O.S.; Fridjónsson, O.H. The sequence of the single 16S rRNA gene of the thermophilic eubacterium Rhodothermus marinus reveals a distant relationship to the group containing Flexibacter, Bacteroides, and Cytophaga species. J. Bacteriol. 1994, 176, 6165–6169. [Google Scholar] [CrossRef]
- Nolan, M.; Tindall, B.J.; Pomrenke, H.; Lapidus, A.; Copeland, A.; Glavina Del Rio, T.; Lucas, S.; Chen, F.; Tice, H.; Cheng, J.F.; et al. Complete genome sequence of Rhodothermus marinus type strain (R-10). Stand. Genom. Sci. 2009, 1, 283–290. [Google Scholar] [CrossRef][Green Version]
- Sigmund, C.D.; Ettayebi, M.; Morgan, E.A. Antibiotic resistance mutations in 16S and 23S ribosomal RNA genes of Escherichia coli. Nucleic Acids Res. 1984, 12, 4653–4663. [Google Scholar] [CrossRef]
- Asai, T.; Zaporojets, D.; Squires, C.; Squires, C.L. An Escherichia coli strain with all chromosomal rRNA operons inactivated: Complete exchange of rRNA genes between bacteria. Proc. Natl. Acad. Sci. USA 1999, 96, 1971–1976. [Google Scholar] [CrossRef]
- Hobbie, S.N.; Bruell, C.; Kalapala, S.; Akshay, S.; Schmidt, S.; Pfister, P.; Böttger, E.C. A genetic model to investigate drug-target interactions at the ribosomal decoding site. Biochimie 2006, 88, 1033–1043. [Google Scholar] [CrossRef]
- Gregory, S.T.; Dahlberg, A.E. Genetic and structural analysis of base substitutions in the central pseudoknot of Thermus thermophilus 16S ribosomal RNA. RNA 2009, 15, 215–223. [Google Scholar] [CrossRef][Green Version]
- Gregory, S.T.; Carr, J.F.; Rodriguez-Correa, D.; Dahlberg, A.E. Mutational analysis of 16S and 23S rRNA genes of Thermus thermophilus. J. Bacteriol. 2005, 187, 4804–4812. [Google Scholar] [CrossRef] [PubMed]
- Bjornsdottir, S.H.; Thorbjarnardottir, S.H.; Eggertsson, G. Establishment of a gene transfer system for Rhodothermus marinus. Appl. Microbiol. Biotechnol. 2005, 66, 675–682. [Google Scholar] [CrossRef] [PubMed]
- Bjornsdottir, S.H.; Fridjonsson, O.H.; Hreggvidsson, G.O.; Eggertsson, G. Generation of targeted deletions in the genome of Rhodothermus marinus. Appl. Environ. Microbiol. 2011, 77, 5505–5512. [Google Scholar] [CrossRef]
- Bjornsdottir, S.H.; Fridjonsson, O.H.; Kristjansson, J.K.; Eggertsson, G. Cloning and expression of heterologous genes in Rhodothermus marinus. Extremophiles 2007, 11, 283–293. [Google Scholar] [CrossRef] [PubMed]
- Maus, C.E.; Plikaytis, B.B.; Shinnick, T.M. Molecular analysis of cross-resistance to capreomycin, kanamycin, amikacin, and viomycin in Mycobacterium tuberculosis. Antimicrob. Agents Chemother. 2005, 49, 3192–3197. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, B.P. Resistance to rifampicin: A review. J. Antibiot. 2014, 67, 625–630. [Google Scholar] [CrossRef] [PubMed]
- Peek, J.; Lilic, M.; Montiel, D.; Milshteyn, A.; Woodworth, I.; Biggins, J.B.; Ternei, M.A.; Calle, P.Y.; Danziger, M.; Warrier, T.; et al. Rifamycin congeners kanglemycins are active against rifampicin-resistant bacteria via a distinct mechanism. Nat. Commun. 2018, 9, 4147. [Google Scholar] [CrossRef]
- Harms, J.M.; Wilson, D.N.; Schluenzen, F.; Connell, S.R.; Stachelhaus, T.; Zaborowska, Z.; Spahn, C.M.; Fucini, P. Translational regulation via L11: Molecular switches on the ribosome turned on and off by thiostrepton and micrococcin. Mol. Cell 2008, 30, 26–38. [Google Scholar] [CrossRef]
- Ban, N.; Nissen, P.; Hansen, J.; Moore, P.B.; Steitz, T.A. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science 2000, 289, 905–920. [Google Scholar] [CrossRef]
- Gale, E.F.; Cundliffe, P.E.; Reynolds, M.H.; Richmond, M.J. Waring. In The Molecular Basis of Antibiotic Action; John Wiley & Sons: London, UK, 1981. [Google Scholar]
- Vázquez-Laslop, N.; Mankin, A.S. How macrolide antibiotics work. Trends Biochem. Sci. 2018, 43, 668–684. [Google Scholar] [CrossRef] [PubMed]
- Svetlov, M.S.; Plessa, E.; Chen, C.W.; Bougas, A.; Krokidis, M.G.; Dinos, G.P.; Polikanov, Y.S. High-resolution crystal structures of ribosome-bound chloramphenicol and erythromycin provide the ultimate basis for their competition. RNA 2019, 25, 600–606. [Google Scholar] [CrossRef] [PubMed]
- Fyfe, C.; Grossman, T.H.; Kerstein, K.; Sutcliffe, J. Resistance to macrolide antibiotics in public health pathogens. Cold Spring Harb. Perspect. Med. 2016, 6, a025395. [Google Scholar] [CrossRef]
- Diner, E.J.; Hayes, C.S. Recombineering reveals a diverse collection of ribosomal proteins L4 and L22 that confer resistance to macrolide antibiotics. J. Mol. Biol. 2009, 386, 300–315. [Google Scholar] [CrossRef]
- Wittmann, H.G.; Stoffler, G.; Apirion, D.; Rosen, L.; Tanaka, K.; Tamaki, M.; Takata, R.; Dekio, S.; Otaka, E.; Osawa, S. Biochemical and genetic studies of two different types of erythromycin resistant mutants of Escherichia coli with altered ribosomal proteins. Mol. Gen. Genet. 1973, 127, 175–189. [Google Scholar] [CrossRef] [PubMed]
- Chittum, H.S.; Champney, W.S. Ribosomal protein gene sequence changes in erythromycin-resistant mutants of Escherichia coli. J. Bacteriol. 1994, 176, 6192–6198. [Google Scholar] [CrossRef]
- Zengel, J.M.; Jerauld, A.; Walker, A.; Wahl, M.C.; Lindahl, L. The extended loops of ribosomal proteins L4 and L22 are not required for ribosome assembly or L4-mediated autogenous control. RNA 2003, 9, 1188–1197. [Google Scholar] [CrossRef] [PubMed]
- Zaman, S.; Fitzpatrick, M.; Lindahl, L.; Zengel, J. Novel mutations in ribosomal proteins L4 and L22 that confer erythromycin resistance in Escherichia coli. Mol. Microbiol. 2007, 66, 1039–1050. [Google Scholar] [CrossRef]
- Lawrence, M.G.; Shamsuzzaman, M.; Kondopaka, M.; Pascual, C.; Zengel, J.M.; Lindahl, L. The extended loops of ribosomal proteins uL4 and uL22 of Escherichia coli contribute to ribosome assembly and protein translation. Nucleic Acids Res. 2016, 44, 5798–5810. [Google Scholar] [CrossRef]
- Murakami, K.S.; Masuda, S.; Darst, S.A. Structural basis of transcription initiation: RNA polymerase holoenzyme at 4 Å resolution. Science 2002, 296, 1280–1284. [Google Scholar] [CrossRef]
- Vassylyev, D.G.; Sekine, S.; Laptenko, O.; Lee, J.; Vassylyeva, M.N.; Borukhov, S.; Yokoyama, S. Crystal structure of a bacterial RNA polymerase holoenzyme at 2.6 Å resolution. Nature 2002, 417, 712–719. [Google Scholar] [CrossRef]
- Murakami, K.S.; Darst, S.A. Bacterial RNA polymerases: The wholo story. Curr. Opin. Struct. Biol. 2003, 13, 31–39. [Google Scholar] [CrossRef]
- Campbell, E.A.; Korzheva, N.; Mustaev, A.; Murakami, K.; Nair, S.; Goldfarb, A.; Darst, S.A. Structural mechanism for rifampicin inhibition of bacterial RNA polymerase. Cell 2001, 104, 901–912. [Google Scholar] [CrossRef]
- Tu, D.; Blaha, G.; Moore, P.B.; Steitz, T.A. Structures of MLSBK antibiotics bound to mutated large ribosomal subunits provide a structural explanation for resistance. Cell 2005, 121, 257–270. [Google Scholar] [CrossRef] [PubMed]
- O’Connor, M.; Gregory, S.T.; Dahlberg, A.E. Multiple defects in translation associated with altered ribosomal protein L4. Nucleic Acids Res. 2004, 32, 5750–5756. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Gregory, S.T.; Dahlberg, A.E. Erythromycin resistance mutations in ribosomal protein L22 and L4 perturb the higher order structure of 23S ribosomal RNA. J. Mol. Biol. 1999, 289, 827–834. [Google Scholar] [CrossRef]
- Gabashvili, I.S.; Gregory, S.T.; Valle, M.; Grassucci, R.; Worbs, M.; Wahl, M.C.; Dahlberg, A.E.; Frank, J. The polypeptide tunnel system in the ribosome and its gating in erythromycin resistance mutants of L4 and L22. Mol. Cell 2001, 8, 181–188. [Google Scholar] [CrossRef]
- Bryan, L.E.; Kowand, S.K.; Van Den Elzen, H.M. Mechanism of aminoglycoside antibiotic resistance in anaerobic bacteria: Clostridium perfringens and Bacteroides fragilis. Antimicrob. Agents Chemother. 1979, 15, 7–13. [Google Scholar] [CrossRef]
- Cannone, J.J.; Subramanian, S.; Schnare, M.N.; Collett, J.R.; D’Souza, L.M.; Du, Y.; Feng, B.; Lin, N.; Madabusi, L.V.; Müller, K.M.; et al. The comparative RNA web (CRW) site: An online database of comparative sequence and structure information for ribosomal, intron, and other RNAs. BMC Bioinform. 2002, 3, 2. [Google Scholar]
- The RNAcentral Consortium; Petrov, A.I.; Kay, S.J.E.; Kalvari, I.; Howe, K.L.; Gray, K.A.; Bruford, E.A.; Kersey, P.J.; Cochrane, G.; Finn, R.D.; et al. RNAcentral: A comprehensive database of non-coding RNA sequences. Nucleic Acids Res. 2017, 45, D128–D134. [Google Scholar] [PubMed]
- Li, M.; Tzagoloff, A.; Underbrink-Lyon, K.; Martin, N.C. Identification of the paromomycin-resistance mutation in the 15 S rRNA gene of yeast mitochondria. J. Biol. Chem. 1982, 257, 5921–5928. [Google Scholar] [CrossRef]
- Spangler, E.A.; Blackburn, E.H. The nucleotide sequence of the 17S ribosomal RNA gene of Tetrahymena thermophila and the identification of point mutations resulting in resistance to the antibiotics paromomycin and hygromycin. J. Biol. Chem. 1985, 260, 6334–6340. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic Class, (Target) | Antibiotic | Response |
|---|---|---|
| aminoglycoside (ribosome, 30S) | streptomycin, apramycin, hygromycin B, gentamicin, neomycin, neamine, ribostamycin, kanamycin, tobramycin, paromomycin | resistant |
| aminocyclitols (ribosome, 30S) | kasugamycin, spectinomycin | resistant |
| tuberactinomycin (ribosome, 70S) | capreomycin | resistant |
| 14-atom macrolides (ribosome, 50S) | erythromycin, oleandomycin, roxithromycin, clarithromycin | sensitive |
| 15-atom macrolide (ribosome, 50S) | azithromycin | sensitive |
| 16-atom macrolides (ribosome, 50S) | spiramycin, chalcomycin, tylosin, carbomycin | sensitive |
| lincosamides (ribosome, 50S) | lincomycin, clindamycin | sensitive |
| streptogramin B (ribosome, 50S) | pristinamycin | sensitive |
| pleuromutilin (ribosome, 50S) | tiamulin | sensitive |
| amphenicol (ribosome, 50S) | chloramphenicol | sensitive |
| thiopeptide, (ribosome, 50S) | thiostrepton | sensitive |
| elfamycin (EF-Tu) 1 | kirromycin | sensitive |
| fusidane (EF-G) 2 | fusidic acid | sensitive |
| rifamycin (RNA polymerase) | rifampicin | sensitive |
| quinolone (DNA gyrase) | nalidixic acid | resistant |
| fluoroquinolones (DNA gyrase) | ofloxacin, ciprofloxacin | sensitive |
| Allele | Rep Strain | Mutation | Selection |
|---|---|---|---|
| rpoB1 | SOP89 | RNA pol β subunit-V146F | Rif50, 100 |
| rpoB2 | SOP90 | RNA pol β subunit-A522V | Rif50, 100 |
| rpoB3 | SOP91 | RNA pol β subunit-H526Y | Rif50 |
| rrlA1 | SOP9 | 23S rRNA-G2057A | Ery50, 100, 200 |
| rrlA2 | SOP23 | 23S rRNA-A2058C | Ery200 |
| rrlA3 | SOP11 | 23S rRNA-A2058G | Ery50, 100, 200/Spi100/Tyl100/Lnc100 |
| rrlA4 | SOP26 | 23S rRNA-A2059G | Ery200/Chl100 |
| rrlA5 | SOP56 | 23S rRNA-G2061C | Chl100/Lnc100 |
| rrlA6 | SOP29 | 23S rRNA-A2062C | Chl25/Ery50/Spi100 |
| rrlA7 | SOP38 | 23S rRNA-A2062G | Chl25/Ery50 |
| rrlA8 | SOP74 | 23S rRNA-A2453C | Chl25 |
| rrlA9 | SOP5 | 23S rRNA-U2500A | Chl50 |
| rrlA10 | SOP4 | 23S rRNA-A2503C | Chl50 |
| rrlA11 | SOP1 | 23S rRNA-A2503G | Chl25, 50 |
| rrlA12 | SOP3 | 23S rRNA-U2504G | Chl25, 50 |
| rrlA13 | SOP14 | 23S rRNA-U2611G | Ery50 |
| rrlA14 | SOP7 | 23S rRNA-A2453C/U2500A | Chl50 |
| rrlA15 | SOP24 | 23S rRNA-G2057A/A2062G | Ery200 |
| rrlA16 | SOP60 | 23S rRNA-A2062G/A2503G | Spi100 |
| rrlA17 | SOP72 | 23S rRNA-A1067C | Thi200 |
| rrlA18 | SOP79 | 23S rRNA-ΔA1069 | Thi100 |
| rrlA19 | SOP73 | 23S rRNA-A1069AA | Thi200 |
| rrlA20 | SOP77 | 23S rRNA-G1071GG | Thi100 |
| rplD1 | SOP57 | uL4-K58N, ΔL59-Y60 | Ery100, 200 |
| rplD2 | SOP25 | uL4-ΔA50-R69 | Ery200 |
| rplD3 | SOP16 | uL4-ΔT65-G82 | Ery100 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Silvia, S.; Donahue, S.A.; Killeavy, E.E.; Jogl, G.; Gregory, S.T. A Survey of Spontaneous Antibiotic-Resistant Mutants of the Halophilic, Thermophilic Bacterium Rhodothermus marinus. Antibiotics 2021, 10, 1384. https://doi.org/10.3390/antibiotics10111384
Silvia S, Donahue SA, Killeavy EE, Jogl G, Gregory ST. A Survey of Spontaneous Antibiotic-Resistant Mutants of the Halophilic, Thermophilic Bacterium Rhodothermus marinus. Antibiotics. 2021; 10(11):1384. https://doi.org/10.3390/antibiotics10111384
Chicago/Turabian StyleSilvia, Sophia, Samantha A. Donahue, Erin E. Killeavy, Gerwald Jogl, and Steven T. Gregory. 2021. "A Survey of Spontaneous Antibiotic-Resistant Mutants of the Halophilic, Thermophilic Bacterium Rhodothermus marinus" Antibiotics 10, no. 11: 1384. https://doi.org/10.3390/antibiotics10111384
APA StyleSilvia, S., Donahue, S. A., Killeavy, E. E., Jogl, G., & Gregory, S. T. (2021). A Survey of Spontaneous Antibiotic-Resistant Mutants of the Halophilic, Thermophilic Bacterium Rhodothermus marinus. Antibiotics, 10(11), 1384. https://doi.org/10.3390/antibiotics10111384