
Antimicrobial Resistance of Listeria monocytogenes Strains Isolated from Humans, Animals, and Food Products in Russia in 1950–1980, 2000–2005, and 2018–2021

 , , ,
, , ,  and
and
Abstract
:1. Introduction
2. Results
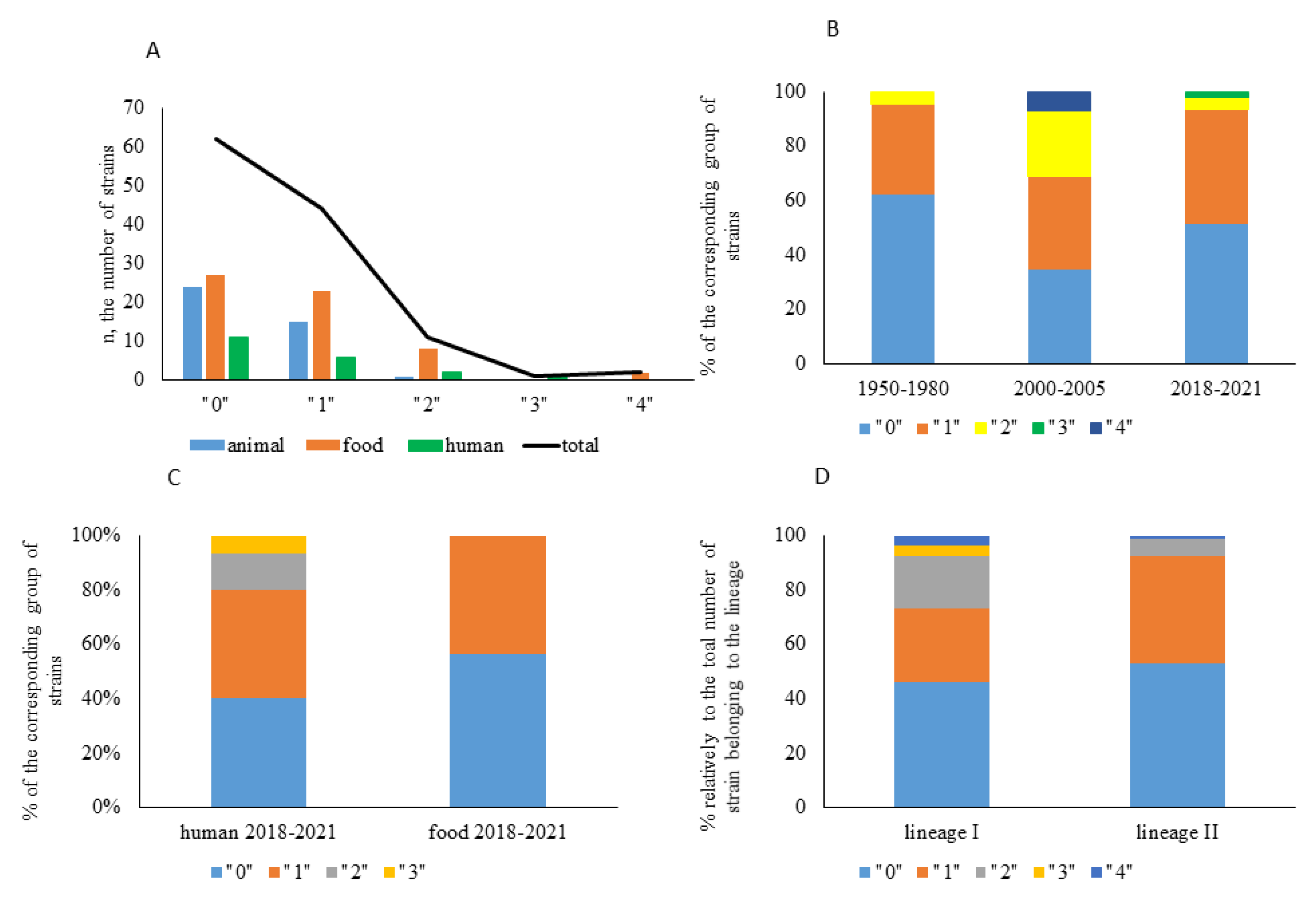
2.1. Strain Characterization
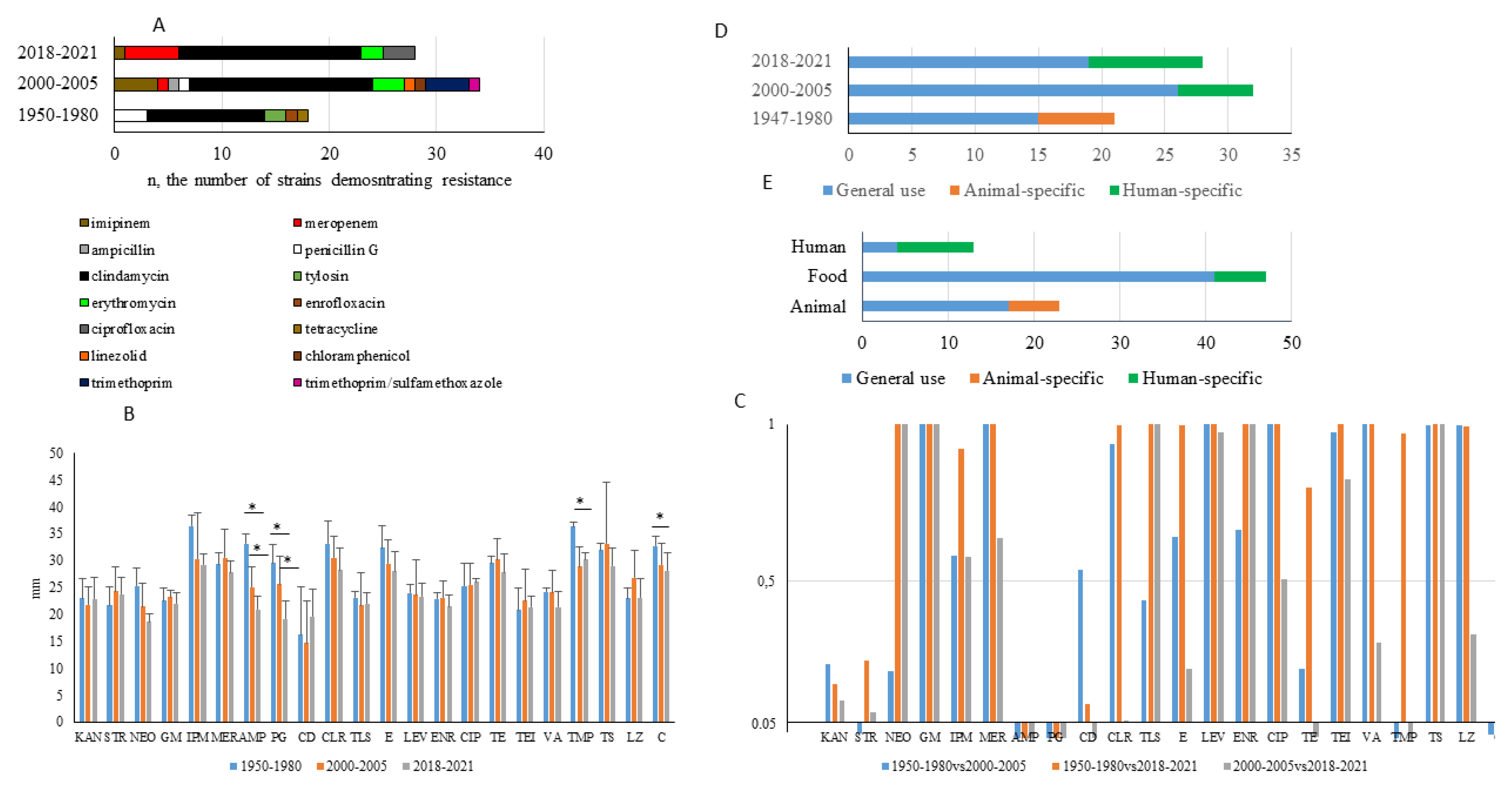
2.2. Frequency of Antibiotic Resistance among Strains Tested
2.3. Characterization of Antibiotic Resistance
2.4. Temporal Changes in Antibiotic Resistance Patterns
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Characterization Methods
4.2. Antibiotic Susceptibility Testing
4.3. Antibiotic Resistance Index (ARI)
4.4. Statistics
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Allerberger, F.; Wagner, M. Listeriosis: A resurgent foodborne infection. Clin. Microbiol. Infect. 2010, 16, 16–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Noordhout, C.M.; Devleesschauwer, B.; Angulo, F.J.; Verbeke, G.; Haagsma, J.; Kirk, M.; Havelaar, A.; Speybroeck, N. The global burden of listeriosis: A systematic review and meta-analysis. Lancet Infect. Dis. 2014, 14, 1073–1082. [Google Scholar] [CrossRef] [Green Version]
- Van Walle, I.; Björkman, J.; Cormican, M.; Dallman, T.; Mossong, J.; Moura, A.; Pietzka, A.; Ruppitsch, W.; Takkinen, J. Retrospective validation of whole genome sequencing-enhanced surveillance of listeriosis in Europe, 2010 to 2015. Euro Surveill. 2018, 23, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McLauchlin, J.; Grant, K.; Amar, C. Human foodborne listeriosis in England and Wales, 1981 to 2015. Epidemiol. Infect. 2020, 148, e54. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamont, R.; Sobel, J.; Mazaki-Tovi, S.; Kusanovic, J.; Vaisbuch, E.; Kim, S.; Uldbjerg, N.; Romero, R. Listeriosis in human pregnancy: A systematic review. J. Perinat. Med. 2011, 39, 227–236. [Google Scholar] [CrossRef]
- Charlier, C.; Kermorvant-Duchemin, E.; Perrodeau, E.; Moura, A.; Maury, M.; Bracq-Dieye, H.; Thouvenot, P.; Valès, G.; Leclercq, A.; Ravaud, P.; et al. Neonatal listeriosis presentation and outcome: A prospective study of 189 cases. Clin. Infect. Dis. 2021. [CrossRef]
- Filipello, V.; Amato, E.; Gori, M.; Huedo, P.; Ciceri, G.; Lomonaco, S.; Pontello, M. Epidemiology and Molecular Typing of Pregnancy-Associated Listeriosis Cases in Lombardy, Italy, over a 10-Year Period (2005–2014). Infect. Dis. Obstet. Gynecol. 2017, 2017, 6479121. [Google Scholar] [CrossRef] [Green Version]
- Mylonakis, E.; Hohmann, E.; Calderwood, S. Central nervous system infection with Listeria monocytogenes. 33 years’ experience at a general hospital and review of 776 episodes from the literature. Medicine 1998, 77, 313–336. [Google Scholar] [CrossRef]
- Pires, S.M.; Desta, B.N.; Mughini-Gras, L.; Mmbaga, B.T.; Fayemi, O.E.; Salvador, E.M.; Gobena, T.; Majowicz, S.E.; Hald, T.; Hoejskov, P.S.; et al. Burden of foodborne diseases: Think global, act local. Curr. Opin. Food Sci. 2021, 39, 152–159. [Google Scholar] [CrossRef]
- Hald, T.; Aspinall, W.; Devleesschauwer, B.; Cooke, R.; Corrigan, T.; Havelaar, A.; Gibb, H.; Torgerson, P.; Kirk, M.; Angulo, F.J.; et al. World Health Organization Estimates of the Relative Contributions of Food to the Burden of Disease Due to Selected Foodborne Hazards: A Structured Expert Elicitation. PLoS ONE 2016, 11, e0145839. [Google Scholar] [CrossRef] [Green Version]
- Orsi, R.H.; den Bakker, H.C.; Wiedmann, M. Listeria monocytogenes lineages: Genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 2011, 301, 79–96. [Google Scholar] [CrossRef]
- Hain, T.; Ghai, R.; Billion, A.; Kuenne, C.T.; Steinweg, C.; Izar, B.; Mohamed, W.; Mraheil, M.A.; Domann, E.; Schaffrath, S.; et al. Comparative genomics and transcriptomics of lineages I, II, and III strains of Listeria monocytogenes. BMC Genomics 2012, 13, 144. [Google Scholar] [CrossRef] [Green Version]
- Oevermann, A.; Zurbriggen, A.; Vandevelde, M. Rhombencephalitis Caused by Listeria monocytogenes in Humans and Ruminants: A Zoonosis on the Rise? Interdiscip. Perspect. Infect. Dis. 2010, 2010. [Google Scholar] [CrossRef] [Green Version]
- Dell’Armelina Rocha, P.R.; Lomonaco, S.; Bottero, M.T.; Dalmasso, A.; Dondo, A.; Grattarola, C.; Zuccon, F.; Iulini, B.; Knabel, S.J.; Capucchio, M.T.; et al. Ruminant rhombencephalitis-associated Listeria monocytogenes strains constitute a genetically homogeneous group related to human outbreak strains. Appl. Environ. Microbiol. 2013, 79, 3059–3066. [Google Scholar] [CrossRef] [Green Version]
- Voronina, O.; Ryzhova, N.; Kunda, M.; Kurnaeva, M.; Semenov, A.; Aksenova, E.; Egorova, I.Y.; Kolbasov, D.; Ermolaeva, S.; Gintsburg, A.L. Diversity and Pathogenic Potential of Listeria monocytogenes Isolated from Environmental Sources in the Russian Federation. Int. J. Mod. Emg. Res. 2015, 5, 5–13. [Google Scholar]
- Psareva, E.; Egorova, I.; Liskova, E.; Razheva, I.; Gladkova, N.; Sokolova, E.; Potemkin, E.; Zhurilov, P.; Mikhaleva, T.; Blokhin, A.; et al. Retrospective Study of Listeria Monocytogenes Isolated in the Territory of Inner Eurasia from 1947 to 1999. Pathogens 2019, 8, 184. [Google Scholar] [CrossRef] [Green Version]
- Voronina, O.; Kunda, M.; RyzhYushchukova, N.; Kutuzova, A.; Aksenova, E.; Karpova, T.; Tartakovskij, I.; Yushchuk, N.; Klimova, E.; Karetkina, G.; et al. Listeriosis: Genotyping as a key for identification a possible source of infection. Clin. Microbiol. Antimicrob. Chemother. 2019, 21, 261–273. [Google Scholar] [CrossRef]
- Voronina, O.; Tartakovsky, I.; Yuyshchuk, N.; Ryzhova, N.; Kunda, M.; Aksenova, E.; Kutuzova, A.; Melkumyan, A.; Karpova, T.; Gruzdeva, O.; et al. Analysis of sporadic cases of invasivelisteriosis in a metropolis. J. Microbiol. Epidemiol. Immunobiol. 2020, 97, 547–555. [Google Scholar] [CrossRef]
- Scortti, M.; Lacharme-Lora, L.; Wagner, M.; Chico-Calero, I.; Losito, P.; Vázquez-Boland, J.A. Coexpression of virulence and fosfomycin susceptibility in Listeria: Molecular basis of an antimicrobial in vitro-in vivo paradox. Nat. Med. 2006, 12, 515–517. [Google Scholar] [CrossRef]
- Troxler, R.; von Graevenitz, A.; Funke, G.; Wiedemann, B.; Stock, I. Natural antibiotic susceptibility of Listeria species: L. grayi, L. innocua, L. ivanovii, L. monocytogenes, L. seeligeri and L. welshimeri strains. Clin. Microbiol. Infect. 2000, 6, 525–535. [Google Scholar] [CrossRef] [Green Version]
- Temple, M.; Nahata, M. Treatment of listeriosis. Ann. Pharmacother. 2000, 34, 656–661. [Google Scholar] [CrossRef]
- Pagliano, P.; Arslan, F.; Ascione, T. Epidemiology and treatment of the commonest form of listeriosis: Meningitis and bacteraemia. Le Infez. Med. 2017, 25, 210–216. [Google Scholar]
- Morvan, A.; Moubareck, C.; Leclercq, A.; Hervé-Bazin, M.; Bremont, S.; Lecuit, M.; Courvalin, P.; Le Monnier, A. Antimicrobial resistance of Listeria monocytogenes strains isolated from humans in France. Antimicrob. Agents Chemother. 2010, 54, 2728–2731. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noll, M.; Kleta, S.; Al Dahouk, S. Antibiotic susceptibility of 259 Listeria monocytogenes strains isolated from food, food-processing plants and human samples in Germany. J. Infect. Public Health 2018, 11, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Bakulov, I. Listeriosis of animals: Problems and research. Acta Microbiol. Hung. 1989, 36, 145–147. [Google Scholar] [PubMed]
- Seeliger, H. Listeriosis--history and actual developments. Infection 1988, 16 (Suppl. 2), S80–S84. [Google Scholar] [CrossRef] [PubMed]
- State standard of the Russian Federation food products. Methods for the detection and determination of bacteria Listeria monocytogenes. GOST R 51921-2002. 2002. Available online: https://internet-law.ru/gosts/gost/2375/ (accessed on 26 April 2021).
- Krumperman, P. Multiple antibiotic resistance indexing of Escherichia coli to identify high-risk sources of fecal contamination of foods. Appl. Environ. Microbiol. 1983, 46, 165–170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tehrani, A.H.; Gilbride, K.A. A closer look at the antibiotic-resistant bacterial community found in urban wastewater treatment systems. Microbiologyopen 2018, 7, e00589. [Google Scholar] [CrossRef]
- Bauditz, R. Results of clinical studies with Baytril in calves and pigs. Vet. Med. Rev. 1987, 2, 122–129. [Google Scholar]
- Powers, J.H. Antimicrobial drug development – the past, the present, and the future. Clin. Microbiol. Infect. 2004, 10, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Tahoun, A.; Abou Elez, R.; Abdelfatah, E.; Elsohaby, I.; El-Gedawy, A.; Elmoslemany, A. Listeria monocytogenes in raw milk, milking equipment and dairy workers: Molecular characterization and antimicrobial resistance patterns. J. Glob. Antimicrob. Resist. 2017, 10, 264–270. [Google Scholar] [CrossRef]
- Maia, D.; Haubert, L.; Würfel, S.; Kroning, I.; Cardoso, M.; Lopes, G.; Fiorentini, A.; da Silva, W. Listeria monocytogenes in sliced cheese and ham from retail markets in southern Brazil. FEMS Microbiol. Lett. 2019, 366. [Google Scholar] [CrossRef]
- Escolar, C.; Gómez, D.; Del Carmen Rota García, M.; Conchello, P.; Herrera, A. Antimicrobial Resistance Profiles of Listeria monocytogenes and Listeria innocua Isolated from Ready-to-Eat Products of Animal Origin in Spain. Foodborne Pathog. Dis. 2017, 14, 357–363. [Google Scholar] [CrossRef]
- Jones, E.; MacGowan, A. Antimicrobial chemotherapy of human infection due to Listeria monocytogenes. Eur. J. Clin. Microbiol. Infect. Dis. 1995, 14, 165–175. [Google Scholar] [CrossRef]
- Fischer, M.; Wamp, S.; Fruth, A.; Allerberger, F.; Flieger, A.; Halbedel, S. Population structure-guided profiling of antibiotic resistance patterns in clinical Listeria monocytogenes isolates from Germany identifies pbpB3 alleles associated with low levels of cephalosporin resistance. Emerg. Microbes Infect. 2020, 9, 1804–1813. [Google Scholar] [CrossRef]
- Capita, R.; Felices-Mercado, A.; García-Fernández, C.; Alonso-Calleja, C. Characterization of Listeria Monocytogenes Originating from the Spanish Meat-Processing Chain. Foods 2019, 8, 542. [Google Scholar] [CrossRef] [Green Version]
- Rezai, R.; Ahmadi, E.; Salimi, B. Prevalence and Antimicrobial Resistance Profile of Listeria Species Isolated from Farmed and On-Sale Rainbow Trout (Oncorhynchus mykiss) in Western Iran. J. Food Prot. 2018, 81, 886–891. [Google Scholar] [CrossRef]
- Maćkiw, E.; Stasiak, M.; Kowalska, J.; Kucharek, K.; Korsak, D.; Postupolski, J. Occurrence and Characteristics of Listeria monocytogenes in Ready-to-Eat Meat Products in Poland. J. Food Prot. 2020, 83, 1002–1009. [Google Scholar] [CrossRef]
- Meletis, G. Carbapenem resistance: Overview of the problem and future perspectives. Ther. Adv. Infect. Dis. 2016, 3, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Hadorn, K.; Hächler, H.; Schaffner, A.; Kayser, F. Genetic characterization of plasmid-encoded multiple antibiotic resistance in a strain of Listeria monocytogenes causing endocarditis. Eur. J. Clin. Microbiol. Infect. Dis. 1993, 12, 928–937. [Google Scholar] [CrossRef]
- Tsakris, A.; Papa, A.; Douboyas, J.; Antoniadis, A. Neonatal meningitis due to multi-resistant Listeria monocytogenes. J. Antimicrob. Chemother. 1997, 39, 553–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pesavento, G.; Ducci, B.; Nieri, D.; Comodo, N.; Lo Nostro, A. Prevalence and antibiotic susceptibility of Listeria spp. isolated from raw meat and retail foods. Food Control 2010, 21, 708–713. [Google Scholar] [CrossRef]
- Iwu, C.; Okoh, A. Characterization of antibiogram fingerprints in Listeria monocytogenes recovered from irrigation water and agricultural soil samples. PLoS One 2020, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keet, R.; Rip, D. Listeria monocytogenes isolates from Western Cape, South Africa exhibit resistance to multiple antibiotics and contradicts certain global resistance patterns. AIMS Microbiol. 2021, 7, 40–58. [Google Scholar] [CrossRef]
- European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters; Version 7.0; European Committee on Antimicrobial Susceptibility Testing: Växjö, Sweden, 2017. [Google Scholar]
- CLSI. Performance Standards for Antimicrobial Disk Susceptibility Tests, M100S, 29th ed.; CLSI: Wayne, PA, USA, 2019; Volume 39. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Period of Isolation | Source of the Strain | Phylogenetic Position | All Collection | |||
|---|---|---|---|---|---|---|
| Human | Food | Animal | Lineage I | Lineage II | ||
| 1950–1980 | 0.016 | n.a. | 0.019 | 0 | 0 | 0.017 |
| 2000–2005 | 0.055 | 0.051 | n.a. | 0.043 | 0.051 | 0.039 |
| 2018–2021 | 0.024 | 0.019 | n.a. | 0.052 | 0.020 | 0.027 |
| total | 0.031 | 0.039 | 0.019 | 0.043 | 0.056 | 0.031 |
| Class | Antibiotic | Total Number of Resistant Strains | Number of Strains with Resistance to Only One Antibiotic |
|---|---|---|---|
| Aminoglycosides | |||
| Gentamicin | 0 | 0 | |
| Kanamycin | 0 | 0 | |
| Neomycin | 0 | 0 | |
| Streptomycin | 0 | 0 | |
| β-Lactams:Penicillins | |||
| Penicillin G | 4 | 2 | |
| Ampicillin | 1 | 0 | |
| Amoxicillin/clavulanic acid | 0 | 0 | |
| β-Lactams:Carbapenems | |||
| Imipenem | 5 | 1 | |
| Meropenem | 6 | 2 | |
| Macrolides | |||
| Clarithromycin | 0 | 0 | |
| Tylosin | 2 | 2 | |
| Erythromycin | 5 | 3 | |
| Lincosamides | |||
| Clindamycin | 45 | 33 | |
| Quinolones | |||
| Ciprofloxacin | 3 | 1 | |
| Levofloxacin | 0 | 0 | |
| Enrofloxacin | 1 | 0 | |
| Glycopeptides | |||
| Teicoplanin | 0 | 0 | |
| Vancomycin | 0 | 0 | |
| Other antibiotics | |||
| Trimethoprim | 4 | 0 | |
| Trimethoprim/sulfamethoxazole | 1 | 0 | |
| Linezolid | 1 | 0 | |
| Chloramphenicol | 1 | 0 | |
| Tetracycline | 1 | 0 |
| N | Strain | Resistance | CC/Lineage | Year | Source |
|---|---|---|---|---|---|
| 1 | 178-P | PG -ENR | CC7/II | 1967 | pig |
| 2 | 3880 | CD-TE | CC7/II | 1970 | pig |
| 3 | 24-T | CD-IPM | CC37/II | 2005 | dairy product |
| 4 | 14-2 | CD-IPM | CC6/I | 2001 | fish |
| 5 | 35-T | CD-TR | CC37/II | 2005 | dairy product |
| 6 | 134/3 | CD-TR | CC2/I | 2005 | dairy product |
| 7 | 1300 | CD-PG | CC1/I | 2005 | dairy product |
| 8 | 44 | CD-AMP | CC7/I | 2002 | meat |
| 9 | 98/20 | CD-LZ | CC9/II | 2005 | dairy product |
| 10 | 13215 | CD-E | CC59/I | 2005 | dairy product |
| 11 | UH18 | CD-CIP | CC155/II | 2019 | perinatal, blood |
| 12 | H67-1 | IMP-MER | CC6/I | 2019 | clinical, adult |
| 13 | UH19 | CD–MER-CIP | CC6/I | 2019 | perinatal, blood |
| 14 | 129/3 | CD–MER–IPM-TMP | CC9/II | 2005 | dairy products |
| 15 | 114/31 | CD–E–C-TMP | CC8/II | 2005 | dairy products |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Andriyanov, P.A.; Zhurilov, P.A.; Liskova, E.A.; Karpova, T.I.; Sokolova, E.V.; Yushina, Y.K.; Zaiko, E.V.; Bataeva, D.S.; Voronina, O.L.; Psareva, E.K.; et al. Antimicrobial Resistance of Listeria monocytogenes Strains Isolated from Humans, Animals, and Food Products in Russia in 1950–1980, 2000–2005, and 2018–2021. Antibiotics 2021, 10, 1206. https://doi.org/10.3390/antibiotics10101206
Andriyanov PA, Zhurilov PA, Liskova EA, Karpova TI, Sokolova EV, Yushina YK, Zaiko EV, Bataeva DS, Voronina OL, Psareva EK, et al. Antimicrobial Resistance of Listeria monocytogenes Strains Isolated from Humans, Animals, and Food Products in Russia in 1950–1980, 2000–2005, and 2018–2021. Antibiotics. 2021; 10(10):1206. https://doi.org/10.3390/antibiotics10101206
Chicago/Turabian StyleAndriyanov, Pavel A., Pavel A. Zhurilov, Elena A. Liskova, Tatyana I. Karpova, Elena V. Sokolova, Yulia K. Yushina, Elena V. Zaiko, Dagmara S. Bataeva, Olga L. Voronina, Ekaterina K. Psareva, and et al. 2021. "Antimicrobial Resistance of Listeria monocytogenes Strains Isolated from Humans, Animals, and Food Products in Russia in 1950–1980, 2000–2005, and 2018–2021" Antibiotics 10, no. 10: 1206. https://doi.org/10.3390/antibiotics10101206
APA StyleAndriyanov, P. A., Zhurilov, P. A., Liskova, E. A., Karpova, T. I., Sokolova, E. V., Yushina, Y. K., Zaiko, E. V., Bataeva, D. S., Voronina, O. L., Psareva, E. K., Tartakovsky, I. S., Kolbasov, D. V., & Ermolaeva, S. A. (2021). Antimicrobial Resistance of Listeria monocytogenes Strains Isolated from Humans, Animals, and Food Products in Russia in 1950–1980, 2000–2005, and 2018–2021. Antibiotics, 10(10), 1206. https://doi.org/10.3390/antibiotics10101206