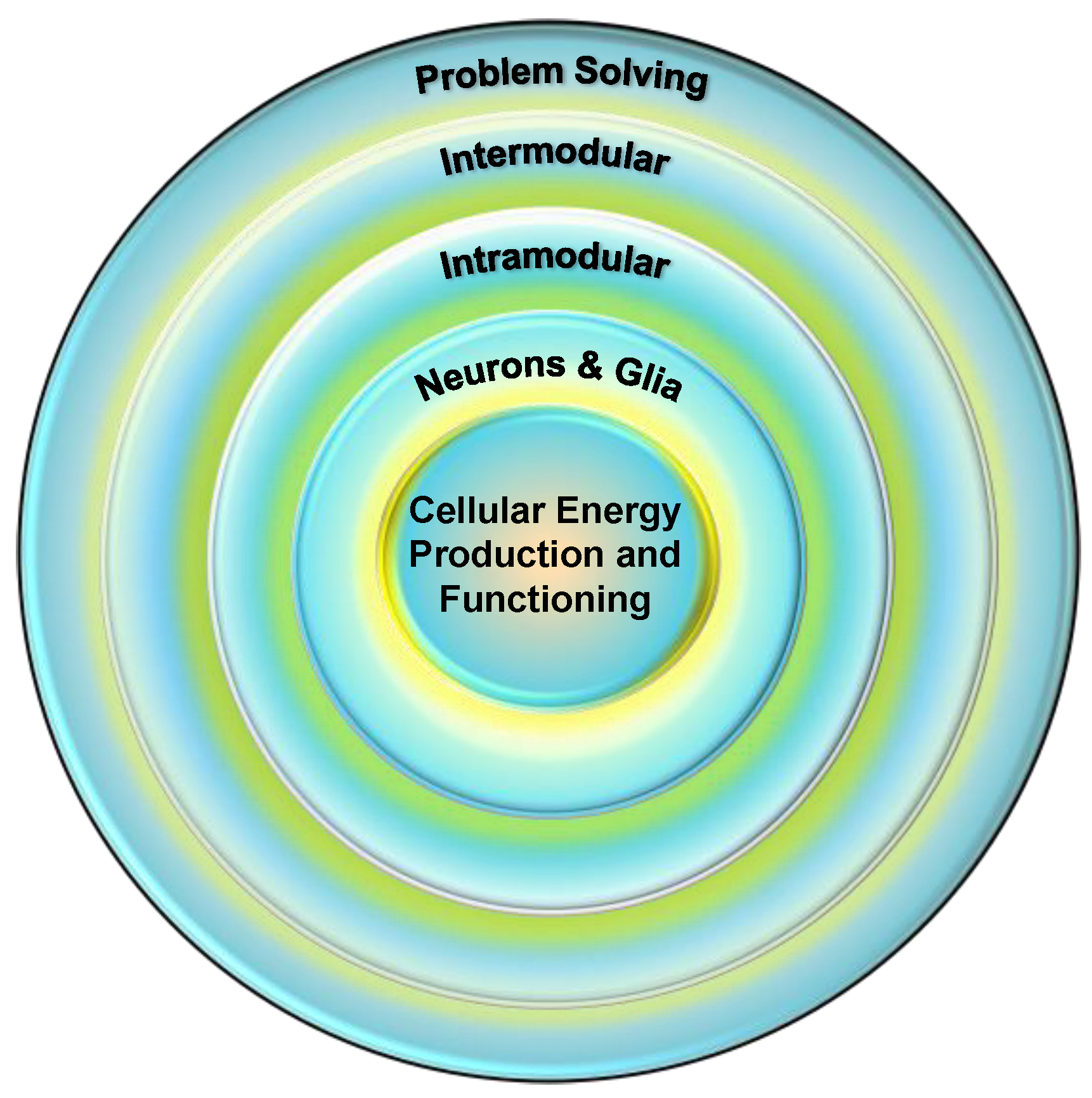
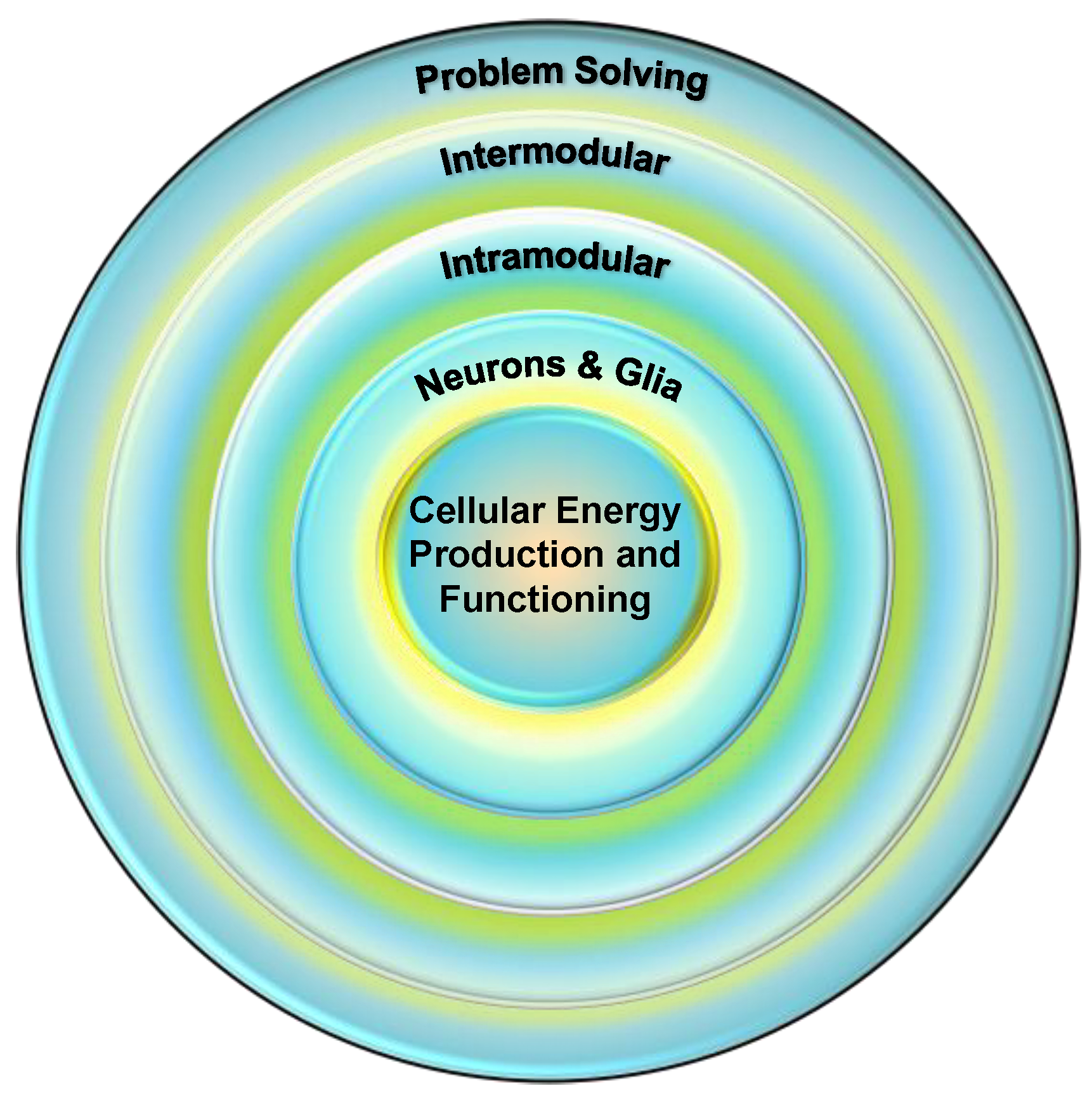
Mitochondria as the Linchpin of General Intelligence and the Link between g, Health, and Aging


{kind=link}
Abstract
:Funding
Conflicts of Interest
References
- Spearman, C. General Intelligence, Objectively Determined and Measured. Am. J. Psychol. 1904, 15, 201–292. [Google Scholar] [CrossRef]
- Warne, R.T.; Burningham, C. Spearman’s g found in 31 non-Western nations: Strong Evidence that g is a Universal Phenomenon. Psychol. Bull. 2019, 145, 237–272. [Google Scholar] [CrossRef] [PubMed]
- Engle, R.W.; Kane, M.J.; Tuholski, S.W. Individual differences in working memory capacity and what they tell us about controlled attention, General Fluid Intelligence, and Functions of the Prefrontal Cortex. In Models of Working Memory: Mechanisms of Active Maintenance and Executive; Miyake, A., Shah, P., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 102–134. [Google Scholar]
- Kranzler, J.H.; Jensen, A.R. The nature of psychometric g: Unitary Process or A Number of Independent Processes? Intelligence 1991, 15, 397–422. [Google Scholar] [CrossRef]
- Jung, R.E.; Haier, R.J. The Parieto-Frontal Integration Theory (P-FIT) of intelligence: Converging Neuroimaging Evidence. Behav. Brain Sci. 2007, 30, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Gottfredson, L.S.; Deary, I.J. Intelligence Predicts Health and Longevity, but why? Curr. Dir. Psychol. Sci. 2004, 13, 1–4. [Google Scholar] [CrossRef]
- Hendrickson, D.E.; Hendrickson, A.E. The Biological Basis of Individual Differences in Intelligence. Personal. Individ. Differ. 1980, 1, 3–33. [Google Scholar] [CrossRef]
- Kovacs, K.; Conway, A.R. Process overlap theory: A Unified Account of the General Factor of Intelligence. Psychol. Inq. 2016, 27, 151–177. [Google Scholar] [CrossRef]
- Krijnen, W.P. Positive Loadings and Factor Correlations from Positive Covariance Matrices. Psychometrika 2004, 69, 655–660. [Google Scholar] [CrossRef]
- Geary, D.C. Efficiency of Mitochondrial Functioning as the Fundamental Biological Mechanism of General Intelligence (g). Psychol. Rev. 2018, 125, 1028–1050. [Google Scholar] [CrossRef]
- Holyoak, K.J.; Thagard, P. The Analogical Mind. Am. Psychol. 1997, 52, 35–44. [Google Scholar] [CrossRef]
- Neubauer, A.C.; Fink, A. Intelligence and Neural Efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef] [PubMed]
- Santarnecchi, E.; Emmendorfer, A.; Pascual-Leone, A. Dissecting the parieto-frontal correlates of fluid intelligence: A Comprehensive ALE Meta-Analysis Study. Intelligence 2017, 63, 9–28. [Google Scholar] [CrossRef]
- Bullmore, E.; Sporns, O. The Economy of Brain Network Organization. Nat. Rev. Neurosci. 2012, 13, 336–349. [Google Scholar] [CrossRef] [PubMed]
- Tseng, P.Y.; Lee, I.H.; Chen, K.C.; Chen, P.S.; Chiu, N.T.; Yao, W.J.; Chu, C.L.; Yeh, T.L.; Yang, Y.K. The Correlation between Mid-Brain Serotonin Transporter Availability and Intelligence Quotient in Healthy Volunteers. Eur. Psychiatry 2015, 30, 193–197. [Google Scholar] [CrossRef] [PubMed]
- Lane, N. Mitonuclear match: Optimizing Fitness and Fertility over Generations Drives Ageing within Generations. Bioessays 2011, 33, 860–869. [Google Scholar] [CrossRef]
- Barbey, A.K.; Colom, R.; Solomon, J.; Krueger, F.; Forbes, C.; Grafman, J. An Integrative Architecture for General Intelligence and Executive Function Revealed by Lesion Mapping. Brain 2012, 135, 1154–1164. [Google Scholar] [CrossRef]
- Lee, J.J.; McGue, M.; Iacono, W.G.; Michael, A.M.; Chabris, C.F. The causal influence of brain size on human intelligence: Evidence from Within–Family Phenotypic Associations and GWAS Modeling. Intelligence 2019, 75, 48–58. [Google Scholar] [CrossRef]
- Savage, J.E.; Jansen, P.R.; Stringer, S.; Watanabe, K.; Bryois, J.; De Leeuw, C.A.; Nagel, M.; Awasthi, S.; Barr, P.B.; Coleman, J.R.; et al. Genome-Wide Association Meta-Analysis in 269,867 Individuals Identifies New Genetic and Functional links to Intelligence. Nat. Genet. 2018, 50, 912–919. [Google Scholar] [CrossRef]
- Latorre-Pellicer, A.; Moreno-Loshuertos, R.; Lechuga-Vieco, A.V.; Sánchez-Cabo, F.; Torroja, C.; Acín-Pérez, R.; Calvo, E.; Aix, E.; González–Guerra, A.; Bernad-Miana, M.L.; et al. Mitochondrial and Nuclear DNA Matching Shapes Metabolism and Healthy Ageing. Nature 2016, 535, 561–565. [Google Scholar] [CrossRef]
- Caito, S.W.; Aschner, M. Mitochondrial Redox Dysfunction and Environmental Exposures. Antioxid. Redox Signal. 2015, 23, 578–595. [Google Scholar] [CrossRef]
- Harman, D. The biologic clock: The Mitochondria? J. Am. Geriatr. Soc. 1972, 20, 145–147. [Google Scholar] [CrossRef] [PubMed]
- Koch, R.E.; Josefson, C.C.; Hill, G.E. Mitochondrial Function, Ornamentation, and Immunocompetence. Biol. Rev. 2017, 92, 1459–1474. [Google Scholar] [CrossRef] [PubMed]
- Stewart, J.B.; Freyer, C.; Elson, J.L.; Larsson, N.G. Purifying Selection of mtDNA and its Implications for Understanding Evolution and Mitochondrial Disease. Nat. Rev. Genet. 2008, 9, 657–662. [Google Scholar] [CrossRef] [PubMed]
- Deary, I.J.; Weiss, A.; Batty, G.D. Intelligence and personality as predictors of illness and death: How Researchers in Differential Psychology and Chronic Disease Epidemiology are Collaborating to Understand and Address Health Inequalities. Psychol. Sci. Public Interest 2010, 11, 53–79. [Google Scholar] [CrossRef] [PubMed]
- Lindenberger, U.; Baltes, P.B. Sensory functioning and intelligence in old age: A Strong Connection. Psychol. Aging 1994, 9, 339–355. [Google Scholar] [CrossRef] [PubMed]
- Lubinski, D.; Humphreys, L.G. Some Bodily and Medical Correlates of Mathematical Giftedness and Commensurate Levels of Socioeconomic Status. Intelligence 1992, 16, 99–115. [Google Scholar] [CrossRef]
- Salthouse, T.A. How Many Causes are there of Aging-Related Decrements in Cognitive Functioning? Dev. Rev. 1994, 14, 413–437. [Google Scholar] [CrossRef]
- Tucker-Drob, E.M.; Brandmaier, A.M.; Lindenberger, U. Coupled cognitive changes in adulthood: A Meta-Analysis. Psychol. Bull. 2019, 145, 273–301. [Google Scholar] [CrossRef]
© 2019 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Geary, D.C. Mitochondria as the Linchpin of General Intelligence and the Link between g, Health, and Aging. J. Intell. 2019, 7, 25. https://doi.org/10.3390/jintelligence7040025
Geary DC. Mitochondria as the Linchpin of General Intelligence and the Link between g, Health, and Aging. Journal of Intelligence. 2019; 7(4):25. https://doi.org/10.3390/jintelligence7040025
Chicago/Turabian StyleGeary, David C. 2019. "Mitochondria as the Linchpin of General Intelligence and the Link between g, Health, and Aging" Journal of Intelligence 7, no. 4: 25. https://doi.org/10.3390/jintelligence7040025
APA StyleGeary, D. C. (2019). Mitochondria as the Linchpin of General Intelligence and the Link between g, Health, and Aging. Journal of Intelligence, 7(4), 25. https://doi.org/10.3390/jintelligence7040025