
Repurposing Eltrombopag for Multidrug Resistant Staphylococcus aureus Infections

 , ,
, ,
Abstract
:1. Introduction
2. Results and Discussion
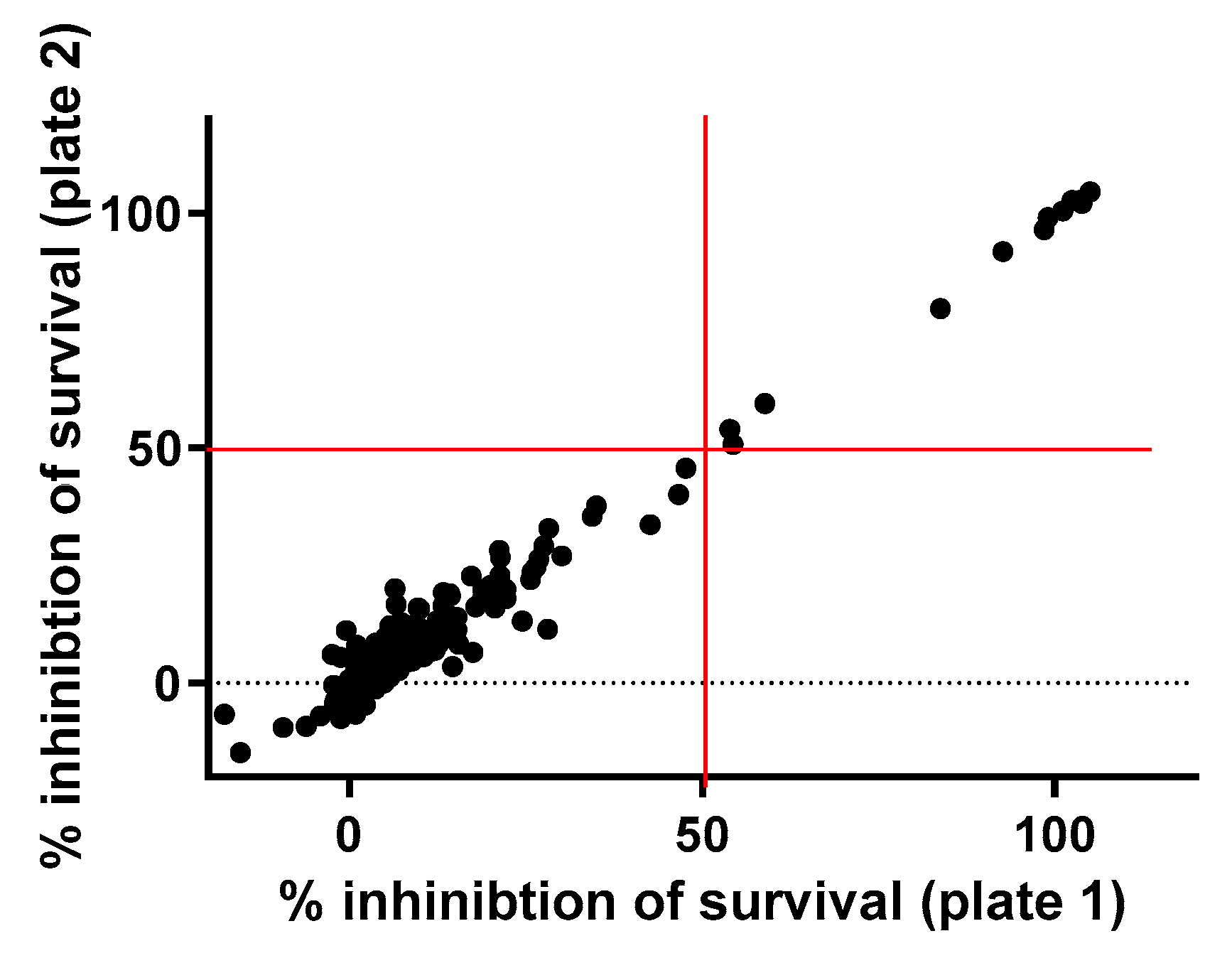
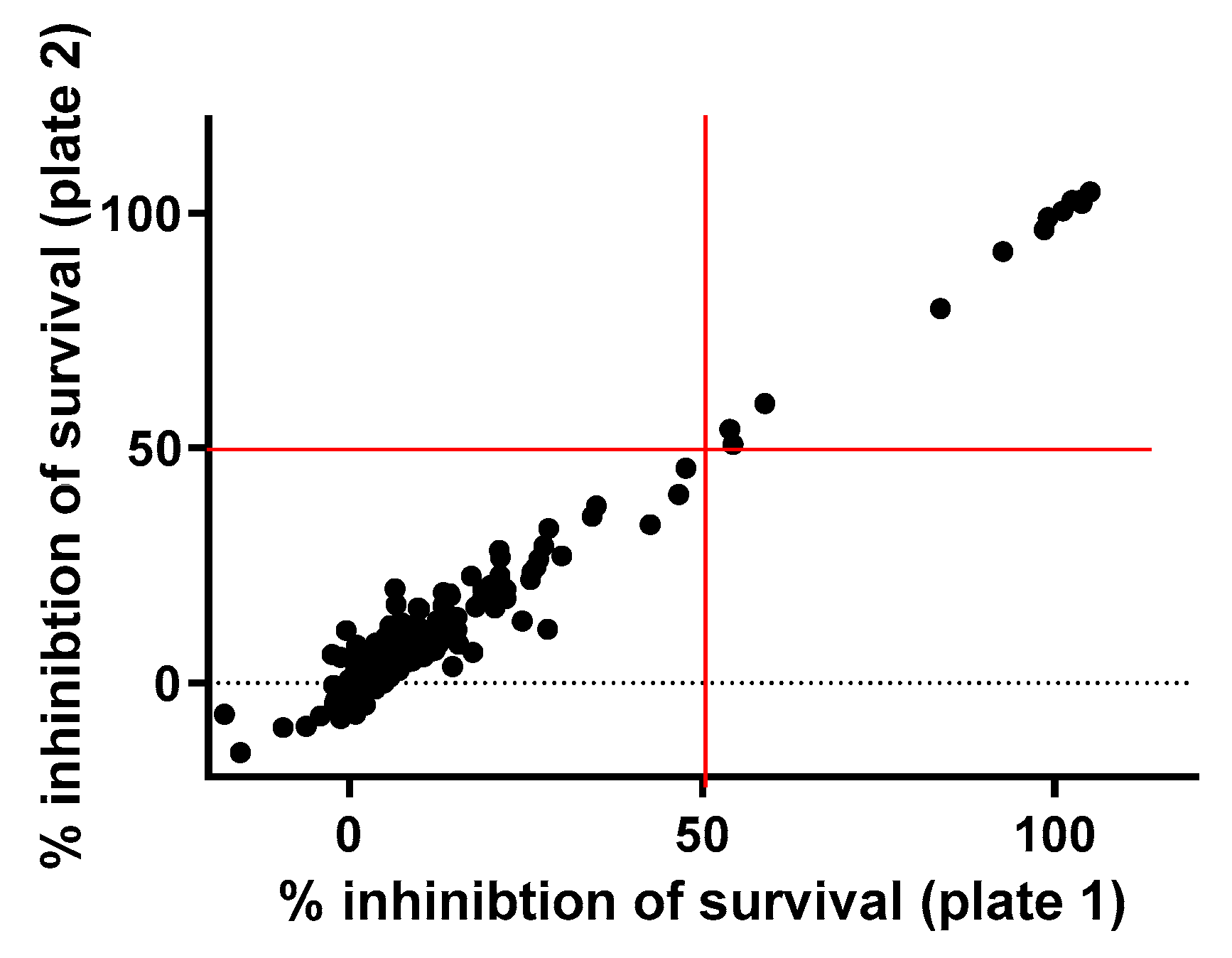
2.1. Primary Screening
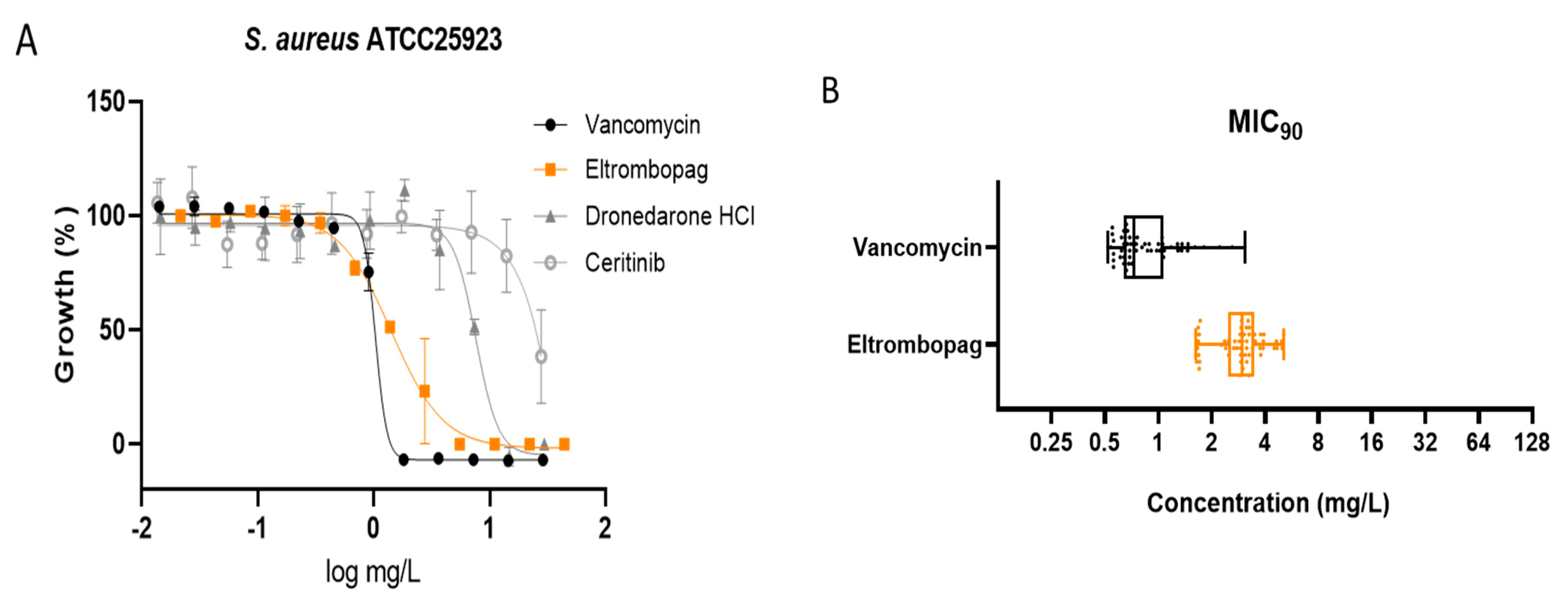
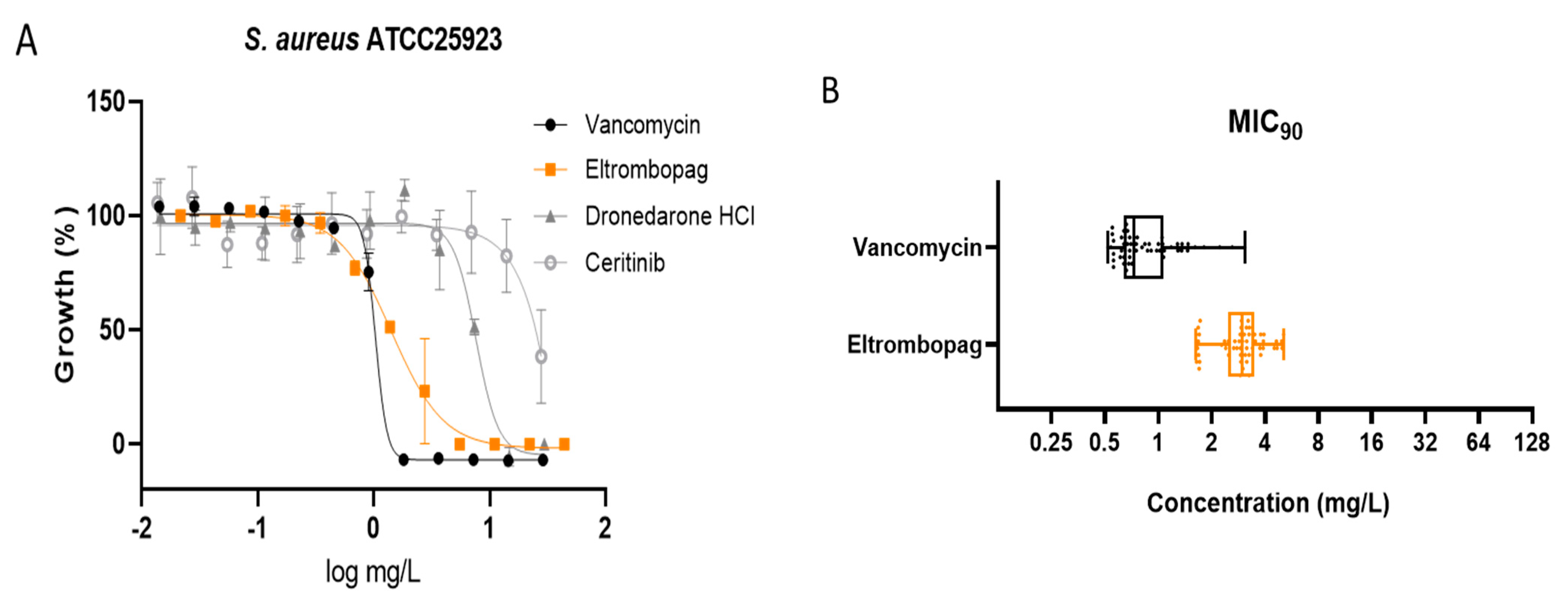
2.2. Dose–Response Assay of Selected Repositionable Candidates
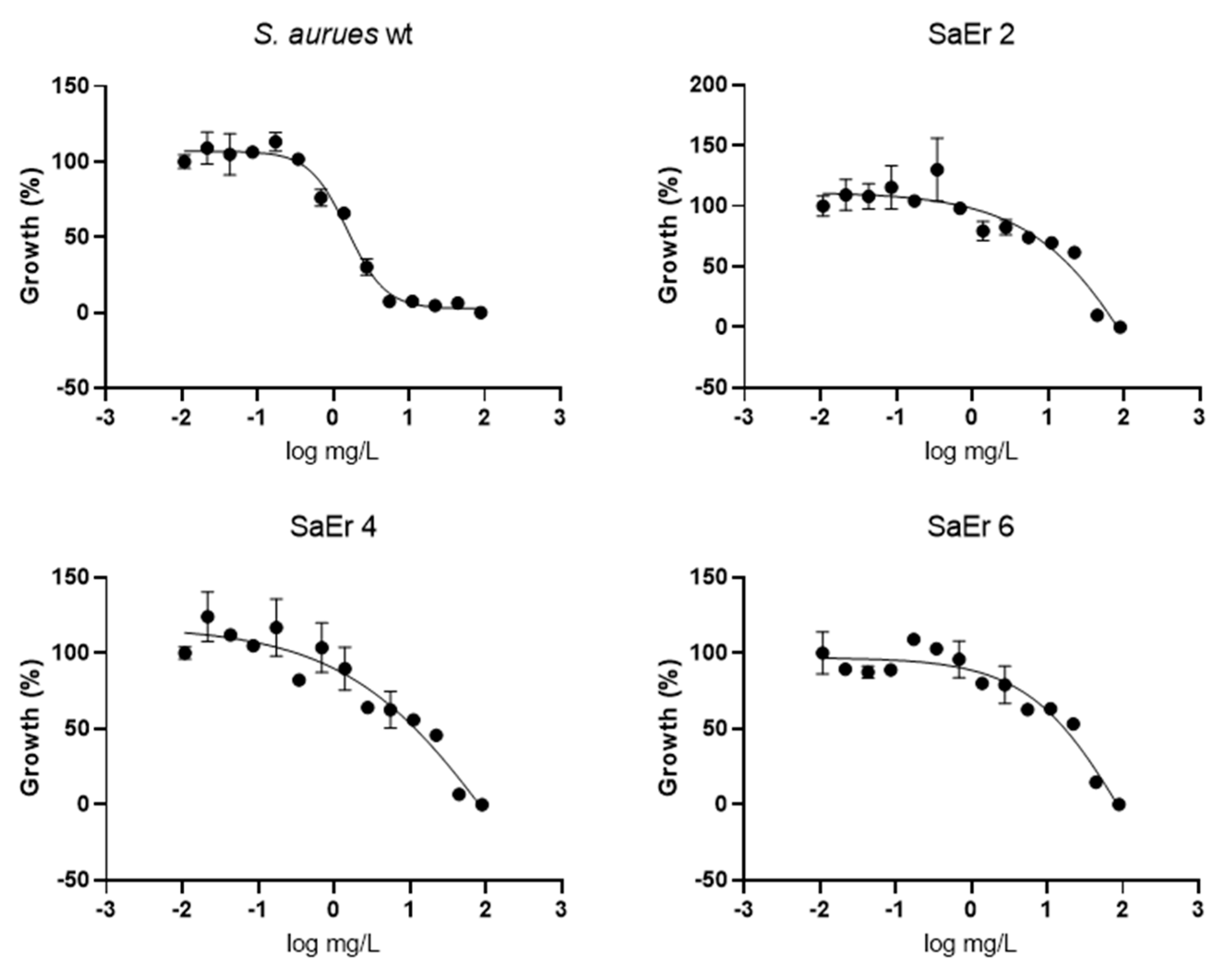
2.3. Confirmation of Eltrombopag Activity against S. aureus Clinical Isolates and Other Bacterial Species
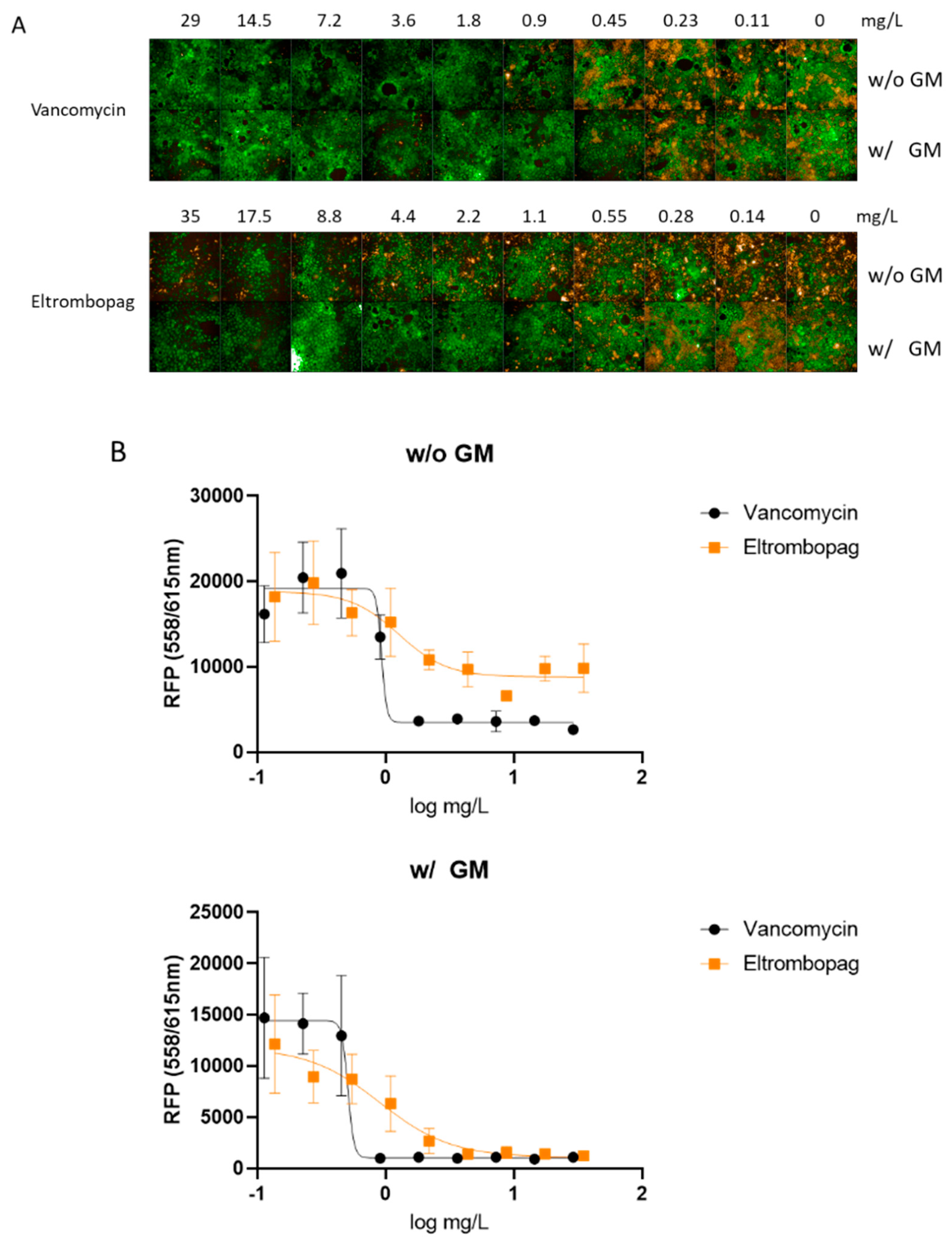
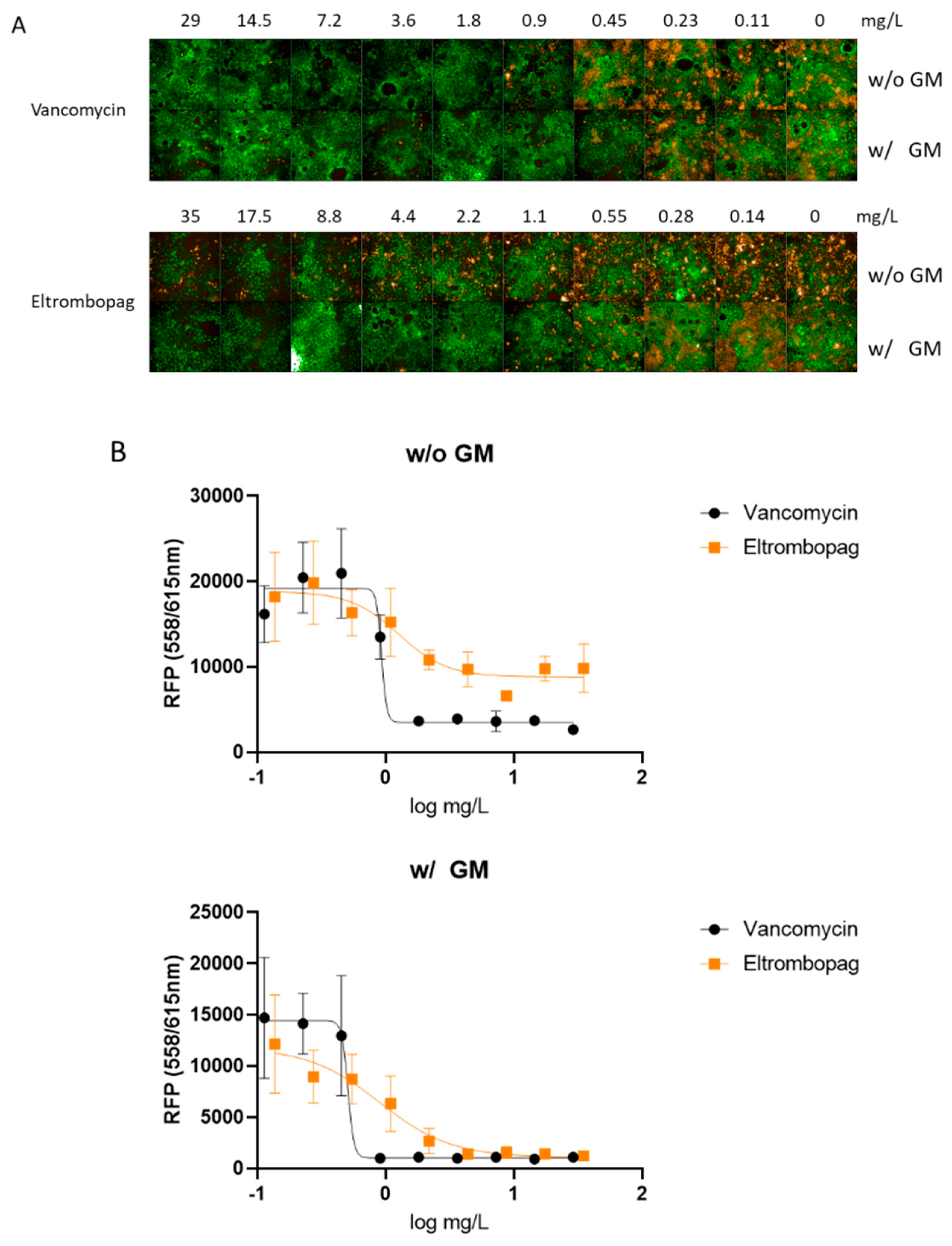
2.4. Eltrombopag Inhibits S. aureus Growth in a Cell Line Infection Model
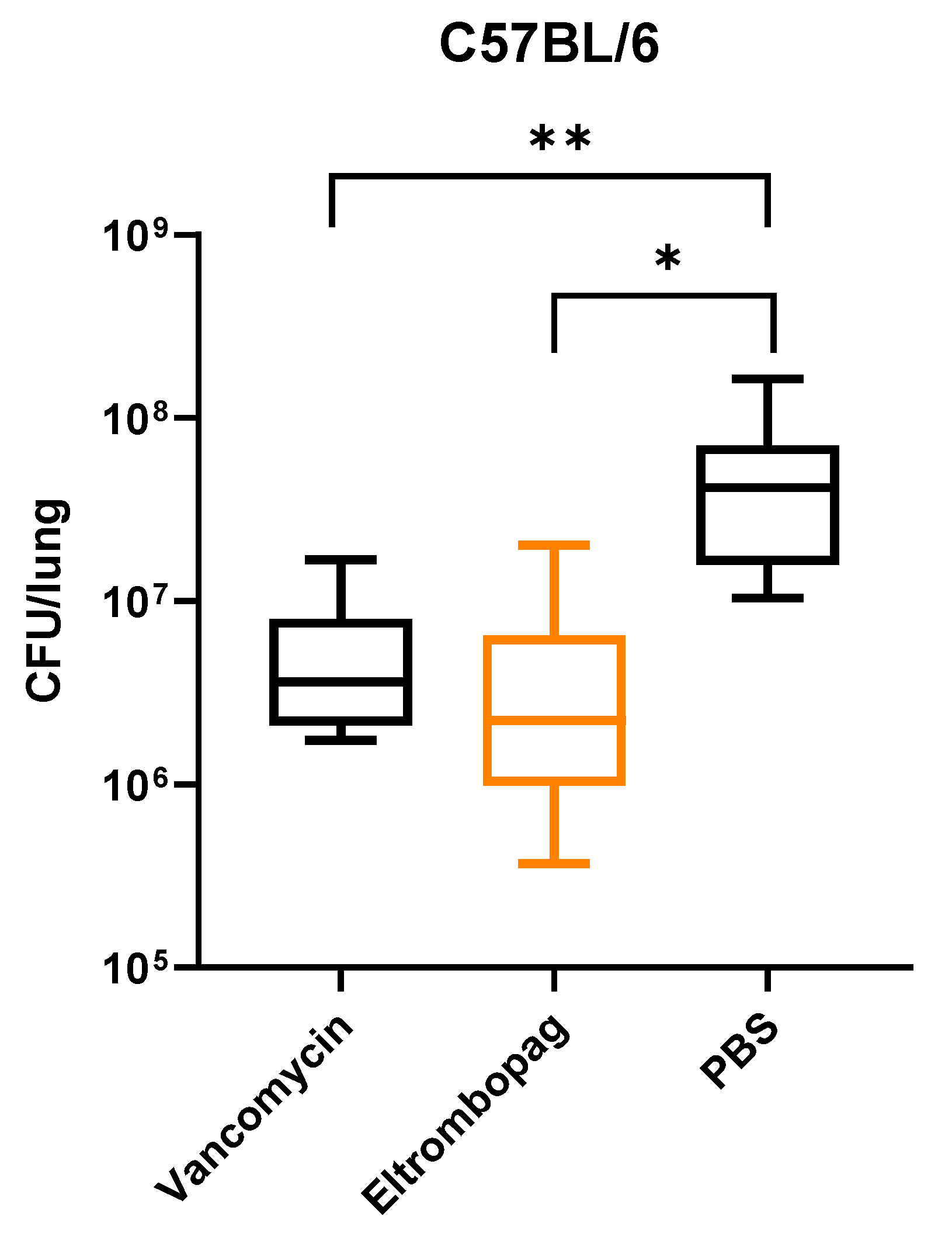
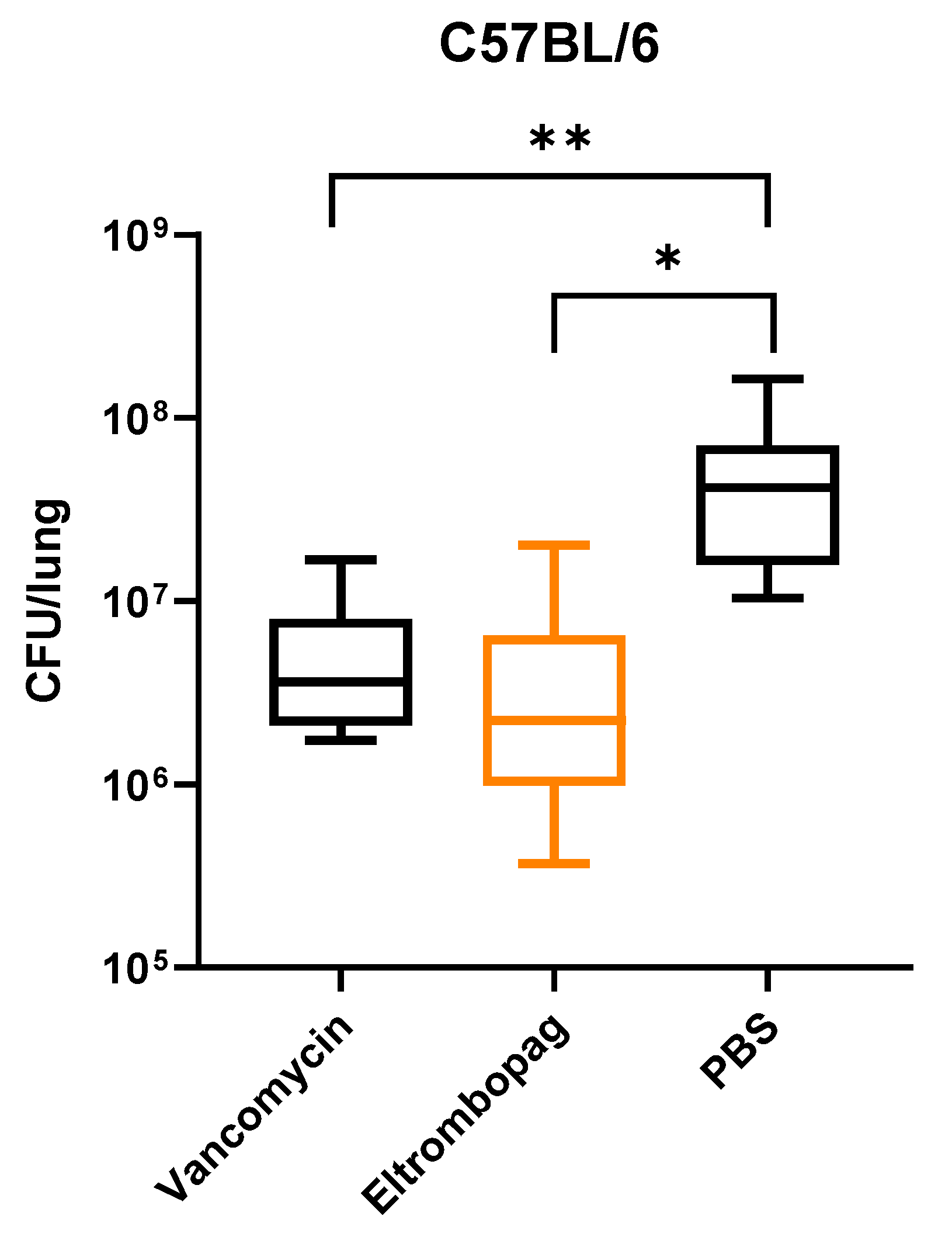
2.5. In Vivo Efficacy of Eltrombopag in a Mouse Infection Model
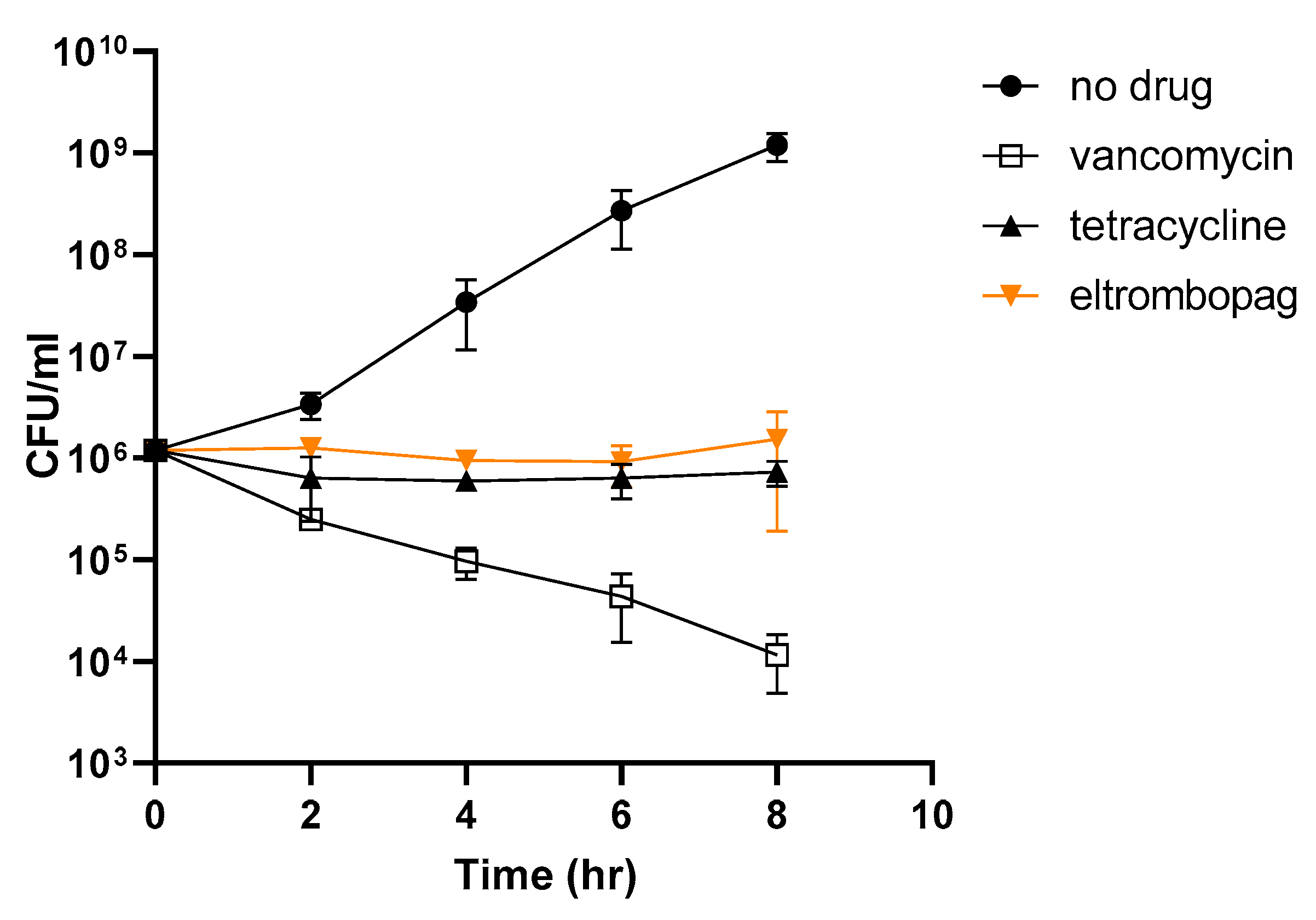
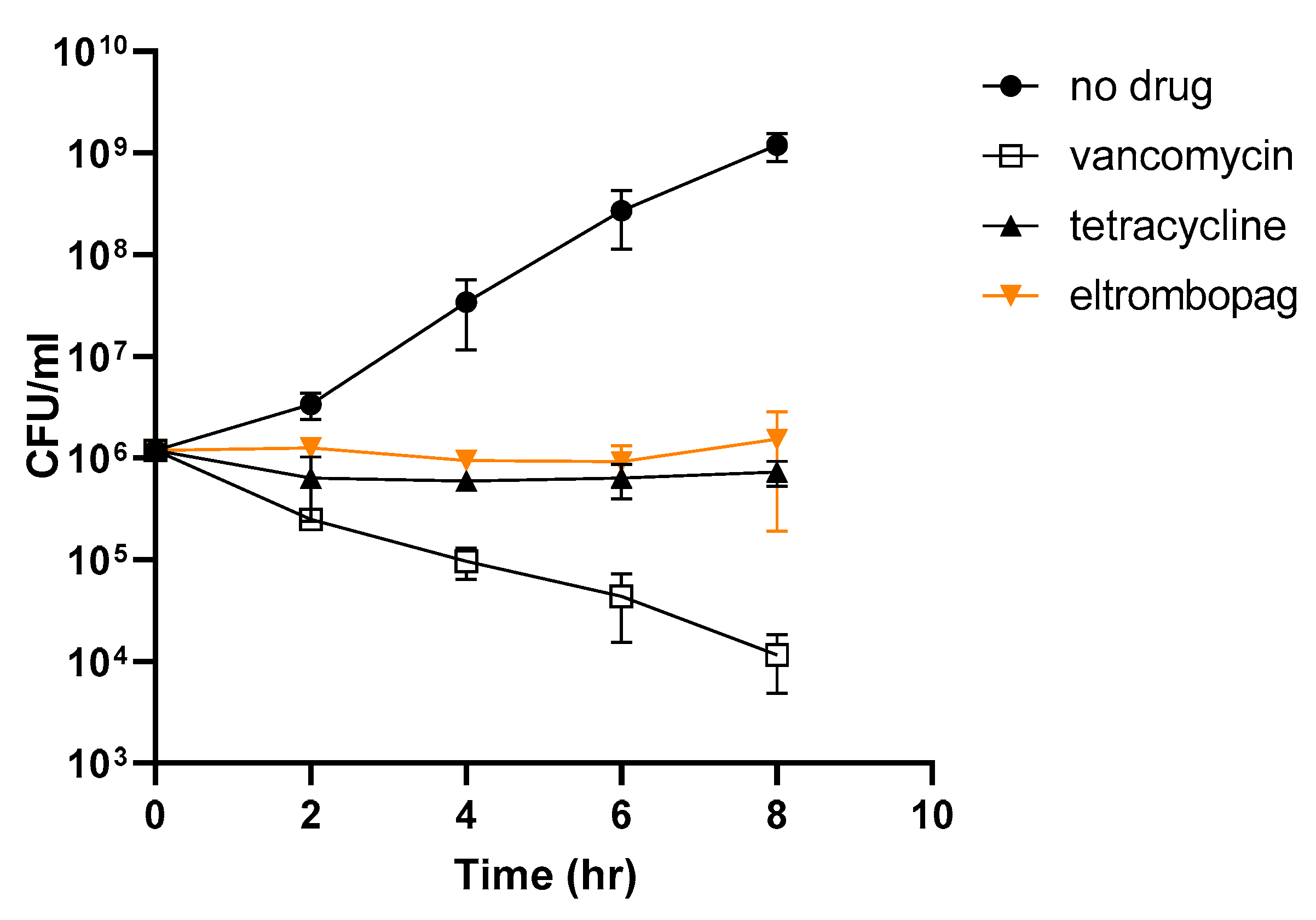
2.6. Elucidating the Antibacterial Mechanism of Eltrombopag
3. Materials and Methods
3.1. Strains and Culture Conditions
3.2. Primary Screening
3.3. Dose–Response of Selected Repositionable Candidates
3.4. Effect of Eltrombopag on S. aureus Infection of Human Caco-2 Cells
3.5. Cytotoxicity Test of Eltrombopag
3.6. Mouse Infection Model
3.7. Kinetics of Time-Dependent Bacterial Killing
3.8. Eltrombopag-Resistant Mutant Selection
3.9. Whole-Genome Sequencing Analysis
3.10. RNA Extraction and qRT-PCR
3.11. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McGuinness, W.A.; Malachowa, N.; DeLeo, F.R. Vancomycin Resistance in Staphylococcus aureus. Yale J. Biol. Med. 2017, 90, 269–281. [Google Scholar]
- Beyer, P.; Paulin, S. Priority pathogens and the antibiotic pipeline: An update. Bullet. World Health Organ. 2020, 98, 151. [Google Scholar] [CrossRef]
- Thakare, R.; Singh, A.K.; Das, S.; Vasudevan, N.; Jachak, G.R.; Reddy, D.S.; Dasgupta, A.; Chopra, S. Repurposing Ivacaftor for treatment of Staphylococcus aureus infections. Int. J. Antimicrob. Agents 2017, 50, 389–392. [Google Scholar] [CrossRef]
- Jeon, S.; Ko, M.; Lee, J.; Choi, I.; Byun, S.Y.; Park, S.; Shum, D.; Kim, S. Identification of Antiviral Drug Candidates against SARS-CoV-2 from FDA-Approved Drugs. Antimicrob. Agents Chemother. 2020, 64, e00819-20. [Google Scholar] [CrossRef]
- Ittzes, B.; Szentkiralyi, E.; Szabo, Z.; Batai, I.Z.; Gyorffy, O.; Kovacs, T.; Batai, I.; Kerenyi, M. Amiodarone that has antibacterial effect against human pathogens may represent a novel catheter lock. Acta Microbiol. Immunol. Hung. 2020, 67, 133–137. [Google Scholar]
- Chang, H.C.; Huang, Y.T.; Chen, C.S.; Chen, Y.W.; Huang, Y.T.; Su, J.C.; Teng, L.J.; Shiau, C.W.; Chiu, H.C. In vitro and in vivo activity of a novel sorafenib derivative SC5005 against MRSA. J. Antimicrob. Chemother. 2016, 71, 449–459. [Google Scholar] [CrossRef]
- Le, P.; Kunold, E.; Macsics, R.; Rox, K.; Jennings, M.C.; Ugur, I.; Reinecke, M.; Chaves-Moreno, D.; Hackl, M.W.; Fetzer, C.; et al. Repurposing human kinase inhibitors to create an antibiotic active against drug-resistant Staphylococcus aureus, persisters and biofilms. Nat. Chem. 2020, 12, 145–158. [Google Scholar] [CrossRef]
- Li, S.; Qi, X.; Huang, Y.; Liu, D.; Zhou, F.; Zhou, C. Ceritinib (LDK378): A potent alternative to crizotinib for ALK-rearranged non-small-cell lung cancer. Clin. Lung Cancer 2015, 16, 86–91. [Google Scholar] [CrossRef]
- Gonzalez-Porras, J.R.; Bastida, J.M. Eltrombopag in immune thrombocytopenia: Efficacy review and update on drug safety. Ther. Adv. Drug Saf. 2018, 9, 263–285. [Google Scholar] [CrossRef]
- Melander, R.J.; Zurawski, D.V.; Melander, C. Narrow-Spectrum Antibacterial Agents. Medchemcomm 2018, 9, 12–21. [Google Scholar] [CrossRef]
- Lane, A.B.; Copeland, N.K.; Onmus-Leone, F.; Lawler, J.V. Methicillin-Resistant Staphylococcus aureus as a Probable Cause of Antibiotic-Associated Enterocolitis. Case Rep. Infect. Dis. 2018, 2018, 3106305. [Google Scholar] [CrossRef] [Green Version]
- Acton, D.S.; Plat-Sinnige, M.J.T.; van Wamel, W.; de Groot, N.; van Belkum, A. Intestinal carriage of Staphylococcus aureus: How does its frequency compare with that of nasal carriage and what is its clinical impact? Eur. J. Clin. Microbiol. 2009, 28, 115–127. [Google Scholar] [CrossRef] [Green Version]
- Squier, C.; Rihs, J.D.; Risa, K.J.; Sagnimeni, A.; Wagener, M.M.; Stout, J.; Muder, R.R.; Singh, N. Staphylococcus aureus rectal carriage and its association with infections in patients in a surgical intensive care unit and a liver transplant unit. Infect. Cont. Hosp. Epidemiol. 2002, 23, 495–501. [Google Scholar] [CrossRef] [PubMed]
- Ray, A.J.; Pultz, N.J.; Bhalla, A.; Aron, D.C.; Donskey, C.J. Coexistence of vancomycin-resistant enterococci and Staphylococcus aureus in the intestinal tracts of hospitalized patients. Clin. Infect. Dis. 2003, 37, 875–881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Erickson-Miller, C.L.; DeLorme, E.; Tian, S.S.; Hopson, C.B.; Stark, K.; Giampa, L.; Valoret, E.I.; Duffy, K.J.; Luengo, J.L.; Rosen, J.; et al. Discovery and characterization of a selective, nonpeptidyl thrombopoietin receptor agonist. Exp. Hematol. 2005, 33, 85–93. [Google Scholar] [CrossRef] [PubMed]
- Linzner, N.; Van Loi, V.; Fritsch, V.N.; Antelmann, H. Thiol-based redox switches in the major pathogen Staphylococcus aureus. Biol. Chem. 2021, 402, 333–361. [Google Scholar] [CrossRef] [PubMed]
- Vollmer, W.; Joris, B.; Charlier, P.; Foster, S. Bacterial peptidoglycan (murein) hydrolases. FEMS Microbiol. Rev. 2008, 32, 259–286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sakamoto, J.J.; Sasaki, M.; Tsuchido, T. Purification and characterization of a Bacillus subtilis 168 nuclease, YokF, involved in chromosomal DNA degradation and cell death caused by thermal shock treatments. J. Biol. Chem. 2001, 276, 47046–47051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically, Approved Standard, 9th ed.; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2012.
- Ferruzza, S.; Rossi, C.; Scarino, M.L.; Sambuy, Y. A protocol for differentiation of human intestinal Caco-2 cells in asymmetric serum-containing medium. Toxicol. In Vitro 2012, 26, 1252–1255. [Google Scholar] [CrossRef]
- Mariadason, J.M.; Arango, D.; Corner, G.A.; Aranes, M.J.; Hotchkiss, K.A.; Yang, W.C.; Augenlicht, L.H. A gene expression profile that defines colon cell maturation in vitro. Cancer Res. 2002, 62, 4791–4804. [Google Scholar] [PubMed]
- Sambuy, Y.; Angelis, I.; Ranaldi, G.; Scarino, M.L.; Stammati, A.; Zucco, F. The Caco-2 cell line as a model of the intestinal barrier: Influence of cell and culture-related factors on Caco-2 cell functional characteristics. Cell Biol. Toxicol. 2005, 21, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Ibberson, C.B.; Parlet, C.P.; Kwiecinski, J.; Crosby, H.A.; Meyerholz, D.K.; Horswill, A.R. Hyaluronan Modulation Impacts Staphylococcus aureus Biofilm Infection. Infect. Immun. 2016, 84, 1917–1929. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Name | Compound Structure | % Inhibition ± SD a | Class b |
|---|---|---|---|
| Dronedarone HCl | 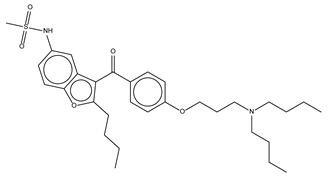 | 59.2 ± 0.4 | Antiarrhythmic |
| Ceritinib | 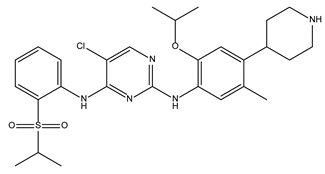 | 81.8 ± 2.9 | Kinase inhibitor |
| Eltrombopag | 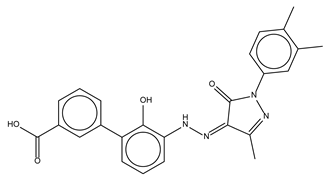 | 100.8 ± 0.5 | Thrombopoietin receptor agonist |
| Ivacaftor | 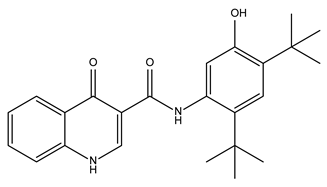 | 104.8 ± 0.4 | Cystic fibrosis transmembrane conductance regulator (CFTR) potentiator |
| Tedizolid (phosphate) | 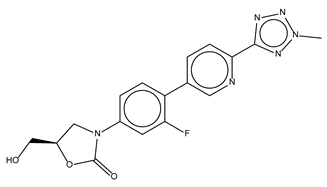 | 52.6 ± 2.6 | Antibiotic |
| Fosfomycin sodium | 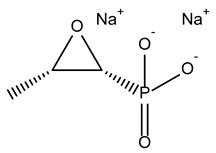 | 53.9 ± 0.1 | Antibiotic |
| Rosoxacin | 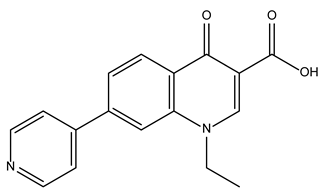 | 92.2 ± 0.6 | Antibiotic |
| Fidaxomicin | 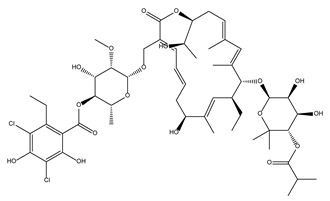 | 99.0 ± 0.1 | Antibiotic |
| Retapamulin | 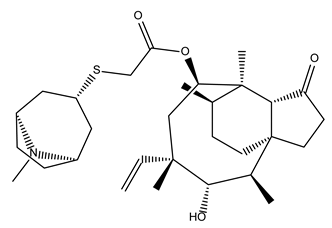 | 97.5 ± 1.4 | Antibiotic |
| Cefditoren Pivoxil | 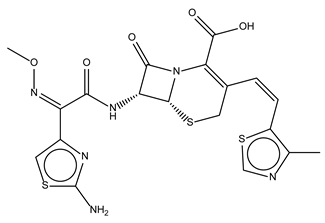 | 102.6 ± 0.1 | Antibiotic |
| Ticarcillin sodium |  | 103.0 ± 1.2 | Antibiotic |
| Cefpodoxime (free acid) |  | 103.1 ± 0.6 | Antibiotic |
| Gene ID | Gene Name | Mutation |
|---|---|---|
| SASA1_01682 | yedL (pmtR) | Point mutation (P37L) |
| SASA1_01628 | walR | Point mutation (E11G or D83G) |
| SASA1_01072 | yjbH (spxH) | Truncation |
| SASA1_00966 | lytE | Truncation |
| SASA1_02364 | yokF | Truncation |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, H.; Lee, J.; Hwang, J.; Park, S.; Kim, N.; Kim, K.; Lee, H.; Shum, D.; Jang, S. Repurposing Eltrombopag for Multidrug Resistant Staphylococcus aureus Infections. Antibiotics 2021, 10, 1372. https://doi.org/10.3390/antibiotics10111372
Lee H, Lee J, Hwang J, Park S, Kim N, Kim K, Lee H, Shum D, Jang S. Repurposing Eltrombopag for Multidrug Resistant Staphylococcus aureus Infections. Antibiotics. 2021; 10(11):1372. https://doi.org/10.3390/antibiotics10111372
Chicago/Turabian StyleLee, Hyunjung, Jaehoan Lee, Juchan Hwang, Sinyoung Park, Namyoul Kim, Kideok Kim, Honggun Lee, David Shum, and Soojin Jang. 2021. "Repurposing Eltrombopag for Multidrug Resistant Staphylococcus aureus Infections" Antibiotics 10, no. 11: 1372. https://doi.org/10.3390/antibiotics10111372
APA StyleLee, H., Lee, J., Hwang, J., Park, S., Kim, N., Kim, K., Lee, H., Shum, D., & Jang, S. (2021). Repurposing Eltrombopag for Multidrug Resistant Staphylococcus aureus Infections. Antibiotics, 10(11), 1372. https://doi.org/10.3390/antibiotics10111372