
Seed Priming with Single-Walled Carbon Nanotubes Grafted with Pluronic P85 Preserves the Functional and Structural Characteristics of Pea Plants

 , ,
, ,  ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Pluronic P85 and P85-SWCNT Dispersions
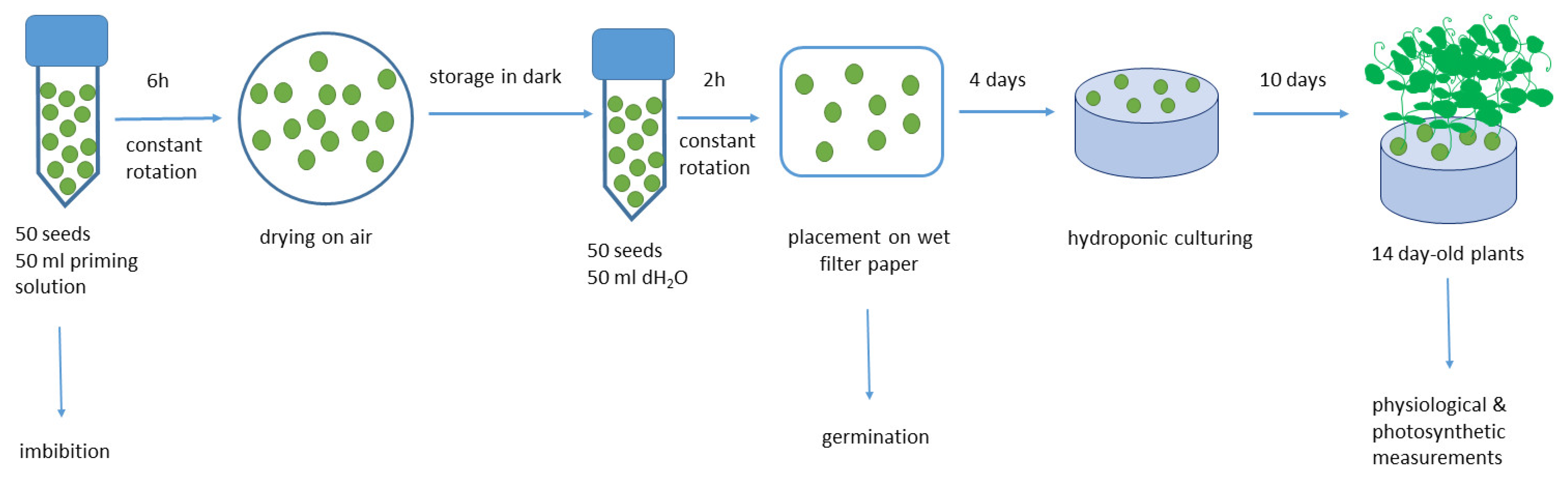
2.3. Seed Priming
2.4. Seed Germination and Plant Development
2.5. Leaf Anatomy
2.6. Leaf Pigments and Chlorophyll Fluorescence
2.7. Statistical Evaluation
3. Results
3.1. Seed Germination
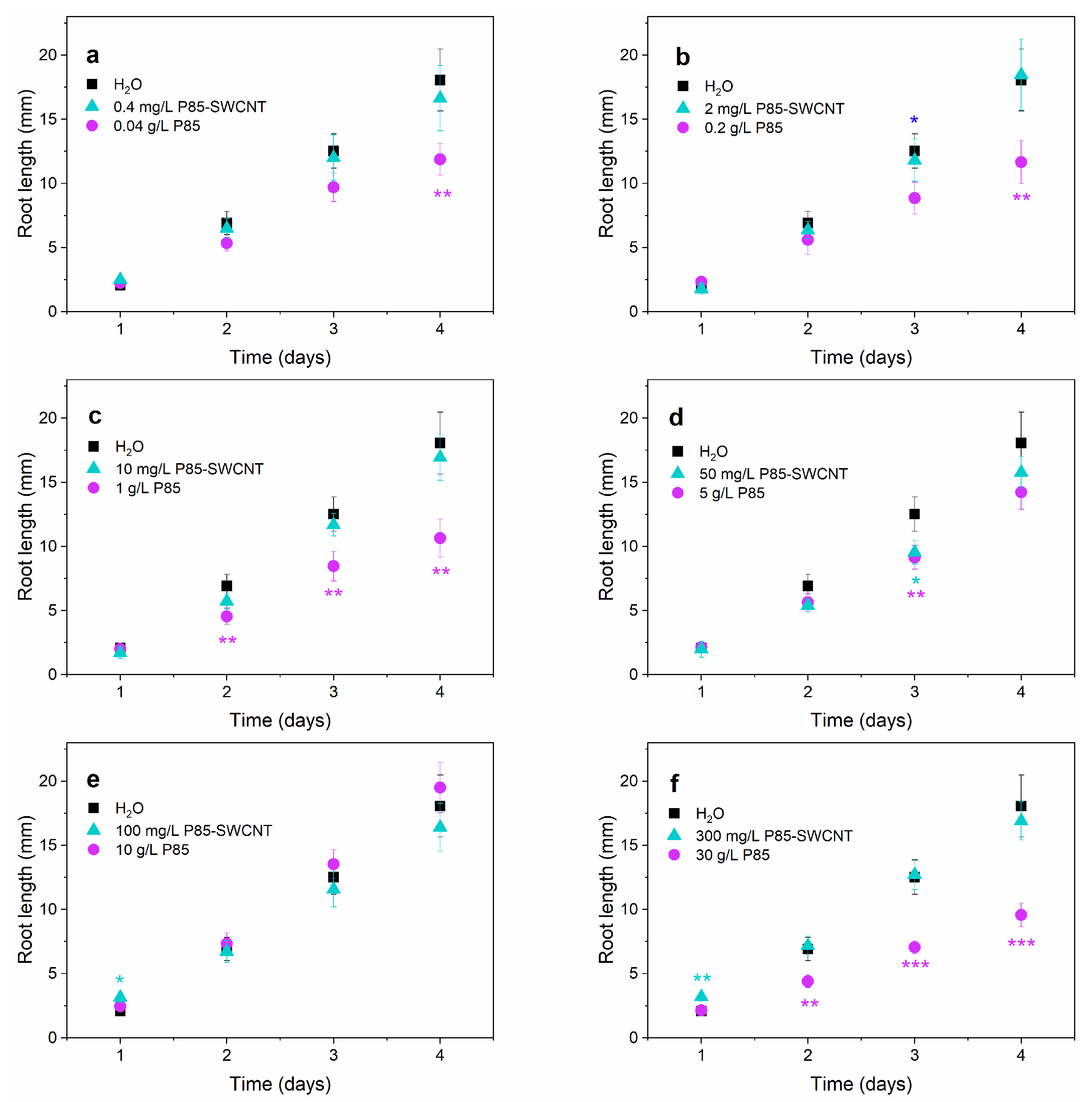
3.2. Plant Development
3.3. Leaf Anatomical Traits
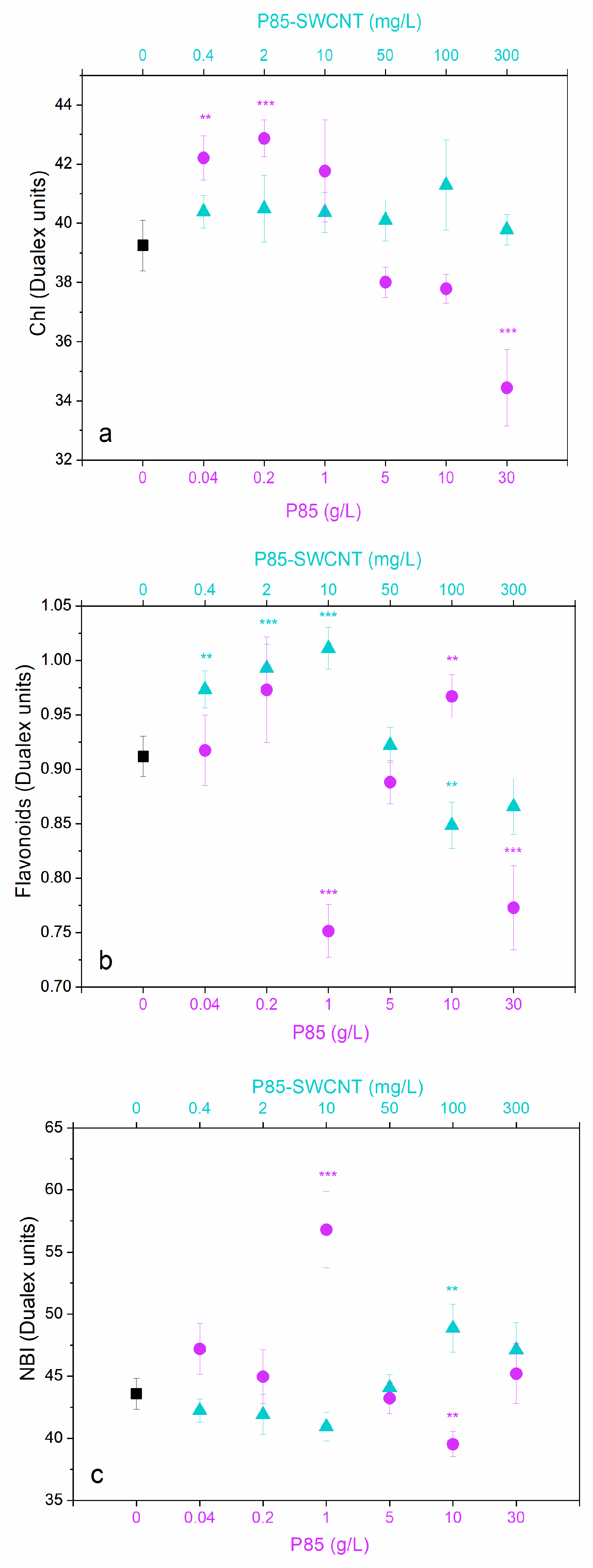
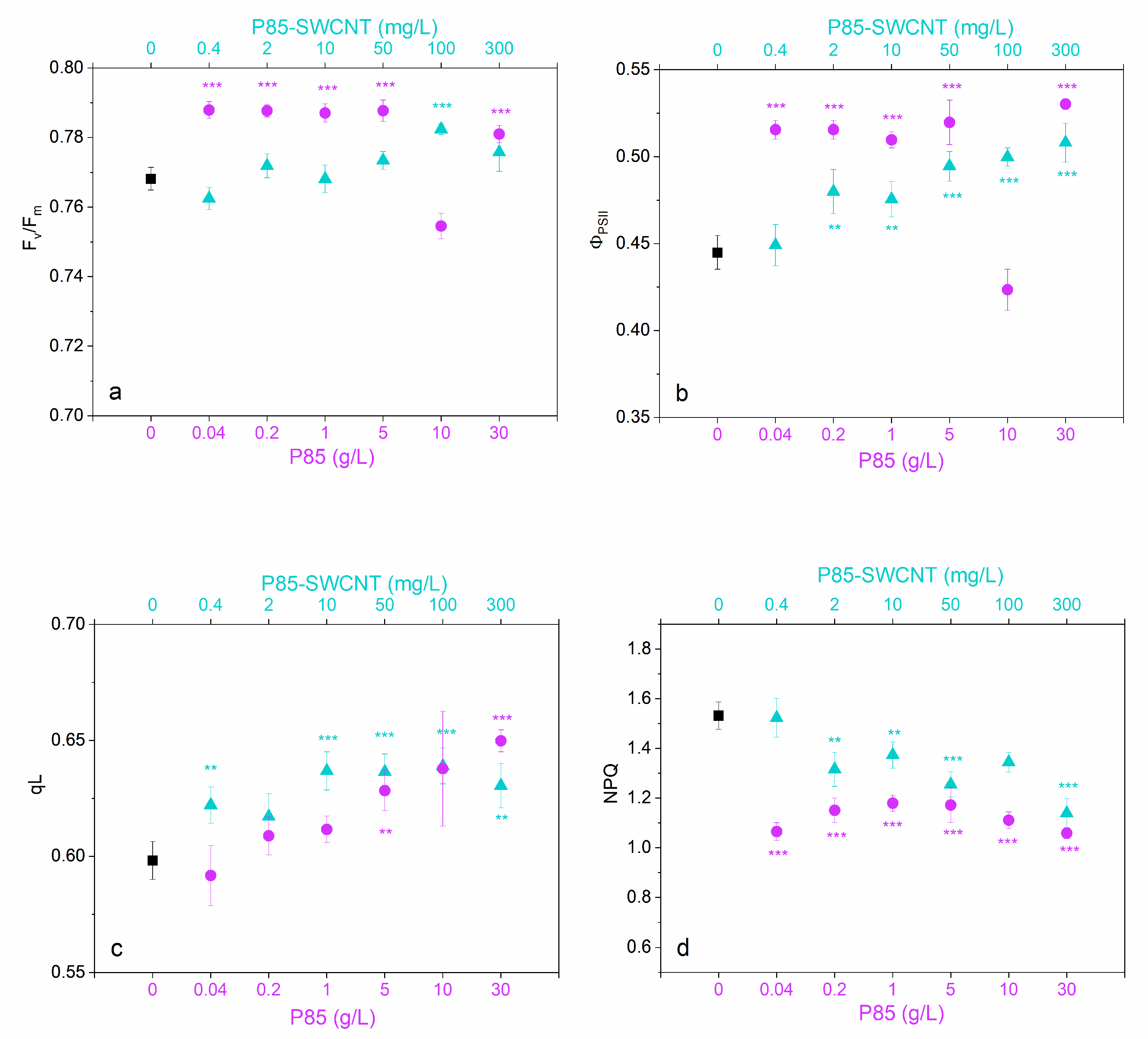
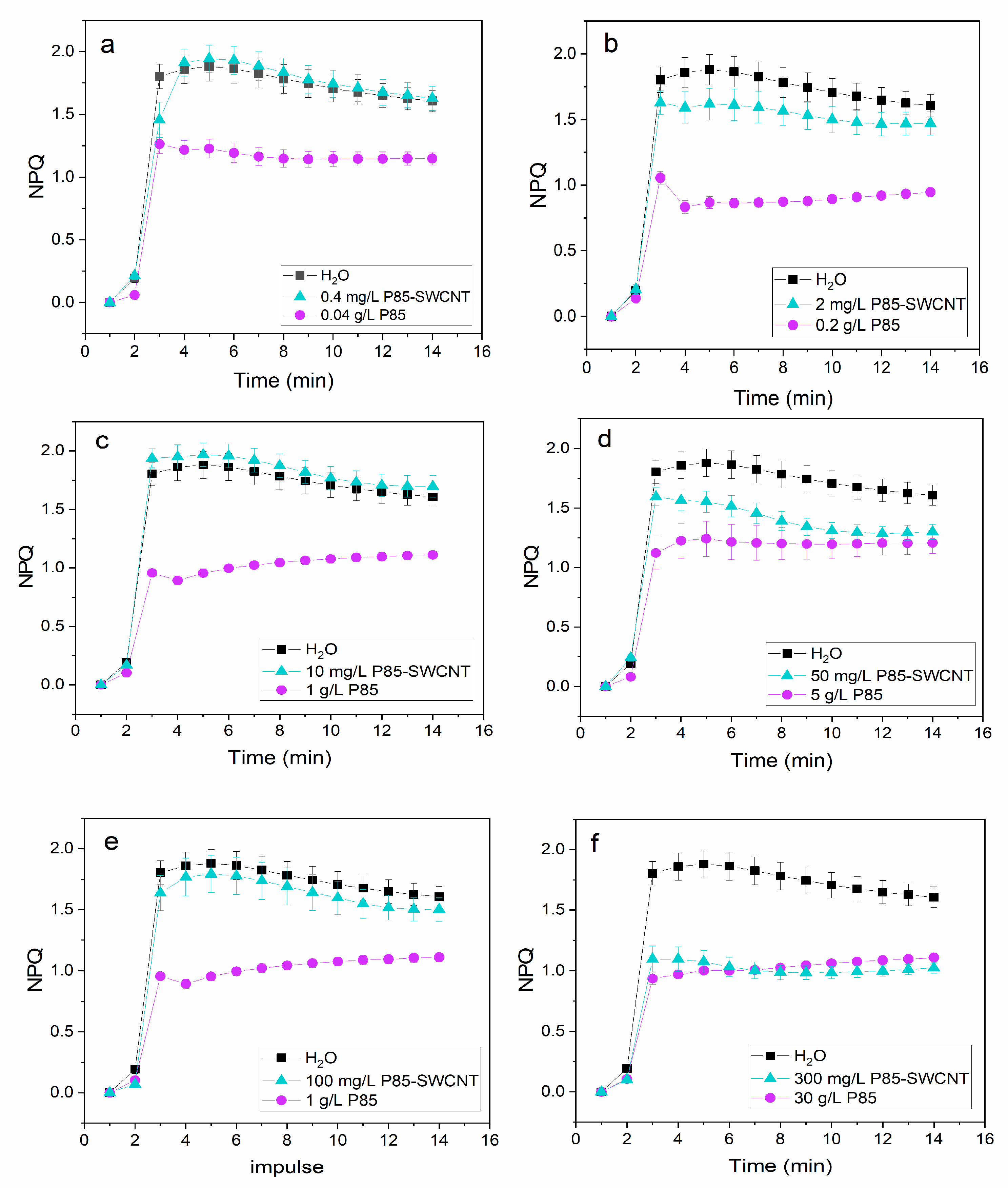
3.4. Leaf Physiological Characteristics
4. Discussion
4.1. Advancements in Nanopriming as New Tool for Germination Improvement
4.2. Optimal Conditions for Pea Seed Priming with P85-SWCNT
4.3. Harmful Effects of Priming with P85 Polymer
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Mondal, S.; Bose, B. Seed Priming: An Interlinking technology between seeds, seed germination and seedling establishment. In Plant Reproductive Ecology—Recent Adwances; Rustagi, A., Chaudhry, B., Eds.; IntechOpen: London, UK, 2021; pp. 1–16. [Google Scholar]
- Chetyrkina, M.R.; Fedorov, F.S.; Nasibulin, A.G. In vitro toxicity of carbon nanotubes: A systematic Review. RSC Adv. 2022, 12, 16235–16256. [Google Scholar] [CrossRef] [PubMed]
- Nile, S.H.; Thiruvengadam, M.; Wang, Y.; Samynathan, R.; Shariati, M.A.; Rebezov, M.; Nile, A.; Sun, M.; Venkidasamy, B.; Xiao, J.; et al. Nano-priming as emerging seed priming technology for sustainable agriculture–recent developments and futureperspectives. J. Nanobiotechnol. 2022, 20, 254. [Google Scholar] [CrossRef] [PubMed]
- Safdar, M.; Kim, W.; Park, S.; Gwon, Y.; Kim, Y.O.; Kim, J. Engineering plants with carbon nanotubes: A sustainable agriculture approach. J. Nanobiotechnol. 2022, 20, 275. [Google Scholar] [CrossRef] [PubMed]
- Shelar, A.; Nile, S.H.; Singh, A.V.; Rothenstein, D.; Bill, J.; Xiao, J.; Chaskar, M.; Kai, G.; Patil, R. Recent advances in nano-enabled seed treatment strategies for sustainable agriculture: Challenges, risk assessment, and future perspectives. Nano-Micro Lett. 2023, 15, 54. [Google Scholar] [CrossRef]
- Yan, S.; Zhao, L.; Li, H.; Zhang, Q.; Tan, J.; Huang, M.; He, S.; Li, L. Single-walled carbon nanotubes selectively influence maize root tissue development accompanied by the change in the related gene expression. J. Hazard. Mater. 2013, 246-247, 110–118. [Google Scholar] [CrossRef]
- Haghighi, M.; da Silva, J.A.T. The effect of carbon nanotubes on the seed germination and seedling growth of four vegetable species. J. Crop Sci. Biotechnol. 2014, 17, 201–208. [Google Scholar] [CrossRef]
- Pourkhaloee, A.; Haghigh, M.; Saharkhiz, M.J.; Jouzi, H.; Doroodmand, M.M. Carbon nanotubes can promote seed germination via seed coat penetration. Seed Technol. 2011, 33, 155–169. [Google Scholar]
- Hatami, M.; Hadian, J.; Ghorbanpour, M. Mechanisms underlying toxicity and stimulatory role of single-walled carbon nanotubes in Hyoscyamus niger during drought stress simulated by polyethylene glycol. J. Hazard. Mater. 2017, 324, 306–320. [Google Scholar] [CrossRef]
- Cañas, J.E.; Long, M.; Nations, S.; Vadan, R.; Dai, L.; Luo, M.; Ambikapathi, R.; Lee, E.H.; Olszyk, D. Effects of functionalized and nonfunctionalized single-walled carbon nanotubes on root elongation of select crop species. Environ. Toxicol. Chem. 2008, 27, 1922–1931. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; Dervishi, E.; Mahmood, M.; Xu, Y.; Li, Z.; Watanabe, F.; Biris, A.S. Carbon nanotubes are able to penetrate plant seed Coat and dramatically affect seed germination and plant growth. ACS Nano 2009, 3, 3221–3227. [Google Scholar] [CrossRef]
- Lahiani, M.H.; Dervishi, E.; Chen, J.; Nima, Z.; Gaume, A.; Biris, A.S.; Khodakovskaya, M.V. Impact of carbon nanotube exposure to seeds of valuable crops. ACS Appl. Mater. Interfaces 2013, 5, 7965–7973. [Google Scholar] [CrossRef]
- Khodakovskaya, M.V.; Kim, B.S.; Kim, J.N.; Alimohammadi, M.; Dervishi, E.; Mustafa, T.; Cernigla, C.E. Carbon nanotubes as plant growth regulators: Effects on tomato growth, reproductive system, and soil microbial community. Small 2013, 9, 115–123. [Google Scholar] [CrossRef]
- Liu, Q.; Chen, B.; Wang, Q.; Shi, X.; Xiao, Z.; Lin, J.; Fang, X. Carbon nanotubes as molecular transporters for walled plant cells. Nano Lett. 2009, 9, 1007–1010. [Google Scholar] [CrossRef]
- Lew, T.T.S.; Wong, M.H.; Kwak, S.-Y.; Sinclair, R.; Koman, V.B.; Strano, M.S. Rational design principles for the transport and subcellular distribution of nanomaterials into plant protoplasts. Small 2018, 14, 1802086. [Google Scholar] [CrossRef] [PubMed]
- Giraldo, J.P.; Landry, M.P.; Faltermeier, S.M.; McNicholas, T.P.; Iverson, N.M.; Boghossian, A.A.; Reuel, N.F.; Hilmer, A.J.; Sen, F.; Brew, J.A.; et al. Plant nanobionics approach to augment photosynthesis and biochemical sensing. Nat. Mater. 2014, 13, 400–408. [Google Scholar] [CrossRef]
- Ghasemi-Kooch, M.; Dehestani, M. Interaction of photosynthetic pigments with single-walled carbon nanotube (15, 15): A molecular dynamics study. Adsorption 2018, 24, 43–51. [Google Scholar] [CrossRef]
- Dorogi, M.; Balint, Z.; Mikó, C.; Vileno, B.; Milas, M.; Hernadi, K.; Forró, L.; Varó, G.; Nagy, L. Stabilization effect of single-walled carbon nanotubes on the functioning of photosynthetic reaction centers. J. Phys. Chem. B 2006, 110, 21473–21479. [Google Scholar] [CrossRef]
- Mackowski, S. Hybrid nanostructures for efficient light harvesting. J. Phys. Condens. Matter 2010, 22, 193102. [Google Scholar] [CrossRef] [PubMed]
- Hajdu, K.; Szabó, T.; Magyar, M.; Bencsik, G.; Németh, Z.; Nagy, K.; Magrez, A.; Forró, L.; Váró, G.; Hernádi, K.; et al. Photosynthetic reaction center protein in nanostructures. Phys. Stat. Sol. B 2011, 248, 2700–2703. [Google Scholar] [CrossRef]
- Nagy, L.; Magyar, M.; Szabó, T.; Hajdu, K.; Giotta, L.; Dorogi, M.; Milano, F. Photosynthetic machineries in nano-systems. Curr. Protein Pept. Sci. 2014, 15, 363–373. [Google Scholar] [CrossRef]
- Wiwatowski, K.; Duzynska, A.; Swiniarski, M.; Szalkowski, M.; Zdrojek, M.; Judek, J.; Mackowski, S.; Kaminska, I. Energy transfer from natural photosynthetic complexes to single-wall carbon nanotubes. J. Lumin. 2016, 170, 855–859. [Google Scholar] [CrossRef]
- Orlanducci, S.; Fulgenzi, G.; Margonelli, A.; Rea, G.; Antal, T.K.; Lambreva, M.D. Mapping single walled carbon nanotubes in photosynthetic algae by single-cell confocal raman microscopy. Materials 2020, 13, 5121. [Google Scholar] [CrossRef] [PubMed]
- Antal, T.K.; Volgusheva, A.A.; Kukarskikh, G.G.; Lukashev, E.P.; Bulychev, A.A.; Margonelli, A.; Orlanducci, S.; Leo, G.; Cerri, L.; Tyystjärvi, E.; et al. Single-walled carbon nanotubes protect photosynthetic reactions in Chlamydomonas reinhardtii against photoinhibition. Plant Physiol. Biochem. 2022, 192, 298–307. [Google Scholar] [CrossRef] [PubMed]
- Lambreva, M.D.; Akhtar, P.; Sipka, G.; Margonelli, A.; Lambrev, P.H. Fluorescence quenching in thylakoid membranes induced by single-walled carbon nanotubes. Photochem. Photobiol. Sci 2023. Epub ahead of print. [Google Scholar] [CrossRef] [PubMed]
- Kwak, S.Y.; Lew, T.T.S.; Sweeney, C.J.; Koman, V.B.; Wong, M.H.; Bohmert-Tatarev, K.; Snell, K.D.; Seo, J.S.; Chua, N.H.; Strano, M.S. Chloroplast-selective gene delivery and expression in planta using chitosan-complexed singlewalled carbon nanotube carriers. Nat. Nanotechnol. 2019, 14, 447–455. [Google Scholar] [CrossRef]
- Velikova, V.; Petrova, N.; Kovács, L.; Petrova, A.; Koleva, D.; Tsonev, T.; Taneva, S.; Petrov, P.; Krumova, S. Single-walled carbon nanotubes modify leaf micromorphology, chloroplast ultrastructure and photosynthetic activity of pea plants. Int. J. Mol. Sci. 2021, 22, 4878. [Google Scholar] [CrossRef]
- Petrova, N.; Paunov, M.; Petrov, P.; Velikova, V.; Goltsev, V.; Krumova, S. Polymer-modified single-walled carbon nanotubes affect photosystem II photochemistry, intersystem electron transport carriers and photosystem I end acceptors in pea plants. Molecules 2021, 26, 5958. [Google Scholar] [CrossRef]
- Karkanis, A.; Ntatsi, G.; Kontopoulou, C.K.; Pristeri, A.; Bilalis, D.; Savvas, D. Field pea in European cropping systems: Adaptability, biological nitrogen fixation and cultivation practices. Not. Bot. Horti. Agrobo Cluj Napoca 2016, 44, 325–336. [Google Scholar] [CrossRef]
- Petrov, P.D.; Georgiev, G.L. Fabrication of super-macroporous nanocomposites by deposition of carbon nanotubes onto polymer cryogels. Eur. Polym. J. 2012, 48, 1366–1373. [Google Scholar] [CrossRef]
- Ranal, M.A.; de Santana, D.G.; Ferreira, W.R.; Mendes-Rodrigues, C. Calculating germination measurements and organizing spreadsheets. Rev. Bras. Botânica 2009, 32, 849–855. [Google Scholar] [CrossRef]
- Velikova, V.; Arena, C.; Izzo, L.G.; Tsonev, T.; Koleva, D.; Tattini, M.; Roeva, O.; De Maio, A.; Loreto, F. Functional and Structural Leaf Plasticity Determine Photosynthetic Performances during Drought Stress and Recovery in Two Platanus orientalis Populations from Contrasting Habitats. Int. J. Mol. Sci. 2020, 21, 3912. [Google Scholar] [CrossRef]
- Genty, B.; Briantais, J.M.; Baker, N.R. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence. Biochim. Biophys. Acta 1989, 990, 87–92. [Google Scholar] [CrossRef]
- Bilger, W.; Björkman, O. Temperature dependence of violaxanthin de-epoxidation and non-photochemical fluorescence quenching in intact leaves of Gossypium hirsutum L. and Malva parviflora L. Planta 1991, 184, 226–234. [Google Scholar] [CrossRef] [PubMed]
- Baker, N.R. Chlorophyll fluorescence: A probe of photosynthesis in vivo. Ann. Rev. Plant Biol. 2008, 59, 89–113. [Google Scholar] [CrossRef] [PubMed]
- Yousefi, S.; Kartoolinejad, D.; Naghdi, R. Effects of priming with multi-walled carbon nanotubes on seed physiological characteristics of hopbush (Dodonaea viscosa L.) under drought stress. Int. J. Environ. Res. 2017, 74, 528–539. [Google Scholar] [CrossRef]
- López-Vargas, E.R.; González-García, Y.; Pérez-Álvarez, M.; Cadenas-Pliego, G.; González-Morales, S.; Benavides-Mendoza, A.; Cabrera, R.I.; Juárez-Maldonado, A. Seed priming with carbon nanomaterials to modify the germination, growth, and antioxidant status of tomato seedlings. Agronomy 2020, 10, 639. [Google Scholar] [CrossRef]
- Wu, Q.; Fan, C.; Wang, H.; Han, Y.; Tai, F.; Wu, J.; Li, H.; He, R. Biphasic impacts of graphite-derived engineering carbon-based nanomaterials on plant performance: Effectiveness vs. nanotoxicity. Adv. Agrochem. 2023, in press. [Google Scholar] [CrossRef]
- Batrakova, E.V.; Kabanov, A.V. Pluronic block copolymers: Evolution of drug delivery concept from inert nanocarriers to biological response modifiers. J. Control. Release 2008, 130, 98–106. [Google Scholar] [CrossRef]
- Samal, A.; Mohanty, S.; Das, S.; Das, B.C.; Beura, J.K. Polymer coating of cowpea seeds (Vigna unguiculata) for improving its storability and performance. J. Pharm. Innov. 2021, 10, 592–597. [Google Scholar]
- Su, L.Q.; Li, J.G.; Xue, H.; Wang, X.F. Super absorbent polymer seed coatings promote seed germination and seedling growth of Caragana korshinskii in drought. J. Zhejiang Univ. Sci B. 2017, 18, 696–706. [Google Scholar] [CrossRef]
- Farias, B.V.; Pirzada, T.; Mathew, R.; Sit, T.L.; Opperman, C.; Khan, S.A. Electrospun polymer nanofibers as seed coatings forcrop protection. ACS Sustain. Chem. Eng. 2019, 7, 19848–19856. [Google Scholar] [CrossRef]
- Choong, M.F.; Lucas, P.W.; Ong, J.S.Y.; Pereira, B.; Tan, H.T.W.; Turner, I.M. Leaf fracture toughness and sclerophylly: Their correlations and ecological implications. New Phytol. 1992, 121, 597–610. [Google Scholar] [CrossRef]
- Wright, I.; Reich, P.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Onoda, Y.; Wright, I.J.; Evans, J.R.; Hikosaka, K.; Kitajima, K.; Niinemets, Ü.; Poorter, H.; Tosens, T.; Westoby, M. Physiological and structural tradeoffs underlying the leaf economics spectrum. New Phytol. 2017, 214, 1447–1463. [Google Scholar] [CrossRef]
- Vogelmann, T.C.; Martin, G. The functional significance of palisade tissue: Penetration of directional versus diffuse light. Plant Cell Environ. 1993, 16, 65–72. [Google Scholar] [CrossRef]
- Terashima, I.; Saeki, T. Light environment within a leaf. I. Optical properties of paradermal sections of Camellia leaves with special reference to differences in the optical properties of palisade and spongy tissues. Plant Cell Physiol. 1984, 24, 1493–1501. [Google Scholar] [CrossRef]
- Karabourniotis, G.; Liakopoulos, G.; Bresta, P.; Nikolopoulos, D. The Optical Properties of Leaf Structural Elements and Their Contribution to Photosynthetic Performance and Photoprotection. Plants 2021, 10, 1455. [Google Scholar] [CrossRef] [PubMed]
- Cui, M.; Vogelmann, T.C.; Smith, W.K. Chlorophyll and light gradients in sun and shade leaves of Spinacia oleracea. Plant Cell Environ. 1991, 14, 493–500. [Google Scholar] [CrossRef]
- Théroux-Rancourt, G.; Roddy, A.B.; Earles, J.M.; Gilbert, M.E.; Zwieniecki, M.A.; Boyce, C.K.; Tholen, D.; McElrone, A.J.; Simonin, K.A.; Brodersen, C.R. Maximum CO2 diffusion inside leaves is limited by the scaling of cell size and genome size. Proc. Biol. Sci. B. 2021, 288, 20203145. [Google Scholar] [CrossRef]
- Davis, P.A.; Caylor, S.; Whippo, C.W.; Hangarter, R.P. Changes in leaf optical properties associated with light-dependent chloroplast movements. Plant Cell Environ. 2011, 34, 2047–2059. [Google Scholar] [CrossRef]
- Matos, F.S.; Wolfgramm, R.; Goncёalves, F.V.; Cavatte, P.C.; Ventrella, M.C.; DaMatta, F.M. Phenotypic plasticity in response to light in the coffee tree. Environ. Exp. Bot. 2009, 67, 421–427. [Google Scholar] [CrossRef]
- Kalmatskaya, O.A.; Karavaev, V.A.; Tikhonov, A.N. Slow induction of chlorophyll a fluorescence excited by blue and red light in Tradescantia leaves acclimated to high and low light. Photosynth. Res. 2019, 142, 265–282. [Google Scholar] [CrossRef] [PubMed]
- Kromdijk, J.; Głowacka, K.; Leonelli, L.; Gabilly, S.T.; Iwai, M.; Niyogi, K.K.; Long, S.P. Improving photosynthesis and crop productivity by accelerating recovery from photoprotection. Science 2016, 354, 857–861. [Google Scholar] [CrossRef] [PubMed]
- De Souza, A.P.; Burgess, S.J.; Doran, L.; Hansen, J.; Manukyan, L.; Maryn, N.; Gotarkar, D.; Leonelli, L.; Niyogi, K.K.; Long, S.P. Soybean photosynthesis and crop yield are improved by accelerating recovery from photoprotection. Science 2022, 377, 851–854. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, D.; Mohapatra, S.; Dogra, V. Improving photosynthetic efficiency by modulating non-photochemical quenching. Trends Plant Sci. 2023, 28, 264–266. [Google Scholar] [CrossRef] [PubMed]
- Dziergowska, K.; Michalak, I. The role of nanoparticles in sustainable agriculture. In Smart Agrochemicals for Sustainable Agriculture; Chojnacka, K., Saeid, A., Eds.; Elsevier Inc.: Amsterdam, The Netherlands, 2022; pp. 225–278. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imbibition (%) | |||
|---|---|---|---|
| Concentration | 2 h | 4 h | 6 h |
| H2O (control) | |||
| 70 ± 5 | 105 ± 8 | 126 ± 6 | |
| (mg/L) | P85-SWCNT | ||
| 0.4 | 60 ± 8 | 94 ± 7 | 123 ± 10 |
| 2 | 67 ± 5 | 105 ± 4 | 129 ± 3 |
| 10 | 63 ± 3 | 100 ± 1 | 125 ± 1 |
| 50 | 59 ± 8 | 101 ± 11 | 124 ± 7 |
| 100 | 59 ± 8 | 103 ± 14 | 123 ± 11 |
| 300 | 54 ± 9 | 96 ± 12 | 120 ± 10 |
| (g/L) | P85 | ||
| 0.04 | 64 ± 11 | 96 ± 11 | 121 ± 12 |
| 0.2 | 75 ± 11 | 106 ± 10 | 127 ± 10 |
| 1 | 68 ± 13 | 95 ± 13 | 118 ± 9 |
| 5 | 72 ± 14 | 104 ± 18 | 122 ± 16 |
| 10 | 72 ± 12 | 106 ± 16 | 123 ± 15 |
| 30 | 71 ± 14 | 104 ± 19 | 121 ± 18 |
| Concentration | G (%) | MT (Day) | Z | MR (Seeds per Day) |
|---|---|---|---|---|
| H2O (control) | ||||
| 55 ± 5 | 2.15 ± 0.1 | 0.42 ± 0.04 | 0.47 ± 0.02 | |
| (mg/L) | P85-SWCNT | |||
| 0.4 | 52 ± 7 | 2.26 ± 0.06 | 0.38 ± 0.03 | 0.44 ± 0.01 |
| 2 | 58 ± 5 | 2.11 ± 0.10 | 0.42 ± 0.02 | 0.48 ± 0.02 |
| 10 | 54 ± 4 | 2.2 ± 0.05 | 0.46 ± 0.04 | 0.45 ± 0.01 |
| 50 | 53 ± 4 | 2.33 ± 0.17 | 0.55 ± 0.07 | 0.44 ± 0.03 |
| 100 | 42 ± 18 | 2.43 ± 0.66 | 0.73 ± 0.10 ** | 0.45 ± 0.12 |
| 300 | 60 ± 6 | 2.31 ± 0.62 | 0.44 ± 0.12 | 0.47 ± 0.13 |
| (g/L) | P85 | |||
| 0.04 | 35 ± 13 | 2.43 ± 0.39 | 0.39 ± 0.08 | 0.42 ± 0.07 |
| 0.2 | 35 ± 7 | 2.61 ± 0.46 | 0.34 ± 0.06 | 0.40 ± 0.07 |
| 1 | 42 ± 14 | 2.66 ± 0.41 | 0.35 ± 0.06 | 0.39 ± 0.06 |
| 5 | 32 ± 22 | 2.09 ± 0.31 | 0.44 ± 0.04 | 0.49 ± 0.07 |
| 10 | 34 ± 18 | 2.06 ± 0.56 | 0.45 ± 0.01 | 0.52 ± 0.14 |
| 30 | 27 ± 23 | 2.40 ± 0.60 | 0.74 ± 0.26 | 0.44 ± 0.11 |
| Concentration | Leaf Thickness (µm) | Adaxial and Abaxial Epidermis Thickness (µm) | Mesophyll Thickness (µm) | Palisade Parenchyma Thickness (µm) | Spongy Parenchyma Thickness (µm) |
|---|---|---|---|---|---|
| H2O (control) | |||||
| 268 ± 18 | 52 ± 7 | 215 ± 16 | 41 ± 7 | 173 ± 10 | |
| (mg/L) | P85-SWCNT | ||||
| 50 | 201 ± 14 ** | 49 ± 6 | 149 ± 14 ** | 38 ± 8 | 111 ± 15 *** |
| 100 | 209 ± 15 ** | 42 ± 5 | 161 ± 10 ** | 45 ± 9 | 122 ± 16 ** |
| 300 | 198 ± 12 *** | 37 ± 5 | 154 ± 11 *** | 44 ± 8 | 107 ± 18 ** |
| (g/L) | P85 | ||||
| 5 | 159 ± 11 *** | 26 ± 4 *** | 129 ± 9 *** | 46 ± 9 | 79 ± 16 *** |
| 10 | 165 ± 12 *** | 25 ± 3 *** | 145 ± 12 *** | 49 ± 5 | 98 ± 16 *** |
| 30 | 158 ± 11 *** | 26 ± 4 *** | 129 ± 9 *** | 38 ± 9 | 93 ± 17 *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Krumova, S.; Petrova, A.; Petrova, N.; Stoichev, S.; Ilkov, D.; Tsonev, T.; Petrov, P.; Koleva, D.; Velikova, V. Seed Priming with Single-Walled Carbon Nanotubes Grafted with Pluronic P85 Preserves the Functional and Structural Characteristics of Pea Plants. Nanomaterials 2023, 13, 1332. https://doi.org/10.3390/nano13081332
Krumova S, Petrova A, Petrova N, Stoichev S, Ilkov D, Tsonev T, Petrov P, Koleva D, Velikova V. Seed Priming with Single-Walled Carbon Nanotubes Grafted with Pluronic P85 Preserves the Functional and Structural Characteristics of Pea Plants. Nanomaterials. 2023; 13(8):1332. https://doi.org/10.3390/nano13081332
Chicago/Turabian StyleKrumova, Sashka, Asya Petrova, Nia Petrova, Svetozar Stoichev, Daniel Ilkov, Tsonko Tsonev, Petar Petrov, Dimitrina Koleva, and Violeta Velikova. 2023. "Seed Priming with Single-Walled Carbon Nanotubes Grafted with Pluronic P85 Preserves the Functional and Structural Characteristics of Pea Plants" Nanomaterials 13, no. 8: 1332. https://doi.org/10.3390/nano13081332
APA StyleKrumova, S., Petrova, A., Petrova, N., Stoichev, S., Ilkov, D., Tsonev, T., Petrov, P., Koleva, D., & Velikova, V. (2023). Seed Priming with Single-Walled Carbon Nanotubes Grafted with Pluronic P85 Preserves the Functional and Structural Characteristics of Pea Plants. Nanomaterials, 13(8), 1332. https://doi.org/10.3390/nano13081332