
Comparing Contemporary Evangelical Models Regarding Human Origins

Abstract
1. Introduction
2. Results
Formed from dust. The most obvious statement about Adam—and the one most important to this discussion—is the statement that God formed (yaṣar) him from the dust (‘apar) of the earth. Is this intended to be a statement about the material origins of the first human being? Traditionally, it has been common to think about this statement as describing a material process of special creation characterized by discontinuity with any previously existing creature.
The Christian view of creation that Darwinism rivaled was not monolithic, even if the popular belief among Christians was that God created the cosmos approximately 4000 years before the coming of the Christ. This belief preceded the publication of The Annals of the World by James Ussher, Archbishop of Armagh, who attempted to identify the time of creation’s beginning with precision. For example, both leading church Reformers Martin Luther and John Calvin believed that creation was not yet 6000 years old and that God created all things within six twenty-four-hour days. Calvin is thoroughly conversant with Augustine, agreeing with him at several points, but also rejecting his belief that God created all things instantaneously.
2.1. Classical Theistic Evolution/Evolutionary Creationist Model
The greatest problem with evolutionary creation is that it does not embrace the traditional literal interpretation of the opening chapters of the Bible. Church history shows that nearly all Christians have understood Gen 1–3 to be a basic record of actual events in the past. Specifically, most have believed that Gen 2 reveals that human history began with Adam and Eve.
[W]e can be confident that finding evidence that we were created independently of other animals or that we descend from only two people just isn’t going to happen. Some ideas in science are so well supported that it is highly unlikely new evidence will substantially modify them, and these are among them. The sun is at the center of our solar system, humans evolved, and we evolved as a population.
2.1.1. Responses
While the story is still unfolding, we conclude that the genomes of humans and other complex organisms are not full of junk but rather are highly compact information suites that are largely devoted to the specification of regulatory RNAs. These RNAs drive the trajectories of differentiation and development, underpin brain function and convey transgenerational memory of experience, much of it contrary to long-held conceptions of genetic programming and the dogmas of evolutionary theory.
2.2. Homo Divinus Model of Denis Alexander
2.2.1. Responses
2.3. Genealogical Adam and Eve Model of S. Joshua Swamidass
No additional miracles allowed. No appeals to divine action are permitted to explain the data or increase confidence in the hypothesis. Yes, one direct act of God is included in the hypothesis itself, but the evidential evaluation of the hypothesis cannot infer or rely upon divine action in any way.
The two findings of evolutionary science. The people outside the Garden would share common descent with the great apes, and the size of their population would never dip down to a single couple.
2.3.1. Responses
Swamidass’ view would seem to suggest logically that those individuals who were biological (but not textual) humans are qualitatively indistinct from other animals. However, in that case it makes no sense to call their deeds evil, or to postulate that they had a sense of right and wrong. Moreover, if they, as Swamidass suggests, “do wrong at times”, then does this not suggest that Adam’s fall is but one of many falls that have occurred in human history? The theological ramifications that accompany this scenario are too severe for me to entertain Swamidass’ proposal.
Swamidass zealously distances his position from earlier racist polygenetic theories. According to those accounts, racial groups that (allegedly) didn’t descend from Adam weren’t full human beings. I agree that his position is a vast improvement. However, the genealogical hypothesis remains polygenetic, at least to some degree. Swamidass defines all those outside the garden as biologically but not textually human.
This move, however, raises a host of questions: for example, in what sense are non-Adamic biological humans fully human? If these biological humans have a different origin from Adam and Eve, do they participate in original sin and salvation? Did Christ live and die for them, and were they able to experience justification by faith? And, if human beings are natural kinds—as Christians have always believed—then how is interbreeding even possible? In chapter 14, Swamidass speculates that these biological humans are made in God’s image, with minds and souls, but “they are not yet affected by Adam’s fall. They have a sense of right and wrong, written on their hearts (Rom. 2:15), but they are not morally perfect. They do wrong at times. They are subject to physical death, which prevents their wrongdoing from growing into true evil (Gen. 6:3)” (175). Although Swamidass is only speculating here, the notion of other people outside the garden, in my view, is nowhere in Scripture. In traditional Christianity, being human and being a descendant of Adam are co-extensive. As far as I can see, Swamidass’s revisionism lacks a convincing exegetical or theological basis.
2.4. Homo Heidelbergensis Model of William Lane Craig
Adam and Eve may therefore be plausibly identified as members of Homo heidelbergensis and as the founding pair at the root of all human species. Challenges to this hypothesis from population genetics fail principally because we cannot rule out on the basis of the genetic divergence exhibited by contemporary humans that our most recent common ancestors, situated more than 500 kya, are the sole genetic progenitors of the entire human race, whether past or present. The challenge of the wide geographic distribution of humanity is similarly met by situating Adam and Eve far in the past, prior to the divergence of Homo sapiens, Neanderthals, and other species, and allowing multispecies cultural evolution to proceed thereafter in response to environmental changes to produce modern human behaviors wherever their descendants are to be found.
One can … postulate instead a de novo creation of Adam and Eve. But then one faces a difficult dilemma. One must explain our genetic similarity to chimps either on the basis of repetitive divine use of a similar design plan or on the basis of considerable interbreeding with nonhumans. The first has difficulty explaining broken pseudogenes that we share with chimps … The second looks as if God condones bestiality for our forebears.
No such appeal to interbreeding [between humans and non-human hominids] is necessary if we envision Adam and Eve as emerging from a hominin population that shared common ancestry with chimpanzees and other great apes. Indeed, on the view proposed here, Adam and Eve could be our sole genetic progenitors, whose descendants never fell into bestial relations with nonhuman hominins or at least produced no descendants from such liaisons.
2.4.1. Responses
2.5. Unique Origins Design Model of Ann Gauger and Other Intelligent Design Advocates
2.5.1. Responses
2.6. Classical Old Earth Creationist Model of Reasons to Believe
2.6.1. Responses
2.7. Classical Young Earth Creationist Model
2.7.1. Responses
2.8. Old Earth/Recent Humans Hybrid Model
2.8.1. Responses
3. Discussion
Comparison of the Models
- If one cares very little about traditional theological beliefs regarding human origins and is convinced that mainstream evolutionary biology is correct, then the TE/EC model may be appealing.
- Conversely, if one places the highest priority upon traditional theological beliefs about Adam and Eve, then the YEC or Hybrid models may be most attractive.
- If one finds evolutionary science persuasive but desires a form of historical Adam and Eve, then the Homo heidelbergensis, GAE, or Homo divinus models may be persuasive.
- If one finds evolutionary claims to be scientifically weak but accepts standard dating methods, then the Unique Origins Design or OEC models seem most viable. The Hybrid model is also available to those who accept an old Earth, though it requires rejecting dating methods as regards many hominid fossils.
4. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
| 1 | The intent of this section is not to discuss or critique all aspects of modern evolutionary biology, but simply to provide scientific commentary on specific common arguments from evolutionary creationists on the specific topic of human origins. |
| 2 | Swamidass’s (2019) descriptions of the GAE model contains various other statements which indicate the close association of his GAE model with an attempt to retain a standard evolutionary viewpoint:
|
| 3 | Schaffner (2021) stated: “we eventually learn that his [Craig’s] model allows for an unspecified amount of admixture from other hominin lineages into the descendants of Adam, eliminating the need for such a tight bottleneck. A more clearly stated hypothesis in this section would have saved the reader time and frustration.” |
| 4 | The concept of intelligent design entails an infusion of information that came from an intelligent mind. In the context of this model, intelligent design could inolve de novo origin/special creation, but it could also include taking the body plan of a pre-existing organism and infusing information into the genome such that a new type of organism suddenly emerged. Thus, at the very minimum, intelligent design requires some input of information which changes the genotype and phenotype of an organism. Again, this could mean de novo origin/special creation, but it does not necessarily require this. However, even in the case that design does not involve de novo origin/special creation, something special is still happening—namely the infusion of information into the genome which produces an informational discontinuity that is beyond the abilities of natural mechanisms to produce. |
| 5 | For example, the Scofield Refererence Bible, highly influential in American Evangelicalism and Fundamentalism, advocated the gap theory and day/age theory, which accept an ancient age of the earth. However, the Scofield Bible did not place a date on Adam and Eve, and seemingly allowed or assumed a recent Adam and Eve. Its theology of origins would probably be most compatible with the Hybrid model (Section 2.8). |
| 6 | |
| 7 | This evidence is controversial to some, including proponents of the classical OEC model. See Rana and Ross (2015). |
References
- Alexander, Denis. 2008. Creation or Evolution: Do We Have to Choose. Oxford: Monarch Books. [Google Scholar]
- Alper, Joe. 2003. Rethinking Neanderthals. Smithsonian Magazine. Available online: https://www.smithsonianmag.com/science-nature/rethinking-neanderthals-83341003/ (accessed on 8 March 2023).
- Arensburg, Baruch, Anne-Marie Tillier, Bernard Vandermeersch, Henri Duday, Lynne A. Schepartz, and Yoel Rak. 1989. A Middle Palaeolithic human hyoid bone. Nature 338: 758–60. [Google Scholar] [CrossRef] [PubMed]
- Ashton, John F., ed. 2000. In Six Days: Why Fifty Scientists Choose to Believe in Creation. Green Forest: Master Books. [Google Scholar]
- Badarinarayan, Smitha Srinivasachar, and Daniel Sauter. 2021. Switching Sides: How Endogenous Retroviruses Protect Us from Viral Infections. Journal of Virology 95: e02299-20. [Google Scholar] [CrossRef]
- Barrett, Matthew, and Ardel B. Caneday. 2013. Adam: To Be or Not To Be? In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 13–36. [Google Scholar]
- Barrick, William D. 2013a. Response from the Young-Earth View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 80–85. [Google Scholar]
- Barrick, William D. 2013b. A Historical Adam: Young-Earth Creation View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 197–227. [Google Scholar]
- Bar-Yosef, Ofer. 2002. The upper paleolithic revolution. Annual Review of Anthropology 31: 363–93. [Google Scholar] [CrossRef]
- Bednarik, Robert G. 1997. The earliest evidence of ocean navigation. The International Journal of Nautical Archaeology 26: 183–91. [Google Scholar] [CrossRef]
- Buggs, Richard. 2017. Comment at Adam, Eve and Population Genetics: A Reply to Dr. Richard Buggs (Part 1). The BioLogos Forum. November 18. Available online: https://discourse.biologos.org/t/adam-eve-and-population-genetics-a-reply-to-dr-richard-buggs-part-1/37039/61 (accessed on 8 March 2023).
- Buggs, Richard. 2018a. Adam and Eve: Lessons Learned. Nature Ecology & Evolution Blog. April 14. Available online: https://ecoevocommunity.nature.com/posts/32171-adam-and-eve-lessons-learned (accessed on 8 March 2023).
- Buggs, Richard. 2018b. Comment at Adam, Eve and Population Genetics: A Reply to Dr. Richard Buggs (Part 1). The BioLogos Forum. April 25. Available online: https://discourse.biologos.org/t/adam-eve-and-population-genetics-a-reply-to-dr-richard-buggs-part-1/37039/1061 (accessed on 8 March 2023).
- Buggs, Richard. 2018c. How Similar Are Human and Chimpanzee Genomes? RichardBuggs.com. July 14. Available online: https://richardbuggs.com/2018/07/14/how-similar-are-human-and-chimpanzee-genomes/ (accessed on 28 May 2023).
- Chase, Philip G., and April Nowell. 1998. Taphonomy of a Suggested Middle Paleolithic Bone Flute from Slovenia. Current Anthropology 39: 549–53. [Google Scholar] [CrossRef]
- Chazan, Michael. 2017. Toward a Long Prehistory of Fire. Current Anthropology 58: S351–S359. [Google Scholar] [CrossRef]
- Cheetham, Seth W., Geoffrey J. Faulkner, and Marcel E. Dinger. 2020. Overcoming challenges and dogmas to understand the functions of pseudogenes. Nature Reviews Genetics 21: 191–201. [Google Scholar] [CrossRef]
- Chuong, Edward B. 2018. The placenta goes viral: Retroviruses control gene expression in pregnancy. PLoS Biology 16: e3000028. [Google Scholar] [CrossRef]
- Cohen, Jon. 2007. Relative Differences: The Myth of 1%. Science 316: 1836. [Google Scholar] [CrossRef]
- Collins, C. John. 2011. Did Adam and Eve Really Exist Who They Were and Why You Should Care. Wheaton: Crossway. [Google Scholar]
- Collins, C. John. 2013. Historical Adam: Old Earth Creation View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 143–75. [Google Scholar]
- Collins, C. John. 2018. Reading Genesis Well: Navigating History, Poetry, Science, and Truth in Genesis 1–11. Grand Rapids: Zondervan Academic. [Google Scholar]
- Collins, Francis. 2006. The Language of God: A Scientist Presents Evidence for Belief. New York: Free Press. [Google Scholar]
- Conley, Andrew B., Jittima Piriyapongsa, and I. King Jordan. 2008. Retroviral promoters in the human genome. Bioinformatics 24: 1563–67. [Google Scholar] [CrossRef]
- Craig, William Lane. 2000. The Kalām Cosmological Argument. Eugene: Wipf and Stock. [Google Scholar]
- Craig, William Lane. 2001. Time and the Metaphysics of Relativity. Dordecht: Kluwer Academic. [Google Scholar]
- Craig, William Lane. 2021. In Quest of the Historical Adam: A Biblical and Scientific Exploration. Grand Rapids: Eerdmans. [Google Scholar]
- Craig, William Lane, and James Porter Moreland. 2012. The Blackwell Companion to Natural Theology. Sussex: Wiley-Blackwell. [Google Scholar]
- Craig, William Lane, and Quentin Smith. 1995. Theism, Atheism, and Big Bang Cosmology. Oxford: Clarendon Press. [Google Scholar]
- D’Anastasio, Ruggero, Stephen Wroe, Claudio Tuniz, Lucia Mancini, Deneb T. Cesana, Diego Dreossi, Mayoorendra Ravichandiran, Marie Attard, William C. H. Parr, Anne Agur, and et al. 2013. Micro-Biomechanics of the Kebara 2 Hyoid and Its Implications for Speech in Neanderthals. PLoS ONE 8: e82261. [Google Scholar] [CrossRef] [PubMed]
- Daae, H. Donald. 2010. Bridging the Gap: The 7th Day. Bloomington: Westbow Press. [Google Scholar]
- Daae, H. Donald. 2012. Bridging the Gap: The First 6 Days (Revised Edition). Bloomington: Westbow Press. [Google Scholar]
- Davis, Ted. 2012. Theistic Evolution: History and Beliefs. BioLogos. October 15. Available online: https://biologos.org/series/science-and-the-bible/articles/theistic-evolution-history-and-beliefs (accessed on 9 March 2023).
- Dembski, William A. 2009. The End of Christianity: Finding a Good God in an Evil World. Nashville: B&H Academic. [Google Scholar]
- Donald, Alistair. 2009. Evolution and the Church. In Should Christians Embrace Evolution: Biblical and Scientific Responses. Edited by Norman C. Nevin. Phillipsburg: P&R Publishing, pp. 15–26. [Google Scholar]
- Durrett, Rick, and Deena Schmidt. 2008. Waiting for Two Mutations: With Applications to Regulatory Sequence Evolution and the Limits of Darwinian Evolution. Genetics 180: 1501–9. [Google Scholar] [CrossRef] [PubMed]
- Dykes, Sue. 2020a. What about the Genetic Similarity between Humans and Chimps? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 167–75. [Google Scholar]
- Dykes, Sue. 2020b. Are the Hominin Fossils Evidence for Human Evolution? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 177–87. [Google Scholar]
- Dykes, Sue. 2020c. Did Humans and Neanderthals Interbreed? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 189–99. [Google Scholar]
- Dykes, Sue. 2020d. Did Neanderthals Create Art? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 201–12. [Google Scholar]
- ENCODE Project Consortium. 2012. An Integrated Encyclopedia of DNA Elements in the Human Genome. Nature 489: 57–74. [Google Scholar] [CrossRef]
- Everett, Daniel. 2018. Did Homo erectus speak? Aeon. February 28. Available online: https://aeon.co/essays/tools-and-voyages-suggest-that-homo-erectus-invented-language (accessed on 24 March 2023).
- Falk, Darrel. 2004. Coming to Peace with Science: Bridging the Worlds Between Faith and Biology. Downers Grove: IVP Academic. [Google Scholar]
- Falk, Darrel. 2017. Hominid Fossils (evolutionary view). In Dictionary of Christianity and Science. Edited by Paul Copan, Tremper Longman III, Christopher L. Reese and Michael G. Strauss. Grand Rapids: Zondervan, pp. 355–58. [Google Scholar]
- Fiddes, Ian T., Gerrald A. Lodewijk, Meghan Mooring, Colleen M. Bosworth, Adam D. Ewing, Gary L. Mantalas, Adam M. Novak, Anouk van den Bout, Alex Bishara, Jimi L. Rosenkrantz, and et al. 2018. Human-Specific NOTCH2NL Genes Affect Notch Signaling and Cortical Neurogenesis. Cell 173: 1356–69. [Google Scholar] [CrossRef] [PubMed]
- Folger, Tim, and Shanti Menon. 1997. …Or Much Like Us? Discover Magazine. Available online: https://www.discovermagazine.com/planet-earth/or-much-like-us (accessed on 8 March 2023).
- Gates, Alexander J., Deisy Morselli Gysi, Manolis Kellis, and Albert-László Barabási. 2022. A wealth of discovery built on the Human Genome Project—By the numbers. Nature 590: 212–15. [Google Scholar] [CrossRef]
- Gauger, Ann, Douglas Axe, and Casey Luskin. 2012. Science and Human Origins. Seattle: Discovery Institute Press. [Google Scholar]
- Gibbons, Ann. 1998. Ancient Island Tools Suggest Homo erectus Was a Seafarer. Science 279: 1635–37. [Google Scholar] [CrossRef]
- Gray, Gorman. 1997. The Age of the Universe: What Are the Biblical Limits? Washougal: Morning Star. [Google Scholar]
- Grudem, Wayne. 2017a. Biblical and Theological Introduction: The Incompatibility of Theistic Evolution with the Biblical Account of Creation and with Important Christian Doctrines. In Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Edited by J. P. Moreland, Stephen C. Meyer, Christopher Shaw, Ann K. Gauger and Wayne Grudem. Wheaton: Crossway, pp. 61–77. [Google Scholar]
- Grudem, Wayne. 2017b. Theistic Evolution Undermines Twelve Creation Events and Several Crucial Christian Doctrines. In Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Edited by J. P. Moreland, Stephen C. Meyer, Christopher Shaw, Ann K. Gauger and Wayne Grudem. Wheaton: Crossway, pp. 783–837. [Google Scholar]
- Habib, Adella M., Andrei L. Okorokov, Matthew N. Hill, Jose T. Bras, Man-Cheung Lee, Shengnan Li, Samuel J Gossage, Marie van Drimmelen, Maria Morena, Henry Houlden, and et al. 2019. Microdeletion in a FAAH pseudogene identified in a patient with high anandamide concentrations and pain insensitivity. British Journal of Anaesthesia 123: e249–e253. [Google Scholar] [CrossRef]
- Ham, Ken. 2006. Could God Really Have Created Everything in Six Days? In The New Answers Book 1: Over 25 Questions on Creation/Evolution and the Bible. Edited by Ken Ham. Green Forest: Master Books, pp. 88–112. [Google Scholar]
- Hawks, John, Keith Hunley, Sang-Hee Lee, and Milford Wolpoff. 2000. Population bottlenecks and Pleistocene human evolution. Molecular Biology and Evolution 17: 2–22. [Google Scholar] [CrossRef]
- Hayashi, H., T. Arao, Y. Togashi, H. Kato, Y. Fujita, M. A. De Velasco, H. Kimura, K. Matsumoto, K. Tanaka, I. Okamoto, and et al. 2015. The OCT4 pseudogene POU5F1B is amplified and promotes an aggressive phenotype in gastric cancer. Oncogene 34: 199–208. [Google Scholar] [CrossRef]
- Henry, Hugh, and Daniel Dyke. 2012. From Noah to Abraham to Moses: Evidence of Genealogical Gaps in Mosaic Literature. Reasons to Believe. July 23. Available online: https://reasons.org/explore/publications/articles/from-noah-to-abraham-to-moses-evidence-of-genealogical-gaps-in-mosaic-literature-part-1 (accessed on 9 March 2023).
- Hirotsune, Shinji, Noriyuki Yoshida, Amy Chen, Lisa Garrett, Fumihiro Sugiyama, Satoru Takahashi, Ken-ichi Yagami, Anthony Wynshaw-Boris, and Atsushi Yoshiki. 2003. An expressed pseudogene regulates the messenger-RNA stability of its homologous coding gene. Nature 423: 91–96. [Google Scholar] [CrossRef]
- Hlubik, Sarah. Francesco Berna, Craig Feibel, David Braun, and John W. K. Harris. 2017. Researching the Nature of Fire at 1.5 Mya on the Site of FxJj20 AB, Koobi Fora, Kenya, Using High-Resolution Spatial Analysis and FTIR Spectrometry. Current Anthropology 58: S243–S257. [Google Scholar] [CrossRef]
- Hössjer, Ola, and Ann Gauger. 2019. A Single-Couple Human Origin is Possible. BIO-Complexity 2019: 1–20. [Google Scholar] [CrossRef]
- Hössjer, Ola, Ann Gauger, and Colin Reeves. 2016a. Genetic Modeling of Human History Part 1: Comparison of Common Descent and Unique Origin Approaches. BIO-Complexity 2016: 1–15. [Google Scholar] [CrossRef]
- Hössjer, Ola, Ann Gauger, and Colin Reeves. 2016b. Genetic Modeling of Human History Part 2: A Unique Origin Algorithm. BIO-Complexity 2016: 1–36. [Google Scholar] [CrossRef]
- Hössjer, Ola, Günter Bechly, and Ann Gauger. 2021. On the waiting time until coordinated mutations get fixed in regulatory sequences. Journal of Theoretical Biology 524: 110657. [Google Scholar] [CrossRef]
- Jacques, Pierre-Étienne, Justin Jeyakani, and Guillaume Bourque. 2013. The Majority of Primate-Specific Regulatory Sequences Are Derived from Transposable Elements. PLoS Genetics 9: e1003504. [Google Scholar] [CrossRef] [PubMed]
- Jeanson, Nathanial T. 2017. Replacing Darwin: The New Origin of Species. Green Forest: Master Books. [Google Scholar]
- Ji, Zhe, Ruisheng Song, Aviv Regev, and Kevin Struhl. 2015. Many lncRNAs, 5′UTRs, and Pseudogenes Are Translated and Some Are Likely to Express Functional Proteins. eLife 4: e08890. [Google Scholar] [CrossRef]
- Jönsson, Marie E., Raquel Garza, Yogita Sharma, Rebecca Petri, Erik Södersten, Jenny G. Johansson, Pia A. Johansson, Diahann A. M. Atacho, Karolina Pircs, Sofia Madsen, and et al. 2021. Activation of endogenous retroviruses during brain development causes an inflammatory response. The EMBO Journal 40: e106423. [Google Scholar] [CrossRef] [PubMed]
- Keathley, Kenneth, and Mark Rooker. 2014. 40 Questions about Creation and Evolution. Grand Rapids: Kregel Academic. [Google Scholar]
- Kelly, Robert L., and David Hurst Thomas. 2010. Archaeology, 5th ed. Belmont: Wadsworth Cengage Learning. [Google Scholar]
- Kidner, Derek. 1967. Genesis: An Introduction and Commentary. Downers Grove: InterVarsity Press. [Google Scholar]
- Kim, Min-Sik, Sneha M. Pinto, Derese Getnet, Raja Sekhar Nirujogi, Srikanth S. Manda, Raghothama Chaerkady, Anil K. Madugundu, Dhanashree S. Kelkar, Ruth Isserlin, Shobhit Jain, and et al. 2014. A Draft Map of the Human Proteome. Nature 509: 575–81. [Google Scholar] [CrossRef]
- Kitchen, Kenneth Anderson. 1966. Ancient Orient and Old Testament. Downers Grove: InterVarsity Press. [Google Scholar]
- Kovalenko, T. F., and L. I. Patrushev. 2018. Pseudogenes as Functionally Significant Elements of the Genome. Biochemistry (Moscow) 83: 1332–49. [Google Scholar] [CrossRef]
- Lamoureux, Denis O. 2009. I Love Jesus & Accept Evolution. Eugene: Wipf and Stock. [Google Scholar]
- Lieberman, Daniel E., David R. Pilbeam, and Richard W. Wrangham. 2009. The Transition from Australopithecus to Homo. In Transitions in prehistory: Essays in Honor of Ofer Bar-Yosef. Edited by John J. Shea and Daniel E. Lieberman. Cambridge: Oxbow Books, pp. 1–22. [Google Scholar]
- Luskin, Casey. 2011. BioLogos Blog Author Helps Darwin’s Universal Acid Burn God Right Out of Religion. Evolution News. November 3. Available online: https://evolutionnews.org/2011/11/biologos_blog_author_helps_dar/ (accessed on 24 March 2023).
- Luskin, Casey. 2014. The New Theistic Evolutionists: BioLogos and the Rush to Embrace the ‘Consensus’. Christian Research Journal 37: 32–41. Available online: https://www.equip.org/article/new-theistic-evolutionists-biologos-rush-embrace-consensus/ (accessed on 8 March 2023).
- Luskin, Casey. 2022. Evolutionary models of palaeoanthropology, genetics and psychology fail to account for human origins: A review. In Science and Faith in Dialogue. Edited by Frikkie Van Niekerk and Nico Vorster. Cape Town: AOSIS, pp. 243–81. [Google Scholar] [CrossRef]
- Ma, Yanni, Siqi Liu, Jie Gao, Chunyan Chen, Xin Zhang, Hao Yuan, Zhongyang Chen, Xiaolin Yin, Chenguang Sun, Yanan Mao, and et al. 2021. Genome-wide Analysis of Pseudogenes Reveals HBBP1’s Human-specific Essentiality in Erythropoiesis and Implication in β-Thalassemia. Developmental Cell 56: 478–93. [Google Scholar] [CrossRef]
- Madueme, Hans. 2020. Evolution and Historical Adam? A Provocative but Unconvincing Attempt. The Gospel Coalition. March 2. Available online: https://www.thegospelcoalition.org/reviews/genealogical-adam-eve-swamidass/ (accessed on 8 March 2023).
- Mattick, John, and Paulo Amaral. 2023. RNA: The Epicenter of Genetic Information. Boca Raton: CRC/Taylor & Francis Press. [Google Scholar]
- Mayr, Ernst. 2004. What Makes Biology Unique?: Considerations on the Autonomy of a Scientific Discipline. Cambridge: Cambridge University Press. [Google Scholar]
- McLatchie, Jonathan. 2020. The Search for Adam and Eve: Human Origins According to Scripture and Science. JonathanMclatchi.com. December 8. Available online: https://jonathanmclatchie.com/the-search-for-adam-and-eve-human-origins-according-to-scripture-and-science/ (accessed on 8 March 2023).
- Mellars, Paul. 2004. Neanderthals and the modern human colonization of Europe. Nature 432: 461–65. [Google Scholar] [CrossRef] [PubMed]
- Meyer, Stephen C. 2009. Signature in the Cell: DNA and the Evidence for Intelligent Design. New York: Harper One. [Google Scholar]
- Miller, Kenneth. 2005. Day 1 AM testimony, Kitzmiller v. Dover trial. Harrisburg, PA: Middle District Court of Pennsylvania, September 26. Available online: https://web.archive.org/web/20220219142105/https://ncse.ngo/files/pub/legal/kitzmiller/trial_transcripts/2005_0926_day1_am.pdf (accessed on 3 June 2023).
- Moleirinho, Ana, Susana Seixas, Alexandra M. Lopes, Celeste Bento, Maria J. Prata, and António Amorim. 2013. Evolutionary Constraints in the β-Globin Cluster: The Signature of Purifying Selection at the δ-Globin (HBD) Locus and Its Role in Developmental Gene Regulation. Genome Biology and Evolution 5: 559–71. [Google Scholar] [CrossRef] [PubMed]
- Morris, Henry M. 1976. The Genesis Record: A Scientific and Devotional Commentary on the Book of Beginnings. Grand Rapids: Baker Book House. [Google Scholar]
- Morris, Henry M. 1977. The Beginning of the World. Denver: Ascent Books. [Google Scholar]
- Muro, Enrique M., Nancy Mah, and Miguel A. Andrade-Navarro. 2011. Functional evidence of post-transcriptional regulation by pseudogenes. Biochimie 93: 1916–21. [Google Scholar] [CrossRef] [PubMed]
- Nowell, April. 2006. From a Paleolithic art to Pleistocene visual cultures (Introduction to two special issues on “Advances in the study of Pleistocene imagery and symbol use’). Journal of Archaeological Method and Theory 13: 239–49. [Google Scholar] [CrossRef]
- Ostling, Richard N. 2011. The Search for the Historical Adam. Christianity Today. June 3. Available online: https://www.christianitytoday.com/ct/2011/june/historicaladam.html (accessed on 8 March 2023).
- Pain, Debleena, Gung-Wei Chirn, Christopher Strassel, and Daniel M. Kemp. 2005. Multiple Retropseudogenes from Pluripotent Cell-specific Gene Expression Indicates a Potential Signature for Novel Gene Identification. The Journal of Biological Chemistry 280: 6265–68. [Google Scholar] [CrossRef]
- Pennisi, Elizabeth. 2012. ENCODE Project Writes Eulogy for Junk DNA. Science 377: 1160–61. [Google Scholar] [CrossRef]
- Piehler, Armin P., Marit Hellum, Jürgen J. Wenzel, Ellen Kaminski, Kari Bente Foss Haug, Peter Kierulf, and Wolfgang E. Kaminski. 2008. The human ABC transporter pseudogene family: Evidence for transcription and gene-pseudogene interference. BMC Genomics 9: 165. [Google Scholar] [CrossRef]
- Pink, Ryan Charles, Kate Wicks, Daniel Paul Caley, Emma Kathleen Punch, Laura Jacobs, and David Raul Francisco Carter. 2011. Pseudogenes: Pseudo-functional or key regulators in health and disease? RNA 17: 792–98. [Google Scholar] [CrossRef]
- Poliseno, Laura. 2012. Pseudogenes: Newly Discovered Players in Human Cancer. Science Signaling 5: re5. [Google Scholar] [CrossRef]
- Poliseno, Laura, Leonardo Salmena, Jiangwen Zhang, Brett Carver, William J. Haveman, and Pier Paolo Pandolfi. 2010. A coding-independent function of gene and pseudogene mRNAs regulates tumour biology. Nature 465: 1033–38. [Google Scholar] [CrossRef]
- Prieto-Godino, Lucia L., Raphael Rytz, Benoîte Bargeton, Liliane Abuin, J. Roman Arguello, Matteo Dal Peraro, and Richard Benton. 2016. Olfactory receptor pseudo-pseudogenes. Nature 539: 93–97. [Google Scholar] [CrossRef] [PubMed]
- Rana, Fazale. 2018. When Did Modern Human Brains—and the Image of God—Appear? Reasons to Believe. November 14. Available online: https://reasons.org/explore/blogs/the-cells-design/when-did-modern-human-brains-and-the-image-of-god-appear (accessed on 3 June 2023).
- Rana, Fazale. 2020a. Is Evolution Really a Problem for the Christian Faith? In Thinking About Evolution: 25 Questions Christians Want Answered. Edited by Anjeanette Roberts, Fazale Rana, Sue Dykes and Mark Perez. Covina: RTB, pp. 65–73. [Google Scholar]
- Rana, Fazale. 2020b. Answering Scientific Questions on Neanderthal-Human Interbreeding, Part 1. Reasons.org Posted August 5. Available online: https://reasons.org/explore/blogs/the-cells-design/answering-scientific-questions-on-neanderthal-human-interbreeding-part-1 (accessed on 11 May 2023).
- Rana, Fazale. 2021. Ancient Cave Art Strengthens Evidence for the Image of God. Reasons to Believe. March 10. Available online: https://reasons.org/explore/blogs/the-cells-design/ancient-cave-art-strengthens-evidence-for-the-image-of-god (accessed on 3 June 2023).
- Rana, Fazale, and Hugh Ross. 2005. Who Was Adam: A Creation Model Approach to the Origin of Man. Colorado Springs: Navpress. [Google Scholar]
- Rana, Fazale, and Hugh Ross. 2015. Who Was Adam: A Creation Model Approach to the Origin of Man (Second Expanded Edition). Covina: RTB Press. [Google Scholar]
- Rapicavoli, Nicole A., Kun Qu, Jiajing Zhang, Megan Mikhail, Remi-Martin Laberge, and Howard Y. Chang. 2013. A mammalian pseudogene lncRNA at the interface of inflammation and anti-inflammatory therapeutics. eLife 2: e00762. [Google Scholar] [CrossRef] [PubMed]
- Reeves, Michael. 2009. Adam and Eve. In Should Christians Embrace Evolution: Biblical and Scientific Responses. Edited by Norman C. Nevin. Phillipsburg: P&R Publishing, pp. 43–56. [Google Scholar]
- Rice, Stanley A. 2007. Encyclopedia of Evolution. New York: Checkmark Books. [Google Scholar]
- Robson, Michael I., and Stefan Mundlos. 2019. Jumping retroviruses nudge TADs apart. Nature Genetics 51: 1304–5. [Google Scholar] [CrossRef] [PubMed]
- Ross, Hugh. 2001. The Genesis Question: Scientific Advances and the Accuracy of Genesis. Colorado Springs: NavPress. [Google Scholar]
- Ross, Hugh. 2014. Navigating Genesis: A Scientist’s Journey through Genesis 1–11. Covina: RTB Press. [Google Scholar]
- Ross, Hugh. 2016. When Did God Create Adam and Eve? Reasons.org Posted October 19. Available online: https://reasons.org/explore/blogs/todays-new-reason-to-believe-es/when-did-god-create-adam-and-eve (accessed on 29 May 2023).
- Rossano, Matt J. 2010. Supernatural Selection: How Religion Evolved. Oxford: Oxford University Press. [Google Scholar]
- Sakashita, Akihiko, Tomohiro Kitano, Hirotsugu Ishizu, Youjia Guo, Harumi Masuda, Masaru Ariura, Kensaku Murano, and Haruhiko Siomi. 2023. Transcription of MERVL retrotransposons is required for preimplantation embryo development. Nature Genetics 55: 484–95. [Google Scholar] [CrossRef]
- Salmena, Leonardo, Laura Poliseno, Yvonne Tay, Lev Kats, and Pier Paolo Pandolfi. 2011. A ceRNA Hypothesis: The Rosetta Stone of a Hidden RNA Language? Cell 146: 353–58. [Google Scholar] [CrossRef]
- Sanford, John. 2005. Genetic Entropy & The Mystery of the Genome. Lima: Elim. [Google Scholar]
- Sanford, John, Rob Carter, Wes Brewer, John Baumgardner, Bruce Potter, and John Potter. 2018. Adam and Eve, Designed Diversity, and Allele Frequencies. In Proceedings of the Eighth International Conference on Creationism. Edited by J.H. Whitmore. Pittsburgh: Creation Science Fellowship, pp. 200–16. [Google Scholar]
- Schaffner, Stephen. 2021. Adam, Eve, and the evolution of humankind. Science 374: 162. [Google Scholar] [CrossRef]
- Scott, Eugenie. 2007. What Do Creationists Believe about Human Evolution?|Dr. Eugenie Scott. The Leakey Foundation, YouTube Posted November 29, 2021. Available online: https://www.youtube.com/watch?v=iZhO7_GhDH0 (accessed on 8 March 2023).
- Seaman, Josiah, and Richard Buggs. 2020. FluentDNA: Nucleotide visualization of whole genomes, annotations, and alignments. Frontiers in Genetics 11: a292. [Google Scholar] [CrossRef]
- Stitz, Maria, Cristian Chaparro, Zhigang Lu, V. Janett Olzog, Christina E. Weinberg, Jochen Blom, Alexander Goesmann, Christoph Grunau, and Christoph G. Grevelding. 2021. Satellite-Like W-Elements: Repetitive, Transcribed, and Putative Mobile Genetic Factors with Potential Roles for Biology and Evolution of Schistosoma mansoni. Genome Biology and Evolution 13: evab204. [Google Scholar] [CrossRef]
- Stott, John R.W. 1999. Understanding the Bible: Expanded Edition. Grand Rapids: Zondervan. First published 1972. [Google Scholar]
- Stringer, Christopher B. 1992. Evolution of Early Humans. In Cambridge Encyclopedia of Human Evolution. Edited by Steve Jones, Robert Martin and David Pilbeam. Cambridge: Cambridge University Press, pp. 241–51. [Google Scholar]
- Suzuki, Ikuo K., David Gacquer, Roxane Van Heurck, Devesh Kumar, Marta Wojno, Angéline Bilheu, Adèle Herpoel, Nelle Lambert, Julian Cheron, Franck Polleux, and et al. 2018. Human-Specific NOTCH2NL Genes Expand Cortical Neurogenesis through Delta/Notch Regulation. Cell 173: 1370–84. [Google Scholar] [CrossRef]
- Swamidass, S. Joshua. 2017. Heliocentric Certainty Against a Bottleneck of Two? Peaceful Science. Available online: https://discourse.peacefulscience.org/t/heliocentric-certainty-against-a-bottleneck-of-two/61 (accessed on 8 March 2023).
- Swamidass, S. Joshua. 2019. The Genealogical Adam and Eve: The Surprising Science of Universal Ancestry. Downers Grove: IVP Academic. [Google Scholar]
- Tam, Oliver H., Alexei A. Aravin, Paula Stein, Angelique Girard, Elizabeth P. Murchison, Sihem Cheloufi, Emily Hodges, Martin Anger, Ravi Sachidanandam, Richard M. Schultz, and et al. 2008. Pseudogene-derived small interfering RNAs regulate gene expression in mouse oocytes. Nature 453: 534–38. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, João C., Guy S. Jacobs, Chris Stringer, Jonathan Tuke, Georgi Hudjashov, Gludhug A. Purnomo, Herawati Sudoyo, Murray P. Cox, Raymond Tobler, Chris S. M. Turney, and et al. 2021. Widespread Denisovan ancestry in Island Southeast Asia but no evidence of substantial super-archaic hominin admixture. Nature Ecology & Evolution 5: 616–24. [Google Scholar] [CrossRef]
- Thorvaldsen, Steinar, and Ola Hössjer. 2020. Using statistical methods to model the fine-tuning of molecular machines and systems. Journal of Theoretical Biology 501: 110352. [Google Scholar] [CrossRef]
- Toth, Nicholas, and Kathy Schick. 2015. Overview of paleolithic archaeology. In Handbook of Paleoanthropology: Principles, Methods, and Approaches, 2nd ed. Edited by Winfried Henke and Ian Tattersall. Heidelberg: Springer, pp. 2441–64. [Google Scholar] [CrossRef]
- Travis, Melissa Cain. 2021. William Lane Craig Explores the Headwaters of the Human Race. Christianity Today. September 20. Available online: https://www.christianitytoday.com/ct/2021/october/william-lane-craig-quest-historical-adam-genesis-origins.html (accessed on 8 March 2023).
- Trinkaus, Erik, and Cidália Duarte. 2003. The Hybrid Child from Portugal. Scientific American 288: 32. [Google Scholar]
- Trinkaus, Erik, and Pat Shipman. 1993. Neandertals: Images of Ourselves. Evolutionary Anthropology 1: 194–201. [Google Scholar] [CrossRef]
- Troskie, Robin-Lee, Geoffrey J. Faulkner, and Seth W. Cheetham. 2021. Processed pseudogenes: A substrate for evolutionary innovation. BioEssays 43: 2100186. [Google Scholar] [CrossRef] [PubMed]
- University of Michigan News Service. 2000. New Study Suggests Big Bang Theory of Human Evolution. January 10. Available online: https://news.umich.edu/new-study-suggests-big-bang-theory-of-human-evolution/ (accessed on 8 March 2023).
- Venema, Dennis. 2018. Comment at Adam, Eve and Population Genetics: A Reply to Dr. Richard Buggs (Part 1). The BioLogos Forum. April 25. Available online: https://discourse.biologos.org/t/adam-eve-and-population-genetics-a-reply-to-dr-richard-buggs-part-1/37039/1063 (accessed on 8 March 2023).
- Venema, Dennis, and Darrel Falk. 2010. Signature in the Pseudogenes. BioLogos. April 19. Available online: https://biologos.org/articles/signature-in-the-pseudogenes (accessed on 8 March 2023).
- Venema, Dennis, and Scot McKnight. 2017. Adam and the Genome: Reading Scripture after Genetic Science. Grand Rapids: Brazos Press. [Google Scholar]
- Villanea, Fernando A., and Joshua G. Schraiber. 2018. Multiple episodes of interbreeding between Neanderthal and modern humans. Nature Ecology & Evolution 3: 39–44. [Google Scholar] [CrossRef]
- Walton, John H. 2013. A Historical Adam: Archetypal Creation View. In Four Views on The Historical Adam. Edited by Gregory A. Boyd and Philip G. Ryken. Grand Rapids: Zondervan, pp. 89–118. [Google Scholar]
- Waters, Guy Prentiss. 2017. Theistic Evolution Is Incompatible with the Teachings of the New Testament. In Theistic Evolution: A Scientific, Philosophical, and Theological Critique. Edited by J. P. Moreland, Stephen C. Meyer, Christopher Shaw, Ann K. Gauger and Wayne Grudem. Wheaton: Crossway, pp. 879–926. [Google Scholar]
- Wen, Yan-Zi, Ling-Ling Zheng, Liang-Hu Qu, Francisco J. Ayala, and Zhao-Rong Lun. 2012. Pseudogenes Are Not Pseudo Any More. RNA Biology 9: 27–32. [Google Scholar] [CrossRef]
- White, Randall. 2003. Prehistoric Art: The Symbolic Journey of Humankind. New York: Harry N Abrams. [Google Scholar]
- Wong, Kate. 2003. Who were the Neandertals? Scientific American 288: 28–37. [Google Scholar] [CrossRef]
- Wood, Bernard, and Terry Harrison. 2011. The evolutionary context of the first hominins. Nature 470: 347–52. [Google Scholar] [CrossRef]
- Wrangham, Richard, and Rachel Carmody. 2010. Human Adaptation to the Control of Fire. Evolutionary Anthropology 19: 187–99. [Google Scholar] [CrossRef]
- 2012. ENCODE: The Rough Guide to the Human Genome. Discover. September 5. Available online: https://www.discovermagazine.com/the-sciences/encode-the-rough-guide-to-the-human-genome (accessed on 8 March 2023).
- Zhang, Jingyu, Xia Wang, Meixiang Li, Jin Han, Bing Chen, Bin Wang, and Jianwu Dai. 2006. NANOGP8 is a retrogene expressed in cancers. FEBS Journal 273: 1723–30. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Maohua, Congling Shi, Xing Gao, Xinzhi Wu, Fuyou Chen, Shuangquan Zhang, Xingkai Zhang, and John W. Olsen. 2014. On the possible use of fire by Homo erectus at Zhoukoudian, China. Chinese Science Bulletin 59: 335–43. [Google Scholar] [CrossRef]


{kind=link}
| Model | Adam and Eve Were Real Historical People Who Fell into Sin. | Adam and Eve Were Ancestral to All Living Humans. | Adam and Eve Were the Sole Initial Progenitors of All Humans. | Adam and Eve Were Specially and Miraculously Created “de novo.” | Adam and Eve Lived Recently (<10,000 Years) as Genealogies Typically Suggest. | No Physical Death before the Fall of Adam and Eve | Humans Only Interbreed with Imago Dei Humans (i.e., Bestiality Not Required). |
|---|---|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | |||||||
| Homo divinus Model | X | X | |||||
| Genealogical Adam and Eve Model | X | X | Possible, if not favored, but not required. | Quite possible, but not required. | |||
| Homo heidelbergensis Model | X | X | Strongly preferred, but not required. | Probably not. | Strongly preferred, but not required. | ||
| Unique Origins Design Model | X | X | X | Allowed, if not favored. | X | ||
| Classical Old Earth Creationist Model | X | X | X | X | This was initially strongly preferred, but was rejected after evidence of human–Neanderthal interbreeding. | ||
| Classical Young Earth Creationist Model | X | X | X | X | X | X | X |
| Old Earth/Recent Origin Hybrid Model | X | X | X | X | X | Possible, but not required. | X |
| Model | Humans Evolved via Standard/Unguided Evolutionary Mechanisms from Apelike Species. | Humans Share a Common Ancestor with Apes. | Humans Had Thousands of Primordial Ancestors and Never a Bottleneck of Two. | Humans (Homo sapiens) Are Related to Neanderthals and Denisovans. | Human Body Plan Appears before Human Intelligence. | Dating (Old Age) of Hominid Fossils Is Accurate. | Dating (Old Age) of Earth and Universe Is Accurate. |
|---|---|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | X | X | X | X | X | X | X |
| Homo divinus Model | X | X | X | X | X | X | X |
| Genealogical Adam and Eve Model | X | X | X | X | X | X | X |
| Homo heidelbergensis Model | Not clear, but seems to be possible. | X | X | X | X | X | |
| Unique Origins Design Model | X | X | X | ||||
| Classical Old Earth Creationist Model | X | X | X | ||||
| Classical Young Earth Creationist Model | X | ||||||
| Old Earth/Recent Origin Hybrid Model | X | X |
| Model | Acknowledges Waiting Times Problem for Evolving Complex Traits and the Possibility of Design. | Incorporates Population Genetics Modeling Showing Humans Can Come from an Initial Pair. | Incorporates Evidence of Pseudogene Function. | Accepts Evidence of Full Humanity of Neanderthals/Denisovans. | Acknowledges Fossil Gap between Human and Apelike Species. | Adam and Eve Aligned with First Appearance of Fossil Evidence of Humanlike Body Plan. | Adam and Eve Aligned with First Appearance of Archaeological Evidence of Humanlike Intelligence. |
|---|---|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | |||||||
| Homo divinus Model | |||||||
| Genealogical Adam and Eve Model | |||||||
| Homo heidelbergensis Model | X | X | |||||
| Unique Origins Design Model | X | X | X | X | X | Possible. | |
| Classical Old Earth Creationist Model | X | X | X | X | X | ||
| Classical Young Earth Creationist Model | X | X | X | X | X | Not if standard dating methods are adopted. | Not if standard dating methods are adopted. |
| Old Earth/Recent Origin Hybrid Model | X | X | X | X | X | Not if standard dating methods are adopted. | Not if standard dating methods are adopted. |
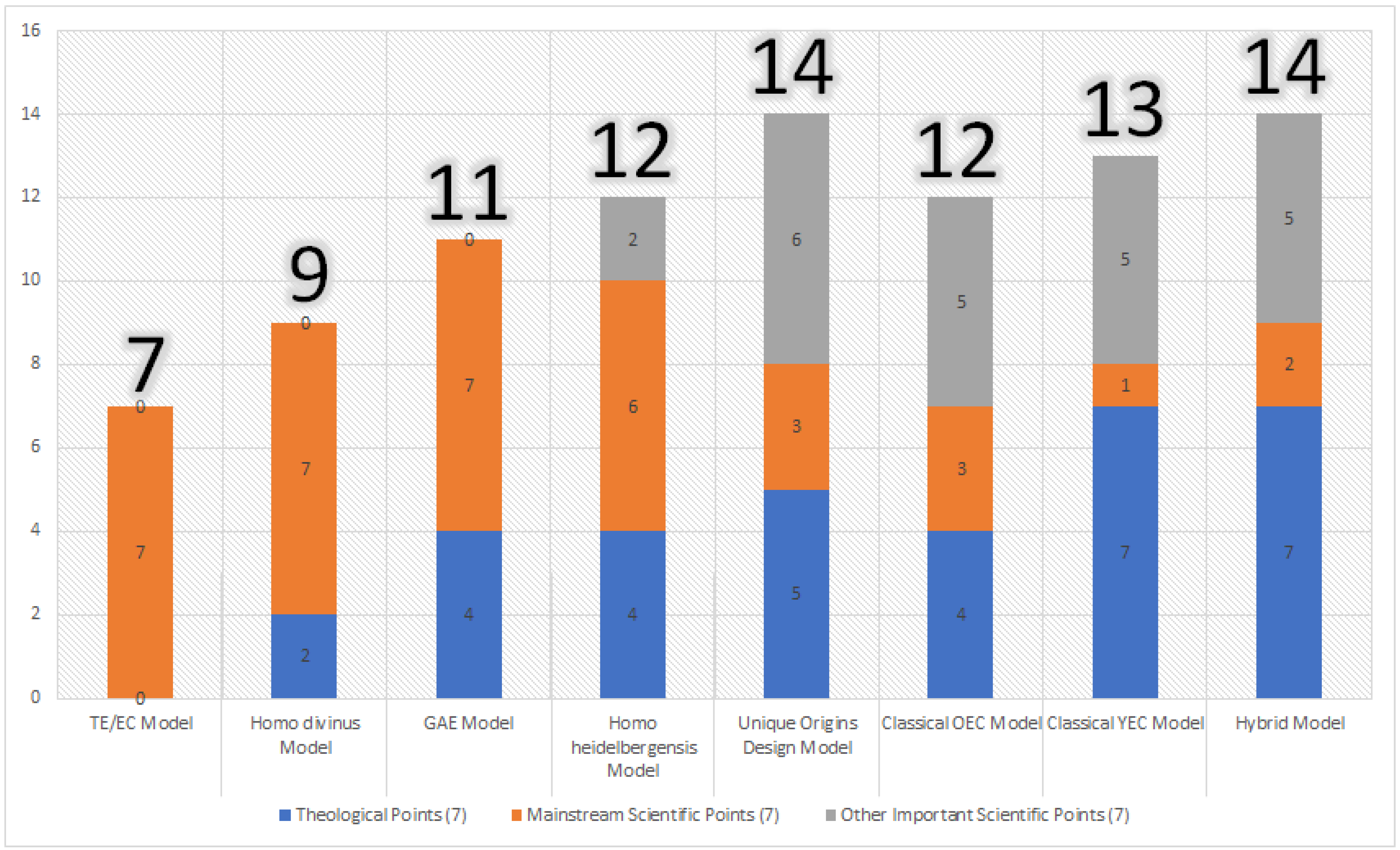
| Model | Theological Points (7) | Mainstream Scientific Points (7) | Subtotal (14) of Table 1 and Table 2 | Other Scientific Points (7) | Grand Total (21) |
|---|---|---|---|---|---|
| Classical Theistic Evolution/Evolutionary Creation Model | 0 | 7 | 7 | 0 | 7 |
| Homo divinus Model | 2 | 7 | 9 | 0 | 9 |
| Genealogical Adam and Eve Model | 4 | 7 | 11 | 0 | 11 |
| Homo heidelbergensis model | 4 | 6 | 10 | 2 | 12 |
| Unique Origins Design Model | 5 | 3 | 8 | 6 | 14 |
| Classical Old Earth Creationist Model | 4 | 3 | 7 | 5 | 12 |
| Classical Young Earth Creationist Model | 7 | 1 | 8 | 5 | 13 |
| Old Earth/Recent Origin Hybrid Model | 7 | 2 | 9 | 5 | 14 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luskin, C. Comparing Contemporary Evangelical Models Regarding Human Origins. Religions 2023, 14, 748. https://doi.org/10.3390/rel14060748
Luskin C. Comparing Contemporary Evangelical Models Regarding Human Origins. Religions. 2023; 14(6):748. https://doi.org/10.3390/rel14060748
Chicago/Turabian StyleLuskin, Casey. 2023. "Comparing Contemporary Evangelical Models Regarding Human Origins" Religions 14, no. 6: 748. https://doi.org/10.3390/rel14060748
APA StyleLuskin, C. (2023). Comparing Contemporary Evangelical Models Regarding Human Origins. Religions, 14(6), 748. https://doi.org/10.3390/rel14060748