A Paradigm Shift in Biology?

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
- Neglecting the relevance of introns seems growingly difficult to maintain. To give an example, the small nucleolar RNAs (snoRNAs) are encoded in the introns of other genes and serve to RNA maturation. This shows the complexity of control mechanisms in the cell. This complexity can be seen also in RNA or protein decay, as also textbooks acknowledge today [6].
- The difficulty to explain with traditional methods certain findings: For instance, the yeast protein and the Drosophila’s engrailed protein have almost the same structure and functionality but share only 17 amino acid residues over 60 [7]. It is to consider that the shared amino acid residues do not constitute the active domains of the proteins.
- Recently, it has been proposed to search the onset of cancer in a global disfunction of the epigenetic network and especially in cell-cell signaling [8].
- The extensively acknowledged relevance of top-down processes in neurosciences. This is remarkably evident in the endogenous production of dopamine in the brain of rats exploring their maze [9].
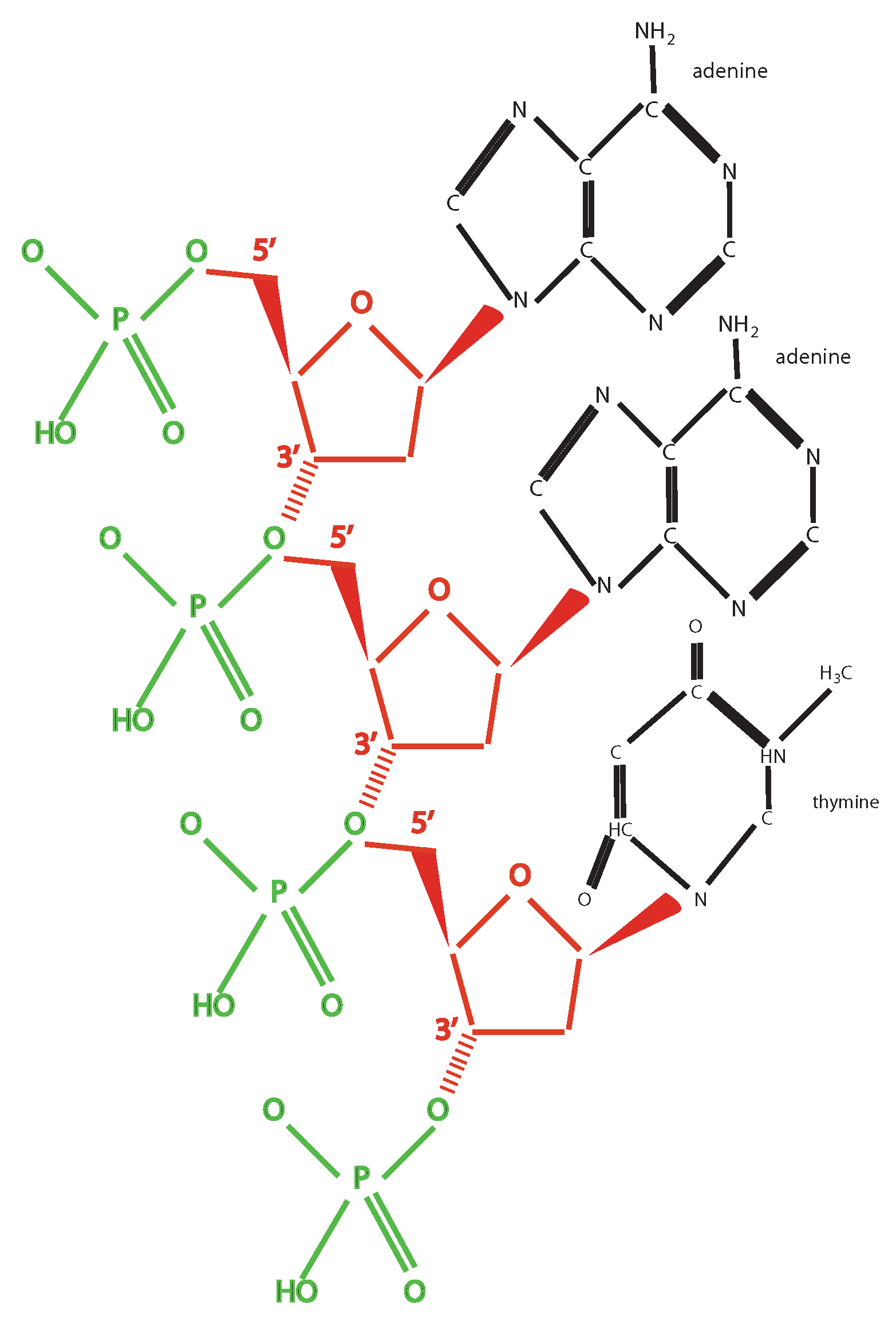
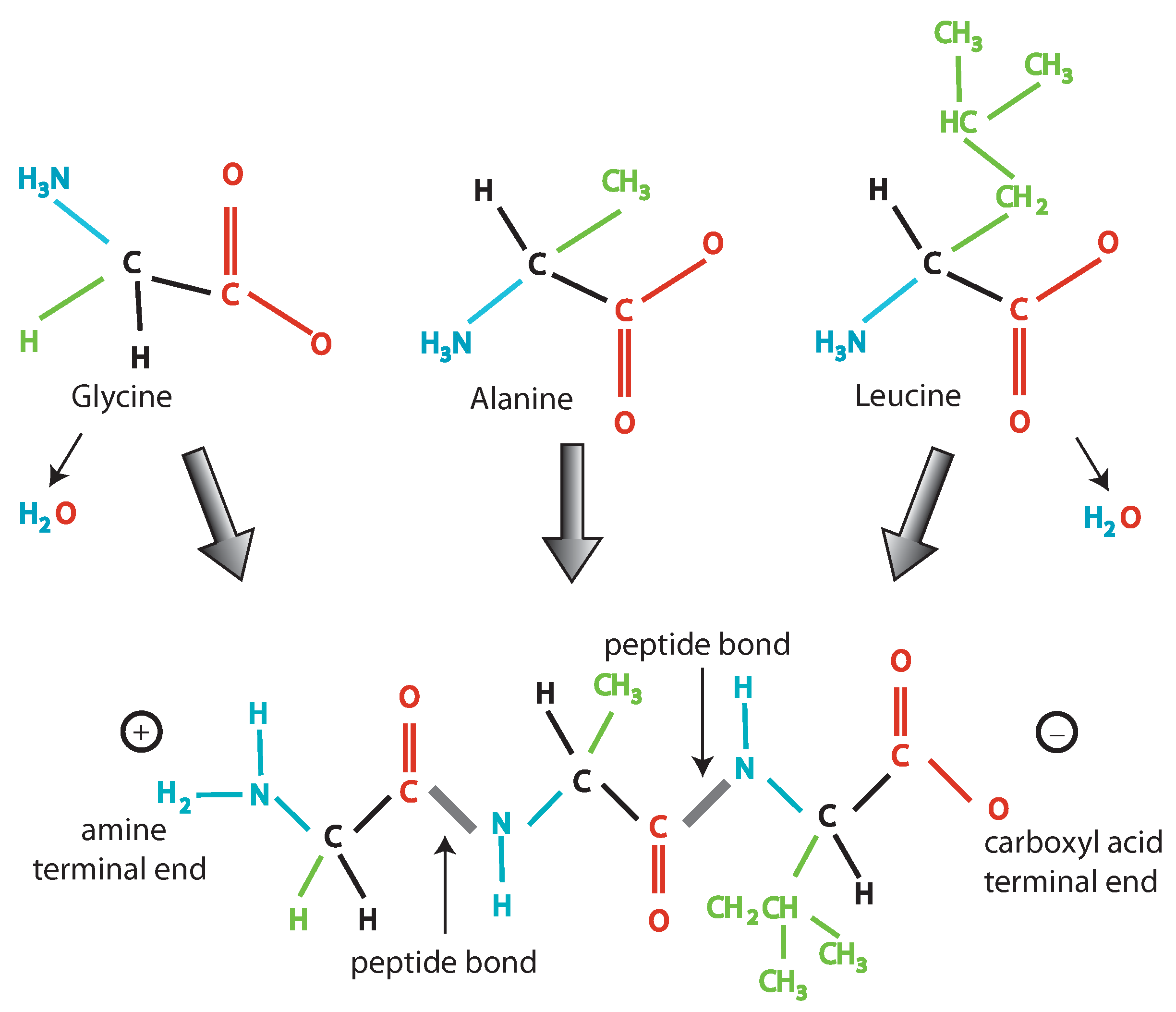
2. Information and Chemistry
- (1)
- The existence of a finite set of mutually exclusive elementary units (A, T, C, G) that
- (2)
- Can be linearly combined (giving rise to a sequence).
- (3)
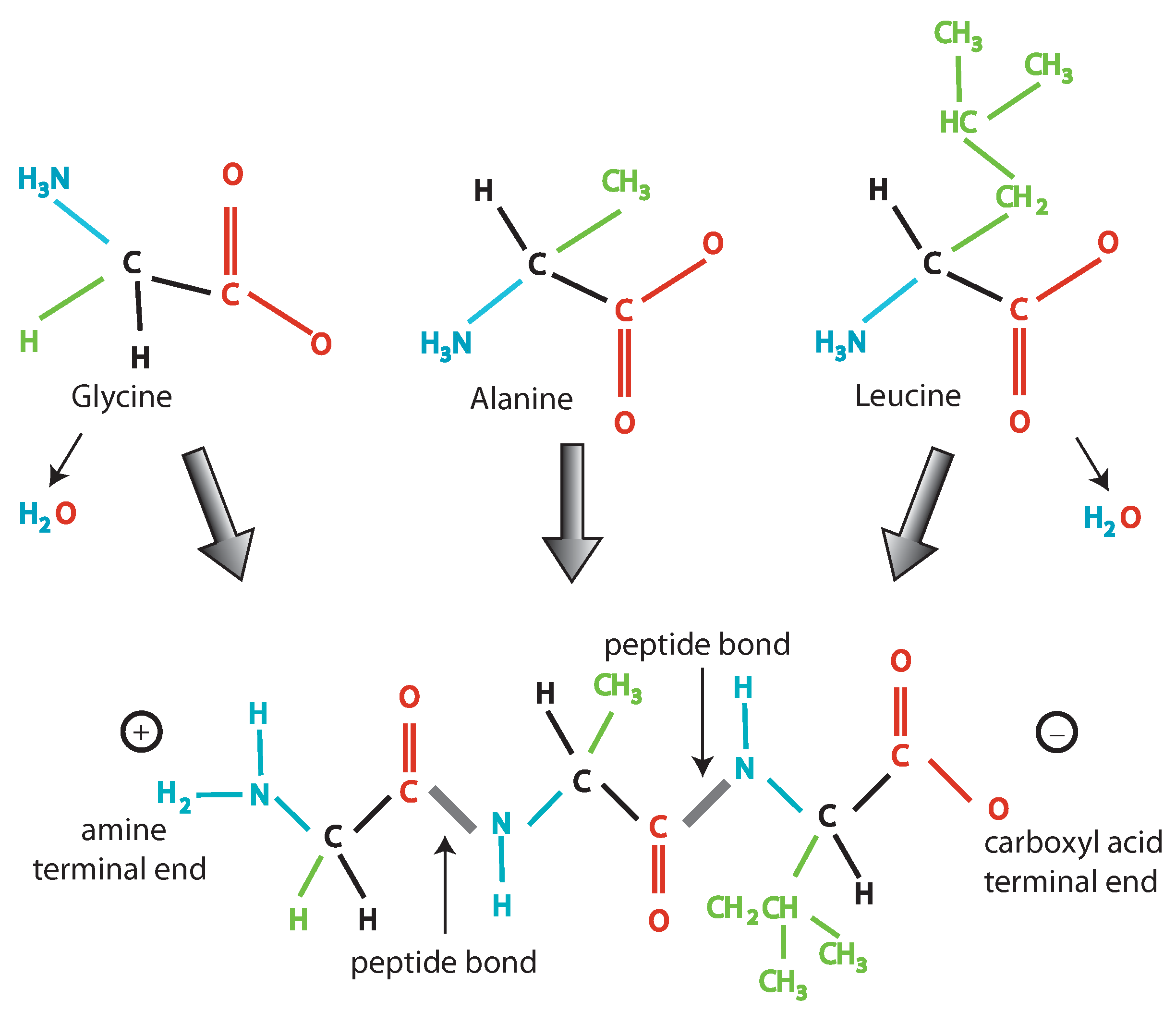
- Every combination must give rise to some “words” (codons) following certain rules: The codons are triplets, there are start and stop signals, AG and CT are subcodes allowing creation of templates.
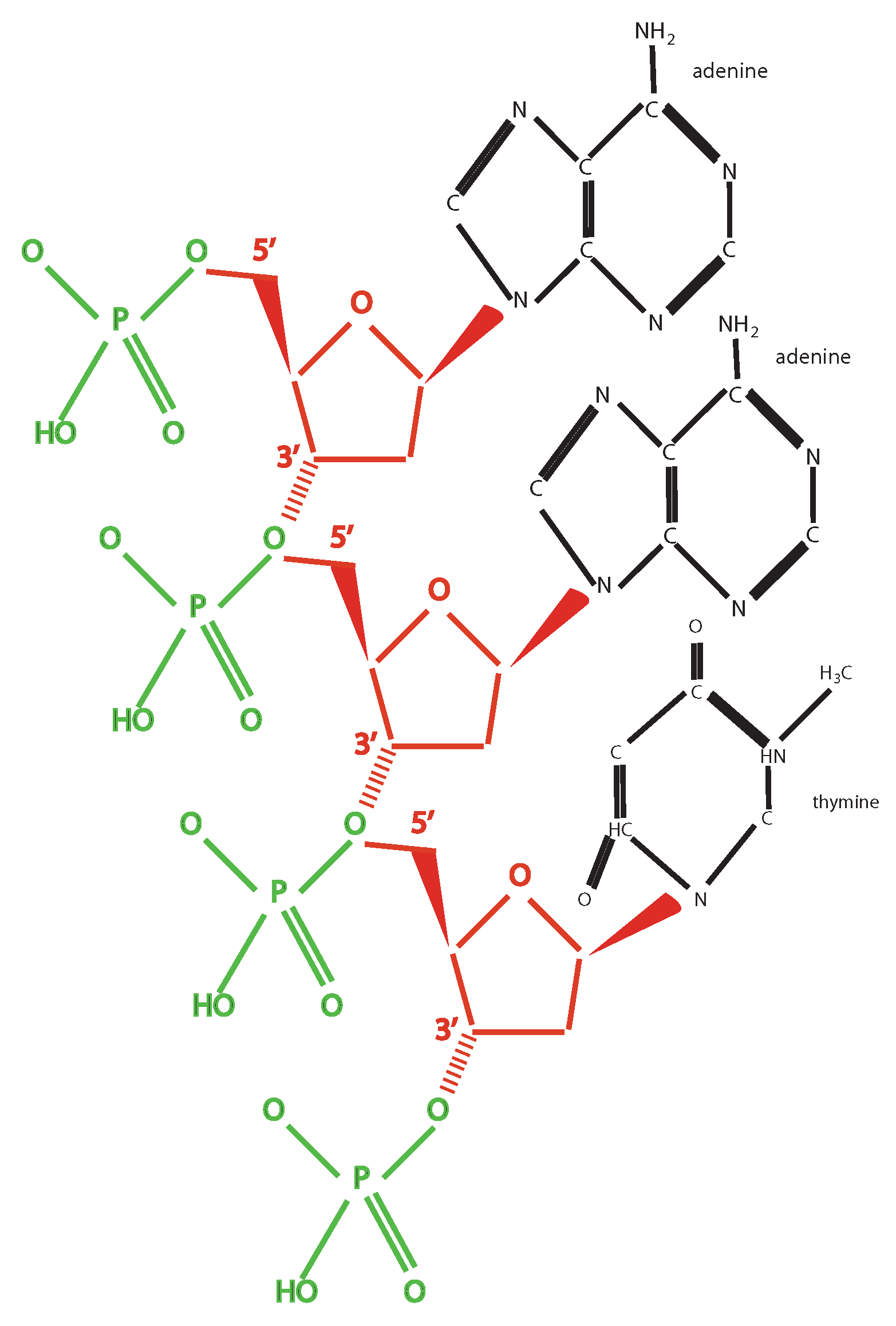
- (4)
- There exist also other elementary units and relative translation rules: The two mentioned subcodes and the RNA code (U instead of T). This suggests a former phase of life with a larger variety of codes, from which, through fine-tuning due to selective pressure, the actual code has been born. This is also shown by the possibility to artificially produce new base pairs: The so-called bases S and Y have been added to the four traditional ones, by producing a sort of six bases code allowing the production of different amino acids [13].
- (a)
- Hydrophobicity,
- (b)
- Energy dependence with respect to volume, and
- (c)
- Correlation between these first two correlations.
3. Complexity
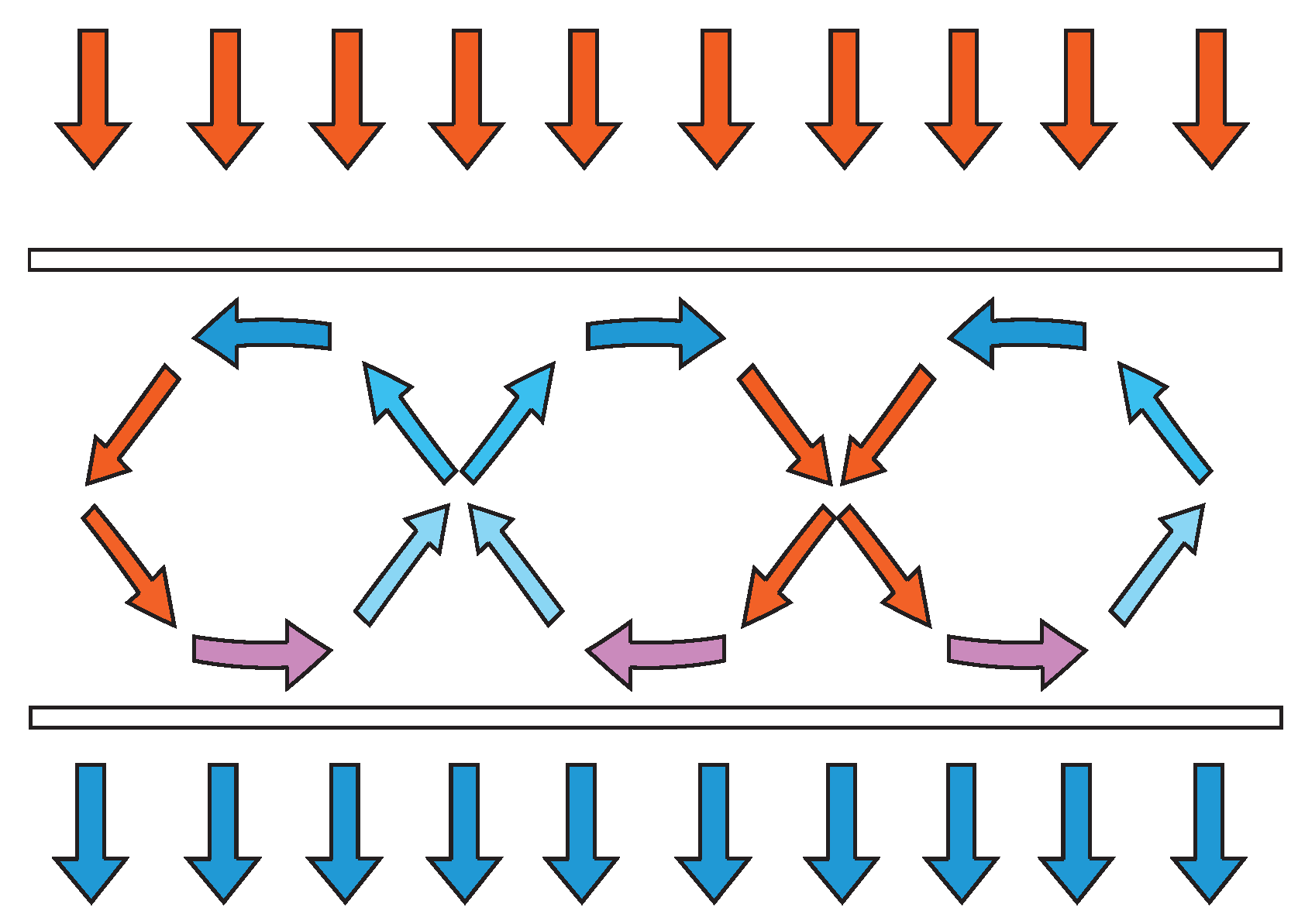
- (1)
- The differentiation of the system in relative independent parts (in some cases, information encapsulation between different levels, like the different levels of DNA packaging from the double strand to the chromosome, and modularity at the same level, like the different segments of DNA exons separated by introns) and
- (2)
- A global order (coherence of the whole).
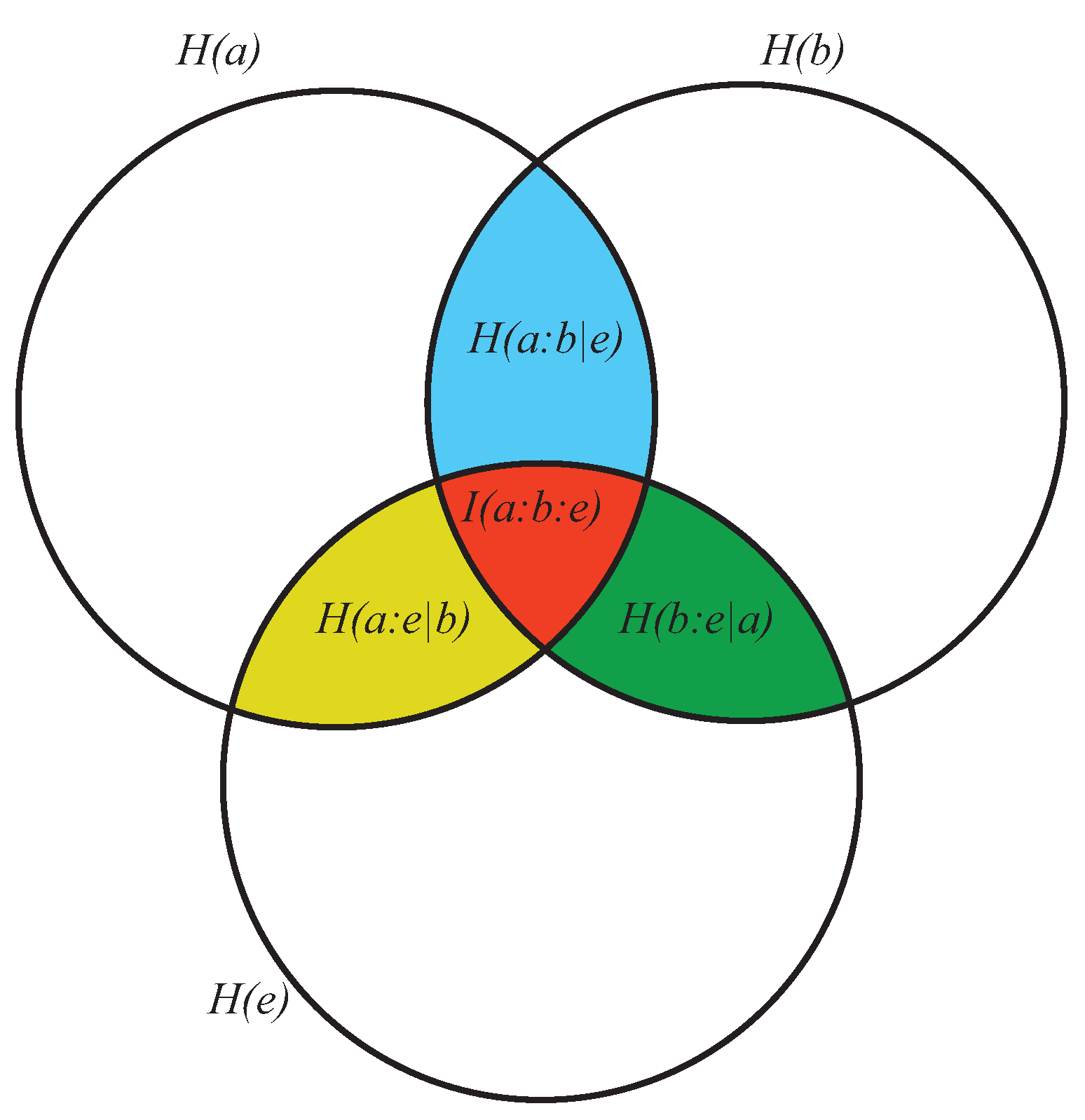
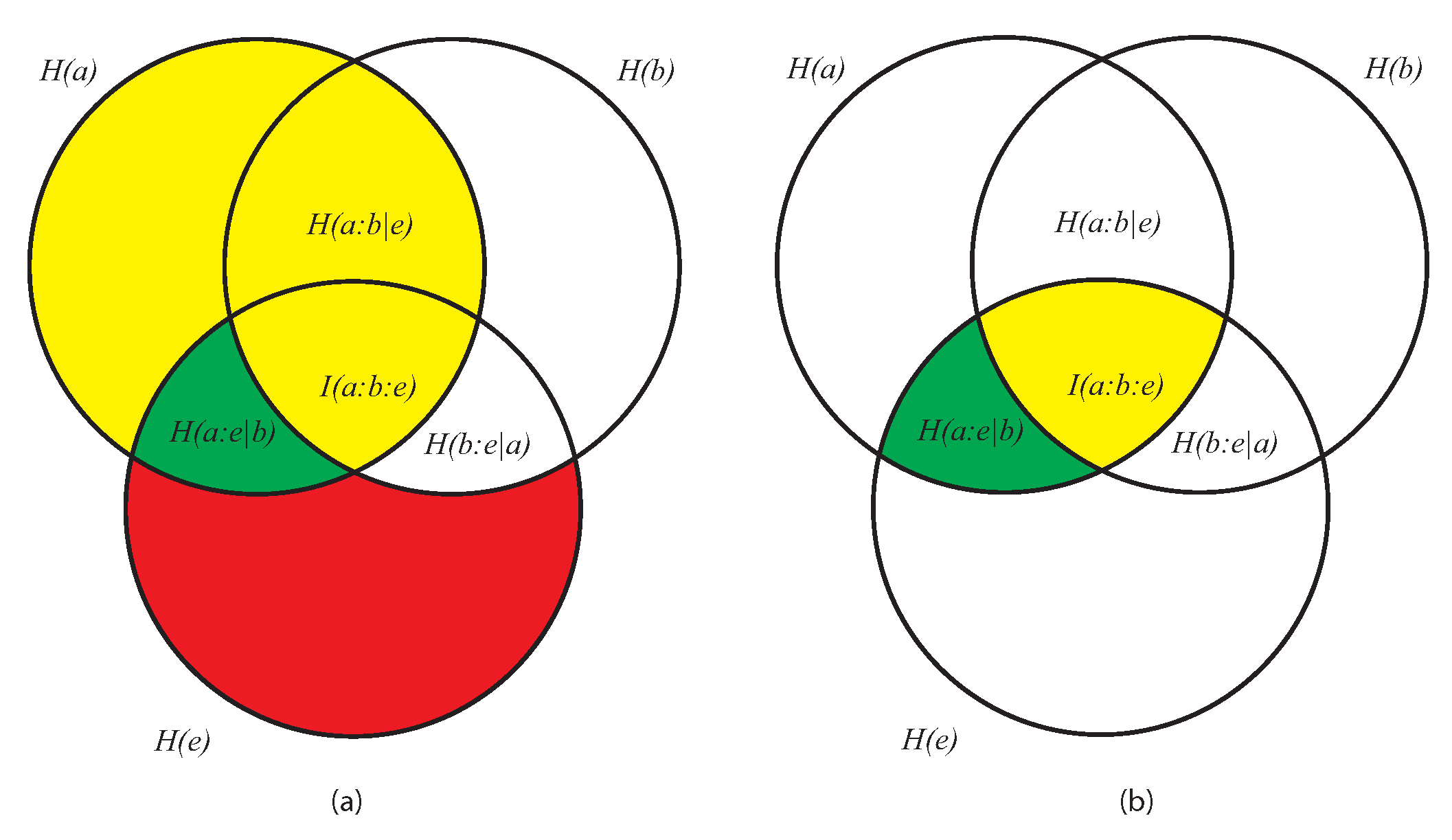
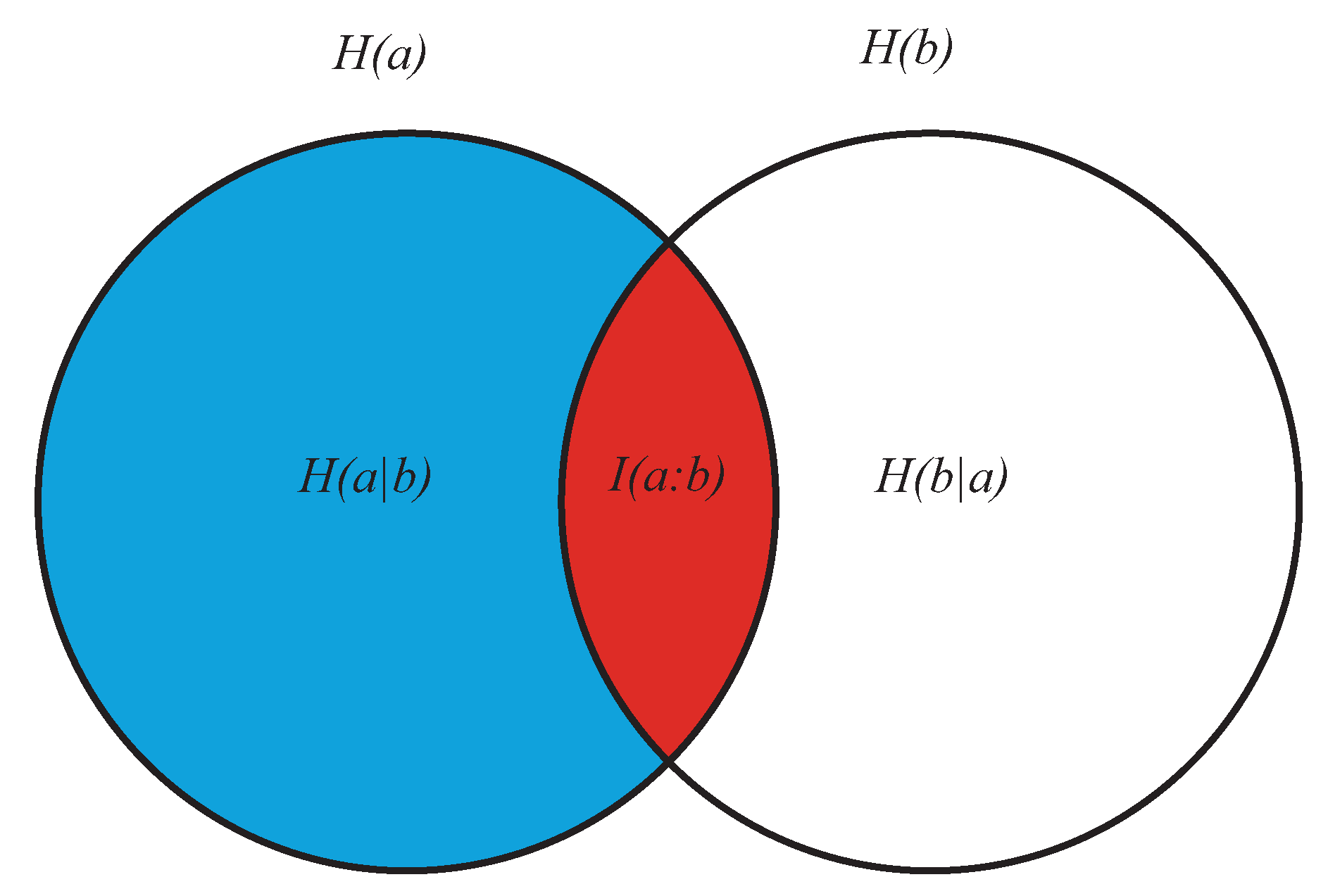
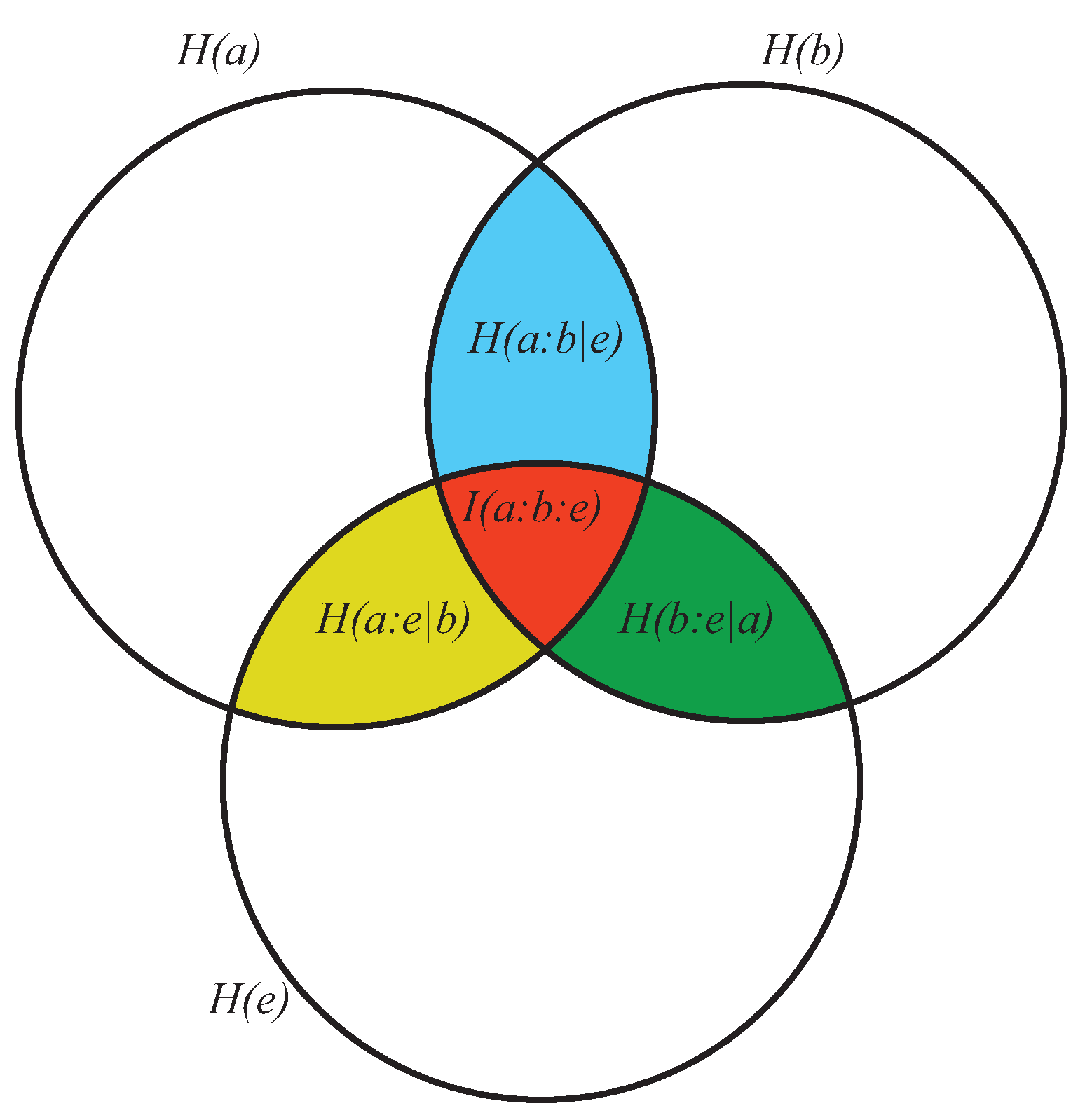
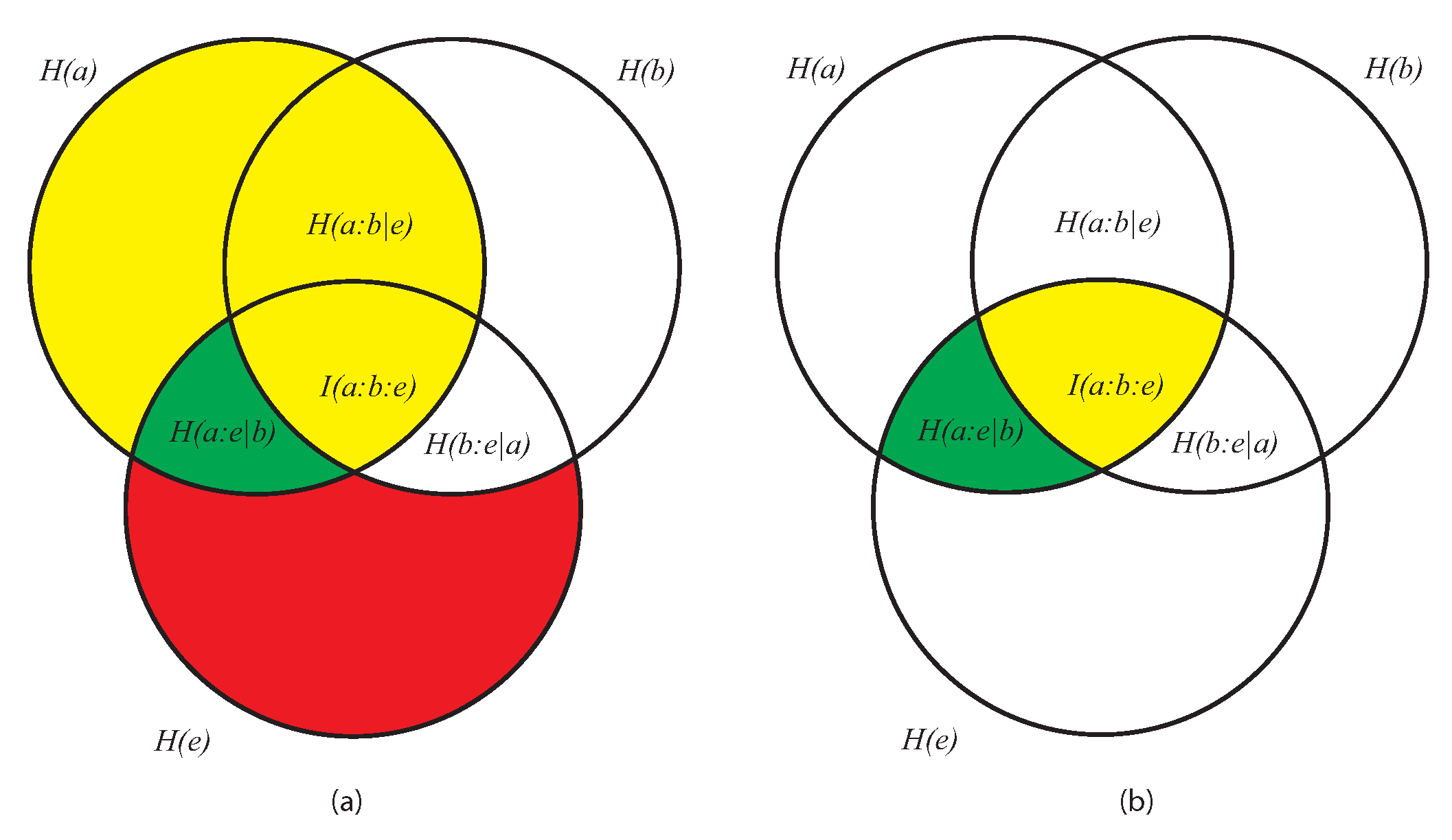
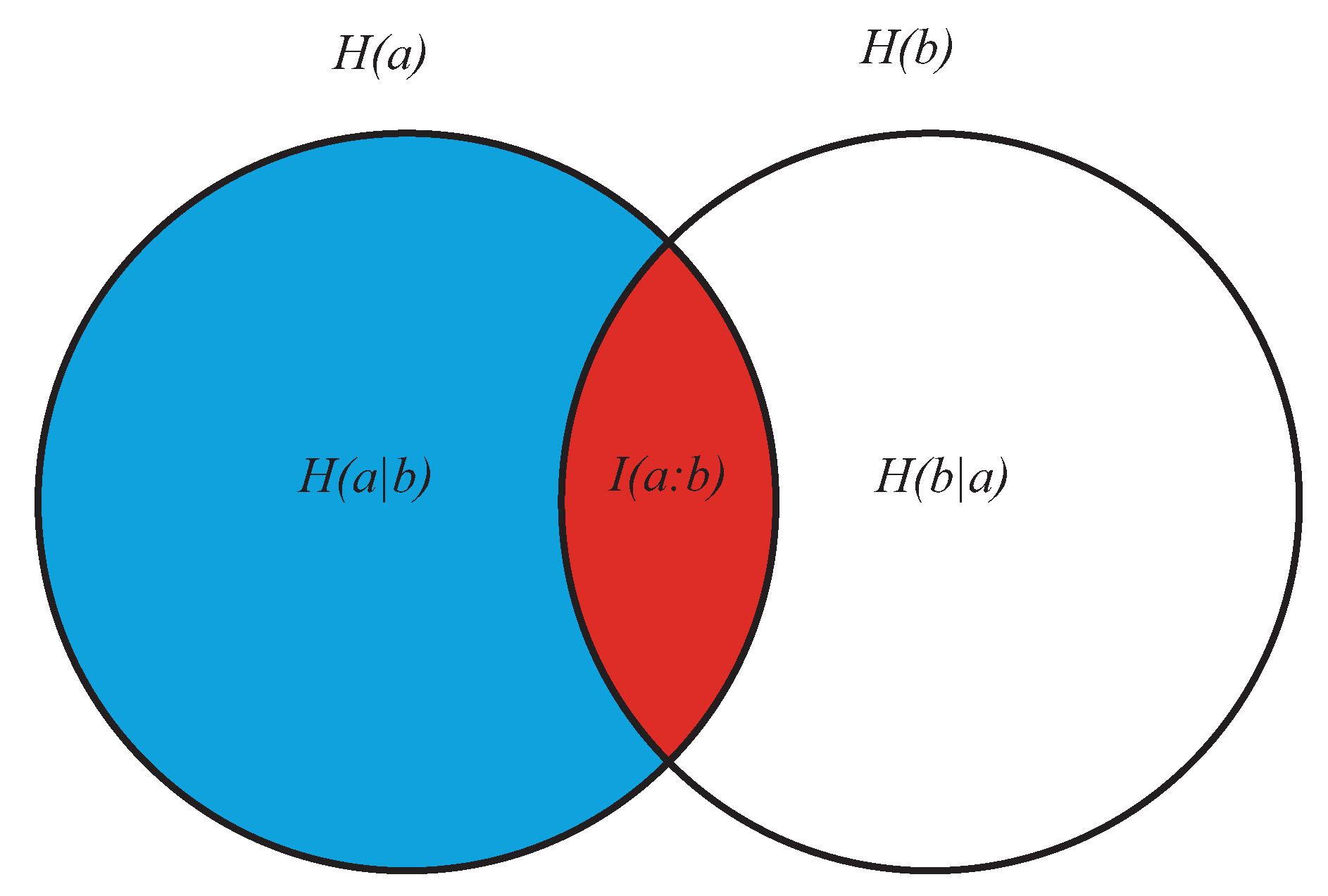
- As the conditional entropy between the information shared by a and e, i.e., , on the one hand, and a third system b, on the other, that is, as , or also
- As the information shared by a, on the one hand, and the conditional entropy between e and b, on the other (), that is, as .
- (1)
- It displays both coherence (order: expressed by the dependencies of the subsystems a, b, and e) and autonomy of the parts (expressed by the lack of the mutual information ). This means that growing of complexity is not proportional to growing of global order, as already anticipated.
- (2)
- At a global level, it does not directly contain informational terms but only conditional-entropic ones. This means that complexity shows global patterns and functions that are independent of any explicit information coding, even if stemming from information codification.
- (3)
- At a local level (the pairwise relations between the three subsystems) it does not require everywhere, but allows for possible information exchange, sharing, and codification.
4. Teleonomy
4.1. Mechanical and Teleonomic Explanations
- Future and past states cannot be deduced from the current state in which the system is.
- There is irreversibility, a feature that is strictly connected with the complexity of biological systems.
4.2. Explanatory Power
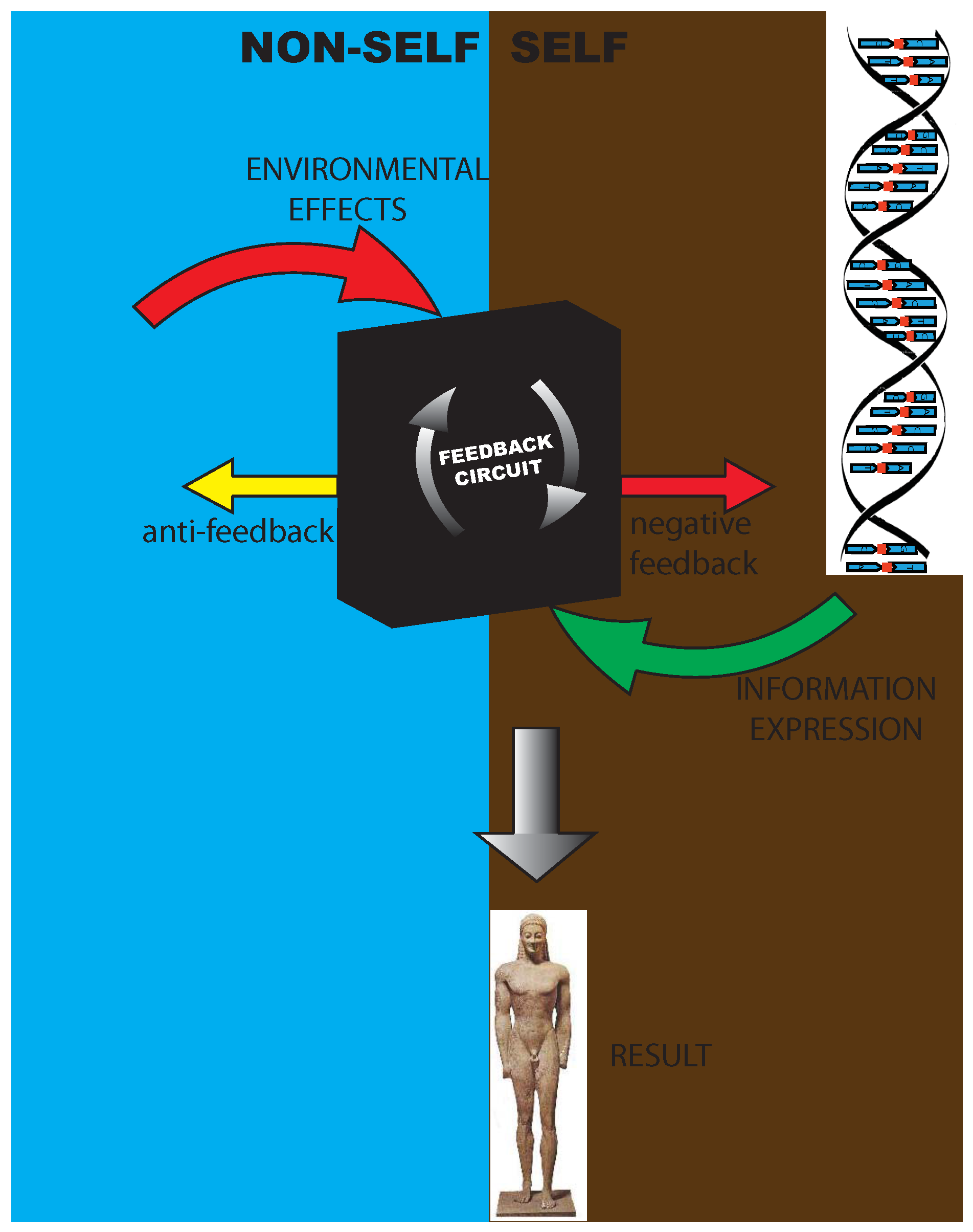
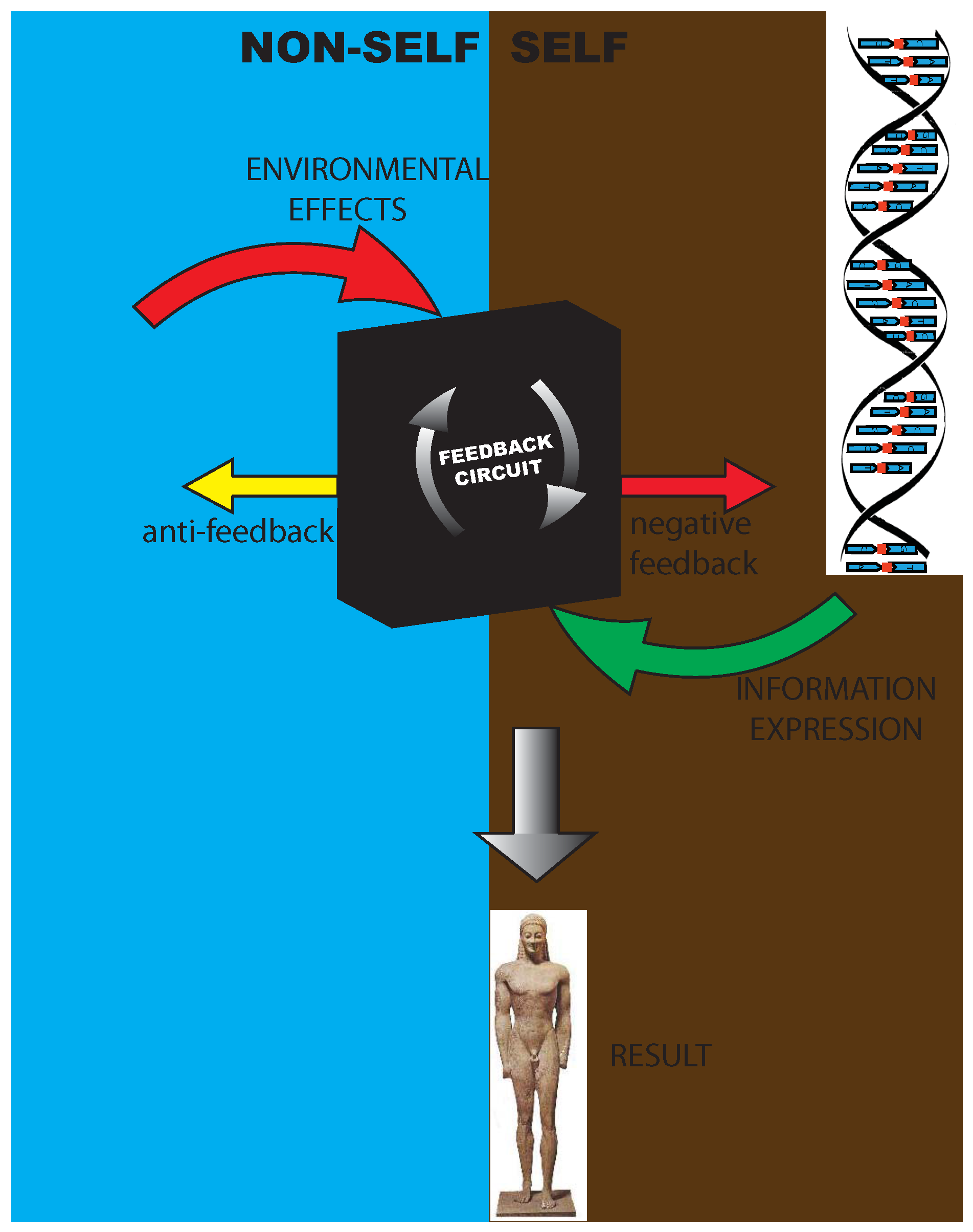
- (1)
- A self-increasing feedback mechanism, coming from the inside of the organism, directed to the building or growth of structures in phenotype or of the three-dimensional configuration of a protein,
- (2)
- A negative feedback [34], whose ultimate source is in some environmental perturbation, mainly addressed at blocking the expression of the activated DNA or at having other damping effects on the phenotype, and
- (3)
- An anti-feedback directed toward the environment, that is, a process spontaneously activated by the organism to prevent dangerous environmental effects and to restore its own homeostasis. This was well understood by Bichat [35], who spoke of an internal principle of organisms able to counteract the spontaneous tendency to destruction.
5. A Selection System
5.1. The Membrane
5.2. Information
- (1)
- A signal triggering the whole process (the so called first messenger) that in most cases binds a receptor (gate protein) without entering itself into the cell;
- (2)
- As a consequence, a second messenger (an intracellular molecule) is delivered inside the cell and may produce amplification of the initial signal (a sort of cascade); this can be assumed to be the proper codification step;
- (3)
- Finally, there is an activation of a response going into the inside of the cell to the metabolic system.
6. Teleology
6.1. Conditions of Teleology
- It cannot be reduced to pure structure, even if it depends on some specific structural elements that are crucial for the function and on the architecture of the whole. Hubs in networks are a manifestation of this property [57,58], something that very much contributes to the robustness of both networks and functions.
- It is always instantiated in some operation: However, it does not depend on it but rather on some formal constraints from above (ultimately, from the needs of the whole organism) and the goal of this upper level. With operation I understand any spatially and temporally coordinated pattern of physical-chemical interactions (it is a complex structure) able to fulfill a function. Operations are therefore a bridge between chemistry and functionality and display both efficacious work and variety.
- It is connected with the needs of the whole organism and therefore relies on the cooperation of different subsystems, like the genetic, metabolic, and selecting systems.
- It depends on information codification but is not its immediate result. The work of natural selection is to have selected the whole process leading from DNA to protein and not single segments.
- Symmetry: If the item a is equivalent to the item b, also b is equivalent to a,
- Reflexivity: a is equivalent to a, and
- Transitivity: If a is equivalent to b and b to c, also a is equivalent to c.
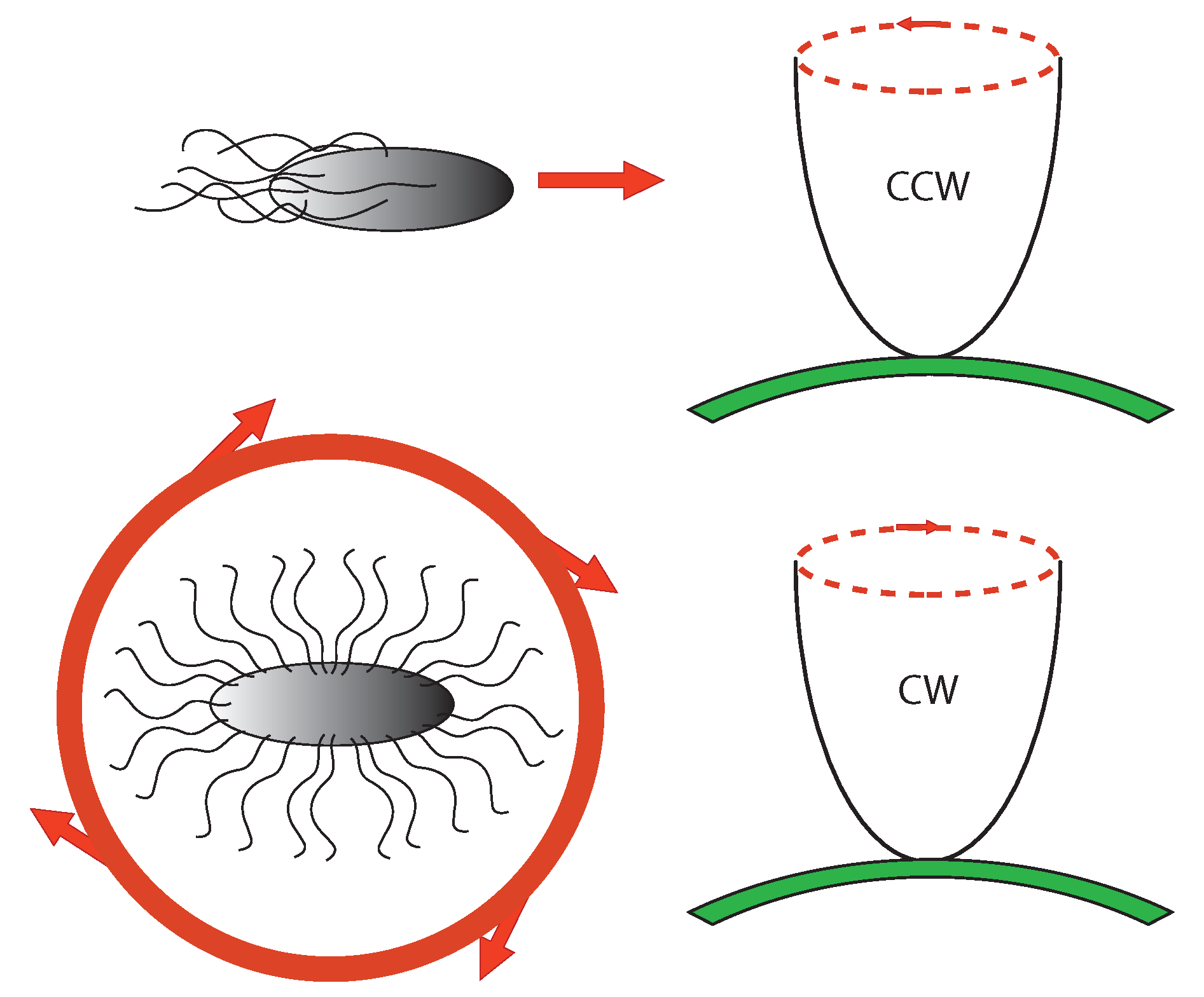
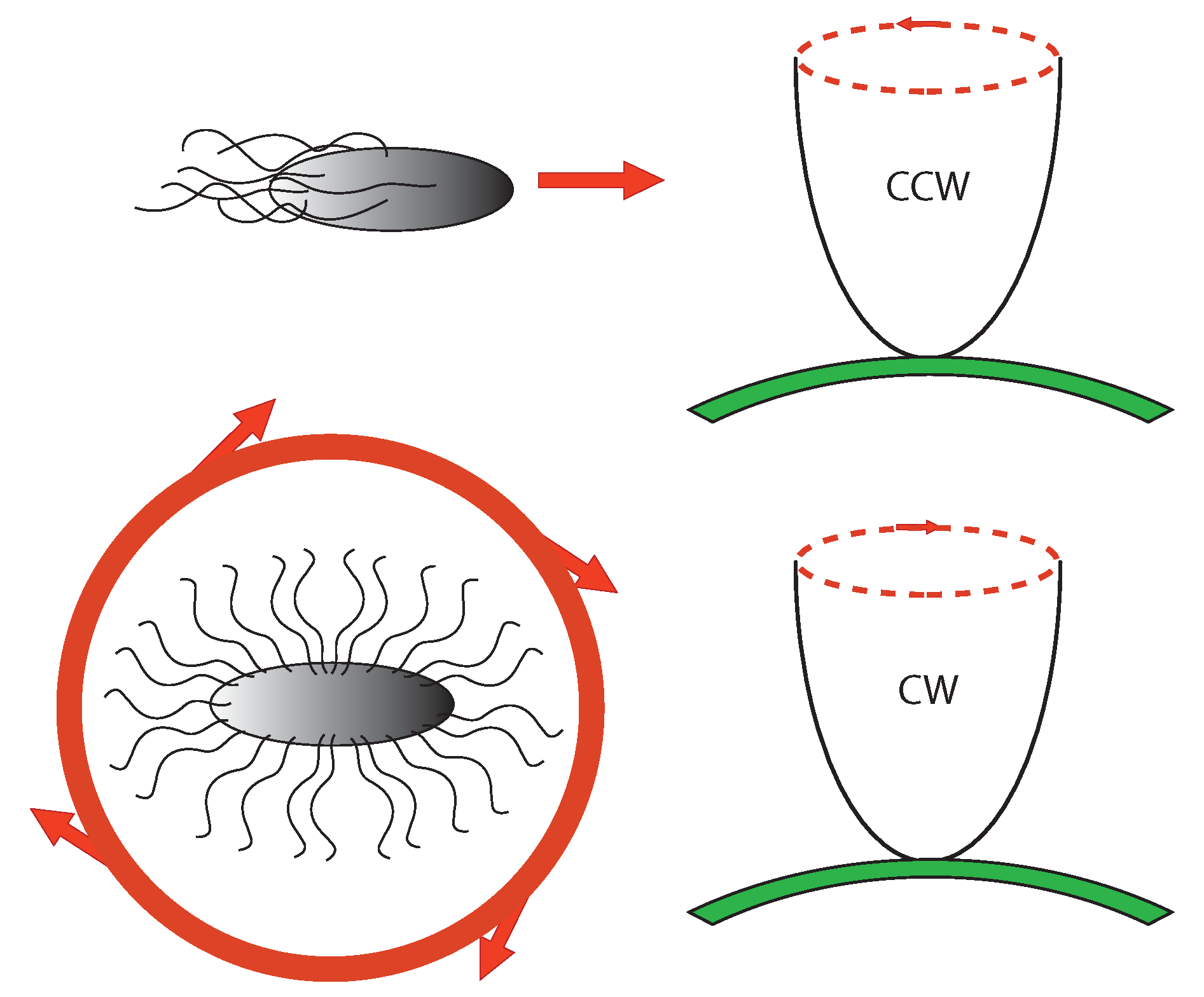
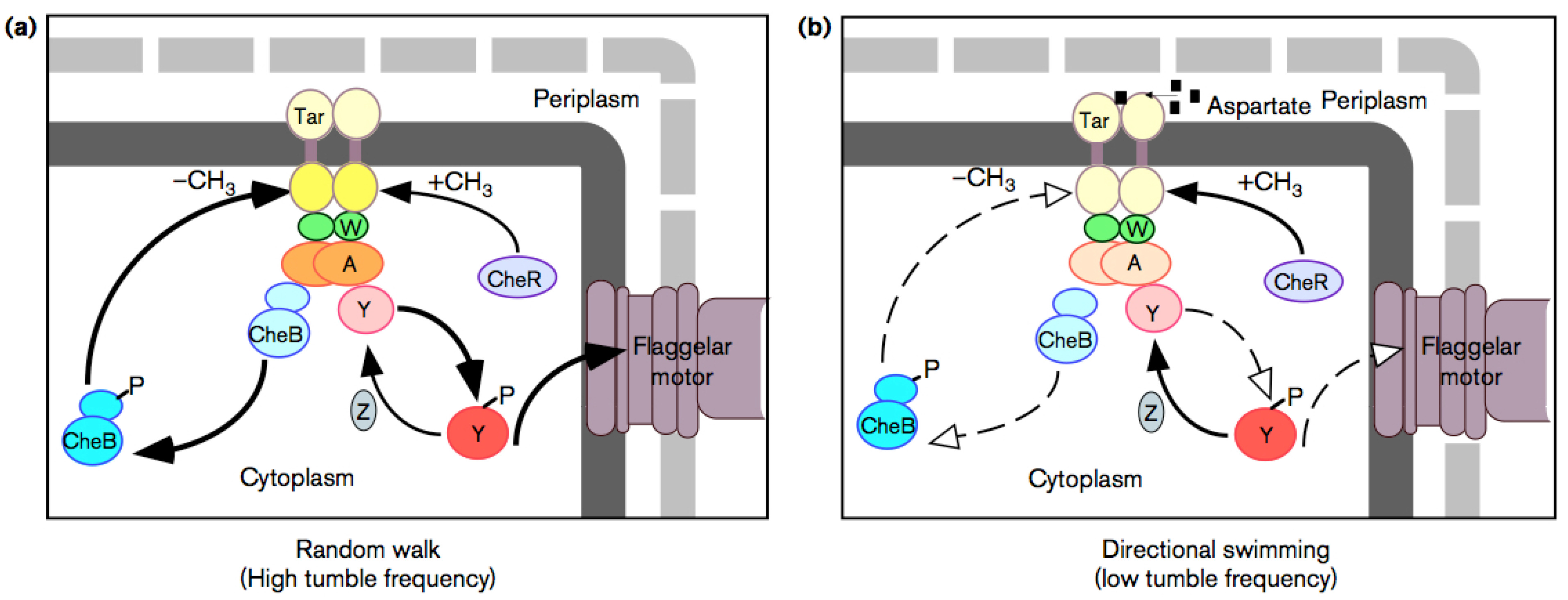
6.2. Chemotaxis as Example of Teleology
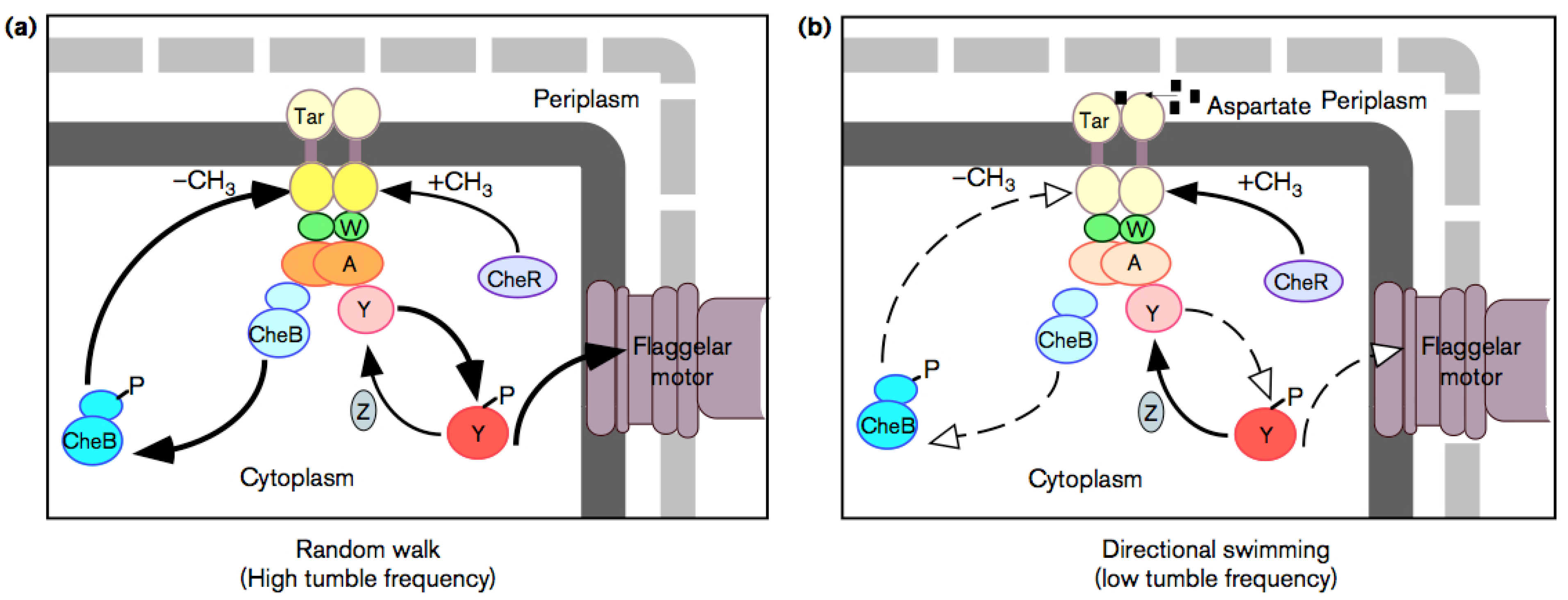
- To make temporal comparison of chemoeffector levels (a pure informational operation), the bacterial cell requires a sensory adaptation mechanism that cancels chemoreceptors signal outputs in static environment, no matter what chemoeffectors may be present [66] (whether attractive or repulsive). This enables the bacterium to reset the threshold sensitivity of the signaling system in order to detect any new change in the chemical environment. It is a true information-erasing mechanism necessary for information acquiring, a kind of very elementary representational function.
- Another very important element is reaction timing: Because Brownian motion of the fluid medium can randomly reorient the bacterium, this requires very short response latencies. It is here that genetic (instructional) factors play a role by enhancing and damping protein production. It has been indeed observed that the protein CheZ plays a very important role in enhancing the rate of CheY (which is directly involved in the change of motion) dephosphorylation [67]. Moreover, in the whole regulation also the transcription factor NarL plays a role.
6.3. Epigenetic Processes
- For the reasons indicated previously, epigeny can indeed be understood as a compromise between cell proliferation (the continuity of the self-reproduction and the own self-production) and the necessity to control cell division (discontinuity, consisting in negative feedback against growth), especially of some specific cells. This has obviously an evolutionary significance [68]. Let me add in this context that early metazoans blocked proliferation by differentiating in a ball of ciliated cells, a primitive blastula [69]. The only cells that could still differentiate were those already in or migrating into the inner cavity of the ball, a sort of gastrulation. In this way, cilia also acquire a developmental role: They help in maintaining the animal shape.
- Epigeny can be understood also as a compromise between environmental inputs (the discontinuous aspect here) and control genes providing anti–feedback (the continuous aspect, from the point of view of the organism). Environmental factors like temperature, nutrition, pressure and gravity, light, presence of predators, presence or absence of conspecifics are very important for the development of an organism. The action of the environment on the organism can be on transcriptional regulation, on the neuroendocrine system, or be a direct cellular induction: They correspond to an action on the genetic, metabolic, and selection system, respectively. The presence of these factors explains why epigeny is not a full teleologic causal process but also consists of considerable teleonomic processes. However, this does not mean that the environment somehow guides or instructs the organism. The environment only provides negative feedback. It is thanks to those teleonomic processes that the organism is able to canalize and even make positive those environmental stimuli for its own growth.
- Continuity and discontinuity are also important from a further perspective. It is perhaps convenient to distinguish between initial information (the continuity across the generation) and information conditioned by the specific metabolism of an individual (the discontinuity). An evidence for this is the fact that differences in position and surroundings in presence of a faithful genetic duplication that result in differences in phenotypic patterns are regulatory differences that are responsible for divergences in developmental processes.
- Cellular memory: Initially, memory almost completely consists of genetic information; as epigeny goes on, epigenetic memory progressively grows. It is, therefore, a parallel process and, to a certain extent, also a cyclic (wave-like) one.
- Cellular machinery, which in turn also depends on cellular memory. Every cell starts its own version of life anew, since its configuration depends on (both temporally and spatially) local context much more than on the genetic information it brings (this represents the point–like, discontinuous aspect).
- (1)
- The set of initial instructions [Section 2] allows for the building of the first elements that give rise to a cascade process (positive feedback) through which further genes and signals are activated and propagated.
- (2)
- The cellular multiplication process proceeds by a successive and parallel but hierarchical building of different levels of commands and containment (body plan, organs, tissues, single cells), where negative feedback and anti-feedback effects are at work. The process is governed by a principle of information accessibility allowing for different levels of information encapsulation, as shown in Section 3.
- (3)
- During this process, many events happen that have multiple effects establishing new interconnections and therefore a huge network of shared information, both horizontally and vertically [Section 4].
- (4)
- The organism is characterized by the fact that it actively searches for the environmental cues (temperature, light, foods, and so on) that allow its own development [Section 5], and it is here that (through anti–feedback) information control and teleologic causation come into play (as explained in this section) assisting epigeny in the process (cognition assists epigenetic processes).
- (5)
- The whole can be seen as a process tending to a final stable state through a trajectory where the distance from the final species-specific steady state is minimized through the active concourse of the organism. However, since each stable state is provisional, we have an itinerant dynamics.
6.4. A Generalization
This principle means, in particular, that the organism maintains its independence from the environment precisely through changing. Indeed, the perturbation, being out of the system’s control (otherwise, it would not happen at all), cannot ever be completely washed out. This means that the system will never fully recover the previous equilibrium state. Therefore, it is a general principle of dynamicity. To this extent, life is deeply rooted in thermodynamics (even if the bridge between entropy growing and entropy lowering processes is constituted through an informational step). Indeed, the Le Chatelier–Braun principle [? ] states that any perturbation of a factor contributing to equilibrium induces a compensating change in an opposing factor. The specificity of organisms is, however, in the fact that such a dynamic integration is or can be adaptive, that is, a change for the better. The concept of itinerancy, which is very widely used in the theory of complex and chaotic systems can be very useful here helping us to overcome the traditional fixed–point attractors in optimal control theory.In appropriate conditions, any system is pushed to minimize the distance between the selected option and the less expensive one or to lower the level of perturbation of its constitutive web of interdependencies provoked by the selection.
7. Conclusions
Acknowledgements
References and Notes
- Lander, E.S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome. Nature 2001, 409, 860–921. [Google Scholar] [CrossRef] [PubMed]
- Wilkins, A.S. The Evolution of Developmental Pathways; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Changeux, J.-P. The Physiology of Truth: Neuroscience and Human Knowledge; Harvard University Press: Cambridge, MA, USA, 2002. [Google Scholar]
- Wolpert, L.; Beddington, R.; Jessell, T.; Lawrence, P.; Meyerowitz, E.; Smith, J. Principles of Development; Oxford University Press: New York, NY, USA, 2005. [Google Scholar]
- Gilbert, S.F. Devolopmental Biology, 8th ed.; Sinauer Associates Inc.: Sunderland, MA, USA, 2006. [Google Scholar]
- Alberts, B.; Johnson, A.; Lewis, J.; Raff, M.; Roberts, K.; Walter, P. The Molecular Biology of the Cell, 5th ed.; Garland Science: New York, NY, USA, 2008. [Google Scholar]
- Wolberger, C.; Vershon, A.K.; Liu, B.; Johnson, A.D.; Pabo, C.O. Crystal structure of a MATa2 Homeodomain–Operator complex suggests a general model for Homeodomain–DNA interactions. Cell 1991, 67, 517–528. [Google Scholar] [CrossRef]
- Bizzarri, M.; Cucina, A.; Conti, F.; D’Anselmi, F. Beyond the oncogene paradigm: Understanding complexity in cancerogenesis. Acta Biotheoretica 2008, 56, 173–196. [Google Scholar] [CrossRef] [PubMed]
- Kentros, C.G.; Agnihotri, N.T.; Streater, S.; Hawkins, R.D.; Kandel, E.R. Increased attention to spatial context increases both place field stability and spatial memory. Neuron 2004, 42, 283–295. [Google Scholar] [CrossRef]
- Kuhn, T.S. The Structure of Scientific Revolutions; University Of Chicago Press: Chicago, IL, USA, 1962. [Google Scholar]
- Pattee, H.H. Evolving Self–reference: Matter, Symbols, and Semantic Closure. Commun. Cognition—Artif. Intell. 1995, 12, 9–28. [Google Scholar]
- Pattee, H.H. The physics of symbols and the evolution of semiotic controls. In The Physics of Symbols and The Evolution of Semiotic Controls; Addison-Wesley: Redwood City, CA, USA, 1997. [Google Scholar]
- Hirao, I.; Ohtsuki, T.; Fujiwara, T.; Mitsui, T.; Yokogawa, T.; Okuni, T.; Nakayama, H.; Takio, K.; Yabuki, T.; Kigawa, T.; Kodama, K.; Yokogawa, T.; Nishikawa, K.; Yokoyama, S. An unnatural base pair for incorporating amino acids analogs to proteins. Nat. Biotechnol. 2002, 20, 177–82. [Google Scholar] [CrossRef] [PubMed]
- Griffiths, P.E. Genetic information: A metaphor in search of a theory. Phil. Sci. 2001, 68, 394–412. [Google Scholar] [CrossRef]
- Polanyi, M. Lifes irreducible structure. Science 1968, 160, 1308–312. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, J. Physico–chemical constraints connected with the coding properties of the genetic system. J. Theor. Biol. 2002, 202, 129–44. [Google Scholar] [CrossRef] [PubMed]
- Holland, S.K.; Blake, C.C.F. Proteins, exons, and molecular evolution. Biosystems 1987, 20, 181–206. [Google Scholar] [CrossRef]
- Gibbs, W.W. The unseen genome: Gems among the junk. Sci. Am. 2003, 11, 48–53. [Google Scholar] [CrossRef]
- Shannon, C.E. A mathematical theory of communication. Bell Syst. Tech. J. 1949, 27, 379–423, 623–56. [Google Scholar] [CrossRef]
- Nielsen, M.A.; Chuang, I.L. Quantum Computation and Quantum Information; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Auletta, G. Quantum information as a general paradigm. Found. Phys. 2005, 35, 787–815. [Google Scholar] [CrossRef]
- Kauffman, S.A. The Origins of Order; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Kauffman, S. At Home in the Universe: The Search for the Laws of Self–Organization and Complexity; Oxford University Press: Oxford, UK, 1995. [Google Scholar]
- Ott, E. Chaos in Dynamical Systems; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Monod, J. Le hasard et la nécessité; Seuil: Paris, France, 1970. [Google Scholar]
- Barbieri, M. The Organic Codes: An Introduction to Semantic Biology; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Arthur, W. Mechanisms of Morphological Evolution; John Wiley: New York, NY, USA, 1984. [Google Scholar]
- Brooks, C.L. III; Gruebele, M.; Onuchic, J.N.; Wolynes, P.G. Chemical physics of protein folding. PNAS 1998, 95, 11037–38. [Google Scholar] [CrossRef] [PubMed]
- Russell, E.S. The Interpretation of Development and Heredity; Clarendon Press: Oxford, UK, 1930. [Google Scholar]
- Robert, J.S. Embryology, Epigenesis, and Evolution: Taking Development Seriously; Cambridge University Press: Cambridge, UK, 2004. [Google Scholar]
- Lotka, A.J. Elements of Physical Biology; Williams and Wilkins: Baltimore, MA, USA, 1925; rep. under the title Elements of Mathematical Biology; Dover: New York, NY, USA, 1956. [Google Scholar]
- Parisi, G. Complex systems: A physicist’s viewpoint. Physica 1999, A263, 557–64. [Google Scholar] [CrossRef]
- Woodger, J.H. Biological Principles: A Critical Study? Kegan Paul: London, UK, 1929; Routledge and Kegan Paul: London, UK, 1967. [Google Scholar]
- Rosenblueth, A.; Wiener, N.; Bigelow, J.H. Behavior, purpose, and teleology. Philos. Sci. 1943, 10, 18–24. [Google Scholar] [CrossRef]
- Bichat, F.X. Recherches physiologiques sur la vie et la mort; Brosson: Paris, France, 1800. [Google Scholar]
- Gilbert, S.F.; Epel, D. Ecological developmental biology: Integrating epigenetics, medicine, and evolution; Sinauer: Sunderland, MA, USA, 2009. [Google Scholar]
- Gould, S.J.; Vrba, E.S. Exaptation: A missing term in the science of form. Paleobiology 1982, 8, 4–15. [Google Scholar]
- Blobel, G. Intracellular protein topogenesis. PNAS 1980, 77, 1496–500. [Google Scholar] [CrossRef] [PubMed]
- Monod, J.; Changeux, J.–P.; Jacob, F. Allosteric proteins and cellular control systems. J. Mol. Biol. 1963, 6, 306–329. [Google Scholar] [CrossRef]
- Monod, J.; Wyman, J.; Changeux, J.–P. On the nature of allosteric transitions: A plausible model. J. Mol. Biol. 1965, 12, 88–118. [Google Scholar] [CrossRef]
- Hunter, Tony signaling—2000 and beyond. Cell 2000, 100, 113–27.
- Ma, L.; Cantley, L.C.; Janmey, P.A.; Kirschner, M.W. Corequirement of specific phosphoinositides and small GTP–Binding protein Cdc42 in inducing actin assembly in Xenopus egg extracts. J. Cell Biol. 1998, 140, 1125–136. [Google Scholar] [PubMed]
- Martin, T.F. Phosphoinositide lipids as signaling molecules: Common themes for signal transduction, cytoskeletal regulation, and membrane trafficking. Annu. Rev. Cell Dev. Bi. 1998, 14, 231–64. [Google Scholar] [CrossRef] [PubMed]
- Dippold, H.C.; Ng, M.M.; Farber–Katz, S.E.; Lee, S.–K.; Kerr, M.L.; Peterman, M.C.; Sim, R.; Wiharto, P.A.; Galbraith, K.A.; Madhavarapu, S.; Fuchs, G.J.; Meerloo, T.; Farquhar, M.G.; Zhou, H.; Field, S.J. GOLPH3 bridges phosphatidylinositol–4– phosphate and actomyosin to stretch and shape the Golgi to promote budding. Cell 2009, 139, 337–51. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J. A theory of Cortical Responses. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2005, B360, 815–836. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.; Kiebel, S. Predictive coding under the free–Energy principle. Phil. Trans. R. Soc. B 2009, B364, 1211–21. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Stephan, K.E. Free–Energy and the brain. Synthese 2007, 159, 417–58. [Google Scholar] [CrossRef] [PubMed]
- Friston, K.J.; Kilner, J.; Harrison, L. A free energy principle for the brain. J. Physiol. Paris. 2006, 100, 70–87. [Google Scholar] [CrossRef] [PubMed]
- McKay, D.J.C. A free–Energy minimization algorithm for decoding and cryptanalysis. Electron. Lett. 1995, 31, 445–47. [Google Scholar]
- Friston, K.J.; Daunizeau, J.; Kilner, J.; Kiebel, S.J. Action and behavior: A free–energy formulation. Biol. Cybern. 2010, 102, 227–260. [Google Scholar] [CrossRef] [PubMed]
- Ayala, F.J. Teleological explanations in evolutionary biology. Phil. Scie. 1970, 37, 1–15. [Google Scholar] [CrossRef]
- Ayala, F.J. Teleological explanations versus teleology. Hist. Phil. Life Sci. 1998, 20, 41–50. [Google Scholar]
- Oparin, A.I. The Origin of Life on the Earth; Academic Press: New York, NY, USA, 1957. [Google Scholar]
- van Gelder, T. Compositionality: A connectionist variation on a classical theme. Cognitive Sci. 1990, 14, 355–84. [Google Scholar] [CrossRef]
- Berg, J.M.; Tymoczko, J.L.; Stryer, L. Biochemistry, 6th ed.; Freeman and Co.: New York, NY, USA, 2006. [Google Scholar]
- Crick, F.H.C. The central dogma of molecular biology. Nature 1970, 227, 561–63. [Google Scholar] [CrossRef] [PubMed]
- Jeong, H.; Tombor, B.; Albert, R.; Oltvai, Z.N.; Barabási, A.–L. The large–scale organization of metabolic networks. Nature 2000, 407, 651–654. [Google Scholar] [PubMed]
- Jeong, H.; Mason, S.P.; Barabási, A.–L.; Oltvai, Z.N. Lethality and centrality in protein networks. Nature 2001, 411, 41–42. [Google Scholar] [CrossRef] [PubMed]
- Wagner, A. Robustness and Evolvability in Living Systems; Princeton University Press: Princeton, NJ, USA, 2005. [Google Scholar]
- Auletta, G.; Ellis, G.; Jaeger, L. Top-down causation by information control: From a philosophical problem to a scientific research program. J. R. Soc. Interface 2008, 5, 1159–1172. [Google Scholar] [CrossRef] [PubMed]
- Wegscheid, B.; Condon, C.; Hartmann, R.K. Type A and B RNase P RNAs are interchangeable in vivo despite substantial biophysical differences. EMBO Reports 2006, 7, 411–17. [Google Scholar] [CrossRef] [PubMed]
- Gobert, A.; Gutmann, B.; Taschner, A.; Gössringer, M.; Holzmann, J.; Hartmann, R.K.; Rossmanith, W.; Giegé, P. A single Arabidopsis organellar protein has RNase P activity. Nat. Struct. Mol. Biol. 2010, 17, 740–46. [Google Scholar] [CrossRef] [PubMed]
- Weiner, O.D. Regulation of cell polarity during eukaryotic chemotaxis: The chemotactic compass. Curr. Opin. Cell Biol. 2002, 14, 196–202. [Google Scholar] [CrossRef]
- Alon, U. Simplicity in biology. Nature 2007, 446, 497. [Google Scholar] [CrossRef] [PubMed]
- Jurica, M.S.; Stoddard, B.L. Mind your Bs and Rs: Bacterial chemotaxis, signal transduction and protein recognition. Structure 1998, 6, 809–813. [Google Scholar] [CrossRef]
- Parkinson, J.S. Signal transduction schemes of bacteria. Cell 1993, 73, 857–71. [Google Scholar] [CrossRef]
- Rao, C.V.; Kirby, J.R.; Arkin, A.P. Design and diversity in bacterial chemotaxis: A comparative study in Escherichia coli and Bacillus subtilis. PLOS Biol. 2004, 2, 0239–52. [Google Scholar] [CrossRef] [PubMed]
- Minelli, A. The Development of Animal Form: Ontogeny, Morphology, and Evolution; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Buss, L.W. The Evolution of Individuality; Princeton University Press: Princeton, NJ, USA, 1987. [Google Scholar]
- Arthur, W. The Origin of Animal Body Plans: A Study in Evolutionary Developmental Biology; Cambridge University Press: Cambridge, UK, 1997. [Google Scholar]
- Schlichting, C.D.; Pigliucci, M. Phenotypic Evolution: A Reaction Norm Perspective; Sinauer: Sunderland, MA, USA, 1998. [Google Scholar]
- Wright, S. Evolution in mendelian populations. Genetics 1931, 16, 97–159. [Google Scholar] [PubMed]
- Wright, S. The roles of mutation, interbreeding, crossbreeding and selection in evolution. In Proceedings of the Sixth International Congress on Genetics, Ithaca, New York, NY, USA; 1932; pp. 56–66. [Google Scholar]
© 2010 by the author; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/.)
Share and Cite
Auletta, G. A Paradigm Shift in Biology? Information 2010, 1, 28-59. https://doi.org/10.3390/info1010028
Auletta G. A Paradigm Shift in Biology? Information. 2010; 1(1):28-59. https://doi.org/10.3390/info1010028
Chicago/Turabian StyleAuletta, Gennaro. 2010. "A Paradigm Shift in Biology?" Information 1, no. 1: 28-59. https://doi.org/10.3390/info1010028
APA StyleAuletta, G. (2010). A Paradigm Shift in Biology? Information, 1(1), 28-59. https://doi.org/10.3390/info1010028