A Luciferase Mutant with Improved Brightness and Stability for Whole-Cell Bioluminescent Biosensors and In Vitro Biosensing

 , , and
, , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Plasmid Construction
2.3. Expression and Purification of Luciferase Mutants
2.4. Heat Inactivation Studies
2.5. Luciferase Emission Spectra, Thermal and pH Stability Studies
2.6. Determination of Kinetic Parameters and ATP Detection
2.7. Whole-Cell Biosensor for Inflammation Activity
3. Result and Discussion
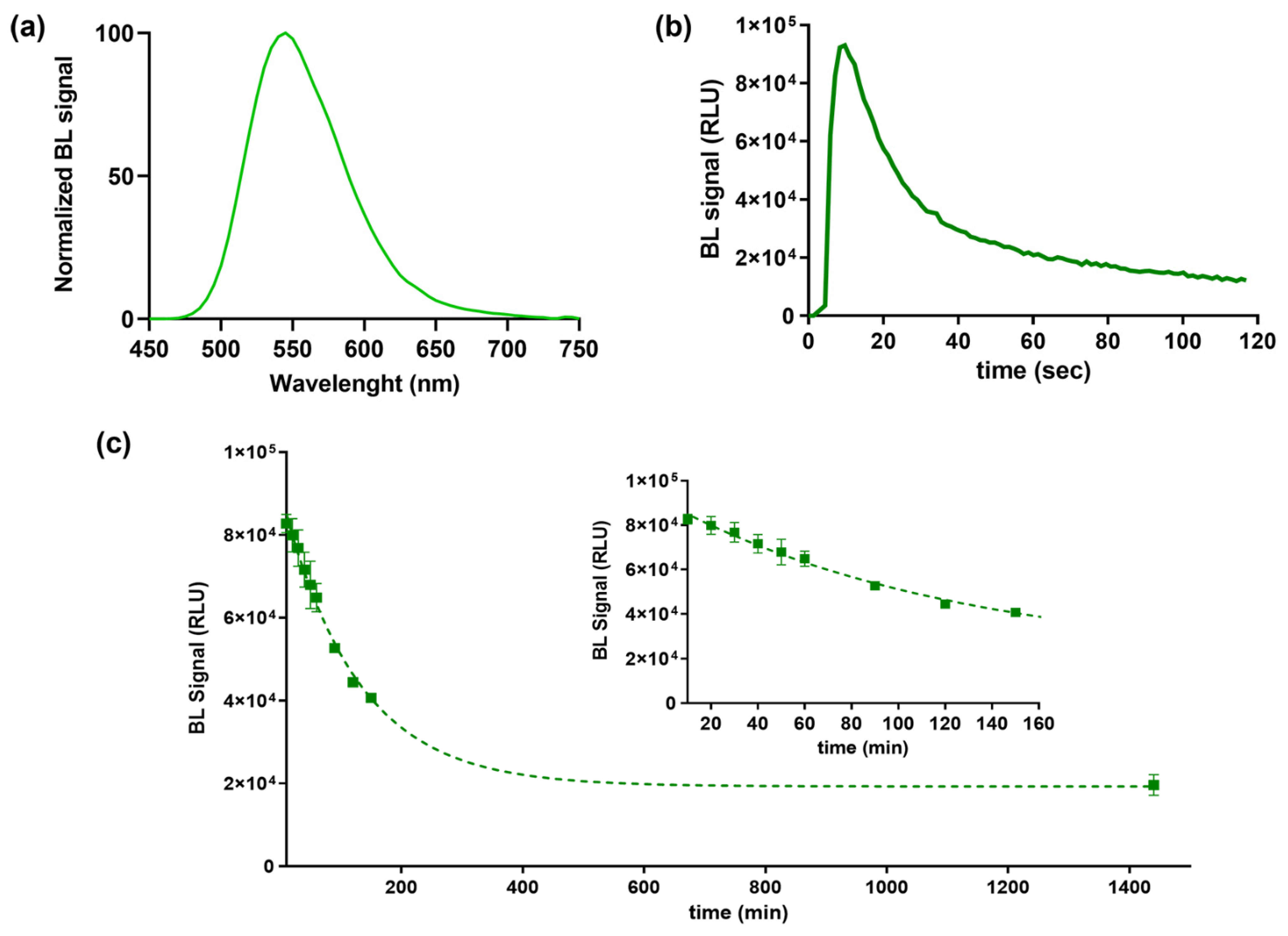
3.1. Luciferase Characterization: Bioluminescence Emission Kinetics and Spectra
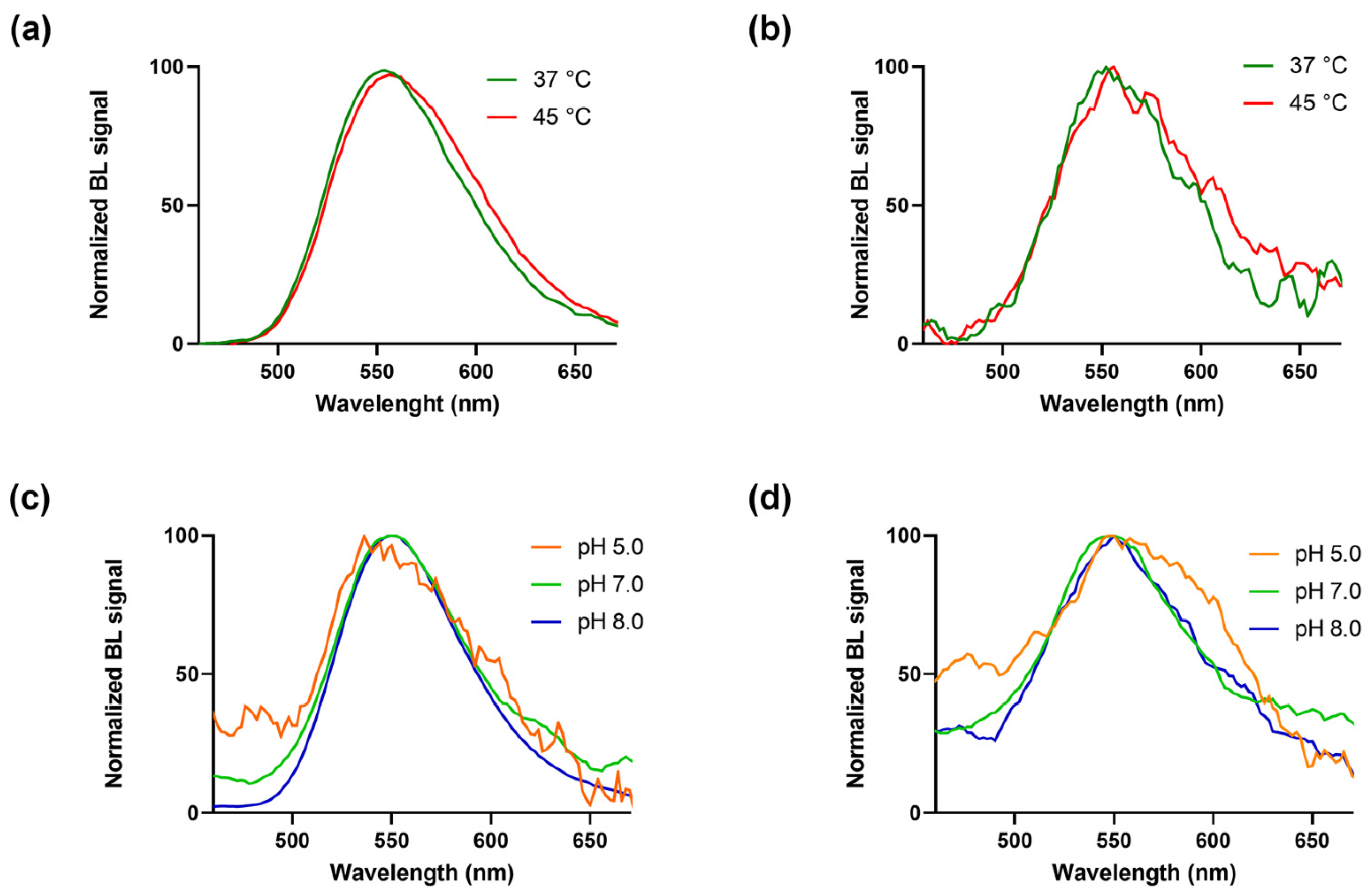
3.2. Thermal and pH Stability Measurements of BgLuc
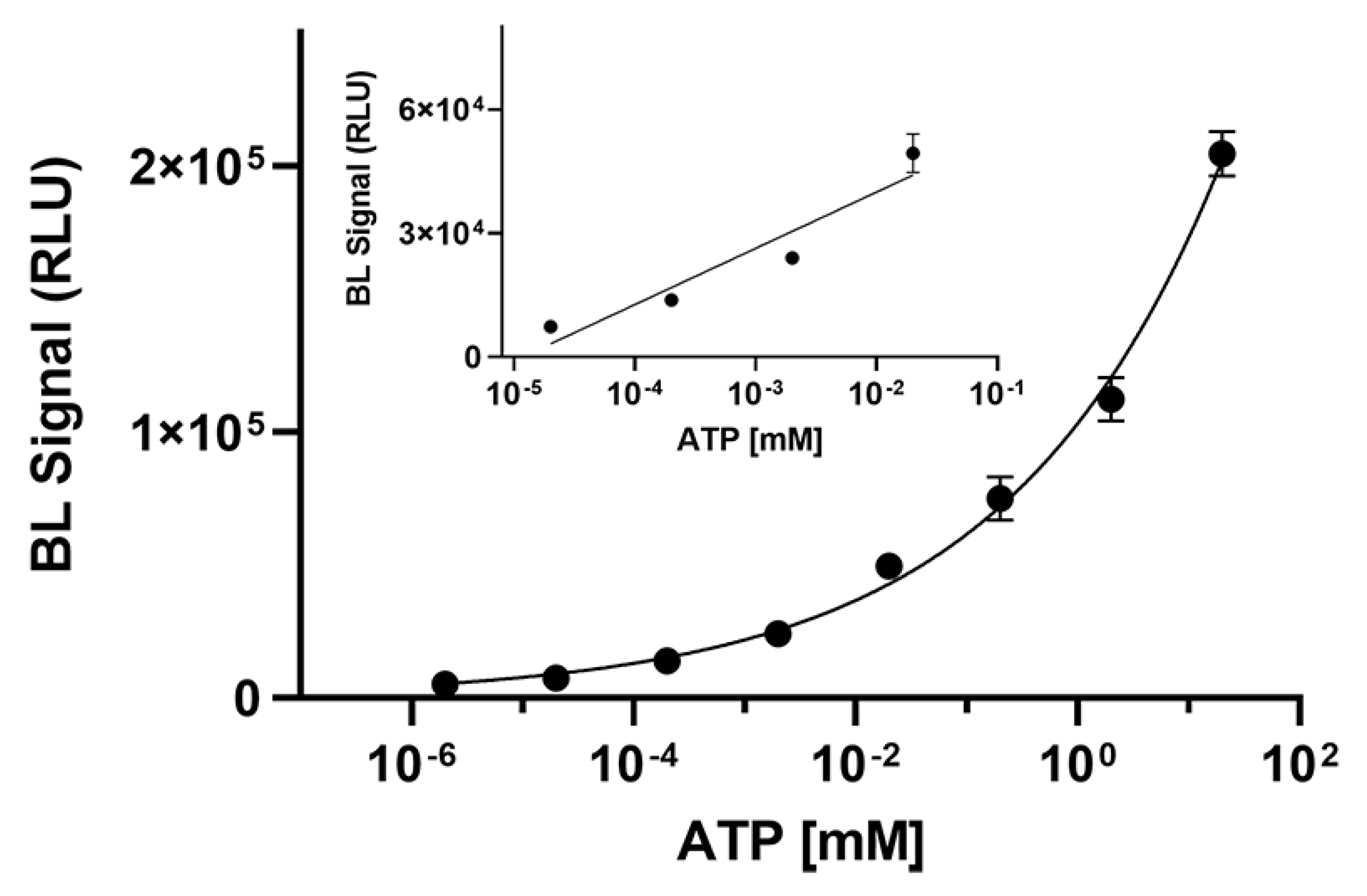
3.3. Measurement of Kinetic Parameters and ATP Detection
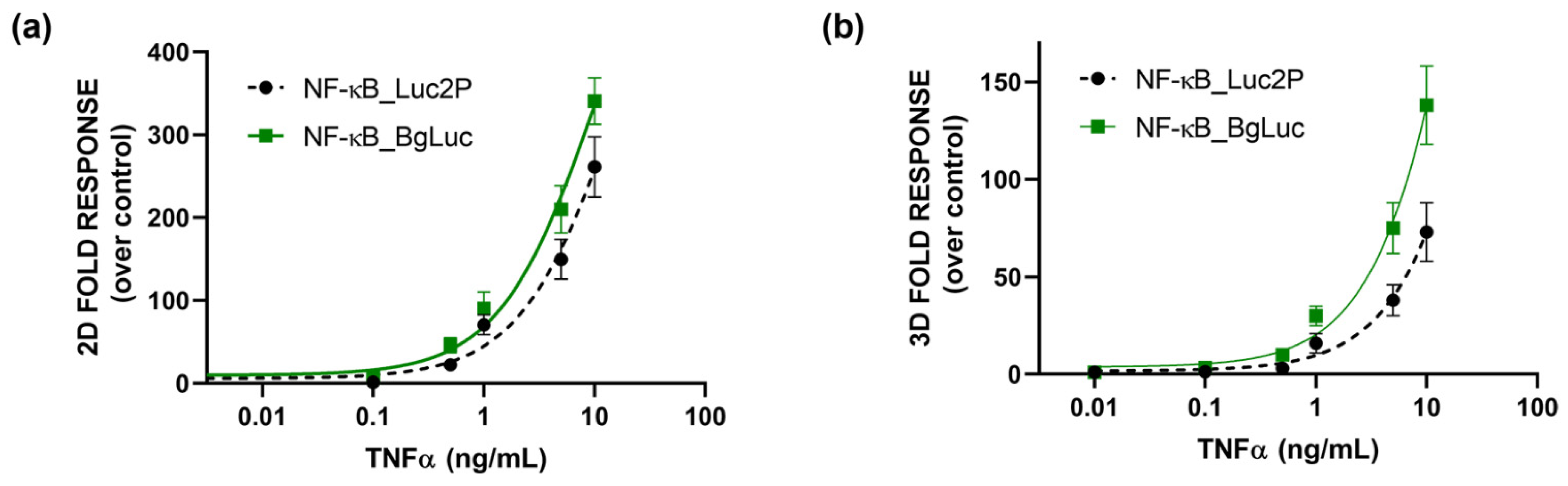
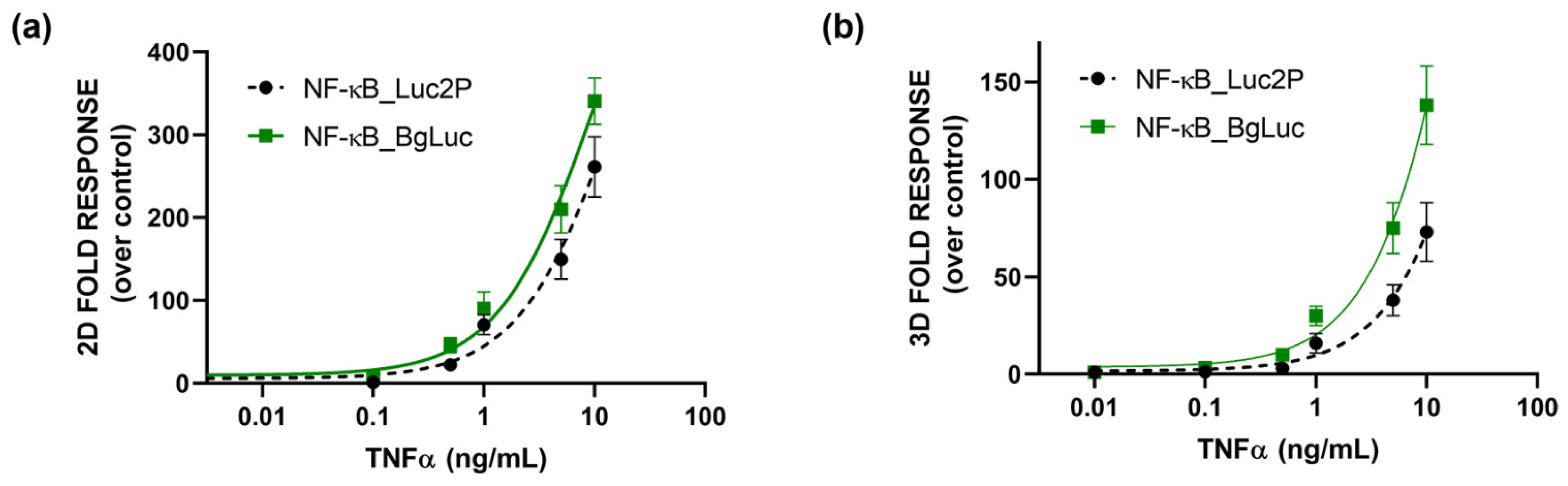
3.4. Whole-Cell Biosensor for Inflammation Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oba, Y.; Schultz, D.T. Firefly genomes illuminate the evolution of beetle bioluminescent systems. Curr. Opin. Insect Sci. 2022, 50, 100879. [Google Scholar] [CrossRef] [PubMed]
- Viviani, V.R. The origin, diversity, and structure function relationships of insect luciferases. Cell. Mol. Life Sci. 2002, 59, 1833–1850. [Google Scholar] [CrossRef] [PubMed]
- Viviani, V.R.; Uchida, A.; Viviani, W.; Ohmiya, Y. The Influence of Ala243 (Gly247), Arg215 and Thr226 (Asn230) on the Bioluminescence Spectra and pH-Sensitivity of Railroad Worm, Click Beetle and Firefly Luciferases. Photochem. Photobiol. 2002, 76, 538. [Google Scholar] [CrossRef]
- Sadikot, R.T.; Blackwell, T.S. Bioluminescence imaging. Proc. Am. Thorac. Soc. 2005, 2, 537–540. [Google Scholar] [CrossRef]
- Rumyantsev, K.A.; Turoverov, K.K.; Verkhusha, V.V. Near-infrared bioluminescent proteins for two-color multimodal imaging. Sci. Rep. 2016, 6, 36588. [Google Scholar] [CrossRef] [PubMed]
- Lundin, A. Optimized Assay of Firefly Luciferase with Stable Light Emission; Szalay, A., Kricka, L.J.S.P., Eds.; John Wiley and Sons: Chichester, UK, 1993; ISBN 9781119130536. [Google Scholar]
- de Wet, J.R.; Wood, K.V.; Helinski, D.R.; DeLuca, M. Cloning of firefly luciferase cDNA and the expression of active luciferase in Escherichia coli. Proc. Natl. Acad. Sci. USA 1985, 82, 7870–7873. [Google Scholar] [CrossRef] [PubMed]
- Branchini, B.R.; Behney, C.E.; Southworth, T.L.; Fontaine, D.M.; Gulick, A.M.; Vinyard, D.J.; Brudvig, G.W. Experimental Support for a Single Electron-Transfer Oxidation Mechanism in Firefly Bioluminescence. J. Am. Chem. Soc. 2015, 137, 7592–7595. [Google Scholar] [CrossRef]
- Branchini, B.R.; Ablamsky, D.M.; Murtiashaw, M.H.; Uzasci, L.; Fraga, H.; Southworth, T.L. Thermostable red and green light-producing firefly luciferase mutants for bioluminescent reporter applications. Anal. Biochem. 2007, 361, 253–262. [Google Scholar] [CrossRef]
- Carvalho, M.C.; Tomazini, A.; Amaral, D.T.; Murakami, M.T.; Viviani, V.R. Luciferase isozymes from the Brazilian Aspisoma lineatum (Lampyridae) firefly: Origin of efficient pH-sensitive lantern luciferases from fat body pH-insensitive ancestors. Photochem. Photobiol. Sci. 2020, 19, 1750–1764. [Google Scholar] [CrossRef]
- Viviani, V.R.; Silva, J.R.; Amaral, D.T.; Bevilaqua, V.R.; Abdalla, F.C.; Branchini, B.R.; Johnson, C.H. A new brilliantly blue-emitting luciferin-luciferase system from Orfelia fultoni and Keroplatinae (Diptera). Sci. Rep. 2020, 10, 9608. [Google Scholar] [CrossRef]
- Branchini, B.R.; Southworth, T.L.; Fontaine, D.M.; Kohrt, D.; Talukder, M.; Michelini, E.; Cevenini, L.; Roda, A.; Grossel, M.J. An enhanced chimeric firefly luciferase-inspired enzyme for ATP detection and bioluminescence reporter and imaging applications. Anal. Biochem. 2015, 484, 148–153. [Google Scholar] [CrossRef] [PubMed]
- Lopreside, A.; Wan, X.; Michelini, E.; Roda, A.; Wang, B. Comprehensive Profiling of Diverse Genetic Reporters with Application to Whole-Cell and Cell-Free Biosensors. Anal. Chem. 2019, 91, 15284–15292. [Google Scholar] [CrossRef] [PubMed]
- Calabretta, M.M.; Álvarez-Diduk, R.; Michelini, E.; Roda, A.; Merkoçi, A. Nano-lantern on paper for smartphone-based ATP detection. Biosens. Bioelectron. 2020, 150, 111902. [Google Scholar] [CrossRef]
- Michelini, E.; Calabretta, M.M.; Cevenini, L.; Lopreside, A.; Southworth, T.; Fontaine, D.M.; Simoni, P.; Branchini, B.R.; Roda, A. Smartphone-based multicolor bioluminescent 3D spheroid biosensors for monitoring inflammatory activity. Biosens. Bioelectron. 2019, 123, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Santangelo, M.F.; Libertino, S.; Turner, A.P.F.; Filippini, D.; Mak, W.C. Integrating printed microfluidics with silicon photomultipliers for miniaturised and highly sensitive ATP bioluminescence detection. Biosens. Bioelectron. 2018, 99, 464–470. [Google Scholar] [CrossRef] [PubMed]
- Stowe, C.L.; Burley, T.A.; Allan, H.; Vinci, M.; Kramer-Marek, G.; Ciobota, D.M.; Parkinson, G.N.; Southworth, T.L.; Agliardi, G.; Hotblack, A.; et al. Near-infrared dual bioluminescence imaging in mouse models of cancer using infraluciferin. Elife 2019, 8, e45801. [Google Scholar] [CrossRef] [PubMed]
- Branchini, B.R.; Ablamsky, D.M.; Davis, A.L.; Southworth, T.L.; Butler, B.; Fan, F.; Jathoul, A.P.; Pule, M.A. Red-emitting luciferases for bioluminescence reporter and imaging applications. Anal. Biochem. 2010, 396, 290–297. [Google Scholar] [CrossRef]
- El Khamlichi, C.; Reverchon-Assadi, F.; Hervouet-Coste, N.; Blot, L.; Reiter, E.; Séverine, M.L. Bioluminescence resonance energy transfer as a method to study protein-protein interactions: Application to G protein coupled receptor biology. Molecules 2019, 24, 537. [Google Scholar] [CrossRef]
- Dale, N.C.; Johnstone, E.K.M.; White, C.W.; Pfleger, K.D.G. NanoBRET: The bright future of proximity-based assays. Front. Bioeng. Biotechnol. 2019, 7, 56. [Google Scholar] [CrossRef]
- Calabretta, M.M.; Lopreside, A.; Montali, L.; Cevenini, L.; Roda, A.; Michelini, E. A Genetically Encoded Bioluminescence Intracellular Nanosensor for Androgen Receptor Activation Monitoring in 3D Cell Models. Sensors 2021, 21, 893. [Google Scholar] [CrossRef]
- Cevenini, L.; Calabretta, M.M.; Calabria, D.; Roda, A.; Michelini, E. Luciferase genes as reporter reactions: How to use them in molecular biology? Adv. Biochem. Eng. Biotechnol. 2016, 154, 3–17. [Google Scholar] [CrossRef]
- Roda, A.; Roda, B.; Cevenini, L.; Michelini, E.; Mezzanotte, L.; Reschiglian, P.; Hakkila, K.; Virta, M. Analytical strategies for improving the robustness and reproducibility of bioluminescent microbial bioreporters. Anal. Bioanal. Chem. 2011, 401, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Calabretta, M.M.; Lopreside, A.; Montali, L.; Zangheri, M.; Evangelisti, L.; D’Elia, M.; Michelini, E. Portable light detectors for bioluminescence biosensing applications: A comprehensive review from the analytical chemist’s perspective. Anal. Chim. Acta 2022, 1200, 339583. [Google Scholar] [CrossRef] [PubMed]
- Lopreside, A.; Montali, L.; Wang, B.; Tassoni, A.; Ferri, M.; Calabretta, M.M.; Michelini, E. Orthogonal paper biosensor for mercury(II) combining bioluminescence and colorimetric smartphone detection. Biosens. Bioelectron. 2021, 194, 113569. [Google Scholar] [CrossRef] [PubMed]
- Lopreside, A.; Calabretta, M.M.; Montali, L.; Ferri, M.; Tassoni, A.; Branchini, B.R.; Southworth, T.; D’Elia, M.; Roda, A.; Michelini, E. Prêt-à-porter nanoYESA and nanoYESΒ bioluminescent cell biosensors for ultrarapid and sensitive screening of endocrine-disrupting chemicals. Anal. Bioanal. Chem. 2019, 411, 4937–4949. [Google Scholar] [CrossRef] [PubMed]
- Bergua, J.F.; Álvarez-Diduk, R.; Hu, L.; Hassan, A.H.A.; Merkoçi, A. Improved Aliivibrio fischeri based-toxicity assay: Graphene-oxide as a sensitivity booster with a mobile-phone application. J. Hazard. Mater. 2021, 406, 124434. [Google Scholar] [CrossRef]
- Calabretta, M.M.; Montali, L.; Lopreside, A.; Fragapane, F.; Iacoangeli, F.; Roda, A.; Bocci, V.; D’Elia, M.; Michelini, E. Ultrasensitive On-Field Luminescence Detection Using a Low-Cost Silicon Photomultiplier Device. Anal. Chem. 2021, 93, 7388–7393. [Google Scholar] [CrossRef]
- Syed, A.J.; Anderson, J.C. Applications of bioluminescence in biotechnology and beyond. Chem. Soc. Rev. 2021, 50, 5668–5705. [Google Scholar] [CrossRef]
- Pozzo, T.; Akter, F.; Nomura, Y.; Louie, A.Y.; Yokobayashi, Y. Firefly Luciferase Mutant with Enhanced Activity and Thermostability. ACS Omega 2018, 3, 2628–2633. [Google Scholar] [CrossRef]
- Mortazavi, M.; Hosseinkhani, S. Surface charge modification increases firefly luciferase rigidity without alteration in bioluminescence spectra. Enzym. Microb. Technol. 2017, 96, 47–59. [Google Scholar] [CrossRef]
- Branchini, B.R.; Southworth, T.L.; Fontaine, D.M.; Kohrt, D.; Welcome, F.S.; Florentine, C.M.; Henricks, E.R.; DeBartolo, D.B.; Michelini, E.; Cevenini, L.; et al. Red-emitting chimeric firefly luciferase for in vivo imaging in low ATP cellular environments. Anal. Biochem. 2017, 534, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Mortazavi, M.; Hosseinkhani, S. Design of thermostable luciferases through arginine saturation in solvent-exposed loops. Protein Eng. Des. Sel. 2011, 24, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Said Alipour, B.; Hosseinkhani, S.; Ardestani, S.K.; Moradi, A. The effective role of positive charge saturation in bioluminescence color and thermostability of firefly luciferase. Photochem. Photobiol. Sci. 2009, 8, 847. [Google Scholar] [CrossRef]
- Ando, Y.; Niwa, K.; Yamada, N.; Enomoto, T.; Irie, T.; Kubota, H.; Ohmiya, Y.; Akiyama, H. Firefly bioluminescence quantum yield and colour change by pH-sensitive green emission. Nat. Photonics 2008, 2, 44–47. [Google Scholar] [CrossRef]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Meng, E.C.; Couch, G.S.; Croll, T.I.; Morris, J.H.; Ferrin, T.E. UCSF ChimeraX: Structure visualization for researchers, educators, and developers. Protein Sci. 2021, 30, 70–82. [Google Scholar] [CrossRef] [PubMed]
- Branchini, B.R.; Magyar, R.A.; Murtiashaw, M.H.; Anderson, S.M.; Zimmer, M. Site-Directed Mutagenesis of Histidine 245 in Firefly Luciferase: A Proposed Model of the Active Site. Biochemistry 1998, 37, 15311–15319. [Google Scholar] [CrossRef]
- Cevenini, L.; Calabretta, M.M.; Lopreside, A.; Branchini, B.R.; Southworth, T.L.; Michelini, E.; Roda, A. Bioluminescence Imaging of Spheroids for High-throughput Longitudinal Studies on 3D Cell Culture Models. Photochem. Photobiol. 2017, 93, 531–535. [Google Scholar] [CrossRef]
- Law, G.H.E.; Gandelman, O.A.; Tisi, L.C.; Lowe, C.R.; Murray, J.A.H. Mutagenesis of solvent-exposed amino acids in Photinus pyralis luciferase improves thermostability and pH-tolerance. Biochem. J. 2006, 397, 305–312. [Google Scholar] [CrossRef]
- Baggett, B.; Roy, R.; Momen, S.; Morgan, S.; Tisi, L.; Morse, D.; Gillies, R.J. Thermostability of Firefly Luciferases Affects Efficiency of Detection by In Vivo Bioluminescence. Mol. Imaging 2004, 3, 324–332. [Google Scholar] [CrossRef]
- White, P.J.; Squirrell, D.J.; Arnaud, P.; Lowe, C.R.; Murray, J.A.H. Improved thermostability of the North American firefly luciferase: Saturation mutagenesis at position 354. Biochem. J. 1996, 319, 343–350. [Google Scholar] [CrossRef] [PubMed]
- Branchini, B.R.; Fontaine, D.M.; Southworth, T.L.; Huta, B.P.; Racela, A.; Patel, K.D.; Gulick, A.M. Mutagenesis and Structural Studies Reveal the Basis for the Activity and Stability Properties That Distinguish the Photinus Luciferases scintillans and pyralis. Biochemistry 2019, 58, 4293–4303. [Google Scholar] [CrossRef] [PubMed]
- Available online: https://ita.promega.com/products/luciferase-assays/genetic-reporter-vectors-and-cell-lines/promoterless-firefly-luciferase-vectors-with-neomycin-selection/?catNum=E6721 (accessed on 8 September 2022).
- Liang, Y. Comparison of red-shifted firefly luciferase Ppy RE9 and conventional Luc2 as bioluminescence imaging reporter genes for in vivo imaging of stem cells. J. Biomed. Opt. 2012, 17, 016004. [Google Scholar] [CrossRef] [PubMed]
- Tisi, L.C.; Law, G.H.; Gandelman, O.; Lowe, C.R.; Murray, J. The basis of the bathochromic shift in the luciferase from photinus pyralis. In Bioluminescence and Chemiluminescence; World Scientific: Singapore; River Edge, NJ, USA, 2002; pp. 57–60. [Google Scholar]
- Calabretta, M.M.; Gregucci, D.; Guarnieri, T.; Bonini, M.; Neri, E.; Zangheri, M.; Michelini, E. Bioluminescence Sensing in 3D Spherical Microtissues for Multiple Bioactivity Analysis of Environmental Samples. Sensors 2022, 22, 4568. [Google Scholar] [CrossRef] [PubMed]
- Ravi, M.; Paramesh, V.; Kaviya, S.R.; Anuradha, E.; Paul Solomon, F.D. 3D cell culture systems: Advantages and applications. J. Cell. Physiol. 2015, 230, 16–26. [Google Scholar] [CrossRef]
- Jack, G.D.; Mead, E.A.; Garst, J.F.; Cabrera, M.C.; DeSantis, A.M.; Slaughter, S.M.; Jervis, J.; Brooks, A.I.; Potts, M.; Helm, R.F. Long term metabolic arrest and recovery of HEK293 spheroids involves NF-κB signaling and sustained JNK activation. J. Cell. Physiol. 2006, 206, 526–536. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Luciferase | Organism | Emission λmax (nm) (pH 7.8) | Half-Life (h, 37 °C) | Notes | Ref. |
|---|---|---|---|---|---|
| PpyWT | P. pyralis | 557 | 0.3 | pH-sensitive | [42] |
| Luc2 | P. pyralis | 557 | 3.0 | pH-sensitive | [43,44] |
| BgLuc | Synthetic | 548 | 2.5 | pH-independent |
| BgLuc Mutant | BL Emission (37 °C) | BL Emission (45 °C) | ||
|---|---|---|---|---|
| λmax (nm) | Half Bandwidth (nm) | λmax (nm) | Half Bandwidth (nm) | |
| BrightGlo substrate | 552 | 76 | 556 | 82 |
| D-LH2 substrate | 552 | 78 | 556 | 90 |
| BgLuc Mutant | BL Emission pH 5.0 | BL Emission pH 7.0 | BL Emission pH 8.0 | |||
|---|---|---|---|---|---|---|
| λmax (nm) | Half Bandwidth (nm) | λmax (nm) | Half Bandwidth (nm) | λmax (nm) | Half Bandwidth (nm) | |
| BrightGlo substrate | 550 | 48 | 548 | 72 | 552 | 70 |
| D-LH2 substrate | 548 | 124 | 550 | 94 | 550 | 97 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Calabretta, M.M.; Gregucci, D.; Martínez-Pérez-Cejuela, H.; Michelini, E. A Luciferase Mutant with Improved Brightness and Stability for Whole-Cell Bioluminescent Biosensors and In Vitro Biosensing. Biosensors 2022, 12, 742. https://doi.org/10.3390/bios12090742
Calabretta MM, Gregucci D, Martínez-Pérez-Cejuela H, Michelini E. A Luciferase Mutant with Improved Brightness and Stability for Whole-Cell Bioluminescent Biosensors and In Vitro Biosensing. Biosensors. 2022; 12(9):742. https://doi.org/10.3390/bios12090742
Chicago/Turabian StyleCalabretta, Maria Maddalena, Denise Gregucci, Héctor Martínez-Pérez-Cejuela, and Elisa Michelini. 2022. "A Luciferase Mutant with Improved Brightness and Stability for Whole-Cell Bioluminescent Biosensors and In Vitro Biosensing" Biosensors 12, no. 9: 742. https://doi.org/10.3390/bios12090742
APA StyleCalabretta, M. M., Gregucci, D., Martínez-Pérez-Cejuela, H., & Michelini, E. (2022). A Luciferase Mutant with Improved Brightness and Stability for Whole-Cell Bioluminescent Biosensors and In Vitro Biosensing. Biosensors, 12(9), 742. https://doi.org/10.3390/bios12090742