Can ECIS Biosensor Technology Be Used to Measure the Cellular Responses of Glioblastoma Stem Cells?

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Culture
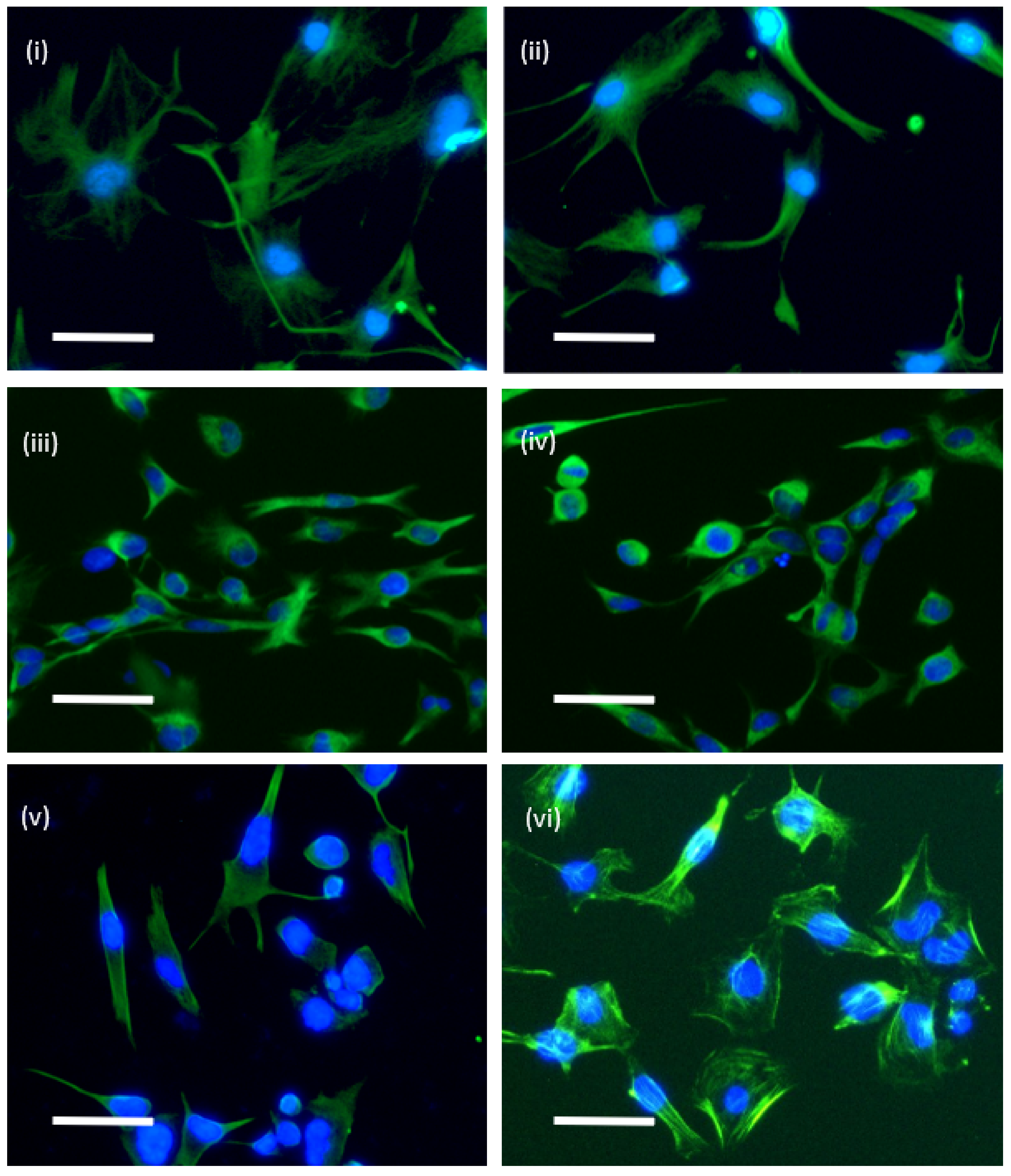
2.2. Immunocytochemistry of Cytoskeletal Proteins
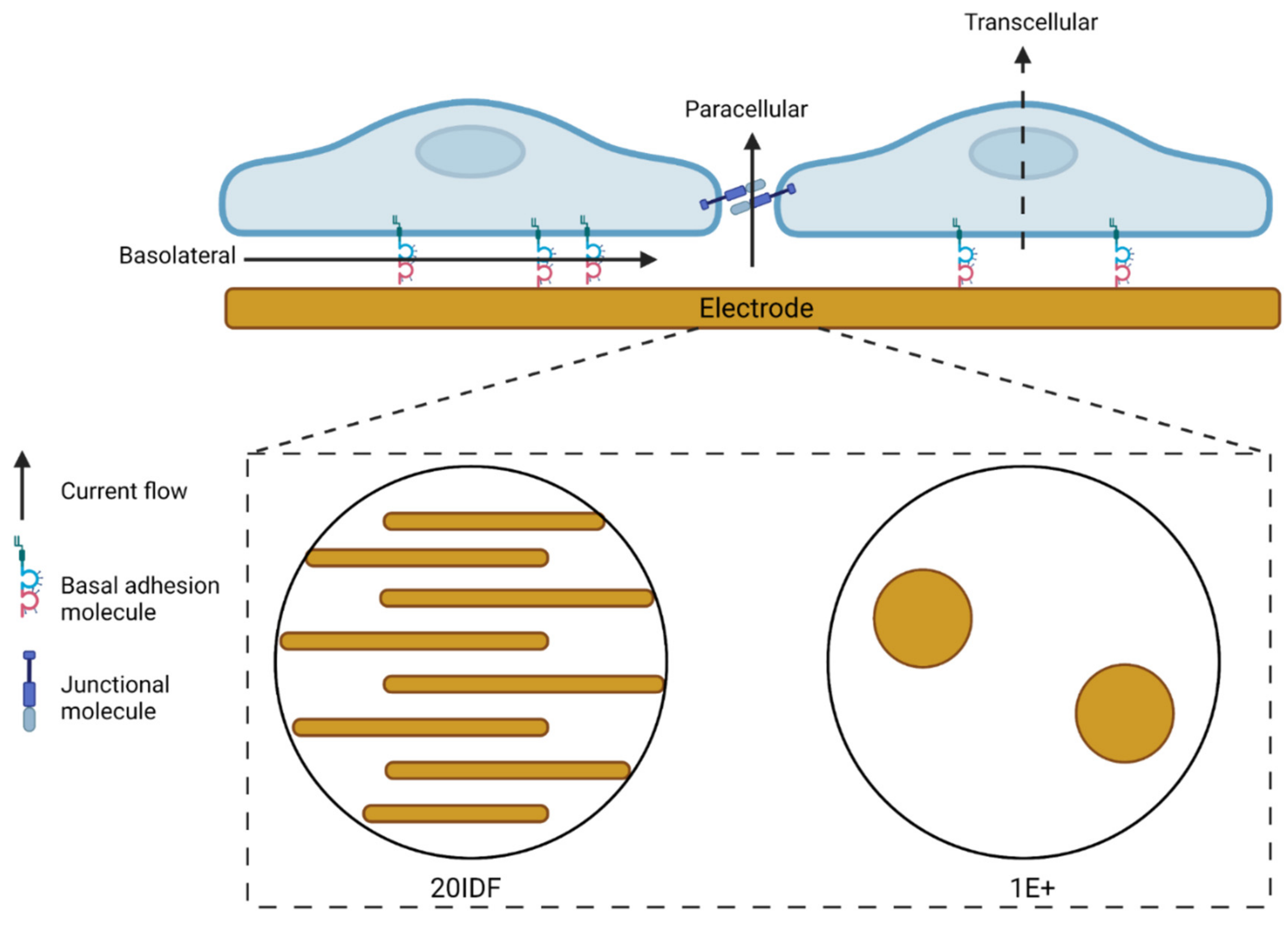
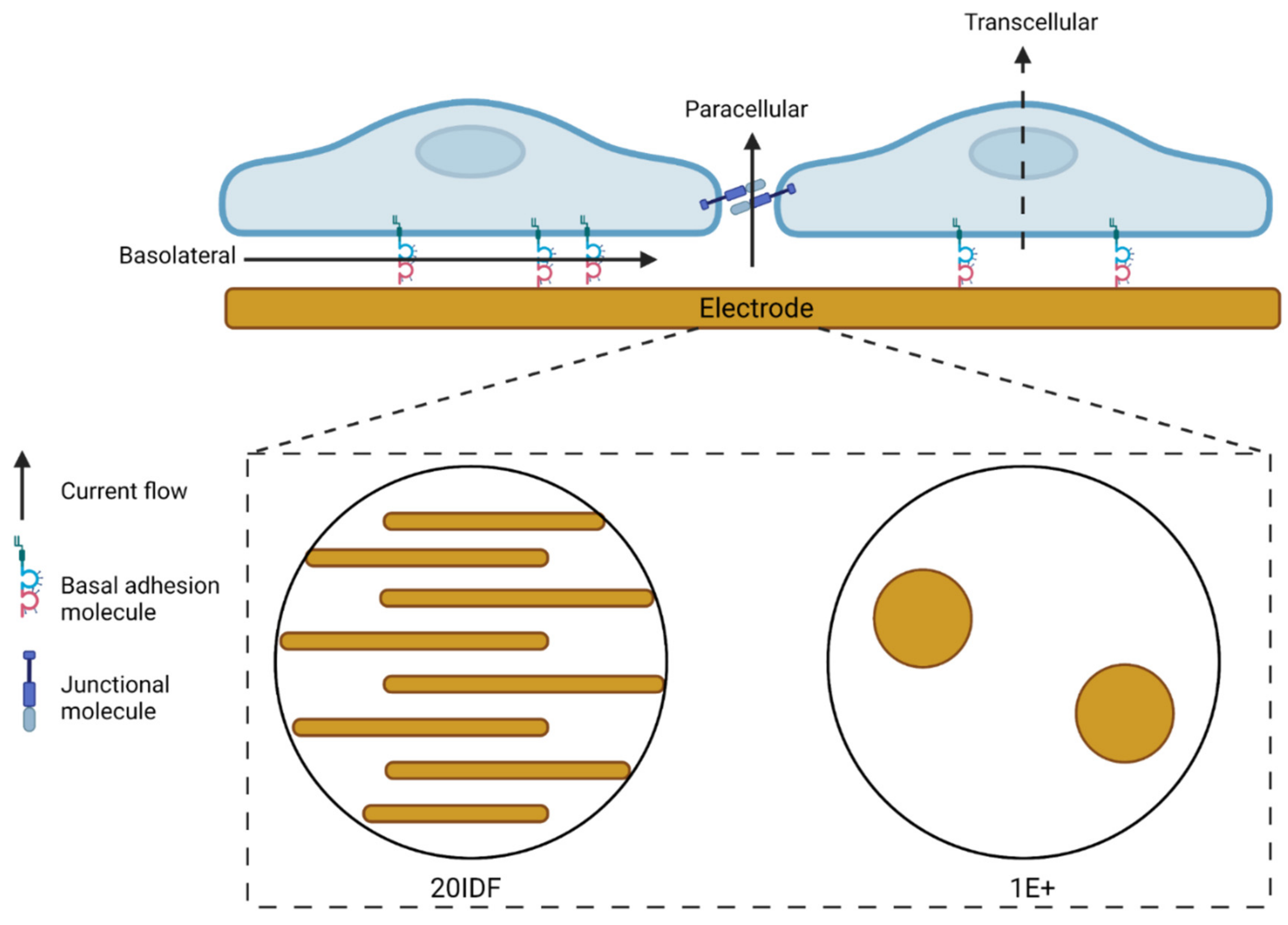
2.3. Electric Cell-Substrate Impedance Sensing (ECIS)
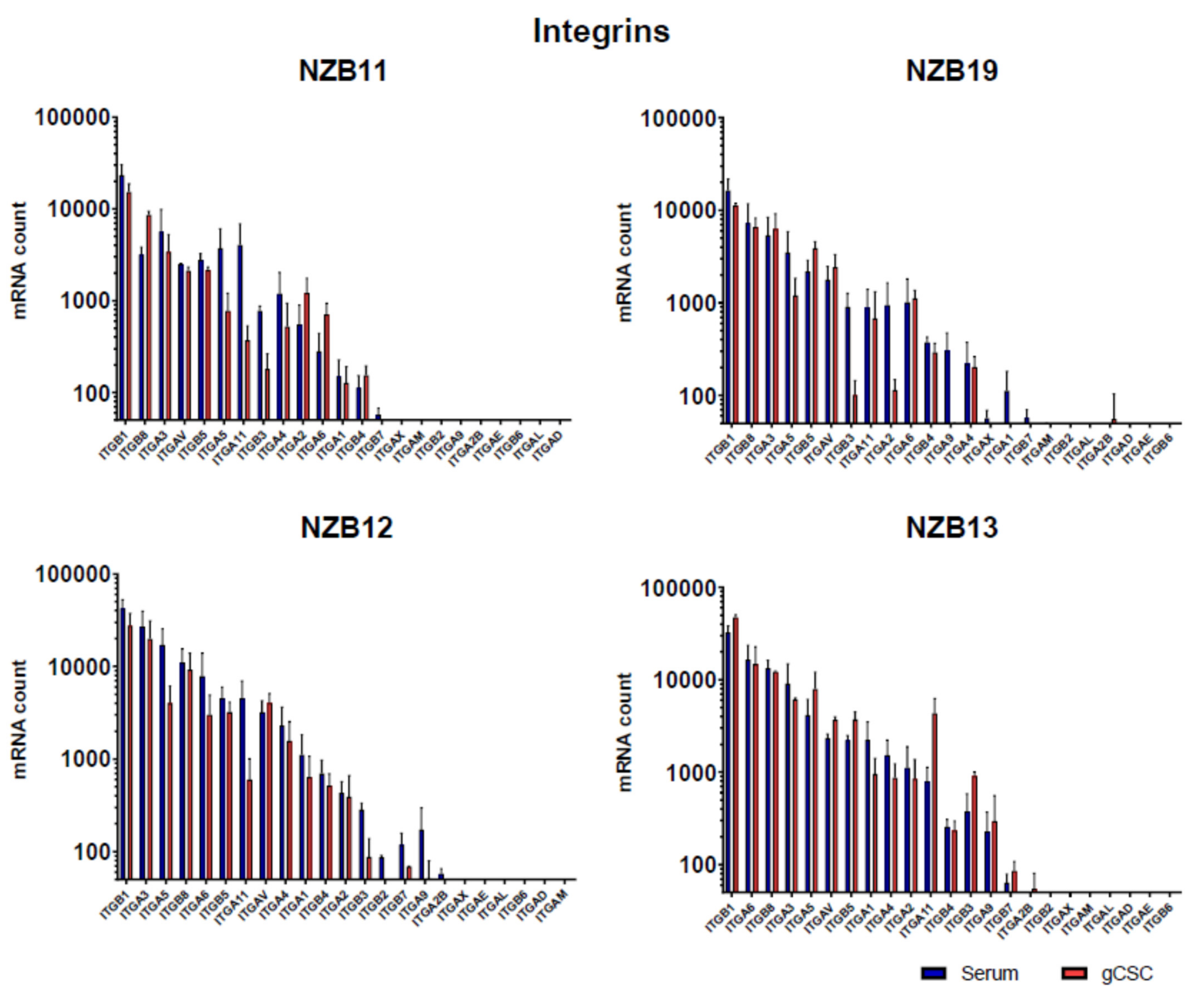
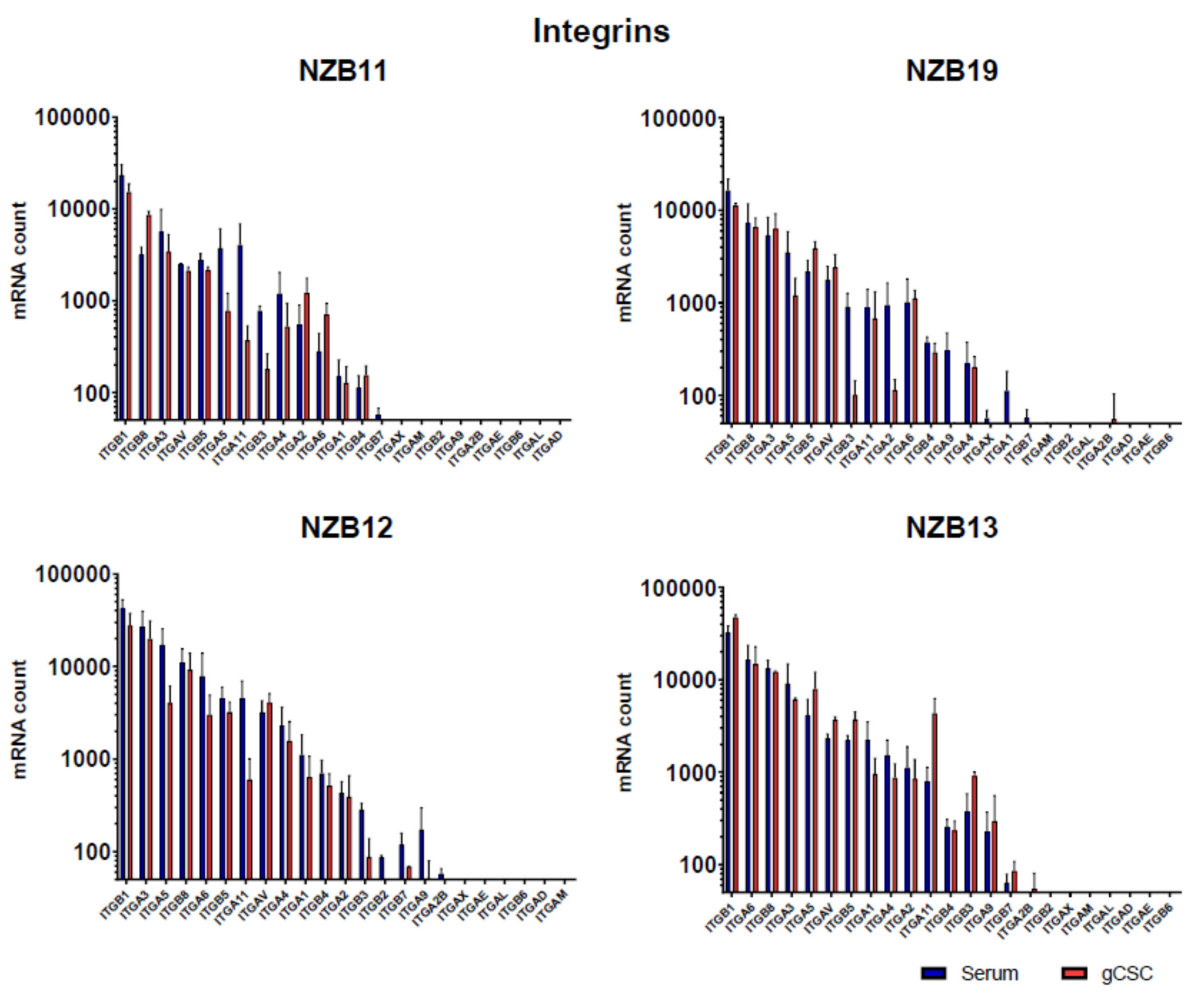
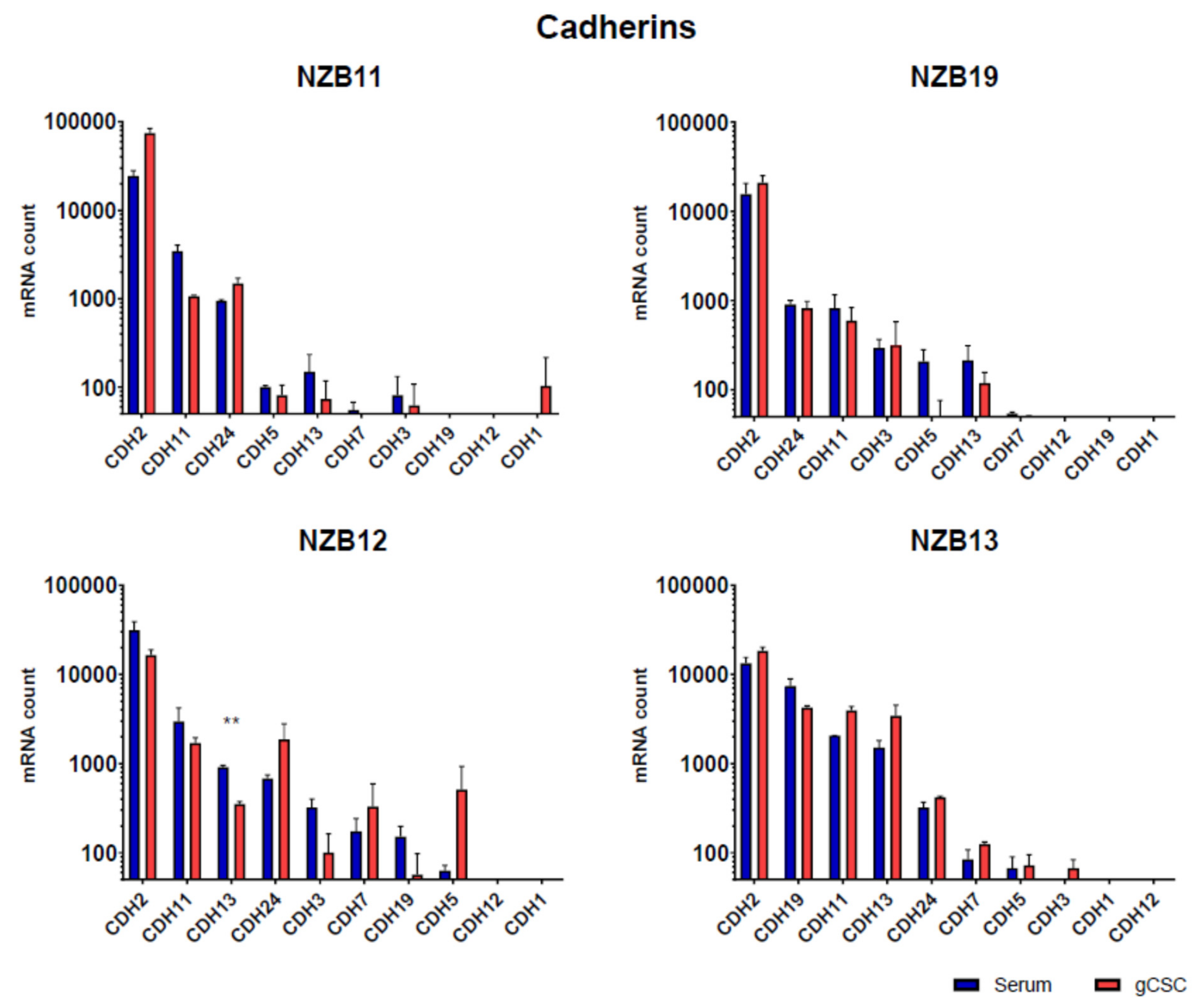
2.4. NanoString Gene Expression Analysis of Glioblastoma Expressed Adhesion Molecules
3. Results
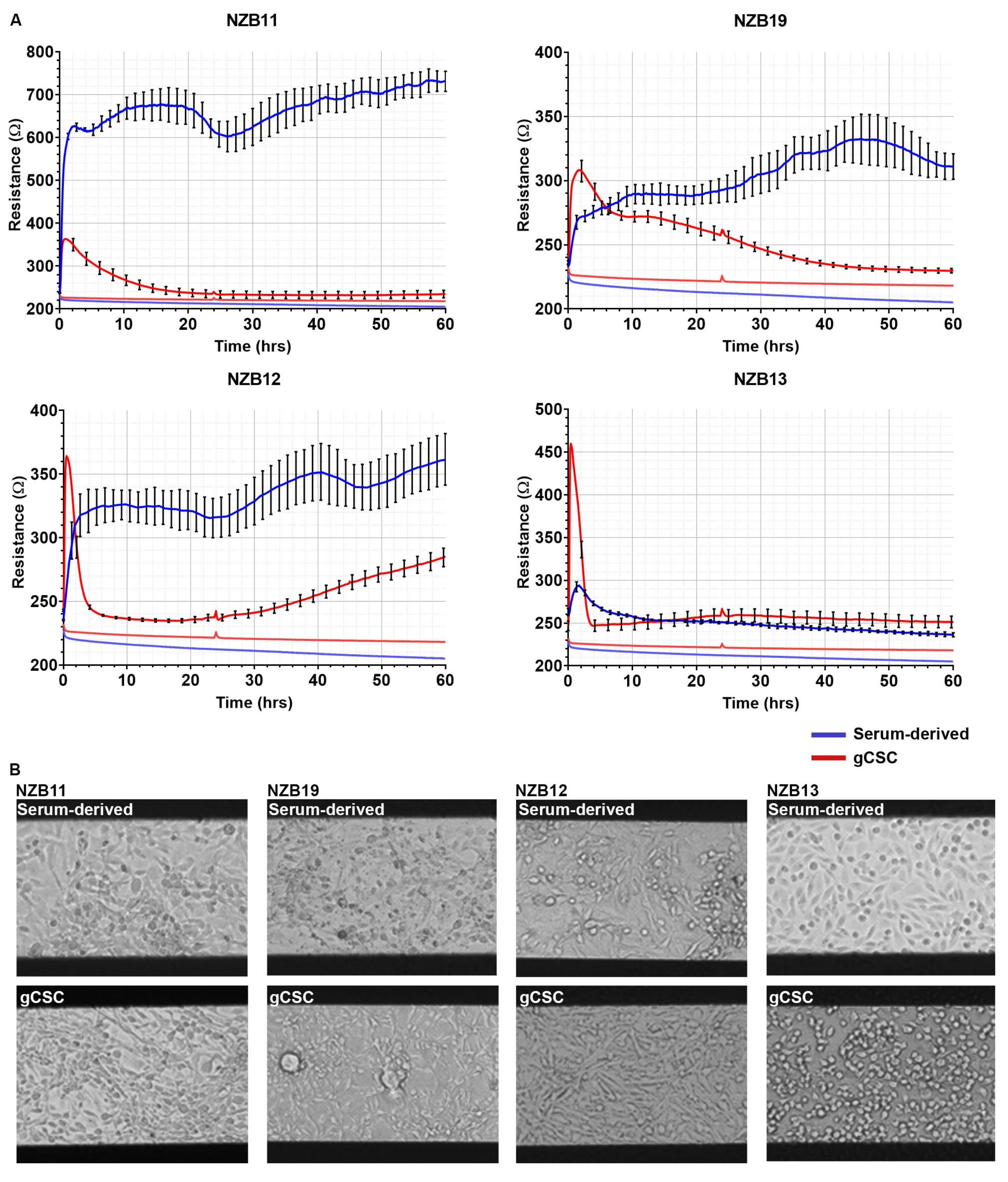
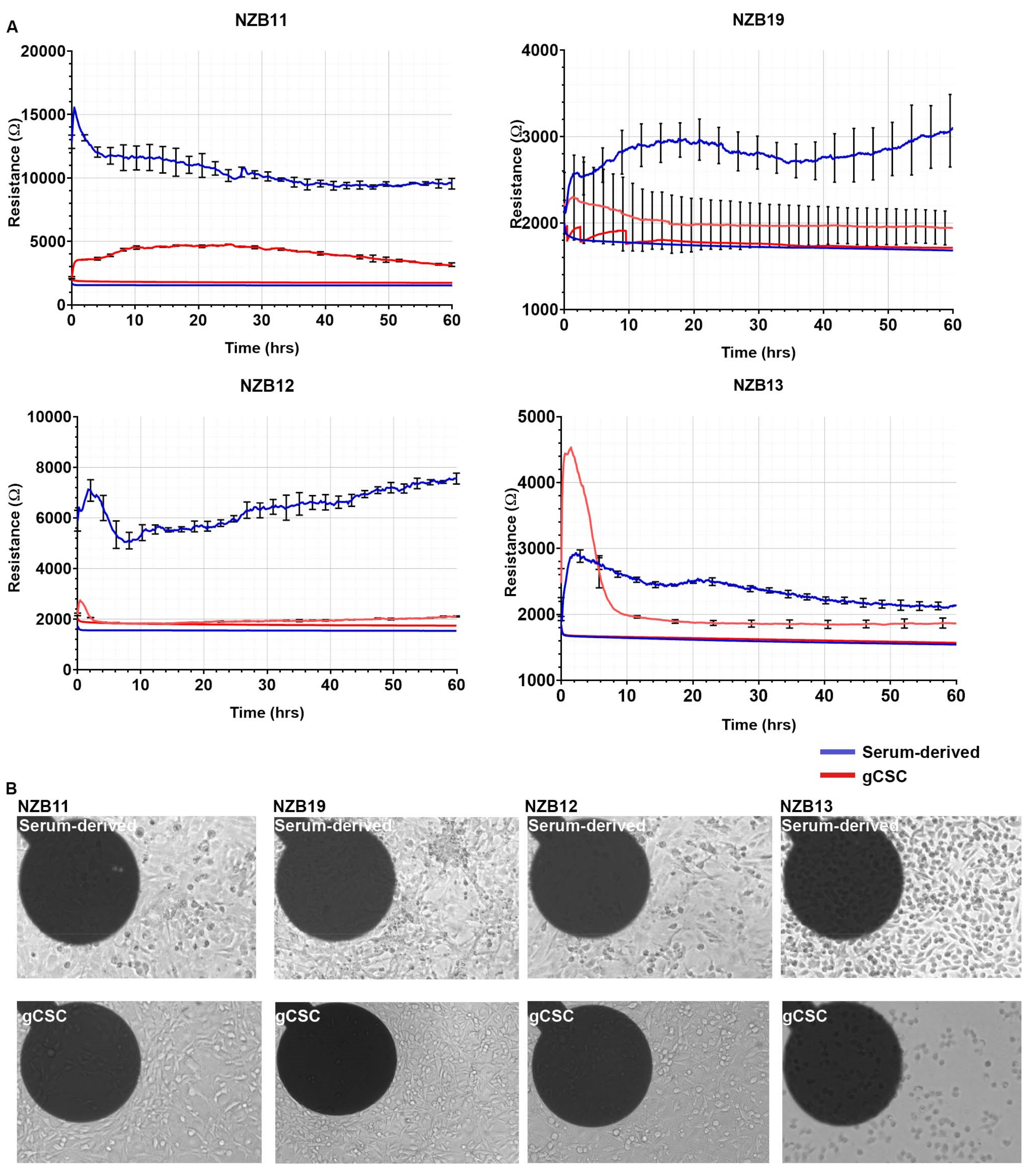
3.1. Serum-Derived Glioblastoma Cell Adhesion Can Be Monitored Using 20IDF ECIS Technology
3.2. Serum Differentiated Glioblastoma Cells Produce Stronger and More Stable Adhesion on 1E+ Arrays
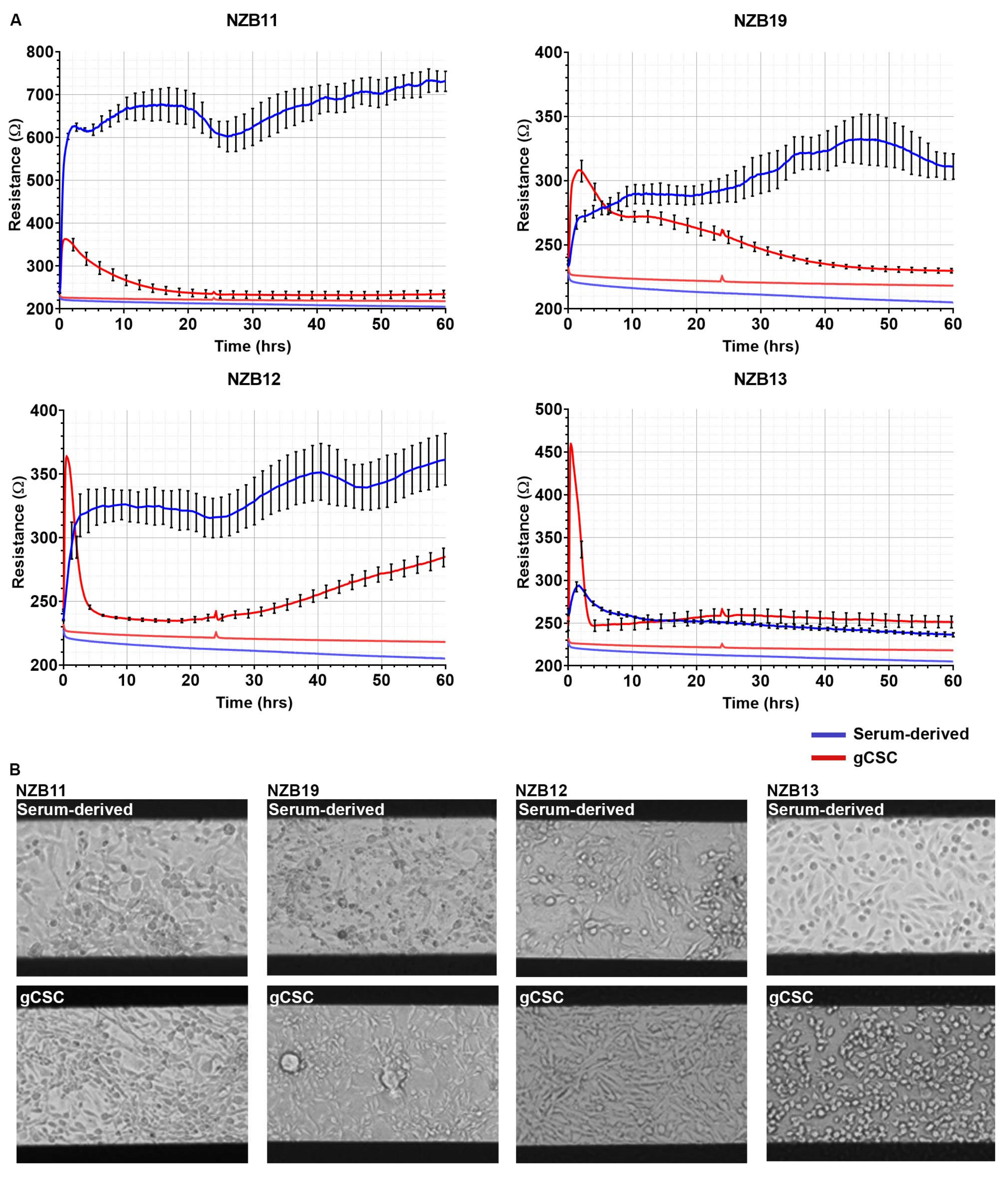
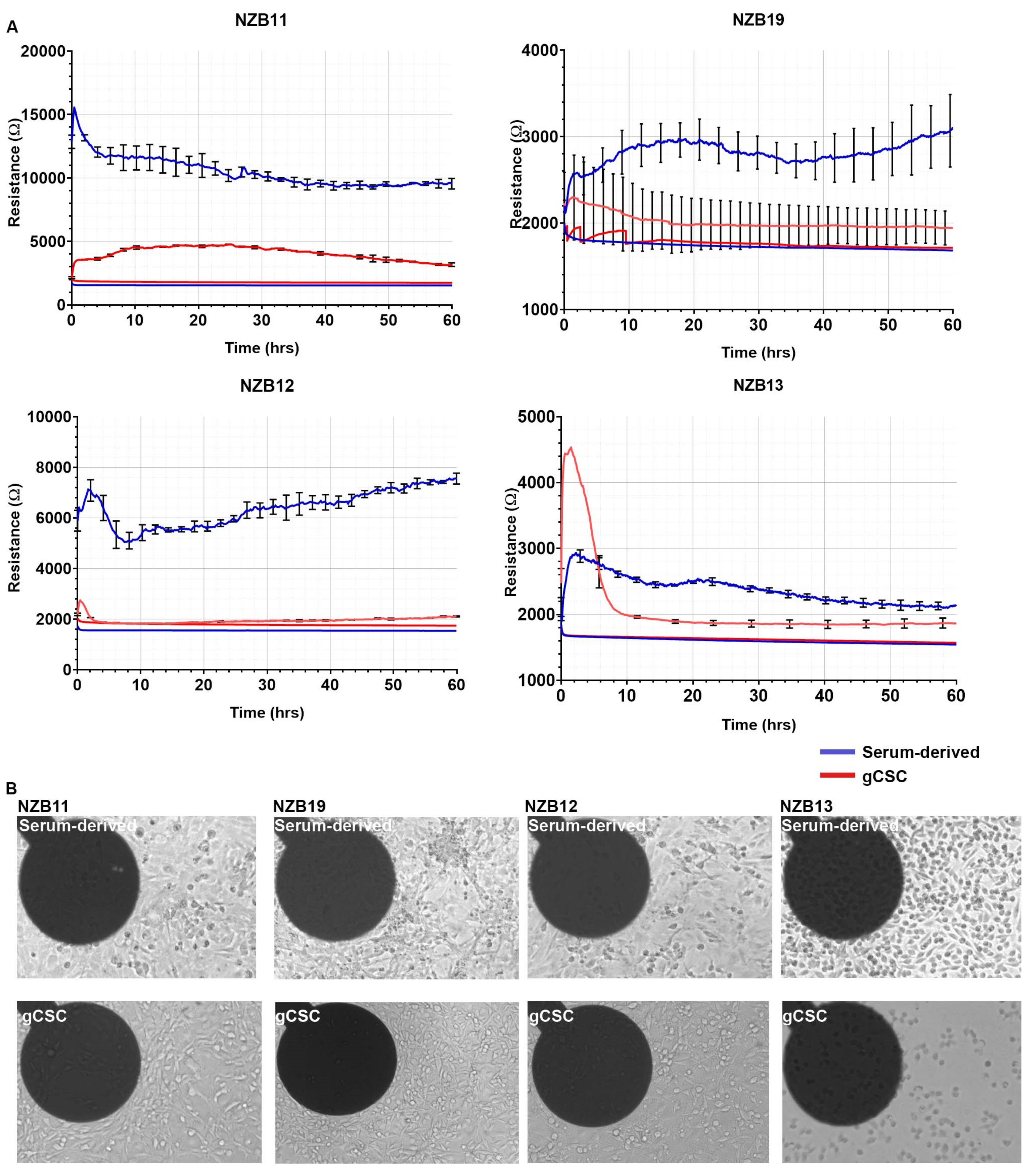
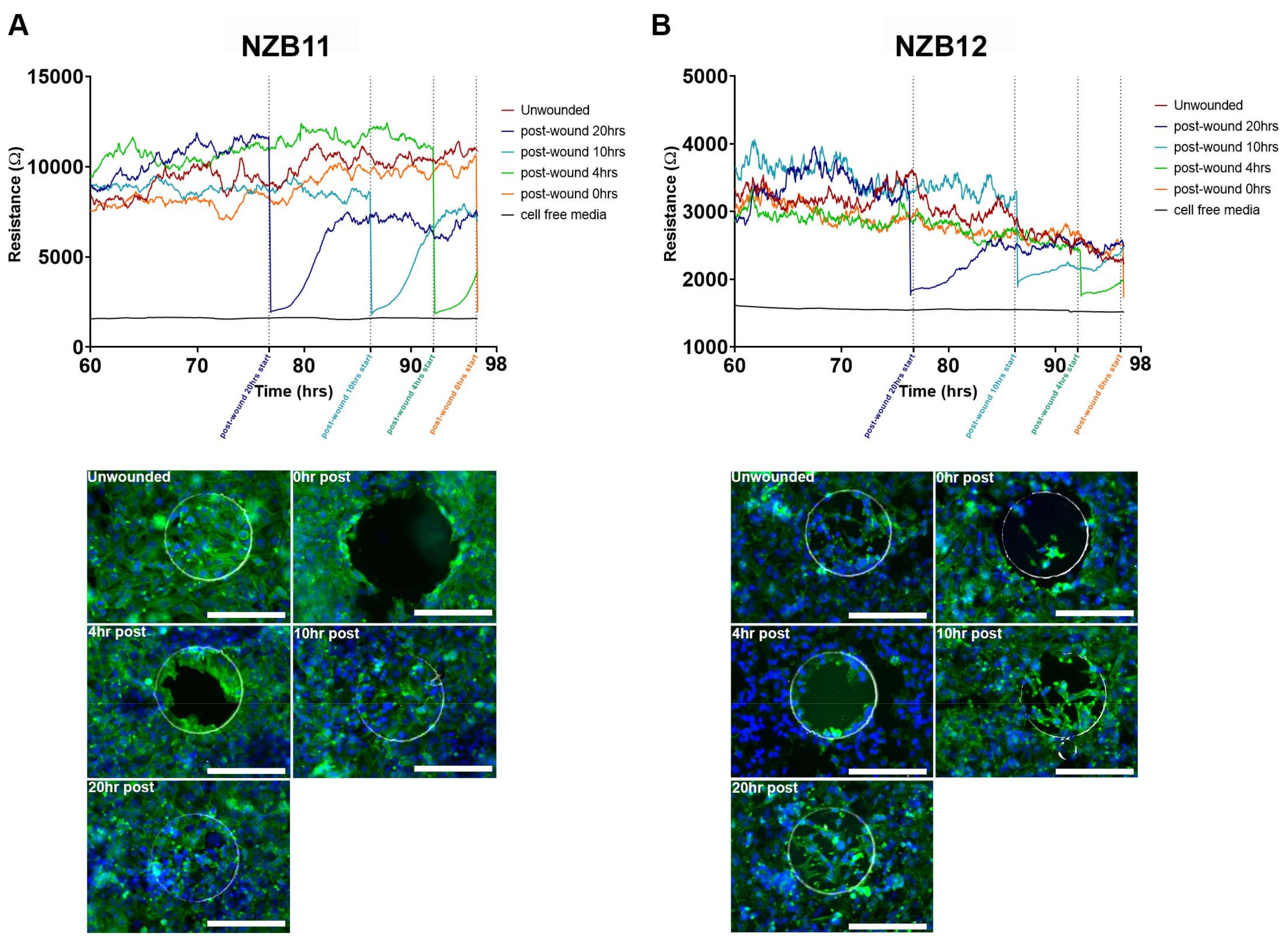
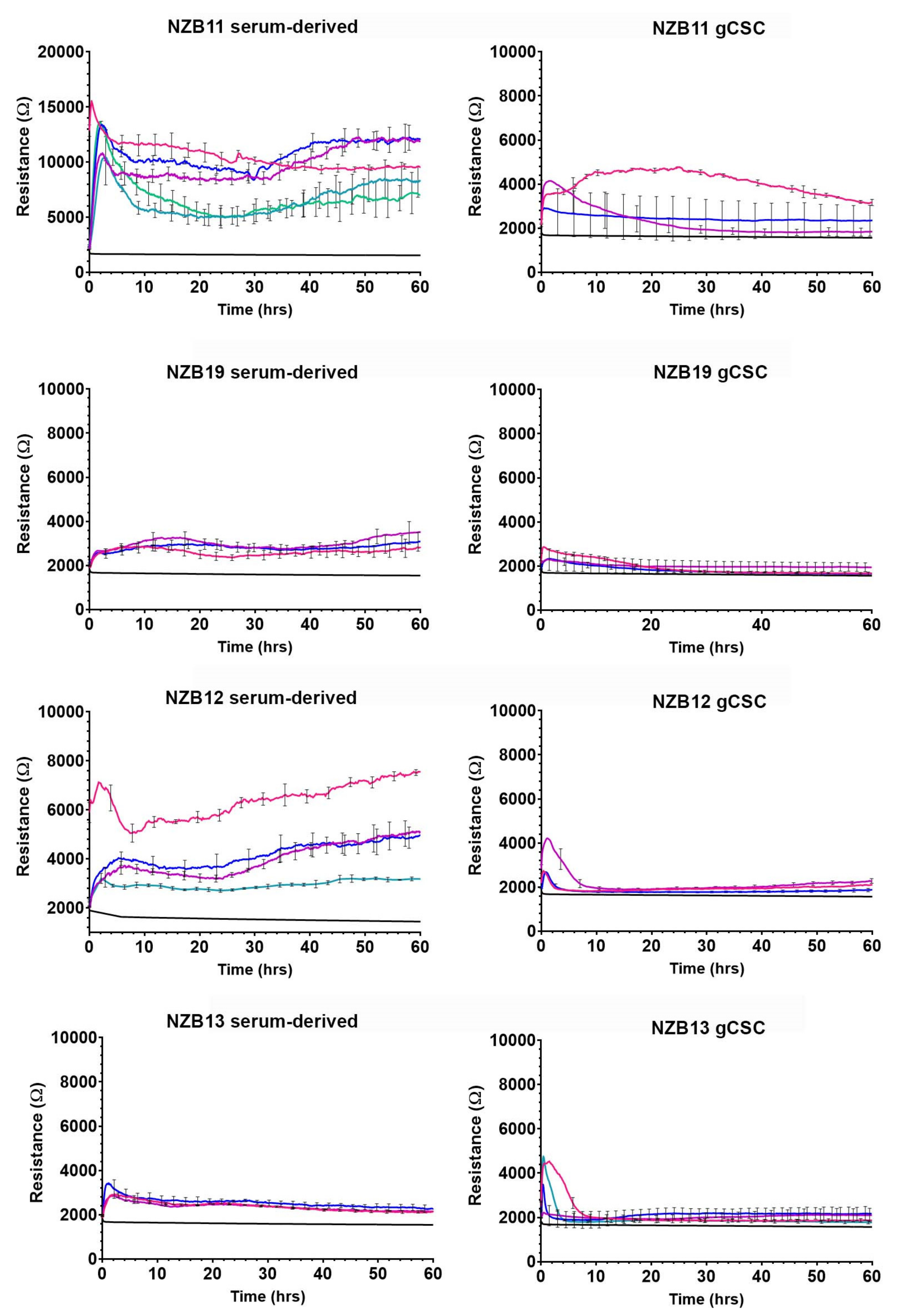
3.3. ECIS Reveals Pronounced Variation in Adhesion Properties across Experiments
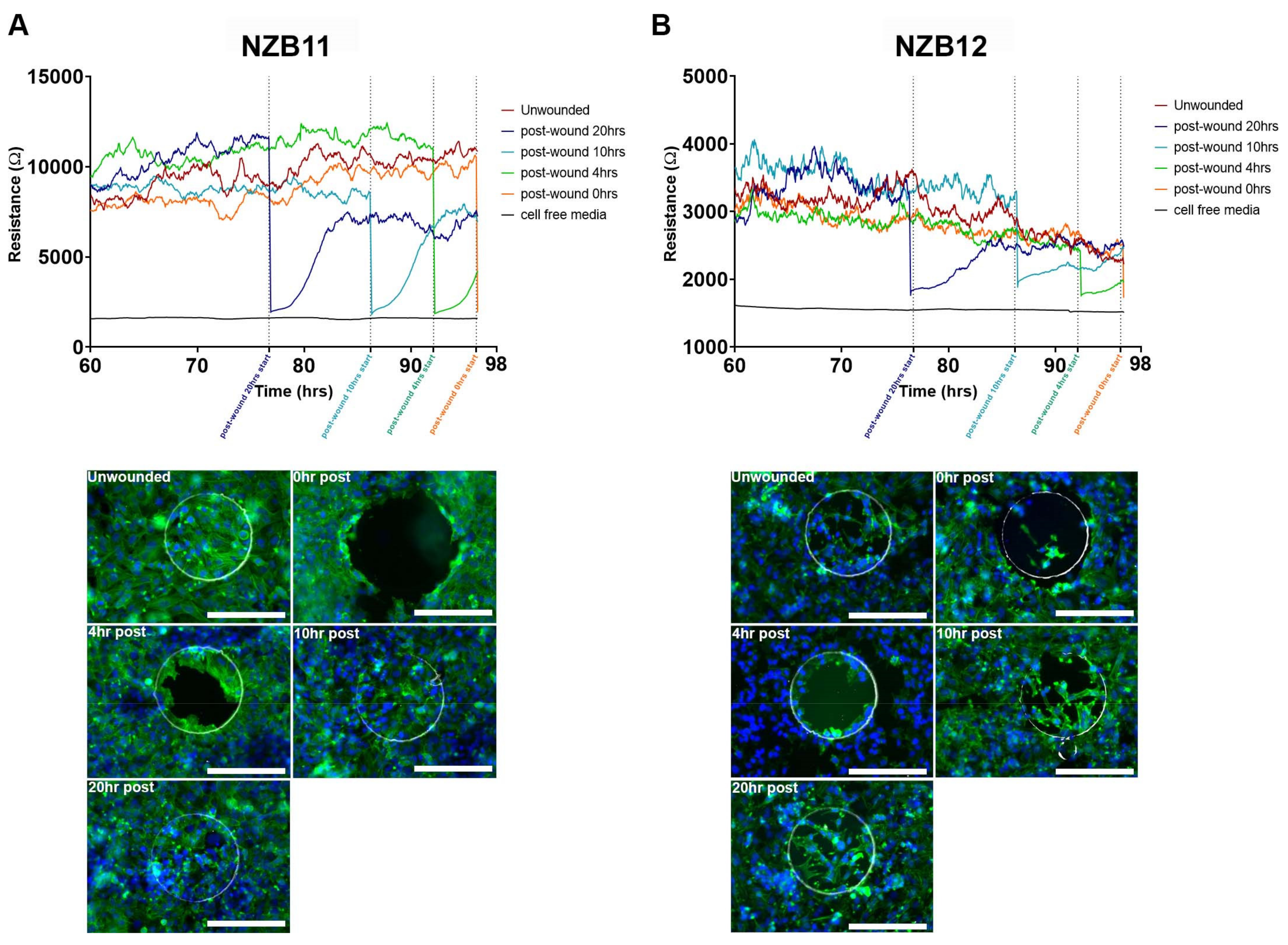
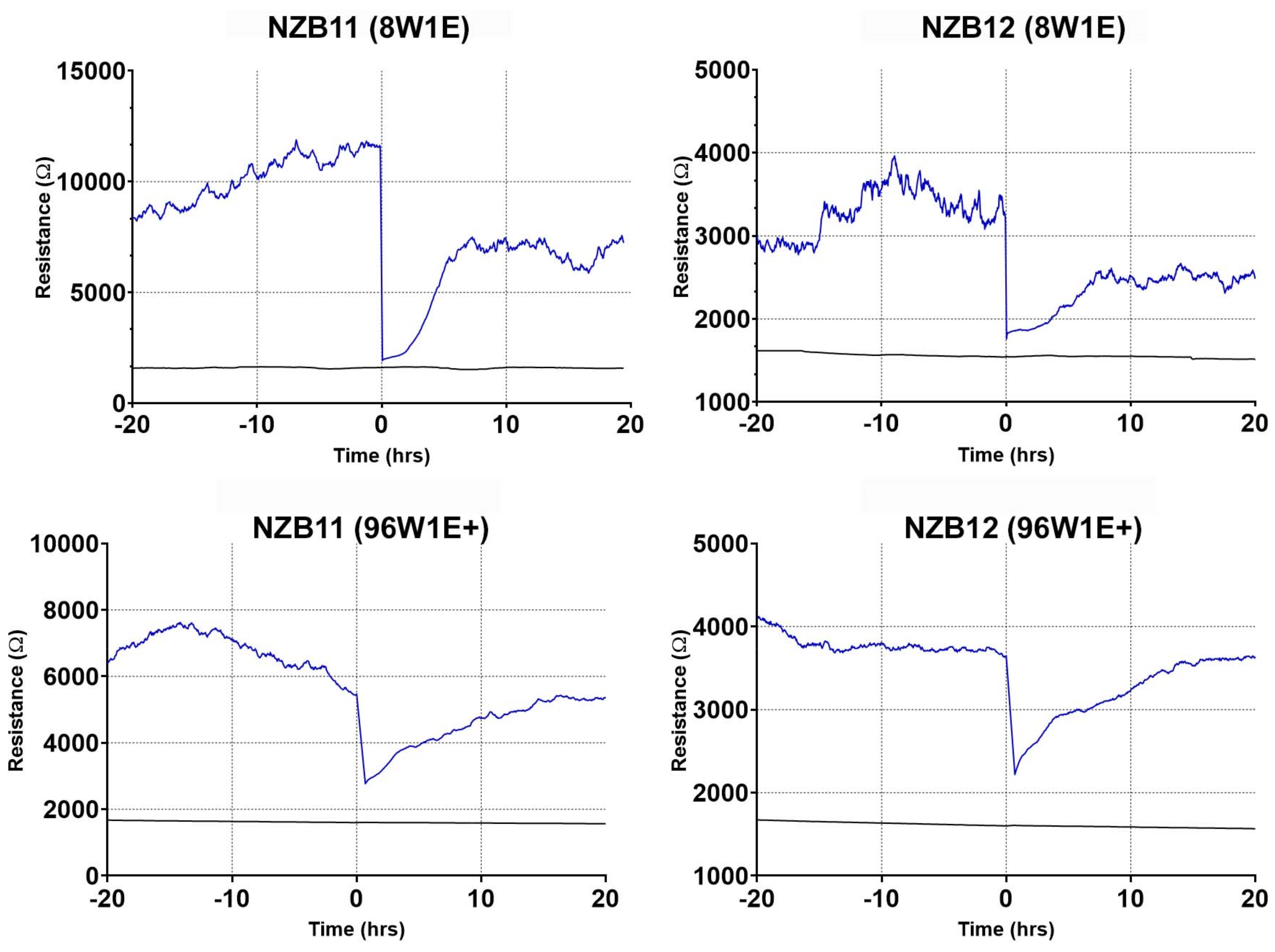
3.4. Ablation of Serum-Derived Glioblastoma Adhesion and Repopulation on ECIS Arrays
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Goenka, A.; Tiek, D.; Song, X.; Huang, T.; Hu, B.; Cheng, S.-Y. The Many Facets of Therapy Resistance and Tumor Recurrence in Glioblastoma. Cells 2021, 10, 484. [Google Scholar] [CrossRef]
- Barcellos-Hoff, M.H.; Newcomb, E.W.; Zagzag, D.; Narayana, A. Therapeutic targets in malignant glioblastoma microenvironment. Semin. Radiat. Oncol. 2009, 19, 163–170. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, H.-M.; Guz-Montgomery, K.; Lowe, D.; Saha, D. Pathogenetic Features and Current Management of Glioblastoma. Cancers 2021, 13, 856. [Google Scholar] [CrossRef] [PubMed]
- Hambardzumyan, D.; Bergers, G. Glioblastoma: Defining Tumor Niches. Trends Cancer 2015, 1, 252–265. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartnett, S.; Kroll, D.; Vogelbaum, M.A. Surgical Adjuncts for Glioblastoma. Neurosurg. Clin. N. Am. 2021, 32, 83–91. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-C.; Lee, I.-C.; Chen, P.-Y. Biomimetic brain tumor niche regulates glioblastoma cells towards a cancer stem cell phenotype. J. Neuro-Oncol. 2018, 137, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Arndt, S.; Seebach, J.; Psathaki, K.; Galla, H.-J.; Wegener, J. Bioelectrical impedance assay to monitor changes in cell shape during apoptosis. Biosens. Bioelectron. 2004, 19, 583–594. [Google Scholar] [CrossRef] [Green Version]
- Opp, D.; Wafula, B.; Lim, J.; Huang, E.; Lo, J.-C.; Lo, C.-M. Use of electric cell–substrate impedance sensing to assess in vitro cytotoxicity. Biosens. Bioelectron. 2009, 24, 2625–2629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Robilliard, L.D.; Kho, D.T.; Johnson, R.H.; Anchan, A.; O’Carroll, S.J.; Graham, E.S. The Importance of Multifrequency Impedance Sensing of Endothelial Barrier Formation Using ECIS Technology for the Generation of a Strong and Durable Paracellular Barrier. Biosensors 2018, 8, 64. [Google Scholar] [CrossRef] [Green Version]
- Wiltshire, R.; Nelson, V.; Kho, D.T.; Angel, C.E.; O’Carroll, S.J.; Graham, E.S. Regulation of human cerebro-microvascular endothelial baso-lateral adhesion and barrier function by S1P through dual involvement of S1P1 and S1P2 receptors. Sci. Rep. 2016, 6, 19814. [Google Scholar] [CrossRef] [Green Version]
- Kho, D.; Johnson, R.H.; Robilliard, L.; Du Mez, E.; McIntosh, J.; O’Carroll, S.J.; Angel, C.; Graham, E.S. ECIS technology reveals that monocytes isolated by CD14+ve selection mediate greater loss of BBB integrity than untouched monocytes, which occurs to a greater extent with IL-1β activated endothelium in comparison to TNFα. PLoS ONE 2017, 12, e0180267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kho, D.T.; Johnson, R.H.; O’Carroll, S.J.; Angel, C.E.; Graham, E.S. Biosensor Technology Reveals the Disruption of the Endothelial Barrier Function and the Subsequent Death of Blood Brain Barrier Endothelial Cells to Sodium Azide and Its Gaseous Products. Biosensors 2017, 7, 41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anchan, A.; Kalogirou-Baldwin, P.; Johnson, R.; Kho, D.T.; Joseph, W.; Hucklesby, J.; Finlay, G.J.; O’Carroll, S.J.; Angel, C.E.; Graham, E.S. Real-Time Measurement of Melanoma Cell-Mediated Human Brain Endothelial Barrier Disruption Using Electric Cell-Substrate Impedance Sensing Technology. Biosensors 2019, 9, 56. [Google Scholar] [CrossRef] [Green Version]
- Robilliard, L.D.; Yu, J.; Anchan, A.; Joseph, W.; Finlay, G.; Angel, C.E.; Graham, E.S. Comprehensive analysis of inhibitory checkpoint ligand expression by glioblastoma cells. Immunol. Cell Biol. 2021, 99, 403–418. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, C.A.; Ross, A.H. Cancer stem cells: Cell culture, markers, and targets for new therapies. J. Cell. Biochem. 2009, 108, 1031–1038. [Google Scholar] [CrossRef] [Green Version]
- Lathia, J.D.; Mack, S.C.; Mulkearns-Hubert, E.E.; Valentim, C.L.; Rich, J.N. Cancer stem cells in glioblastoma. Genes Dev. 2015, 29, 1203–1217. [Google Scholar] [CrossRef] [Green Version]
- Silver, D.J.; Sinyuk, M.; Vogelbaum, M.A.; Ahluwalia, M.S.; Lathia, J.D. The intersection of cancer, cancer stem cells, and the immune system: Therapeutic opportunities. Neuro-Oncology 2016, 18, 153–159. [Google Scholar] [CrossRef] [PubMed]
- Orzan, F.; De Bacco, F.; Crisafulli, G.; Pellegatta, S.; Mussolin, B.; Siravegna, G.; D’Ambrosio, A.; Comoglio, P.M.; Finocchiaro, G.; Boccaccio, C. Genetic Evolution of Glioblastoma Stem-Like Cells from Primary to Recurrent Tumor. Stem Cells 2017, 35, 2218–2228. [Google Scholar] [CrossRef] [Green Version]
- Patel, A.P.; Tirosh, I.; Trombetta, J.J.; Shalek, A.K.; Gillespie, S.M.; Wakimoto, H.; Cahill, D.P.; Nahed, B.V.; Curry, W.T.; Martuza, R.L.; et al. Single-cell RNA-seq highlights intratumoral heterogeneity in primary glioblastoma. Science 2014, 344, 1396–1401. [Google Scholar] [CrossRef] [Green Version]
- Al-Mayhani, T.M.F.; Ball, S.L.; Zhao, J.-W.; Fawcett, J.; Ichimura, K.; Collins, P.V.; Watts, C. An efficient method for derivation and propagation of glioblastoma cell lines that conserves the molecular profile of their original tumours. J. Neurosci. Methods 2009, 176, 192–199. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.; Kotliarova, S.; Kotliarov, Y.; Li, A.; Su, Q.; Donin, N.M.; Pastorino, S.; Purow, B.W.; Christopher, N.; Zhang, W.; et al. Tumor stem cells derived from glioblastomas cultured in bFGF and EGF more closely mirror the phenotype and genotype of primary tumors than do serum-cultured cell lines. Cancer Cell 2006, 9, 391–403. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession | Target Sequence |
|---|---|---|
| Housekeepers | ||
| MRPS5 | NM_031902.3 | ATCCCTACGCCAGCTTGAGCCGTGCACTGCAGACACAATGCTGTATTTCTTCTCCCAGTCACCTGATGAGCCAGCAGTATAGACCATATAGTTTCTTCAC |
| PCNA | NM_002592.2 | GGTGTTGGAGGCACTCAAGGACCTCATCAACGAGGCCTGCTGGGATATTAGCTCCAGCGGTGTAAACCTGCAGAGCATGGACTCGTCCCACGTCTCTTTG |
| PPIA | NM_021130.3 | TCTATGGGGAGAAATTTGAAGATGAGAACTTCATCCTAAAGCATACGGGTCCTGGCATCTTGTCCATGGCAAATGCTGGACCCAACACAAATGGTTCCCA |
| TBP | NM_001172085.1 | ACAGTGAATCTTGGTTGTAAACTTGACCTAAAGACCATTGCACTTCGTGCCCGAAACGCCGAATATAATCCCAAGCGGTTTGCTGCGGTAATCATGAGGA |
| CDH1 | NM_001317184.1 | CCTTCCTCCCAATACATCTCCCTTCACAGCAGAACTAACACACGGGGCGAGTGCCAACTGGACCATTCAGTACAACGACCCAACCCAAGAATCTATCATT |
| CDH11 | NM_001797.2 | CAGGAAGCCAAAGTCCCAGTGGCCATTAGGGTCCTTGATGTCAACGATAATGCTCCCAAGTTTGCTGCCCCTTATGAAGGTTTCATCTGTGAGAGTGATC |
| CDH12 | NM_004061.3 | CGCCTAATCTTCACCCGCTGCTAGGCTCGTTTAATGAGTCTTCTGAGAGCTAAGGAGTCCTCGGATTCATTCAAAGCATTCTACAATGAACGCTAGGGGG |
| CDH13 | NM_001220488.1 | ATCTGCCATGCAAAACGAGGGAGCGTTAGGAAGGAATCCGTCTTGTAAAGCCATTGGTCCTGGTCATCAGCCTCTACCCAATGCTTTCGTGATGCTGCTG |
| CDH19 | NM_021153.2 | TCCAGAAGGAACATTAGTTATCCAGGTGACAGCAAGTGATGCTGACGATCCCTCAAGTGGTAATAATGCTCGTCTCCTCTACAGCTTACTTCAAGGCCAG |
| CDH2 | NM_001792.3 | GGTCATCCCTCCAATCAACTTGCCAGAAAACTCCAGGGGACCTTTTCCTCAAGAGCTTGTCAGGATCAGGTCTGATAGAGATAAAAACCTTTCACTGCGG |
| CDH24 | NM_022478.3 | TCCTCTCCTTCCTCCGTGGCGTTTTGTCTCTGCAGTTCTGAAGCTCACACATAGTCTCCCTGCGTCTTCCTTGCCCATACACATGCTCTGTGTCTGTCTC |
| CDH3 | NM_001317195.1 | AGCTCTGTTTAGCACTGATAATGATGACTTCACTGTGCGGAATGGCGAGACAGTCCAGGAAAGAAGGTCACTGAAGGAAAGGAATCCATTGAAGATCTTC |
| CDH4 | NM_001794.2 | AGAGAAAGTTCAGCAGTACACAGTCATCGTTCAGGCCACAGATATGGAAGGAAATCTCAACTATGGCCTCTCAAACACAGCCACAGCCATCATCACGGTG |
| CDH5 | NM_001795.3 | TCTCCCCTTCTCTGCCTCACCTGGTCGCCAATCCATGCTCTCTTTCTTTTCTCTGTCTACTCCTTATCCCTTGGTTTAGAGGAACCCAAGATGTGGCCTT |
| CDH7 | NM_004361.2 | GTTACACGCTACGGATAGAAGCTGCAAATAAAGATGCCGACCCTCGCTTTCTGAGCTTGGGTCCGTTCAGTGACACGACAACTGTGAAGATAATTGTGGA |
| ITGA1 | NM_181501.1 | AAGTGGCAAGACTATAAGGAAAGAGTATGCACAACGTATTCCATCAGGTGGGGATGGTAAGACACTGAAATTTTTTGGCCAGTCTATCCACGGAGAAATG |
| ITGA11 | NM_012211.3 | CCTGAAAAAGTTTTACATTGGCCCAGGGCAGATCCAGGTTGGAGTTGTGCAGTATGGCGAAGATGTGGTGCATGAGTTTCACCTCAACGACTACAGGTCT |
| ITGA2 | NM_002203.2 | CAACGGGTGTGTGTTCTGACATCAGTCCTGATTTTCAGCTCTCAGCCAGCTTCTCACCTGCAACTCAGCCCTGCCCTTCCCTCATAGATGTTGTGGTTGT |
| ITGA2B | NM_000419.3 | AGTTACCGCCCAGGCATCCTTTTGTGGCACGTGTCCTCCCAGAGCCTCTCCTTTGACTCCAGCAACCCAGAGTACTTCGACGGCTACTGGGGGTACTCGG |
| ITGA3 | NM_005501.2 | CATGATTCAGCGCAAGGAGTGGGACTTATCTGAGTATAGTTACAAGGACCCAGAGGACCAAGGAAACCTCTATATTGGGTACACGATGCAGGTAGGCAGC |
| ITGA4 | NM_000885.4 | GCCCACTGCCAACTGGCTCGCCAACGCTTCAGTGATCAATCCCGGGGCGATTTACAGATGCAGGATCGGAAAGAATCCCGGCCAGACGTGCGAACAGCTC |
| ITGA5 | NM_002205.2 | AGAAGACTTTGTTGCTGGTGTGCCCAAAGGGAACCTCACTTACGGCTATGTCACCATCCTTAATGGCTCAGACATTCGATCCCTCTACAACTTCTCAGGG |
| ITGA6 | NM_000210.1 | CTCATGCGAGCCTTCATTGATGTGACTGCTGCTGCCGAAAATATCAGGCTGCCAAATGCAGGCACTCAGGTTCGAGTGACTGTGTTTCCCTCAAAGACTG |
| ITGA9 | NM_002207.2 | CATGTCTCCAACCTCCTTTGTATATGGCGAGTCCGTGGACGCAGCCAACTTCATTCAGCTGGATGACCTGGAGTGTCACTTTCAGCCCATCAATATCACC |
| ITGAD | NM_005353.2 | ATTGACGGCTCTGGAAGCATTGACCAAAATGACTTTAACCAGATGAAGGGCTTTGTCCAAGCTGTCATGGGCCAGTTTGAGGGCACTGACACCCTGTTTG |
| ITGAE | NM_002208.4 | CTGAATGCAGAGAACCACAGAACTAAGATCACTGTCGTCTTCCTGAAAGATGAGAAGTACCATTCTTTGCCTATCATCATTAAAGGCAGCGTTGGTGGAC |
| ITGAL | NM_001114380.1 | GCAGGATGACACATTTATTGGGAATGAACCATTGACACCAGAAGTGAGAGCAGGCTATTTGGGTTACACCGTGACCTGGCTGCCCTCCCGGCAAAAGACT |
| ITGAM | NM_000632.3 | GCCCTCCGAGGGTGTCCTCAAGAGGATAGTGACATTGCCTTCTTGATTGATGGCTCTGGTAGCATCATCCCACATGACTTTCGGCGGATGAAGGAGTTTG |
| ITGAV | NM_002210.2 | TTTCTTCCGATTCCAAACTGGGAGCACAAGGAGAACCCTGAGACTGAAGAAGATGTTGGGCCAGTTGTTCAGCACATCTATGAGCTGAGAAACAATGGTC |
| ITGAX | NM_000887.3 | CCCCTCAGCCTGTTGGCTTCTGTTCACCAGCTGCAAGGGTTTACATACACGGCCACCGCCATCCAAAATGTCGTGCACCGATTGTTCCATGCCTCATATG |
| ITGB1 | NM_033666.2 | TTTTAACATTACCAAGGTAGAAAGTCGGGACAAATTACCCCAGCCGGTCCAACCTGATCCTGTGTCCCATTGTAAGGAGAAGGATGTTGACGACTGTTGG |
| ITGB2 | NM_000211.2 | CATCGACCTGTACTATCTGATGGACCTCTCCTACTCCATGCTTGATGACCTCAGGAATGTCAAGAAGCTAGGTGGCGACCTGCTCCGGGCCCTCAACGAG |
| ITGB3 | NM_000212.2 | GAATAAGCCTTGGAATTAGATATGGGGCAATGACTGAGCCCTGTCTCACCCATGGATTACTCCTTACTGTAGGGAATGGCAGTATGGTAGAGGGATAAAT |
| ITGB4 | NM_001005731.1 | GGCCCATGTCCATCCCCATCATCCCTGACATCCCTATCGTGGACGCCCAGAGCGGGGAGGACTACGACAGCTTCCTTATGTACAGCGATGACGTTCTACG |
| ITGB5 | NM_002213.3 | TAATCTCTTCTTTACTGCTACCTGCCAAGATGGGGTATCCTATCCTGGTCAGAGGAAGTGTGAGGGTCTGAAGATTGGGGACACGGCATCTTTTGAAGTA |
| ITGB6 | NM_000888.3 | AACATTCTCCAGCTGATCATCTCAGCTTATGAAGAACTGCGGTCTGAGGTGGAACTGGAAGTATTAGGAGACACTGAAGGACTCAACTTGTCATTTACAG |
| ITGB7 | NM_000889.1 | CAACGTGGTACAGCTCATCATGGATGCTTATAATAGCCTGTCTTCCACCGTGACCCTTGAACACTCTTCACTCCCTCCTGGGGTCCACATTTCTTACGAA |
| ITGB8 | NM_002214.2 | GGAAAACTGGAATTGTATGCAATGCCTTCACCCTCACAATTTGTCTCAGGCTATACTTGATCAGTGCAAAACCTCATGTGCTCTCATGGAACAACAGCAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robilliard, L.D.; Yu, J.; Jun, S.-M.; Anchan, A.; Finlay, G.; Angel, C.E.; Graham, E.S. Can ECIS Biosensor Technology Be Used to Measure the Cellular Responses of Glioblastoma Stem Cells? Biosensors 2021, 11, 498. https://doi.org/10.3390/bios11120498
Robilliard LD, Yu J, Jun S-M, Anchan A, Finlay G, Angel CE, Graham ES. Can ECIS Biosensor Technology Be Used to Measure the Cellular Responses of Glioblastoma Stem Cells? Biosensors. 2021; 11(12):498. https://doi.org/10.3390/bios11120498
Chicago/Turabian StyleRobilliard, Laverne Diana, Jane Yu, Sung-Min Jun, Akshata Anchan, Graeme Finlay, Catherine E. Angel, and Euan Scott Graham. 2021. "Can ECIS Biosensor Technology Be Used to Measure the Cellular Responses of Glioblastoma Stem Cells?" Biosensors 11, no. 12: 498. https://doi.org/10.3390/bios11120498
APA StyleRobilliard, L. D., Yu, J., Jun, S.-M., Anchan, A., Finlay, G., Angel, C. E., & Graham, E. S. (2021). Can ECIS Biosensor Technology Be Used to Measure the Cellular Responses of Glioblastoma Stem Cells? Biosensors, 11(12), 498. https://doi.org/10.3390/bios11120498