
Intelligence as a Developing Function: A Neuroconstructivist Approach

Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
“Intelligence constitutes the state of equilibrium towards which tend all the successive adaptations of a sensori-motor and cognitive nature, as well as all assimilatory and accommodatory interactions between the organism and the environment.”
2. Intelligence: General Caveats
2.1. On Some Definitions of Intelligence
2.2. The Importance of Intelligence Testing
3. Intelligence across Development: A Dynamic Concept
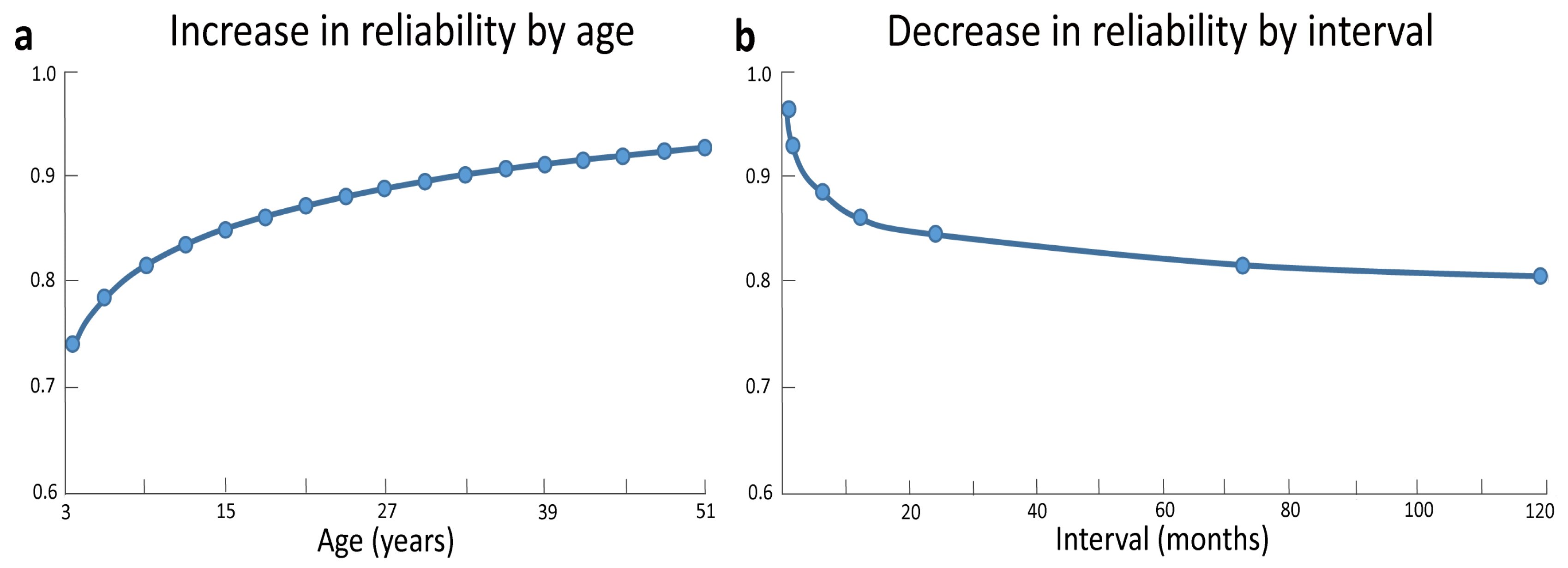
3.1. Intelligence Testing across Development
3.2. Intelligence Fluctuations across Development
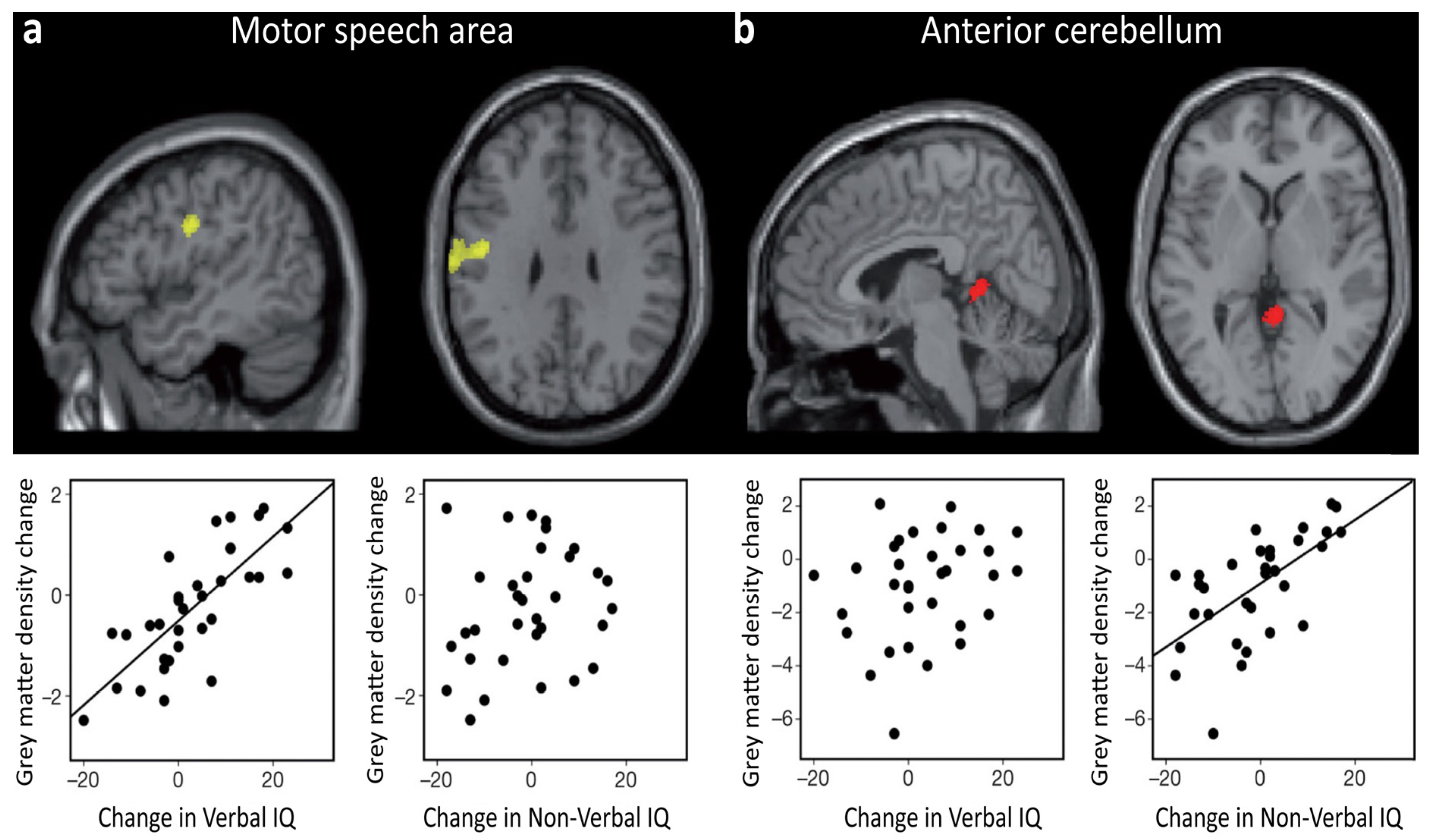
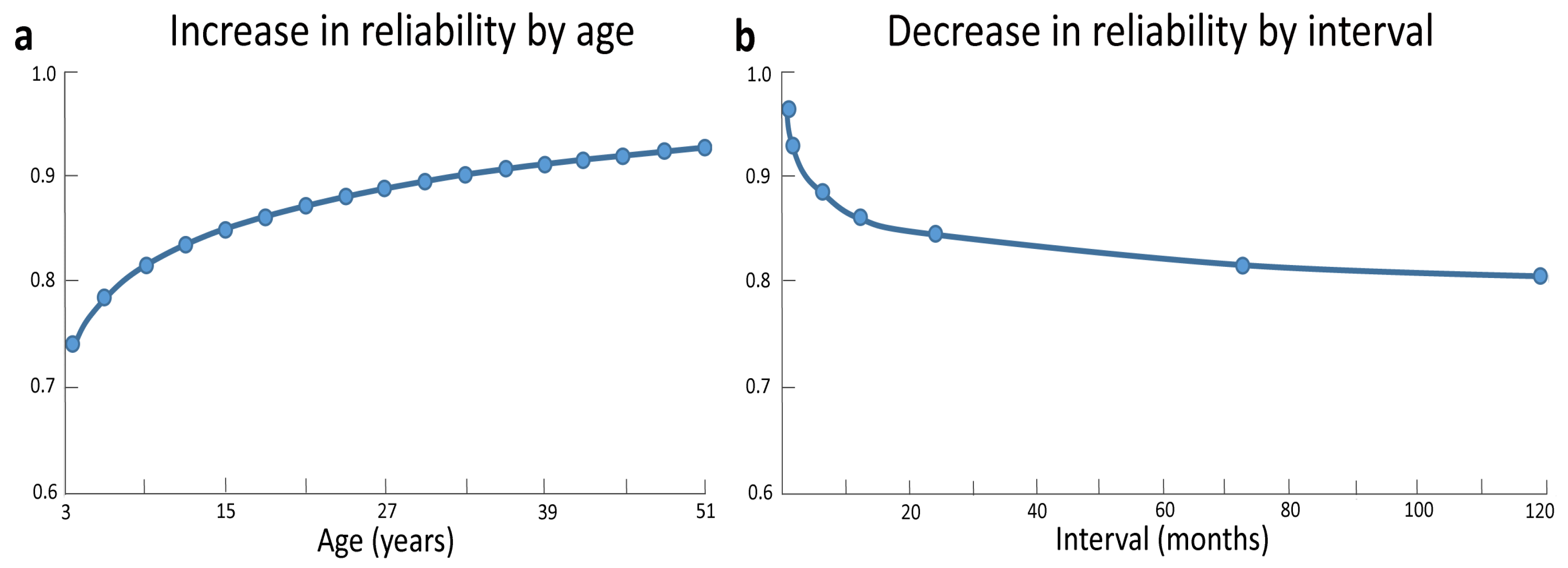
3.3. Neural Changes Associated with Intelligence across Development
3.4. Following Developmental Trajectories
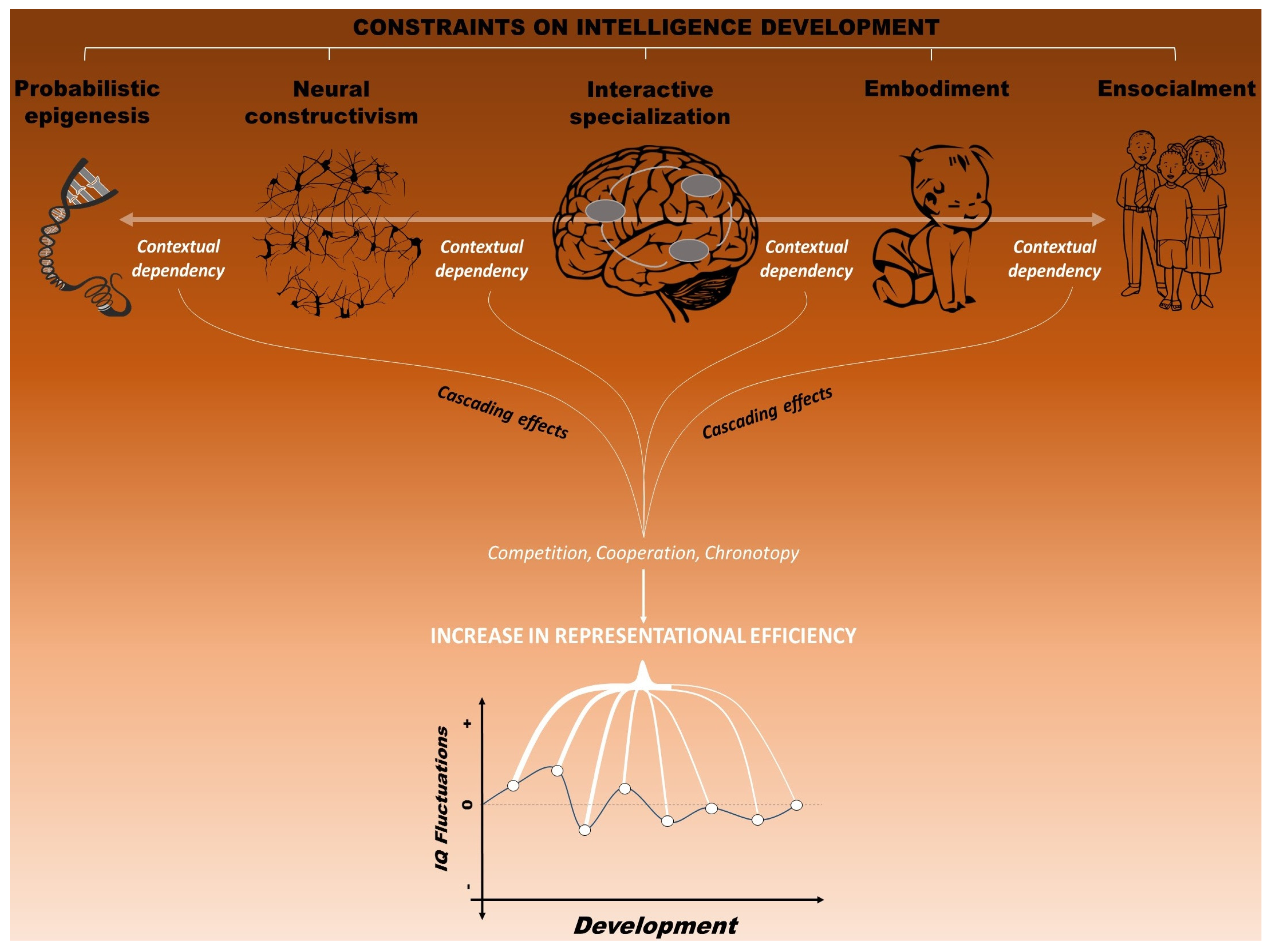
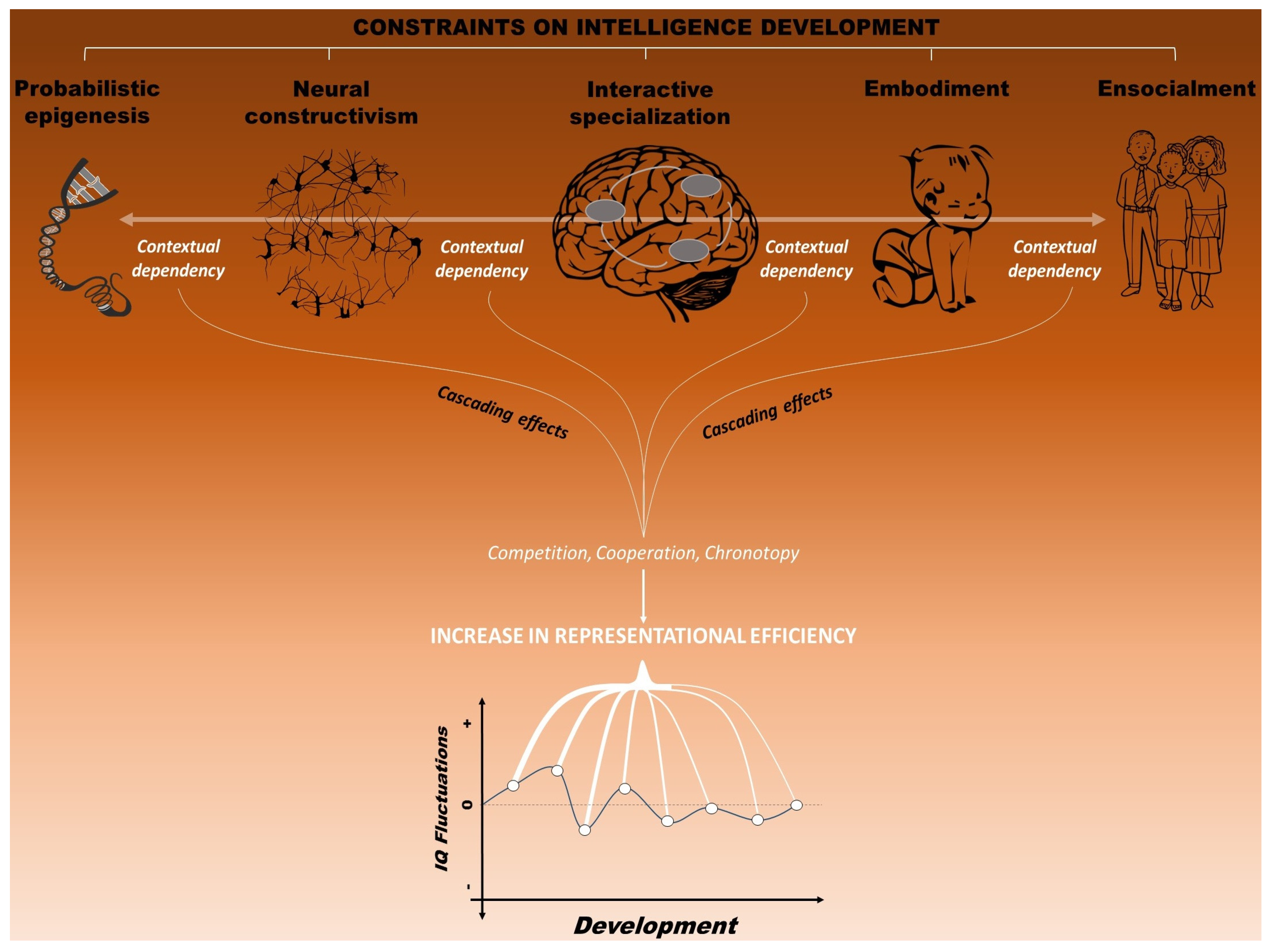
4. A Neuroconstructivist Approach to Intelligence
4.1. Probabilistic Epigenesis
4.2. Neural Constructivism
4.3. Interactive Specialization
4.4. Embodiment
4.5. Ensocialment
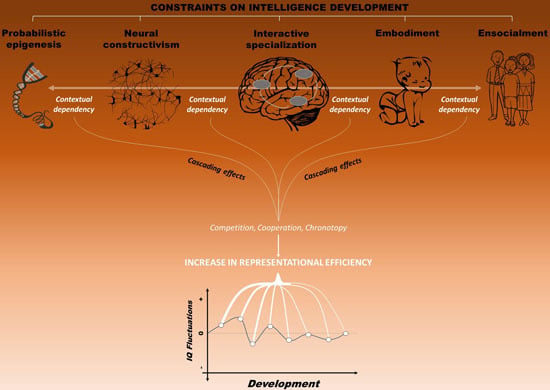
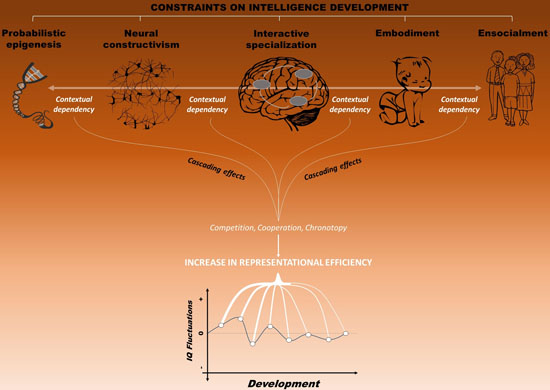
4.6. Toward an Integrated Framework: Reaching a Dynamic Balance
4.7. Predictions and Directions for Future Research
5. Cascading Effects on Life Outcome
6. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Piaget, J. La psychologie de l'intelligence; Armand Colin: Paris, France, 2012. [Google Scholar]
- Deary, I.J. Intelligence. Annu. Rev. Psychol. 2012, 63, 453–482. [Google Scholar] [CrossRef] [PubMed]
- Nisbett, R.E.; Aronson, J.; Blair, C.; Dickens, W.; Flynn, J.; Halpern, D.F.; Turkheimer, E. Intelligence: New findings and theoretical developments. Am. Psychol. 2012, 67, 130–159. [Google Scholar] [CrossRef] [PubMed]
- Kamphaus, R.W. Clinical Assessment of Child and Adolescent Intelligence; Springer Science + Business Media: Berlin, Germany, 2005. [Google Scholar]
- Baron, I.S.; Leonberger, K.A. Assessment of intelligence in the preschool period. Neuropsychol. Rev. 2012, 22, 334–344. [Google Scholar] [CrossRef] [PubMed]
- Westermann, G.; Mareschal, D.; Johnson, M.H.; Sirois, S.; Spratling, M.W.; Thomas, M.S.C. Neuroconstructivism. Dev. Sci. 2007, 10, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Mareschal, D. Neuroconstructivism: How the Brain Constructs Cognition; Oxford University Press: Oxford, UK, 2007; Volume 1. [Google Scholar]
- Karmiloff-Smith, A. An alternative to domain-general or domain-specific frameworks for theorizing about human evolution and ontogenesis. AIMS Neurosci. 2015, 2, 91–104. [Google Scholar] [CrossRef] [PubMed]
- Neisser, U.; Boodoo, G.; Bouchard, T.J., Jr.; Boykin, A.W.; Brody, N.; Ceci, S.J.; Halpern, D.F.; Loehlin, J.C.; Perloff, R.; Sternberg, R.J. Intelligence: Knowns and unknowns. Am. Psychol. 1996, 51, 77–101. [Google Scholar] [CrossRef]
- Sternberg, R.J. Practical Intelligence in Everyday Life; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Spearman, C. “General Intelligence,” objectively determined and measured. Am. J. Psychol. 1904, 15, 201–292. [Google Scholar] [CrossRef]
- Gray, J.R.; Thompson, P.M. Neurobiology of intelligence: Science and ethics. Nat. Rev. Neurosci. 2004, 5, 471–482. [Google Scholar] [CrossRef] [PubMed]
- Cattell, R.B. Theory of fluid and crystallized intelligence: A critical experiment. J. Educ. Psychol. 1963, 54, 1–22. [Google Scholar] [CrossRef]
- Cattell, R.B. Abilities: Their Structure, Growth, and Action; Houghton Mifflin: Boston, MA, USA, 1971. [Google Scholar]
- Kane, M.J.; Engle, R.W. The role of prefrontal cortex in working-memory capacity, executive attention, and general fluid intelligence: An individual-differences perspective. Psychon. Bull. Rev. 2002, 9, 637–671. [Google Scholar] [CrossRef] [PubMed]
- Deary, I.J. Human intelligence differences: Towards a combined experimental–differential approach. Trends Cogn. Sci. 2001, 5, 164–170. [Google Scholar] [CrossRef]
- Waltz, J.A.; Knowlton, B.J.; Holyoak, K.J.; Boone, K.B.; Mishkin, F.S.; de Menezes Santos, M.; Thomas, C.R.; Miller, B.L. A system for relational reasoning in human prefrontal cortex. Psychol. Sci. 1999, 10, 119–125. [Google Scholar] [CrossRef]
- Jung, R.E.; Haier, R.J. The Parieto-Frontal Integration Theory (P-FIT) of intelligence: Converging neuroimaging evidence. Behav. Brain Sci. 2007, 30, 135–154. [Google Scholar] [CrossRef] [PubMed]
- Moffitt, T.E.; Caspi, A.; Harkness, A.R.; Silva, P.A. The Natural History of Change to Intellectual Performance: Who Changes? How Much? Is it Meaningful? J. Child Psychol. Psychiatry 1993, 34, 455–506. [Google Scholar] [CrossRef] [PubMed]
- Deary, I.J.; Penke, L.; Johnson, W. The neuroscience of human intelligence differences. Nat. Rev. Neurosci. 2010, 11, 201–211. [Google Scholar] [CrossRef] [PubMed]
- Wechsler, D. Wechsler Intelligence Scale for Children; Psychological Corporation: San Antonio, TX, USA, 1949. [Google Scholar]
- Raven, J. The Raven’s progressive matrices: Change and stability over culture and time. Cogn. Psychol. 2000, 41, 1–48. [Google Scholar] [CrossRef] [PubMed]
- Evers, A.; Muñiz, J.; Bartram, D.; Boben, D.; Egeland, J.; Fernández-Hermida, J.R.; Frans, Ö.; Gintiliené, G.; Hagemeister, C.; Halama, P. Testing practices in the 21st century. Eur. Psychol. 2012, 17, 300–319. [Google Scholar] [CrossRef]
- Sattler, J.M. Assessment of Children: Cognitive Foundations; JM Sattler: San Diego, CA, USA, 2008. [Google Scholar]
- Bayley, N. Bayley Scales of Infant Development: Manual; Psychological Corporation: San Antonio, TX, USA, 1993. [Google Scholar]
- Wechsler, D. Wechsler Preschool and Primary Scale of Intelligence; Psychological Corporation: New York, NY, USA, 1967. [Google Scholar]
- Haier, R.J. Increased intelligence is a myth (so far). Front. Syst. Neurosci. 2014, 8, 34. [Google Scholar] [CrossRef] [PubMed]
- Baron, I.S. Neuropsychological Evaluation of the Child; Oxford University Press: Oxford, UK, 2004. [Google Scholar]
- Haywood, H.C.; Switzky, H.N. The malleability of intelligence: Cognitive processes as a function of polygenic–experiential interaction. Sch. Psychol. Rev. 1986, 15, 245–255. [Google Scholar]
- Valsiner, J. Conceptualizing intelligence: From an internal static attribution to the study of the process structure of organism-environment relationships. Int. J. Psychol. 1984, 19, 363–389. [Google Scholar] [CrossRef]
- Baltes, P.B.; Staudinger, U.M. Lifespan psychology: Theory and application to intellectual functioning. Annu. Rev. Psychol. 1999, 50, 471–507. [Google Scholar] [CrossRef] [PubMed]
- Horn, J.L.; Cattell, R.B. Refinement and test of the theory of fluid and crystallized general intelligences. J. Educ. Psychol. 1966, 57, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Salthouse, T.A. When does age-related cognitive decline begin? Neurobiol. Aging 2009, 30, 507–514. [Google Scholar] [CrossRef] [PubMed]
- Watkins, M.W.; Smith, L.G. Long-term stability of the Wechsler Intelligence Scale for Children—Fourth Edition. Psychol. Assess. 2013, 25, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Bishop, E.G.; Cherny, S.S.; Corley, R.; Plomin, R.; DeFries, J.C.; Hewitt, J.K. Development genetic analysis of general cognitive ability from 1 to 12 years in a sample of adoptees, biological siblings, and twins. Intelligence 2003, 31, 31–49. [Google Scholar] [CrossRef]
- McCall, R.B.; Hogarty, P.S.; Hurlburt, N. Transitions in infant sensorimotor development and the prediction of childhood IQ. Am. Psychol. 1972, 27, 728–748. [Google Scholar] [CrossRef] [PubMed]
- Johnson, W.; Gow, A.J.; Corley, J.; Starr, J.M.; Deary, I.J. Location in cognitive and residential space at age 70 reflects a lifelong trait over parental and environmental circumstances: The Lothian Birth Cohort 1936. Intelligence 2010, 38, 402–411. [Google Scholar] [CrossRef]
- Chen, Z.; Siegler, R.S. Across the great divide: Bridging the gap between understanding of toddlers’ and older children’s thinking. Monogr. Soc. Res. Child Dev. 2000, 65, 1–96. [Google Scholar] [CrossRef]
- Deary, I.J. The stability of intelligence from childhood to old age. Curr. Dir. Psychol. Sci. 2014, 23, 239–245. [Google Scholar] [CrossRef]
- Schuerger, J.M.; Witt, A.C. The temporal stability of individually tested intelligence. J. Clin. Psychol. 1989, 45, 294–302. [Google Scholar] [CrossRef]
- Waber, D.P.; Forbes, P.W.; Almli, C.R.; Blood, E.A. Four-year longitudinal performance of a population-based sample of healthy children on a neuropsychological battery: The NIH MRI study of normal brain development. J. Int. Neuropsychol. Soc. 2012, 18, 179–190. [Google Scholar] [CrossRef] [PubMed]
- Gottfried, A.W.; Gottfried, A.E.; Guerin, D.W. The Fullerton Longitudinal Study: A long-term investigation of intellectual and motivational giftedness. J. Educ. Gift. 2006, 29, 430–450. [Google Scholar] [CrossRef]
- Schneider, W.; Niklas, F.; Schmiedeler, S. Intellectual development from early childhood to early adulthood: The impact of early IQ differences on stability and change over time. Learn. Individ. Differ. 2014, 32, 156–162. [Google Scholar] [CrossRef]
- Deary, I.J.; Whalley, L.J.; Lemmon, H.; Crawford, J.R.; Starr, J.M. The stability of individual differences in mental ability from childhood to old age: Follow-up of the 1932 Scottish Mental Survey. Intelligence 2000, 28, 49–55. [Google Scholar] [CrossRef]
- Flynn, J.R. What Is Intelligence?: Beyond the Flynn Effect; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Canivez, G.L.; Watkins, M.W. Long-term stability of the Wechsler Intelligence Scale for Children—Third Edition. Psychol. Assess. 1998, 10, 285–291. [Google Scholar] [CrossRef]
- Oakman, S.; Wilson, B. Stability of WISC-R intelligence scores: Implications for 3-year reevaluations of learning disabled students. Psychol. Sch. 1988, 25, 118–120. [Google Scholar] [CrossRef]
- Mortensen, E.L.; Andresen, J.; Kruuse, E.; Sanders, S.A.; Reinisch, J.M. IQ stability: The relation between child and young adult intelligence test scores in low-birthweight samples. Scand. J. Psychol. 2003, 44, 395–398. [Google Scholar] [CrossRef] [PubMed]
- Schneider, W.; Wolke, D.; Schlagmüller, M.; Meyer, R. Pathsways to school achievement in very preterm and full term children. Eur. J. Psychol. Educ. 2004, 19, 385–406. [Google Scholar] [CrossRef]
- Balinsky, B. An analysis of the mental factors of various age groups from nine to sixty. Genet. Psychol. Monogr. 1941, 23, 191–234. [Google Scholar]
- Li, S.-C.; Lindenberger, U.; Hommel, B.; Aschersleben, G.; Prinz, W.; Baltes, P.B. Transformations in the couplings among intellectual abilities and constituent cognitive processes across the life span. Psychol. Sci. 2004, 15, 155–163. [Google Scholar] [CrossRef] [PubMed]
- Garrett, H.E. A developmental theory of intelligence. Am. Psychol. 1946, 1, 372–378. [Google Scholar] [CrossRef] [PubMed]
- Baltes, P.B.; Cornelius, S.W.; Spiro, A.; Nesselroade, J.R.; Willis, S.L. Integration versus differentiation of fluid/crytallized intelligence in old age. Dev. Psychol. 1980, 16, 625–635. [Google Scholar] [CrossRef]
- Tucker-Drob, E.M. Differentiation of cognitive abilities across the life span. Dev. Psychol. 2009, 45, 1097–1118. [Google Scholar] [CrossRef] [PubMed]
- Dickens, W.T.; Flynn, J.R. Heritability estimates versus large environmental effects: The IQ paradox resolved. Psychol. Rev. 2001, 108, 346–369. [Google Scholar] [CrossRef] [PubMed]
- van der Maas, H.L.J.; Dolan, C.V.; Grasman, R.P.P.P.; Wicherts, J.M.; Huizenga, H.M.; Raijmakers, M.E.J. A dynamical model of general intelligence: The positive manifold of intelligence by mutualism. Psychol. Rev. 2006, 113, 842–861. [Google Scholar] [CrossRef] [PubMed]
- McDaniel, M.A. Big-brained people are smarter: A meta-analysis of the relationship between in vivo brain volume and intelligence. Intelligence 2005, 33, 337–346. [Google Scholar] [CrossRef]
- Goh, S.; Bansal, R.; Xu, D.; Hao, X.; Liu, J.; Peterson, B.S. Neuroanatomical correlates of intellectual ability across the life span. Dev. Cogn. Neurosci. 2011, 1, 305–312. [Google Scholar] [CrossRef] [PubMed]
- Narr, K.L.; Woods, R.P.; Thompson, P.M.; Szeszko, P.; Robinson, D.; Dimtcheva, T.; Gurbani, M.; Toga, A.W.; Bilder, R.M. Relationships between IQ and regional cortical gray matter thickness in healthy adults. Cereb. Cortex 2007, 17, 2163–2171. [Google Scholar] [CrossRef] [PubMed]
- Penke, L.; Maniega, S.M.; Bastin, M.E.; Hernandez, M.C.V.; Murray, C.; Royle, N.A.; Starr, J.M.; Wardlaw, J.M.; Deary, I.J. Brain white matter tract integrity as a neural foundation for general intelligence. Mol. Psychiatry 2012, 17, 1026–1030. [Google Scholar] [CrossRef] [PubMed]
- Haier, R.J.; Siegel, B.V.; Nuechterlein, K.H.; Hazlett, E.; Wu, J.C.; Paek, J.; Browning, H.L.; Buchsbaum, M.S. Cortical glucose metabolic rate correlates of abstract reasoning and attention studied with positron emission tomography. Intelligence 1988, 12, 199–217. [Google Scholar] [CrossRef]
- van den Heuvel, M.P.; Stam, C.J.; Kahn, R.S.; Pol, H.E.H. Efficiency of functional brain networks and intellectual performance. J. Neurosci. 2009, 29, 7619–7624. [Google Scholar] [CrossRef] [PubMed]
- Reiss, A.L.; Abrams, M.T.; Singer, H.S.; Ross, J.L.; Denckla, M.B. Brain development, gender and IQ in children. Brain 1996, 119, 1763–1774. [Google Scholar] [CrossRef] [PubMed]
- Van Leeuwen, M.; Peper, J.S.; van den Berg, S.M.; Brouwer, R.M.; Pol, H.E.H.; Kahn, R.S.; Boomsma, D.I. A genetic analysis of brain volumes and IQ in children. Intelligence 2009, 37, 181–191. [Google Scholar] [CrossRef]
- Karama, S.; Colom, R.; Johnson, W.; Deary, I.J.; Haier, R.; Waber, D.P.; Lepage, C.; Ganjavi, H.; Jung, R.; Evans, A.C. Cortical thickness correlates of specific cognitive performance accounted for by the general factor of intelligence in healthy children aged 6 to 18. Neuroimage 2011, 55, 1443–1453. [Google Scholar] [CrossRef] [PubMed]
- Menary, K.; Collins, P.F.; Porter, J.N.; Muetzel, R.; Olson, E.A.; Kumar, V.; Steinbach, M.; Lim, K.O.; Luciana, M. Associations between cortical thickness and general intelligence in children, adolescents and young adults. Intelligence 2013, 41, 597–606. [Google Scholar] [CrossRef] [PubMed]
- Shaw, P.; Greenstein, D.; Lerch, J.; Clasen, L.; Lenroot, R.; Gogtay, N.; Evans, A.; Rapoport, J.; Giedd, J. Intellectual ability and cortical development in children and adolescents. Nature 2006, 440, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Rakic, P. Specification of cerebral cortical areas. Science 1988, 241, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Giedd, J.N.; Blumenthal, J.; Jeffries, N.O.; Castellanos, F.X.; Liu, H.; Zijdenbos, A.; Paus, T.; Evans, A.C.; Rapoport, J.L. Brain development during childhood and adolescence: A longitudinal MRI study. Nat. Neurosci. 1999, 2, 861–863. [Google Scholar] [CrossRef] [PubMed]
- Raznahan, A.; Lerch, J.P.; Lee, N.; Greenstein, D.; Wallace, G.L.; Stockman, M.; Clasen, L.; Shaw, P.W.; Giedd, J.N. Patterns of coordinated anatomical change in human cortical development: A longitudinal neuroimaging study of maturational coupling. Neuron 2011, 72, 873–884. [Google Scholar] [CrossRef] [PubMed]
- Schnack, H.G.; Van Haren, N.E.M.; Brouwer, R.M.; Evans, A.; Durston, S.; Boomsma, D.I.; Kahn, R.S.; Pol, H.E.H. Changes in thickness and surface area of the human cortex and their relationship with intelligence. Cereb. Cortex 2015, 25, 1608–1617. [Google Scholar] [CrossRef] [PubMed]
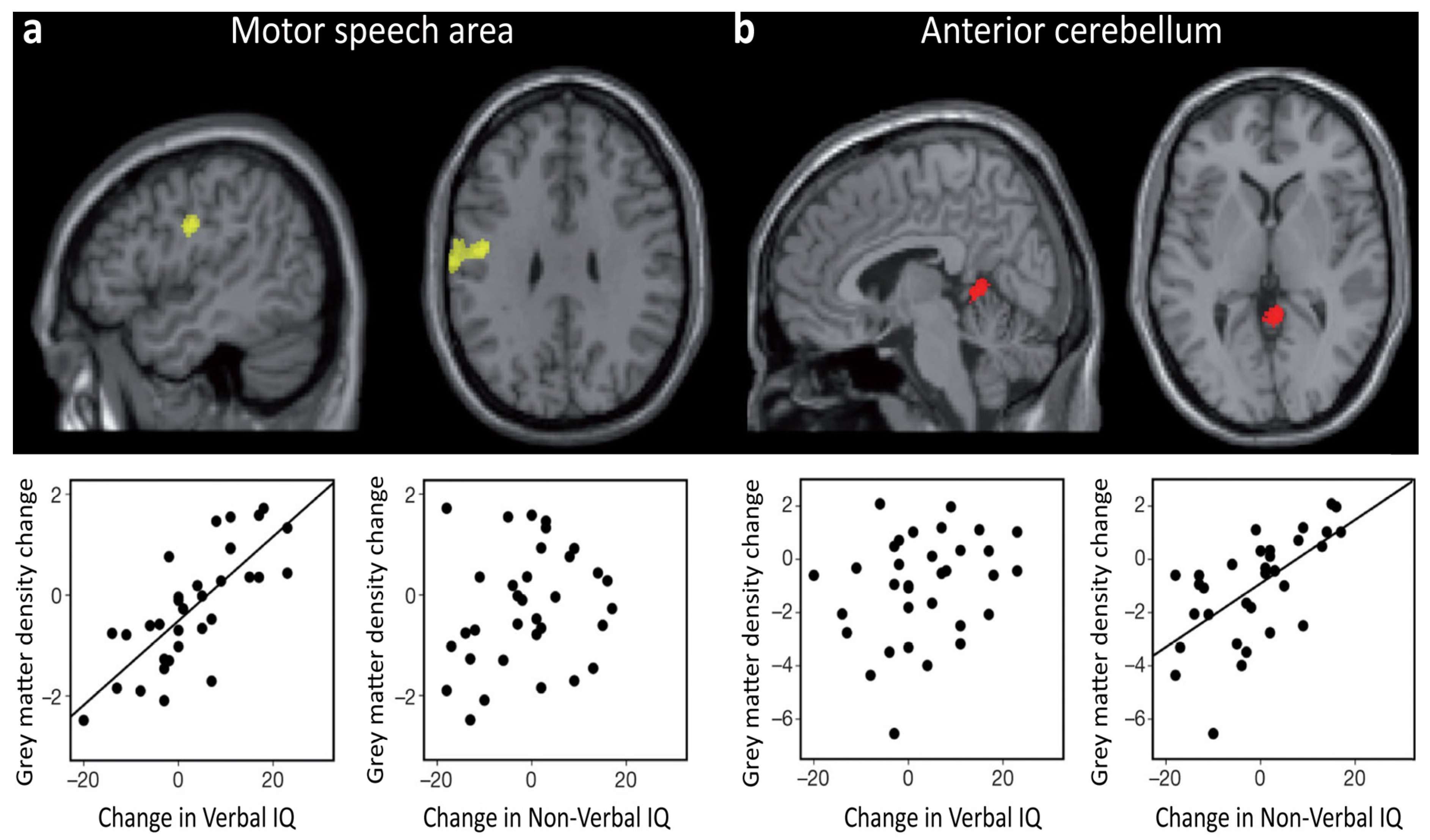
- Ramsden, S.; Richardson, F.M.; Josse, G.; Thomas, M.S.C.; Ellis, C.; Shakeshaft, C.; Seghier, M.L.; Price, C.J. Verbal and non-verbal intelligence changes in the teenage brain. Nature 2011, 479, 113–116. [Google Scholar] [CrossRef] [PubMed]
- Burgaleta, M.; Johnson, W.; Waber, D.P.; Colom, R.; Karama, S. Cognitive ability changes and dynamics of cortical thickness development in healthy children and adolescents. Neuroimage 2014, 84, 810–819. [Google Scholar] [CrossRef] [PubMed]
- Demetriou, A.; Spanoudis, G.; Shayer, M. Inference, reconceptualization, sight, and efficiency along intellectual growth: A general theory. Enfance 2014, 2014, 365–396. [Google Scholar] [CrossRef]
- Demetriou, A.; Shayer, M.; Efklides, A. Neo-Piagetian Theories of Cognitive Development: Implications and Applications for Education; Routledge: Abingdon, UK, 2016. [Google Scholar]
- Demetriou, A.; Spanoudis, G. Cognitive Abilities and Educational Outcomes; Springer: New York, NY, USA, 2017; pp. 39–60. [Google Scholar]
- Demetriou, A.; Spanoudis, G.; Shayer, M. Mapping Mind-Brain Development. In Neurotechnology Direct Brain Communication New Insights Responsibilities Concerning Speechless But Communicative Subjects; Routledge: New York, NY, USA, 2016; Volume 21. [Google Scholar]
- Demetriou, A.; Kyriakides, L. The functional and developmental organization of cognitive developmental sequences. Br. J. Educ. Psychol. 2006, 76, 209–242. [Google Scholar] [CrossRef] [PubMed]
- Demetriou, A.; Spanoudis, G.; Mouyi, A. Educating the Developing Mind: Towards an Overarching Paradigm. Educ. Psychol. Rev. 2011, 23, 601–663. [Google Scholar] [CrossRef]
- Demetriou, A.; Spanoudis, G.; Shayer, M.; Van der Ven, S.; Brydges, C.R.; Kroesbergen, E.; Podjarny, G.; Swanson, H.L. Relations between speed, working memory, and intelligence from preschool to adulthood: Structural equation modeling of 14 studies. Intelligence 2014, 46, 107–121. [Google Scholar] [CrossRef]
- Žebec, M.S.; Demetriou, A.; Kotrla-Topić, M. Changing expressions of general intelligence in development: A 2-wave longitudinal study from 7 to 18years of age. Intelligence 2015, 49, 94–109. [Google Scholar] [CrossRef]
- Spanoudis, G.; Demetriou, A.; Kazi, S.; Giorgala, K.; Zenonos, V. Embedding cognizance in intellectual development. J. Exp. Child Psychol. 2015, 132, 32–50. [Google Scholar] [CrossRef] [PubMed]
- Makris, N.; Tahmatzidis, D.; Demetriou, A.; Spanoudis, G. Mapping the evolving core of intelligence: Changing relations between executive control, reasoning, language, and awareness. Intelligence 2017. [Google Scholar] [CrossRef]
- Demetriou, A.; Spanoudis, G.; Shayer, M. Developmental intelligence: From empirical to hidden constructs. Intelligence 2013, 41, 744–749. [Google Scholar] [CrossRef]
- Demetriou, A.; Spanoudis, G.; Shayer, M.; Mouyi, A.; Kazi, S.; Platsidou, M. Cycles in speed-working memory-G relations: Towards a developmental–differential theory of the mind. Intelligence 2013, 41, 34–50. [Google Scholar] [CrossRef]
- Karmiloff-Smith, A. Development itself is the key to understanding developmental disorders. Trends Cogn. Sci. 1998, 2, 389–398. [Google Scholar] [CrossRef]
- Sirois, S.; Spratling, M.; Thomas, M.S.C.; Westermann, G.; Mareschal, D.; Johnson, M.H. Précis of neuroconstructivism: How the brain constructs cognition. Behav. Brain Sci. 2008, 31, 321–356. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, A.C.; Fink, A. Intelligence and neural efficiency. Neurosci. Biobehav. Rev. 2009, 33, 1004–1023. [Google Scholar] [CrossRef] [PubMed]
- Poldrack, R.A. Is “efficiency” a useful concept in cognitive neuroscience? Dev. Cogn. Neurosci. 2015, 11, 12–17. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, G. Probabilistic epigenesis. Dev. Sci. 2007, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Quartz, S.R.; Sejnowski, T.J. The neural basis of cognitive development: A constructivist manifesto. Behav. Brain Sci. 1997, 20, 537–556. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H.; De Haan, M. Developmental Cognitive Neuroscience: An Introduction; John Wiley & Sons: Hoboken, NJ, USA, 2015. [Google Scholar]
- Clark, A. An embodied cognitive science? Trends Cogn. Sci. 1999, 3, 345–351. [Google Scholar] [CrossRef]
- Smith, L.B. Cognition as a dynamic system: Principles from embodiment. Dev. Rev. 2005, 25, 278–298. [Google Scholar] [CrossRef]
- Bradley, R.H.; Corwyn, R.F. Socioeconomic status and child development. Annu. Rev. Psychol. 2002, 53, 371–399. [Google Scholar] [CrossRef] [PubMed]
- Meltzoff, A.N.; Kuhl, P.K.; Movellan, J.; Sejnowski, T.J. Foundations for a New Science of Learning. Science 2009, 325, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Westermann, G.; Thomas, M.S.C.; Karmiloff-Smith, A. Neuroconstructivism. In The Wiley-Blackwell Handbook of Childhood Cognitive Development, 2nd ed.; Wiley-Blackwell: Oxford, UK, 2010. [Google Scholar]
- Annaz, D.; Karmiloff-Smith, A.; Thomas, M.S.C. The importance of tracing developmental trajectories for clinical child neuropsychology. In Child Neuropsychology Concepts, Theory Practice; Wiley-Blackwell: Oxford, UK, 2008. [Google Scholar]
- Goswami, U. Blackwell Handbook of Childhood Cognitive Development; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]
- Lickliter, R.; Honeycutt, H. Developmental dynamics: Toward a biologically plausible evolutionary psychology. Psychol. Bull. 2003, 129, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Karmiloff-Smith, A.; D’Souza, D.; Dekker, T.M.; Van Herwegen, J.; Xu, F.; Rodic, M.; Ansari, D. Genetic and environmental vulnerabilities in children with neurodevelopmental disorders. Proc. Natl. Acad. Sci. USA 2012, 109 (Suppl. 2), 17261–17265. [Google Scholar] [CrossRef] [PubMed]
- Caspi, A.; Williams, B.; Kim-Cohen, J.; Craig, I.W.; Milne, B.J.; Poulton, R.; Schalkwyk, L.C.; Taylor, A.; Werts, H.; Moffitt, T.E. Moderation of breastfeeding effects on the IQ by genetic variation in fatty acid metabolism. Proc. Natl. Acad. Sci. USA 2007, 104, 18860–18865. [Google Scholar] [CrossRef] [PubMed]
- Plomin, R.; Deary, I.J. Genetics and intelligence differences: Five special findings. Mol. Psychiatry 2015, 20, 98–108. [Google Scholar] [CrossRef] [PubMed]
- Butcher, L.M.; Davis, O.S.P.; Craig, I.W.; Plomin, R. Genome-wide quantitative trait locus association scan of general cognitive ability using pooled DNA and 500K single nucleotide polymorphism microarrays. Genes Brain Behav. 2008, 7, 435–446. [Google Scholar] [CrossRef] [PubMed]
- Davies, G.; Tenesa, A.; Payton, A.; Yang, J.; Harris, S.E.; Liewald, D.; Ke, X.; Le Hellard, S.; Christoforou, A.; Luciano, M. Genome-wide association studies establish that human intelligence is highly heritable and polygenic. Mol. Psychiatry 2011, 16, 996–1005. [Google Scholar] [CrossRef] [PubMed]
- Briley, D.A.; Tucker-Drob, E.M. Genetic and environmental continuity in personality development: A meta-analysis. Psychol. Bull. 2014, 140, 1303–1331. [Google Scholar] [CrossRef] [PubMed]
- Deary, I.J.; Yang, J.; Davies, G.; Harris, S.E.; Tenesa, A.; Liewald, D.; Luciano, M.; Lopez, L.M.; Gow, A.J.; Corley, J. Genetic contributions to stability and change in intelligence from childhood to old age. Nature 2012, 482, 212–215. [Google Scholar] [CrossRef] [PubMed]
- Trzaskowski, M.; Shakeshaft, N.G.; Plomin, R. Intelligence indexes generalist genes for cognitive abilities. Intelligence 2013, 41, 560–565. [Google Scholar] [CrossRef] [PubMed]
- Crone, E.A.; Elzinga, B.M. Changing brains: How longitudinal functional magnetic resonance imaging studies can inform us about cognitive and social-affective growth trajectories. Wiley Interdiscip. Rev. Cogn. Sci. 2015, 6, 53–63. [Google Scholar] [CrossRef] [PubMed]
- Kingsbury, M.A.; Finlay, B.L. The cortex in multidimensional space: Where do cortical areas come from? Dev. Sci. 2001, 4, 125–142. [Google Scholar] [CrossRef]
- Haworth, C.M.A.; Wright, M.J.; Luciano, M.; Martin, N.G.; De Geus, E.J.C.; Van Beijsterveldt, C.E.M.; Bartels, M.; Posthuma, D.; Boomsma, D.I.; Davis, O.S.P. The heritability of general cognitive ability increases linearly from childhood to young adulthood. Mol. Psychiatry 2010, 15, 1112–1120. [Google Scholar] [CrossRef] [PubMed]
- Bartels, M.; Rietveld, M.J.H.; Van Baal, G.C.M.; Boomsma, D.I. Genetic and environmental influences on the development of intelligence. Behav. Genet. 2002, 32, 237–249. [Google Scholar] [CrossRef] [PubMed]
- Brant, A.M.; Munakata, Y.; Boomsma, D.I.; DeFries, J.C.; Haworth, C.M.A.; Keller, M.C.; Martin, N.G.; McGue, M.; Petrill, S.A.; Plomin, R. The nature and nurture of high IQ: An extended sensitive period for intellectual development. Psychol. Sci. 2013, 24, 1487–1495. [Google Scholar] [CrossRef] [PubMed]
- Trzaskowski, M.; Yang, J.; Visscher, P.M.; Plomin, R. DNA evidence for strong genetic stability and increasing heritability of intelligence from age 7 to 12. Mol. Psychiatry 2014, 19, 380–384. [Google Scholar] [CrossRef] [PubMed]
- Briley, D.A.; Tucker-Drob, E.M. Explaining the increasing heritability of cognitive ability across development: A meta-analysis of longitudinal twin and adoption studies. Psychol. Sci. 2013, 24, 1704–1713. [Google Scholar] [CrossRef] [PubMed]
- Plomin, R. Development, Genetics, and Psychology; Erlbaum: Hillsdale, NJ, USA, 1986. [Google Scholar]
- Thomas, M.S.C. Do more intelligent brains retain heightened plasticity for longer in development? A computational investigation. Dev. Cogn. Neurosci. 2016, 19, 258–269. [Google Scholar] [CrossRef] [PubMed]
- Elman, J.L. Rethinking Innateness: A Connectionist Perspective on Development; MIT Press: Cambridge, MA, USA, 1998; Volume 10. [Google Scholar]
- Sale, A.; Berardi, N.; Maffei, L. Enrich the environment to empower the brain. Trends Neurosci. 2009, 32, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Brooks-Gunn, J.; Duncan, G.J. The effects of poverty on children. Future Child. 1997, 7, 55–71. [Google Scholar] [CrossRef]
- Gottfried, A.W.; Gottfried, A.E.; Bathurst, K.; Guerin, D.W.; Parramore, M.M. Socioeconomic Status in Children's Development and Family Environment: Infancy through Adolescence. In Socioeconomic Status, Parenting and Child Development; Erlbaum: Hillsdale, NJ, USA, 2003. [Google Scholar]
- Farah, M.J.; Shera, D.M.; Savage, J.H.; Betancourt, L.; Giannetta, J.M.; Brodsky, N.L.; Malmud, E.K.; Hurt, H. Childhood poverty: Specific associations with neurocognitive development. Brain Res. 2006, 1110, 166–174. [Google Scholar] [CrossRef] [PubMed]
- Tomalski, P.; Moore, D.G.; Ribeiro, H.; Axelsson, E.L.; Murphy, E.; Karmiloff-Smith, A.; Johnson, M.H.; Kushnerenko, E. Socioeconomic status and functional brain development–associations in early infancy. Dev. Sci. 2013, 16, 676–687. [Google Scholar] [CrossRef] [PubMed]
- Noble, K.G.; Norman, M.F.; Farah, M.J. Neurocognitive correlates of socioeconomic status in kindergarten children. Dev. Sci. 2005, 8, 74–87. [Google Scholar] [CrossRef] [PubMed]
- Rao, H.; Betancourt, L.; Giannetta, J.M.; Brodsky, N.L.; Korczykowski, M.; Avants, B.B.; Gee, J.C.; Wang, J.; Hurt, H.; Detre, J.A. Early parental care is important for hippocampal maturation: Evidence from brain morphology in humans. Neuroimage 2010, 49, 1144–1150. [Google Scholar] [CrossRef] [PubMed]
- Piccolo, L.R.; Merz, E.C.; He, X.; Sowell, E.R.; Noble, K.G. Age-Related Differences in Cortical Thickness Vary by Socioeconomic Status. PLoS ONE 2016, 11, e0162511. [Google Scholar] [CrossRef] [PubMed]
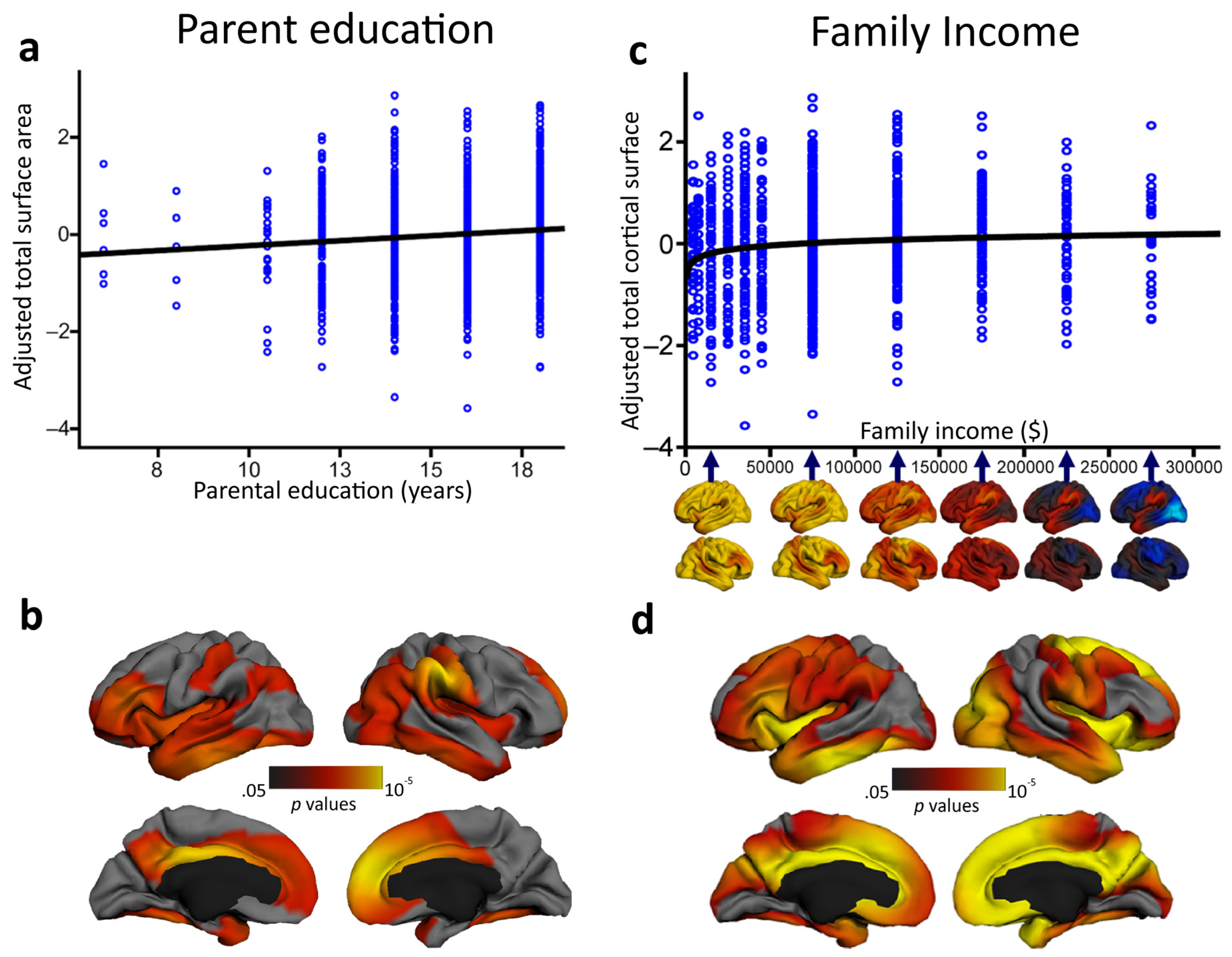
- Noble, K.G.; Houston, S.M.; Brito, N.H.; Bartsch, H.; Kan, E.; Kuperman, J.M.; Akshoomoff, N.; Amaral, D.G.; Bloss, C.S.; Libiger, O. Family income, parental education and brain structure in children and adolescents. Nat. Neurosci. 2015, 18, 773–778. [Google Scholar] [CrossRef] [PubMed]
- Noble, K.G.; Houston, S.M.; Kan, E.; Sowell, E.R. Neural correlates of socioeconomic status in the developing human brain. Dev. Sci. 2012, 15, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Lawson, G.M.; Duda, J.T.; Avants, B.B.; Wu, J.; Farah, M.J. Associations between children’s socioeconomic status and prefrontal cortical thickness. Dev. Sci. 2013, 16, 641–652. [Google Scholar] [CrossRef] [PubMed]
- Ursache, A.; Noble, K.G. Neurocognitive development in socioeconomic context: Multiple mechanisms and implications for measuring socioeconomic status. Psychophysiology 2016, 53, 71–82. [Google Scholar] [CrossRef] [PubMed]
- Hackman, D.A.; Farah, M.J.; Meaney, M.J. Socioeconomic status and the brain: Mechanistic insights from human and animal research. Nat. Rev. Neurosci. 2010, 11, 651–659. [Google Scholar] [CrossRef] [PubMed]
- Evans, G. The environment of childhood poverty. Am. Psychol. 2004, 59, 77–92. [Google Scholar] [CrossRef] [PubMed]
- Karmiloff-Smith, A. Beyond Modularity: A Developmental Perspective on Cognitive Science; MIT Press: Cambridge, MA, USA, 1995. [Google Scholar]
- Johnson, M.H. Functional brain development in humans. Nat. Rev. Neurosci. 2001, 2, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Johnson, M.H. Interactive specialization: A domain-general framework for human functional brain development? Dev. Cogn. Neurosci. 2011, 1, 7–21. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.-J.; Davis, E.P.; Sandman, C.A.; Sporns, O.; O’Donnell, B.F.; Buss, C.; Hetrick, W.P. Children’s intellectual ability is associated with structural network integrity. Neuroimage 2016, 124, 550–556. [Google Scholar] [CrossRef] [PubMed]
- Khundrakpam, B.S.; Lewis, J.D.; Reid, A.; Karama, S.; Zhao, L.; Chouinard-Decorte, F.; Evans, A.C. Brain Development Cooperative Group. Imaging structural covariance in the development of intelligence. Neuroimage 2017, 144, 227–240. [Google Scholar] [PubMed]
- Song, M.; Zhou, Y.; Li, J.; Liu, Y.; Tian, L.; Yu, C.; Jiang, T. Brain spontaneous functional connectivity and intelligence. Neuroimage 2008, 41, 1168–1176. [Google Scholar] [CrossRef] [PubMed]
- Langeslag, S.J.E.; Schmidt, M.; Ghassabian, A.; Jaddoe, V.W.; Hofman, A.; Lugt, A.; Verhulst, F.C.; Tiemeier, H.; White, T.J.H. Functional connectivity between parietal and frontal brain regions and intelligence in young children: The Generation R study. Hum. Brain Mapp. 2013, 34, 3299–3307. [Google Scholar] [CrossRef] [PubMed]
- Fjell, A.M.; Westlye, L.T.; Amlien, I.; Tamnes, C.K.; Grydeland, H.; Engvig, A.; Espeseth, T.; Reinvang, I.; Lundervold, A.J.; Lundervold, A. High-expanding cortical regions in human development and evolution are related to higher intellectual abilities. Cereb. Cortex 2015, 25, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Barsalou, L.W. Grounded cognition. Annu. Rev. Psychol. 2008, 59, 617–645. [Google Scholar] [CrossRef] [PubMed]
- Fagan, J.F.; McGrath, S.K. Infant recognition memory and later intelligence. Intelligence 1981, 5, 121–130. [Google Scholar] [CrossRef]
- Lewis, M.; Brooks-Gunn, J. Visual attention at three months as a predictor of cognitive functioning at two years of age. Intelligence 1981, 5, 131–140. [Google Scholar] [CrossRef]
- McCall, R.B. Early predictors of later IQ: The search continues. Intelligence 1981, 5, 141–147. [Google Scholar] [CrossRef]
- Kavšek, M. Predicting later IQ from infant visual habituation and dishabituation: A meta-analysis. J. Appl. Dev. Psychol. 2004, 25, 369–393. [Google Scholar] [CrossRef]
- Fagan, J.F.; Holland, C.R.; Wheeler, K. The prediction, from infancy, of adult IQ and achievement. Intelligence 2007, 35, 225–231. [Google Scholar] [CrossRef]
- Rose, S.A.; Feldman, J.F.; Jankowski, J.J.; Van Rossem, R. Information processing from infancy to 11years: Continuities and prediction of IQ. Intelligence 2012, 40, 445–457. [Google Scholar] [CrossRef] [PubMed]
- Scerif, G. Attention trajectories, mechanisms and outcomes: At the interface between developing cognition and environment. Dev. Sci. 2010, 13, 805–812. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, M.H. How infant and mother jointly contribute to developing cognitive competence in the child. Proc. Natl. Acad. Sci. USA 1985, 82, 7470–7473. [Google Scholar] [CrossRef] [PubMed]
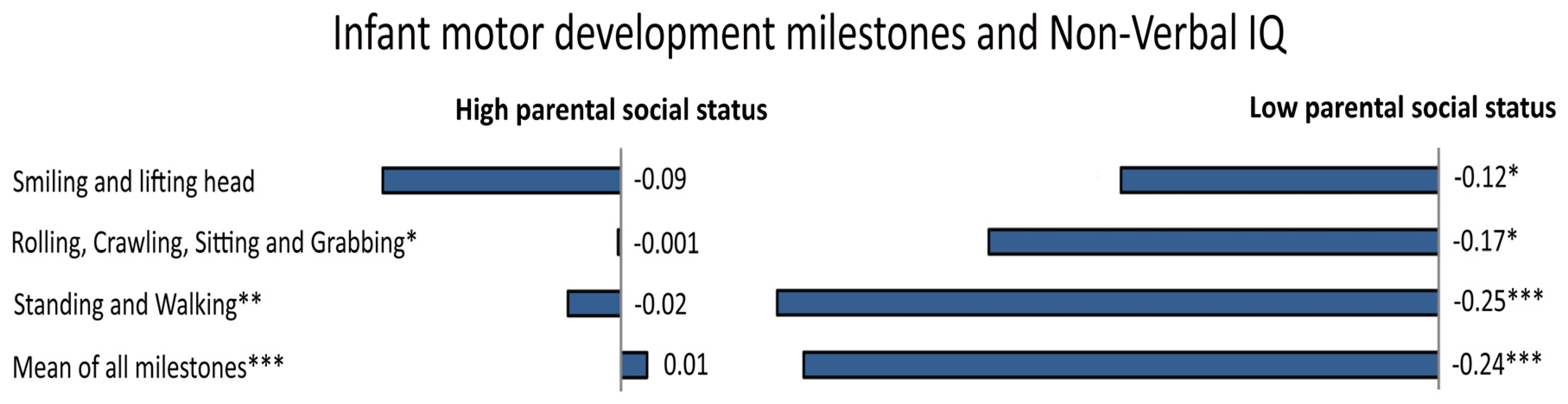
- Murray, G.K.; Jones, P.B.; Kuh, D.; Richards, M. Infant developmental milestones and subsequent cognitive function. Ann. Neurol. 2007, 62, 128–136. [Google Scholar] [CrossRef] [PubMed]
- Hamadani, J.D.; Tofail, F.; Cole, T.; Grantham-McGregor, S. The relation between age of attainment of motor milestones and future cognitive and motor development in Bangladeshi children. Matern. Child Nutr. 2013, 9, 89–104. [Google Scholar] [CrossRef] [PubMed]
- Capute, A.J.; Shapiro, B.K.; Palmer, F.B.; Ross, A.; Wachtel, R.C. Cognitive-Motor Interactions The Relationship of Infant Gross Motor Attainment to IQ at 3 Years. Clin. Pediatr. (Phila). 1985, 24, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Piek, J.P.; Dawson, L.; Smith, L.M.; Gasson, N. The role of early fine and gross motor development on later motor and cognitive ability. Hum. Mov. Sci. 2008, 27, 668–681. [Google Scholar] [CrossRef] [PubMed]
- Taanila, A.; Murray, G.K.; Jokelainen, J.; Isohanni, M.; Rantakallio, P. Infant developmental milestones: A 31-year follow-up. Dev. Med. Child Neurol. 2005, 47, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Ridler, K.; Veijola, J.M.; Tanskanen, P.; Miettunen, J.; Chitnis, X.; Suckling, J.; Murray, G.K.; Haapea, M.; Jones, P.B.; Isohanni, M.K. Fronto-cerebellar systems are associated with infant motor and adult executive functions in healthy adults but not in schizophrenia. Proc. Natl. Acad. Sci. USA 2006, 103, 15651–15656. [Google Scholar] [CrossRef] [PubMed]
- Flensborg-Madsen, T.; Mortensen, E.L. Infant developmental milestones and adult intelligence: A 34-year follow-up. Early Hum. Dev. 2015, 91, 393–400. [Google Scholar] [CrossRef] [PubMed]
- Rigoli, D.; Piek, J.P.; Kane, R.; Oosterlaan, J. Motor coordination, working memory, and academic achievement in a normative adolescent sample: Testing a mediation model. Arch. Clin. Neuropsychol. 2012, 27, 766–780. [Google Scholar] [CrossRef] [PubMed]
- Rigoli, D.; Piek, J.P.; Kane, R.; Oosterlaan, J. An examination of the relationship between motor coordination and executive functions in adolescents. Dev. Med. Child Neurol. 2012, 54, 1025–1031. [Google Scholar] [CrossRef] [PubMed]
- Tabriz, A.A.; Sohrabi, M.R.; Parsay, S.; Abadi, A.; Kiapour, N.; Aliyari, M.; Ahmadi, F.; Roodaki, A. Relation of intelligence quotient and body mass index in preschool children: A community-based cross-sectional study. Nutr. Diabetes 2015, 5, e176. [Google Scholar] [CrossRef] [PubMed]
- Lemos, G.C.; Almeida, L.S.; Colom, R. Intelligence of adolescents is related to their parents’ educational level but not to family income. Pers. Individ. Dif. 2011, 50, 1062–1067. [Google Scholar] [CrossRef]
- Ganzach, Y. Adolescents’ intelligence is related to family income. Pers. Individ. Differ. 2014, 59, 112–115. [Google Scholar] [CrossRef]
- Karmiloff-Smith, A.; Aschersleben, G.; de Schonen, S.; Elsabbagh, M.; Hohenberger, A.; Serres, J. Constraints on the timing of infant cognitive change: Domain-specific or domain-general? Eur. J. Dev. Sci. 2010, 4, 31–45. [Google Scholar]
- De Bellis, M.D.; Hooper, S.R.; Spratt, E.G.; Woolley, D.P. Neuropsychological findings in childhood neglect and their relationships to pediatric PTSD. J. Int. Neuropsychol. Soc. 2009, 15, 868–878. [Google Scholar] [CrossRef] [PubMed]
- Hay, D.F.; Kumar, R. Interpreting the effects of mothers’ postnatal depression on children’s intelligence: A critique and re-analysis. Child Psychiatry Hum. Dev. 1995, 25, 165–181. [Google Scholar] [CrossRef] [PubMed]
- Blair, C.; Diamond, A. Biological processes in prevention and intervention: The promotion of self-regulation as a means of preventing school failure. Dev. Psychopathol. 2008, 20, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Neville, H.J.; Stevens, C.; Pakulak, E.; Bell, T.A.; Fanning, J.; Klein, S.; Isbell, E. Family-based training program improves brain function, cognition, and behavior in lower socioeconomic status preschoolers. Proc. Natl. Acad. Sci. USA 2013, 110, 12138–12143. [Google Scholar] [CrossRef] [PubMed]
- Pietschnig, J.; Penke, L.; Wicherts, J.M.; Zeiler, M.; Voracek, M. Meta-analysis of associations between human brain volume and intelligence differences: How strong are they and what do they mean? Neurosci. Biobehav. Rev. 2015, 57, 411–432. [Google Scholar] [CrossRef] [PubMed]
- Tucker-Drob, E.M.; Briley, D.A. Continuity of genetic and environmental influences on cognition across the life span: A meta-analysis of longitudinal twin and adoption studies. Psychol. Bull. 2014, 140, 949–979. [Google Scholar] [CrossRef] [PubMed]
- Plomin, R.; DeFries, J.C.; Loehlin, J.C. Genotype-environment interaction and correlation in the analysis of human behavior. Psychol. Bull. 1977, 84, 309–322. [Google Scholar] [CrossRef] [PubMed]
- Tucker-Drob, E.M.; Harden, K.P. Intellectual Interest Mediates GenexSocioeconomic Status Interaction on Adolescent Academic Achievement. Child Dev. 2012, 83, 743–757. [Google Scholar] [PubMed]
- Tucker-Drob, E.M.; Briley, D.A.; Engelhardt, L.E.; Mann, F.D.; Harden, K.P. Genetically-Mediated Associations Between Measures of Childhood Character and Academic Achievement. J. Pers. Soc. Psychol. 2016, 111, 790–815. [Google Scholar] [CrossRef] [PubMed]
- Kievit, R.A.; Davis, S.W.; Griffiths, J.; Correia, M.M.; Henson, R.N. A watershed model of individual differences in fluid intelligence. Neuropsychologia 2016, 91, 186–198. [Google Scholar] [CrossRef] [PubMed]
- Strenze, T. Intelligence and socioeconomic success: A meta-analytic review of longitudinal research. Intelligence 2007, 35, 401–426. [Google Scholar] [CrossRef]
- Gottfredson, L.S. Why g matters: The complexity of everyday life. Intelligence 1997, 24, 79–132. [Google Scholar] [CrossRef]
- Wraw, C.; Deary, I.J.; Gale, C.R.; Der, G. Intelligence in youth and health at age 50. Intelligence 2015, 53, 23–32. [Google Scholar] [CrossRef] [PubMed]
- McGurn, B.; Deary, I.J.; Starr, J.M. Childhood cognitive ability and risk of late-onset Alzheimer and vascular dementia. Neurology 2008, 71, 1051–1056. [Google Scholar] [CrossRef] [PubMed]
- Koenen, K.C.; Moffitt, T.E.; Roberts, A.L.; Martin, L.T.; Kubzansky, L.; Harrington, H.; Poulton, R.; Caspi, A. Childhood IQ and adult mental disorders: A test of the cognitive reserve hypothesis. Am. J. Psychiatry 2009, 166, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Lubinski, D.; Humphreys, L.G. Incorporating general intelligence into epidemiology and the social sciences. Intelligence 1997, 24, 159–201. [Google Scholar] [CrossRef]
- Ceci, S.J. How much does schooling influence general intelligence and its cognitive components? A reassessment of the evidence. Dev. Psychol. 1991, 27, 703–722. [Google Scholar] [CrossRef]
- Cahan, S.; Cohen, N. Age versus schooling effects on intelligence development. Child Dev. 1989, 60, 1239–1249. [Google Scholar] [CrossRef] [PubMed]
- Bedard, K.; Dhuey, E. The persistence of early childhood maturity: International evidence of long-run age effects. Q. J. Econ. 2006, 121, 1437–1472. [Google Scholar] [CrossRef]
- Jones, C.; Tarpley, P.L.; Blancero, D. Handbook of Intelligence; Springer: New York, NY, USA, 2015; pp. 459–468. [Google Scholar]
- Karmiloff-Smith, A. Transforming a partially structured brain into a creative mind. Behav. Brain Sci. 1994, 17, 732–745. [Google Scholar] [CrossRef]
| 1 | Piaget was the first psychologist who theorized intelligence within a developmental framework. With admirable foresight, he postulated that intelligence would emerge from a dynamic interplay between the individual and the environment. Ironically, after decades of Nature–Nurture debate, we are still conceptualizing intelligence in similar terms. However, thanks to the recent emergence of the developmental cognitive neuroscience, the neuroepigenetics and the behavioral genetics fields, we can now more accurately define the strict interplay between the developing organism and the environment, which are viewed as active entities exerting cascading effects on intellectual functioning. |

© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rinaldi, L.; Karmiloff-Smith, A. Intelligence as a Developing Function: A Neuroconstructivist Approach. J. Intell. 2017, 5, 18. https://doi.org/10.3390/jintelligence5020018
Rinaldi L, Karmiloff-Smith A. Intelligence as a Developing Function: A Neuroconstructivist Approach. Journal of Intelligence. 2017; 5(2):18. https://doi.org/10.3390/jintelligence5020018
Chicago/Turabian StyleRinaldi, Luca, and Annette Karmiloff-Smith. 2017. "Intelligence as a Developing Function: A Neuroconstructivist Approach" Journal of Intelligence 5, no. 2: 18. https://doi.org/10.3390/jintelligence5020018
APA StyleRinaldi, L., & Karmiloff-Smith, A. (2017). Intelligence as a Developing Function: A Neuroconstructivist Approach. Journal of Intelligence, 5(2), 18. https://doi.org/10.3390/jintelligence5020018