
Does Sand Beach Nourishment Enhance the Dispersion of Non-Indigenous Species?—The Case of the Common Moon Crab, Matuta victor (Fabricius, 1781), in the Southeastern Mediterranean




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Research on Beach Nourishment and Non-Indigenous Species
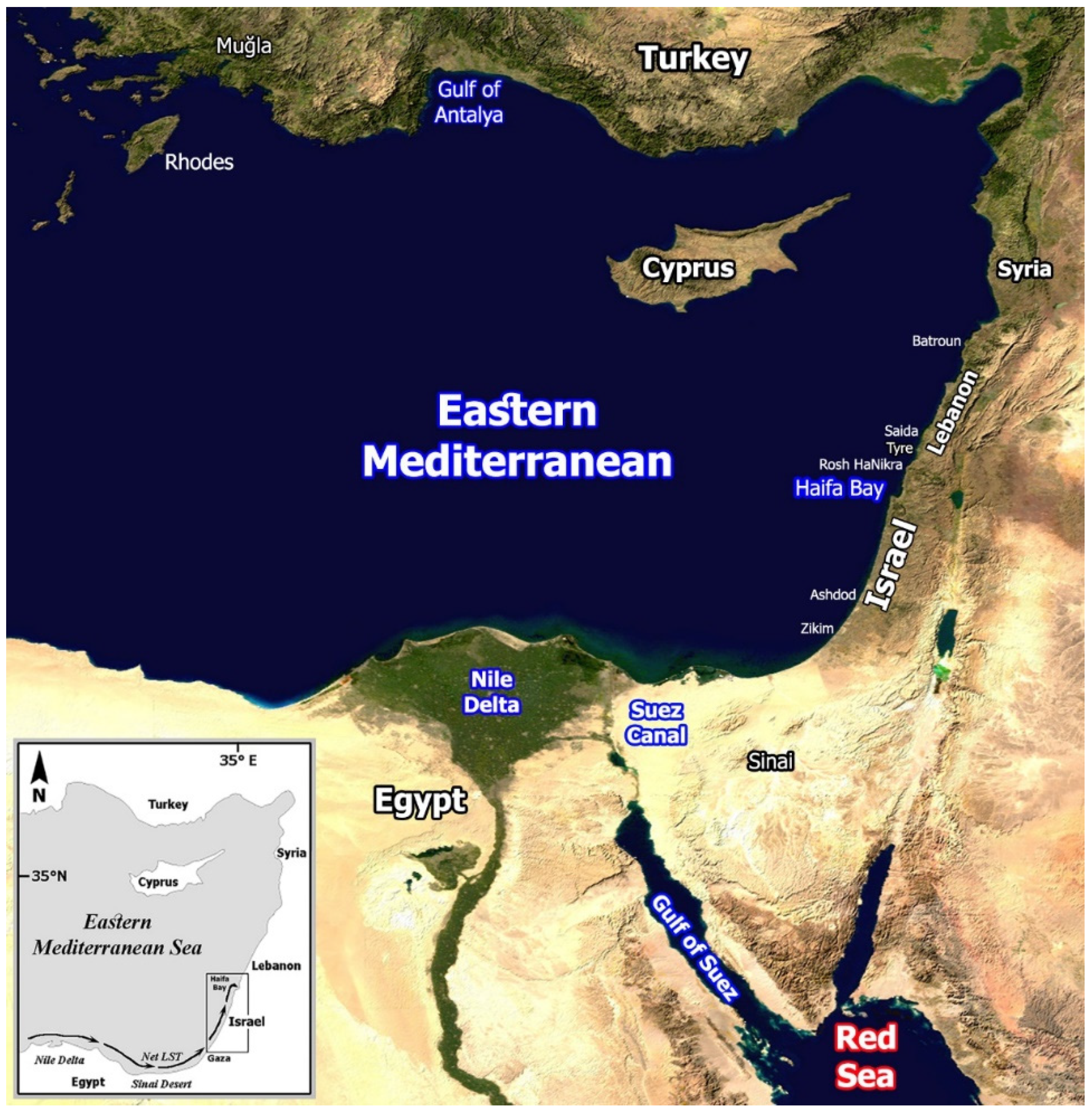
3. Study Area
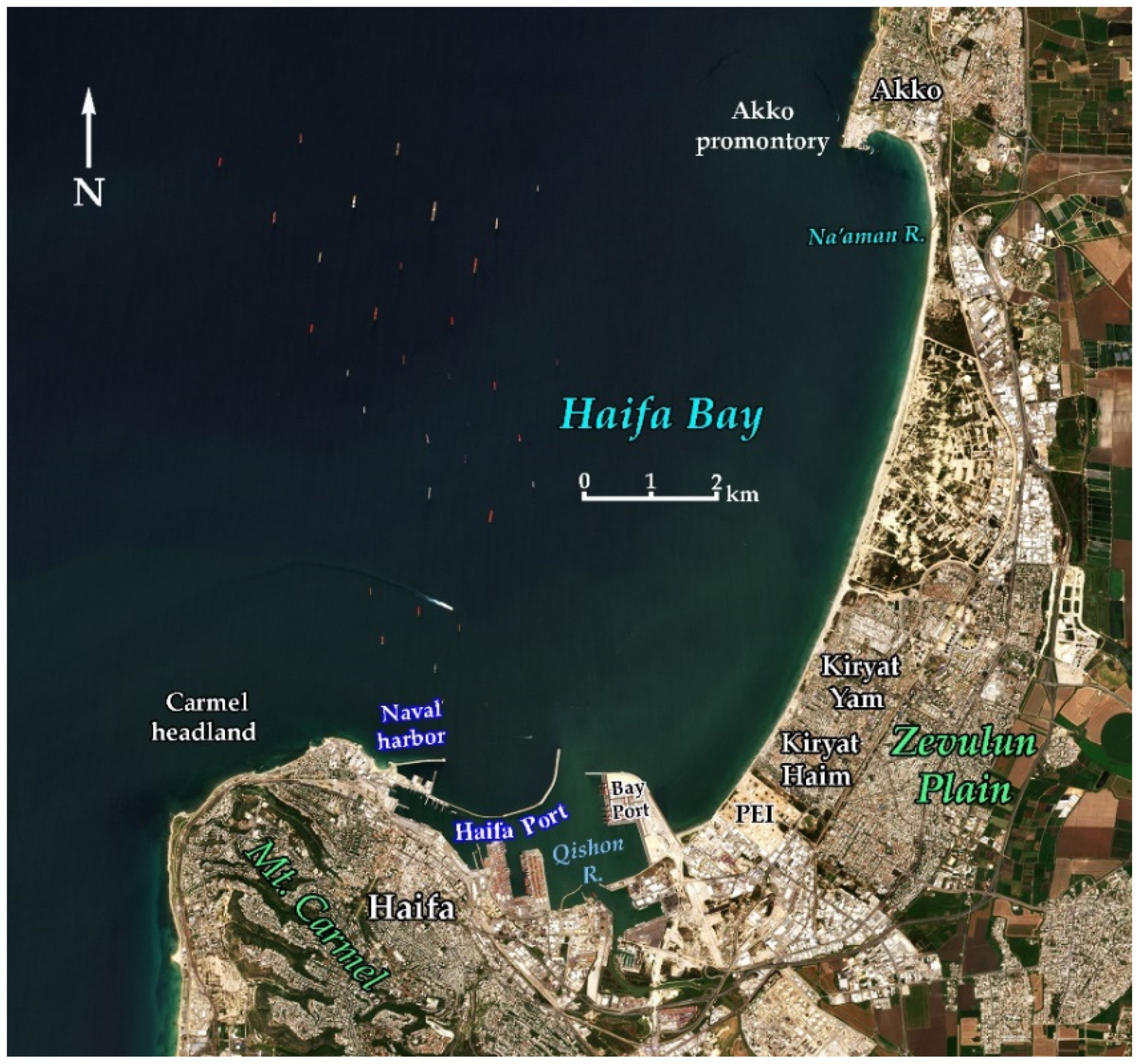
3.1. Haifa Bay Physical Setting
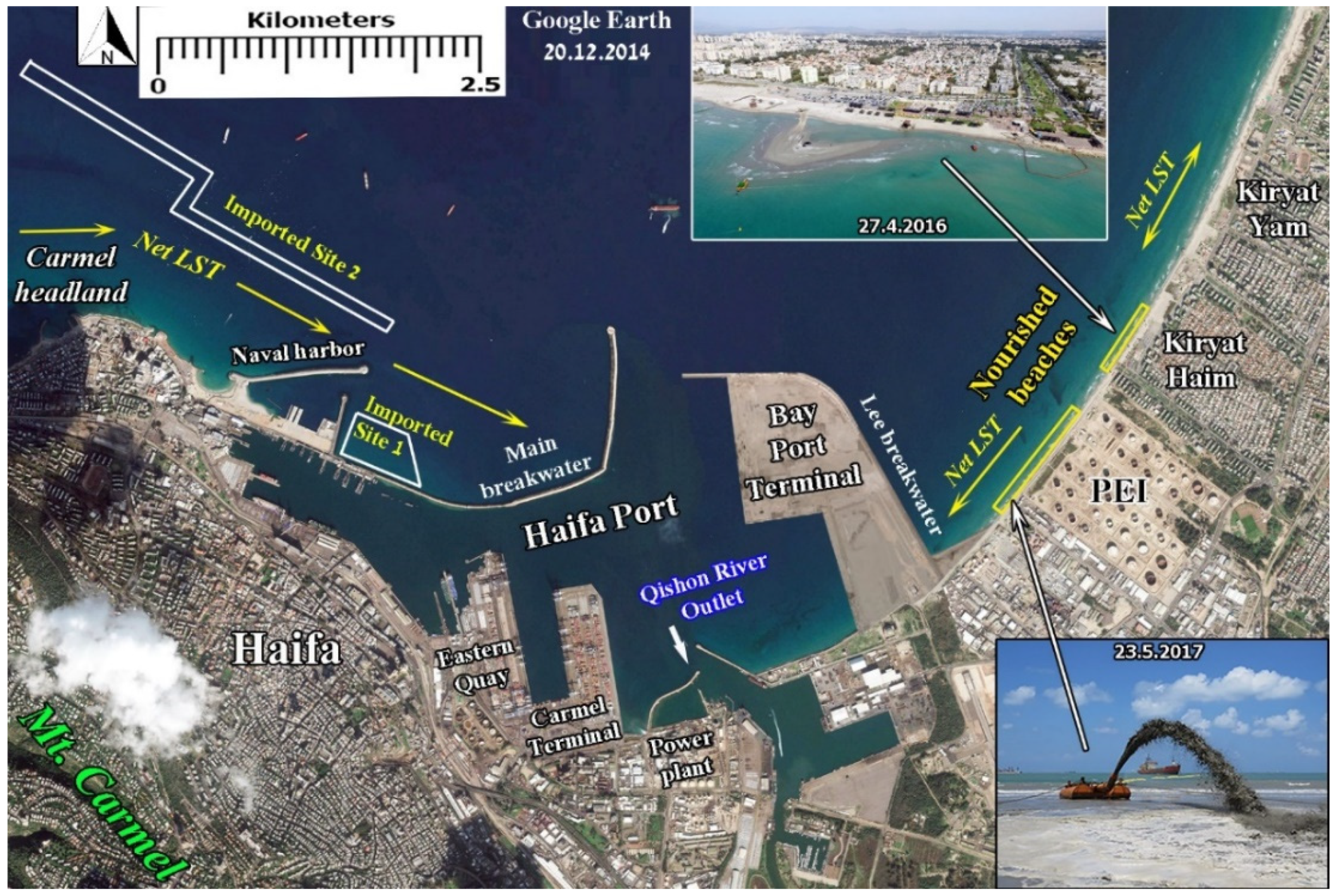
3.2. Haifa Port Main Breakwater Morphological Impact
3.3. Haifa Bay Sand Beach Nourishment Activity
4. The Erythraean Moon Crab Matuta victor and Beach Nourishment
5. The Possible Implications of the Appearance of M. victor in Haifa Bay
6. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hanson, H.; Brampton, A.; Capobianco, M.; Dette, H.H.; Hamm, L.; Laustrup, C.; Lechuga, A.; Spanhoff, R. Beach nourishment projects, practices, and objectives—A European overview. Coast. Eng. 2002, 47, 81–111. [Google Scholar] [CrossRef]
- Özhan, E. Coastal Erosion Management in the Mediterranean; UNEP, MAP, Priority Actions Programme, Regional Activity Centre, Ankara/Split: Ankara, Turkey, 2002. [Google Scholar]
- Waterman, R.E. Integrated Coastal Policy Via Building with Nature; Opmeer BV: The Hague, The Netherlands, 2008. [Google Scholar]
- Anthony, E.J.; Sabatier, F. France. In Coastal Erosion and Protection in Europe; Pranzini, E., Williams, A.T., Eds.; Routledge: London, UK, 2013; Chapter 12; pp. 227–253. [Google Scholar]
- Semeoshenkova, V.; Newton, A. Overview of erosion and beach quality issues in three southern European countries: Portugal, Spain and Italy. Ocean Coast. Manage 2015, 118, 12–121. [Google Scholar] [CrossRef]
- Vanden Eede, S.; Van Tomme, J.; De Busschere, C.; Vandegehuchte, M.L.; Sabbe, K.; Stienen, E.W.; Degraer, S.; Vincx, M.; Bonte, D. Assessing the impact of beach nourishment on the intertidal food web through the development of a mechanistic-envelope model. J. Appl. Ecol. 2014, 51, 1304–1313. [Google Scholar] [CrossRef]
- Pranzini, E.; Wetzel1, L.; Williams, A.T. Aspects of coastal erosion and protection in Europe. J. Coast. Conserv. 2015, 19, 445–459. [Google Scholar] [CrossRef]
- Dean, R.G. Beach Nourishment: Theory and Practice; World Scientific Publishing Company: Singapore, 2003; Volume 18. [Google Scholar]
- Harley, C.D.G.; Hughes, A.R.; Hultgren, K.M.; Miner, B.; Sorte, C.J.B.; Thornber, C.S.; Rodriguez, L.F.; Tomanek, L.; Williams, S.L. The impacts of climate change in coastal marine systems. Ecol. Let. 2006, 9, 228–241. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.; Gladstone, W.; Hacking, N. Australian sandy-beach ecosystems and climate change: Ecology and management. Aust. Zool. 2007, 34, 190–202. [Google Scholar] [CrossRef]
- McLachlan, A.; Brown, A.C. The Ecology of Sandy Shores; Academic Press: Burlington, MA, USA, 2006. [Google Scholar]
- Speybroeck, J.; Bonte, D.; Courtens, W.; Gheskiere, T.; Grootaert, P.; Maelfait, J.-P.; Provoost, S.; Sabbe, K.; Stienen, E.W.M.; Van Lancker, V.; et al. The Belgian sandy beach ecosystem: A review. Mar. Ecol. 2008, 29, 171–185. [Google Scholar] [CrossRef]
- Defeo, O.; McLachlan, A.; Schoeman, D.S.; Schlacher, T.A.; Dugan, J.; Jones, A.; Lastra, M.; Scapini, F. Threats to sandy beach ecosystems: A review. Estuar. Coast. Shelf. Sci. 2009, 81, 1–12. [Google Scholar] [CrossRef]
- Schlacher, T.A.; Schoeman, D.S.; Dugan, J.; Lastra, M.; Jones, A.; Scapini, F.; McLachlan, A. Sandy beach ecosystems: Key features, sampling issues, management challenges and climate change impacts. Mar. Ecol. 2008, 29, 70–90. [Google Scholar] [CrossRef]
- Dean, R.G. Compatibility of borrow material for beach fills. In Proceedings of the 14th International Conference on Coastal Engineering; ASCE: Copenhagen, Denmark, 1974; pp. 1319–1333. [Google Scholar]
- Dean, R.G. Principles of beach nourishment. In Handbook of Coastal Processes and Erosion; Komar, P.D., Ed.; CRC Press: Boca Raton, FL, USA, 1983; Chapter 11; pp. 217–232. [Google Scholar]
- Anthony, E.J. The status of beaches and shoreline development options on the French Riviera: A perspective and a prognosis. J. Coast. Conserv. 1997, 3, 169–178. [Google Scholar] [CrossRef]
- Aragonés, L.; Garcia-Barba, J.; García-Bleda, E.; López, I.; Serra, J.C. Beach nourishment impact on Posidonia oceanica: Case study of Poniente Beach (Benidorm, Spain). Ocean Eng. 2015, 107, 1–12. [Google Scholar] [CrossRef]
- Danovaro, R.; Nepote, E.; Martire, M.L.; Ciotti, C.; De Grandis, G.; Corinaldesi, C.; Carugati, L.; Cerrano, C.; Pica, D.; Di Camillo, C.G.; et al. Limited impact of beach nourishment on macrofaunal recruitment/settlement in a site of community interest in coastal area of the Adriatic Sea (Mediterranean Sea). Mar. Pollut. Bull. 2018, 128, 259–266. [Google Scholar] [CrossRef]
- Naqvi, S.; Pullen, E. Effects of Beach Nourishment and Borrowing on Marine Organisms; Miscellaneous Report No. 82–14; U.S. Army Corps of Engineers, Coastal Engineering Research Center: Fort Belvoir, VA, USA, 1982. [Google Scholar]
- Van Dolah, R.F.; Wendt, P.H.; Martore, R.M.; Levisen, M.V.; Roumillat, W.A. A Physical and Biological Monitoring Study of the Hilton Head Beach Nourishment Project. Unpublished; South Carolina Wildlife and Marine Resources Department for Town of Hilton Head Island: Hilton Head Island, SC, USA, 1992. [Google Scholar]
- Van Dolah, R.F.; Martore, R.M.; Lynch, A.E.; Levisen, M.V.; Wendt, P.H.; Whitaker, D.J.; Anderson, W.D. Final Report: Environmental Evaluation of the Folly Beach Nourishment Project; U.S. Army Corps of Engineers, Charleston District: Charleston, SC, USA, 1994. [Google Scholar]
- Peterson, C.H.; Laney, T.R.W. Biological impacts of beach nourishment. In Workshop on the Science of Beach re Nourishment; Pine Knoll Shores: Carteret County, NC, USA, 2001. [Google Scholar]
- Rice, T. The big picture: An overview of coastal resources and federal projects. In Coastal Ecosystems & Federal Activities Technical Training Symposium; US Fish and Wildlife Service: Gulf Shores, AL, USA, 2001. [Google Scholar]
- Lakshmi, E.; Priya, M.; Achari, V.S. An overview on the treatment of ballast water in ships. Ocean. Coast. Manag. 2021, 199, 105296. [Google Scholar] [CrossRef]
- Minchin, D.; Gollasch, S.; Cohen, A.N.; Hewitt, C.L.; Olenin, S. Characterizing vectors of marine invasion. In Biological Invasions in Marine Ecosystems; Rilov, G., Crooks, J.A., Eds.; Ecological Studies 204; Springer-Verlag: Berlin Heidelberg, Germany, 2009; pp. 109–116. [Google Scholar]
- Franks, S.J.; Peterson, C.J. Burial disturbance leads to facilitation among coastal dune plants. Plant. Ecol. 2003, 16, 13–21. [Google Scholar] [CrossRef]
- Rotter, A.; Klun, K.; Francé, J.; Mozetič, P.; Orlando-Bonaca, M. Non-indigenous species in the Mediterranean Sea: Turning from pest to source by developing the 8Rs model, a new paradigm in pollution mitigation. Front. Mar. Sci. 2020, 7, 178. [Google Scholar] [CrossRef]
- Spanier, E.; Galil, B.S. Lessepsian migrationִ—a continuous biogeographical process. Endeavour 1991, 15, 102–106. [Google Scholar] [CrossRef]
- Edelist, D.; Rilov, G.; Golani, D.; Carlton, J.T.; Spanier, E. Restructuring the sea: Profound shifts in the world’s most invaded marine ecosystem. Divers. Distrib. 2013, 19, 69–77. [Google Scholar] [CrossRef]
- Galil, B.S. The alien crustaceans in the Mediterranean Sea: An historical review. In the Wrong Place—Alien Marine Crustaceans: Distribution, Biology and Impacts; Galil, B.S., Clark, P.F., Carlton, J.T., Eds.; Springer: Dordrecht, The Netherlands, 2011; pp. 377–401. [Google Scholar]
- Katsanevakis, S.; Moustakas, A. Uncertainty in marine invasion science. Front. Mar. Sci. 2018, 5, 38. [Google Scholar] [CrossRef]
- Galil, B.S.; Marchini, A.; Occhipinti-Ambrogi, A. East is east and West is west? Management of marine bioinvasions in the Mediterranean Sea. Estuar. Coast. Shelf Sci. 2018, 201, 7–16. [Google Scholar] [CrossRef]
- Zenetos, A.; Cinar, M.E.; Crocetta, F.; Golani, D.; Rosso, A.; Servello, G.; Shenkar, N.; Turon, X.; Verlaque, M. Uncertainties and validation of alien species catalogues: The Mediterranean as an example. Estuar. Coast. Shelf Sci. 2017, 191, 171–187. [Google Scholar] [CrossRef]
- Galil, B.S.; Mienis, H.K.; Hoffman, R.; Goren, M. Non-indigenous species along the Israeli Mediterranean coast: Tally, policy, outlook. Hydrobiologia 2021, 848, 2011–2029. [Google Scholar] [CrossRef]
- Ben Rais Lasram, F.; Tomasini1, J.A.; Guilhaumon, F.; Romdhane, M.S.; Do Chi, T.; Mouillot, D. Ecological correlates of dispersal success of Lessepsian fishes. Mar. Ecol. Prog. Ser. 2009, 363, 273–286. [Google Scholar] [CrossRef]
- Innocenti, G.; Stasolla, G.; Mendelson, M.; Galil, B.S. Aggressive, omnivorous, invasive: The Erythraean moon crab Matuta victor (Fabricius, 1781) (Crustacea: Decapoda: Matutidae) in the eastern Mediterranean Sea. J. Nat. Hist. 2017, 51, 2133–2142. [Google Scholar] [CrossRef]
- Bitan, M.; Zviely, D. Lost value assessment of bathing beaches due to sea level rise: A case study of the Mediterranean coast of Israel. J. Coast. Conserv. 2019, 23, 773–783. [Google Scholar] [CrossRef]
- Bitan, M.; Zviely, D. Sand beach nourishment: Experience from the Mediterranean coast of Israel. J. Mar. Sci. Eng. 2020, 8, 273. [Google Scholar] [CrossRef]
- Nir, Y. Offshore artificial structures and their influence on the Israel and Sinai Mediterranean beaches. In Proceedings of the 18th International Conference on Coastal Engineering; ASCE: Cape Town, South Africa, 1982; pp. 1837–1856. [Google Scholar]
- Inman, D.L.; Jenkins, S.A. The Nile littoral cell and man’s impact on the coastal zone of the Southeastern Mediterranean. In Proceedings of the 19th International Conference on Coastal Engineering; ASCE: Houston, TX, USA, 1984; pp. 1600–1617. [Google Scholar]
- Stanley, D.J. Sediment transport on the coast and shelf between the Nile Delta and Israeli margin as determined by heavy minerals. J. Coast. Res. 1989, 5, 813–828. [Google Scholar]
- Frihy, O.E.; Fanos, A.M.; Khafagy, A.A.; Komar, P.D. Patterns of sediment transport along the Nile Delta, Egypt. Coast. Eng. 1991, 15, 409–429. [Google Scholar] [CrossRef]
- Sharaf El Din, S.H.; Mahar, A.M. Evaluation of sediment transport along the Nile Delta coast, Egypt. J. Coast. Res. 1997, 13, 23–26. [Google Scholar]
- Frihy, O.E.; Deabes, E.A.; El Gindy, A.A. Wave climate and nearshore processes on the Mediterranean coast of Egypt. J. Coast. Res. 2010, 26, 103–112. [Google Scholar] [CrossRef]
- Khalifa, M.A. Adoption of recent formulae for sediment transport calculations applied on the Egyptian Nile delta coastal area. J. Coast. Conserv. 2012, 16, 37–49. [Google Scholar] [CrossRef]
- Emery, K.O.; Neev, D. Mediterranean beaches of Israel. Israel Geol. Surv. Bull. 1960, 26, 1–24. [Google Scholar]
- Goldsmith, V.; Golik, A. Sediment transport model of the southeastern Mediterranean coast. Mar. Geol. 1980, 37, 135–147. [Google Scholar] [CrossRef]
- Rohrlich, V.; Goldsmith, V. Sediment transport along the southeast Mediterranean: A geological perspective. Geo-Mar. Lett. 1984, 4, 99–103. [Google Scholar] [CrossRef]
- Carmel, Z.; Inman, D.; Golik, A. Directional wave measurements at Haifa, Israel, and sediment transport along the Nile littoral cell. Coast. Eng. 1985, 9, 21–36. [Google Scholar] [CrossRef]
- Perlin, A.; Kit, E. Longshore sediment transport on the Mediterranean coast of Israel. J. Waterw. Port. Coast. Ocean. Eng. 1999, 125, 80–87. [Google Scholar] [CrossRef]
- Zviely, D.; Kit, E.; Klein, M. Longshore sand transport estimates along the Mediterranean coast of Israel in the Holocene. Mar. Geol. 2007, 237, 61–73. [Google Scholar] [CrossRef]
- Kit, E.; Sladkevich, M. Structure of offshore currents on sediment Mediterranean coast of Israel. In Proceedings of the 6th Workshop on Physical Processes in Natural Waters, Girona, Spain, 27–29 June 2001; Casamitjana, X., Ed.; 2001; pp. 97–100. [Google Scholar]
- Kunitsa, D.; Rosentraub, D.; Stiassnie, M. Estimates of winter currents on the Israeli continental shelf. Coast. Eng. 2005, 52, 93–102. [Google Scholar] [CrossRef]
- Zviely, D. Sedimentological Processes in Haifa Bay in Context of the Nile Littoral Cell. Ph.D. Thesis, Department of Geography and Environment Studies, University of Haifa, Haifa, Israel, 2006. (In Hebrew, English abstract).. [Google Scholar]
- Zviely, D.; Sivan, D.; Ecker, A.; Bakler, N.; Rohrlich, V.; Galili, E.; Boaretto, E.; Klein, M.; Kit, E. The Holocene evolution of Haifa Bay area, Israel, and its influence on the ancient human settlements. Holocene 2006, 16, 849–861. [Google Scholar] [CrossRef]
- Pomerancblum, M. The distribution of heavy minerals and their hydraulic equivalents in sediments of the Mediterranean shelf of Israel. J. Sedim. Petrol. 1966, 36, 162–174. [Google Scholar] [CrossRef]
- Almagor, G.; Gill, D.; Perath, I. Marine sand resources offshore Israel. Mar. Georesources Geotech. 2000, 18, 1–42. [Google Scholar] [CrossRef]
- Sandler, A.; Herut, B. Composition of clays along the continental shelf off Israel: Contribution of the Nile versus local sources. Mar. Geol. 2000, 167, 339–354. [Google Scholar] [CrossRef]
- Nir, Y. Recent Sediments of Haifa Bay; Rep. MG/11/80; Geological Survey of Israel: Jerusalem, Israel, 1980; p. 8. [Google Scholar]
- Bakler, N. Gross Lithology of Drilling and Laboratory Data, Haifa Bay, Tel-Aviv and Caesarea; Final summary 1, UNDP-GSI offshore dredging project, ISR/71/682; Geological Survey of Israel: Jerusalem, Israel, 1975; p. 23. [Google Scholar]
- Hall, J.K. Seismic Studies, Haifa Bay, Summary Report; UNDP-GSI offshore dredging project ISR/71/522. Geological Survey of Israel: Jerusalem, Israel, 1976; Volume 1/76, p. 36. [Google Scholar]
- Zviely, D.; Kit, E.; Rosen, B.; Galili, E.; Klein, M. Shoreline migration and beach-nearshore sand balance over the last 200 years in Haifa Bay (SE Mediterranean). Geo Mar. Lett. 2009, 29, 93–110. [Google Scholar] [CrossRef]
- Galil, B.S.; Froglia, C.; Noel, P. Looking back, looking ahead: The CIESM Atlas, Crustaceans. Manag. Biol. Invasions 2015, 6, 171–175. [Google Scholar] [CrossRef]
- Coll, M.; Piroddi, C.; Steenbeek, J.; Kaschner, K.; Lasram, F.B.R.; Aguzzi, J.; Ballesteros, E.; Bianchi, C.N.; Corbera, J.; Dailianis, T.; et al. The biodiversity of the Mediterranean Sea: Estimates, patterns, and threats. PLoS ONE 2010, 5, e11842. [Google Scholar] [CrossRef]
- Gökoğlu, M.; Julian, D.; Dailianis, T.; Akyol, O.; Babali, N.; Bariche, M.; Crocetta, F.; Gerovasileiou, V.; Chanem, R.; Gökoğlu, M.; et al. Occurence of Matuta victor (Crustacea: Decapoda) in Turkey [p.619–620]. In New Mediterranean biodiversity records. Medit. Mar. Sci. 2016, 17, 608–626. [Google Scholar]
- Kondylatos, G.; Corsini-Foka, M.; Perakis, E. First record of the isopod Idotea hectica (Pallas, 1772) (Idoteidae) and of the brachyuran crab Matuta victor (Fabricius, 1781) (Matutidae) in the Hellenic waters. Medit. Mar. Sci. 2018, 19, 656–661. [Google Scholar] [CrossRef]
- Hanim, N.; Wardiatno, Y.; Perwitasari, D.; Suman, A.; Parlindungan, D.; Farajallah, A. Distribution of Matuta purnama J. C. Y. Lai and Galil, 2007 (Brachyura: Matutidae) outside type locality. IOP Conf. Ser. Earth Environ. Sci. 2021, 744, 012023. [Google Scholar] [CrossRef]
- Bellwood, O. The occurrence, mechanics and significance of burying behavior in crabs (Crustacea: Brachyura). J. Nat. Hist. 2002, 36, 1223–1238. [Google Scholar] [CrossRef]
- Ng, P.K.L. Crabs [pp. 1045–1155]. In FAO Species Identification Guide for Fishery Purposes. The Living Marine Resources of the Western Central Pacific. Volume 2: Cephalopods, Crustaceans, Holothurians and Sharks; Carpenter, K.E., Niem, V.H., Eds.; Food and Agriculture Organization of the United Nations: Rome, Italy, 1998; pp. 687–1396. [Google Scholar]
- Galil, B.S.; Mendelson, M.A. Record of the moon crab Matuta victor (Fabricius, 1781) (Crustacea; Decapoda; Matutidae) from the Mediterranean coast of Israel. BioInv. Rec. 2013, 2, 69–71. [Google Scholar] [CrossRef]
- Crocetta, F.; Bariche, M.; Crocetta, F.; Agius, D.; Balistreri, P.; Bariche, M.; Bayhan, Y.; Çakir, M.; Ciriaco, S.; Corsini-Foka, M.; et al. Six new records from Lebanon, with general implications for Mediterranean alien fauna [p. 696–698]. In New Mediterranean biodiversity records. Medit. Mar. Sci. 2015, 16, 682–702. [Google Scholar] [CrossRef]
- Ateş, S.; Katağan, T.; Sert, M.; Özdilek, S.Y. A new locality for common box crab, Matuta victor (Fabricius, 1781), from the eastern Mediterranean Sea. J. Black Sea Medit. Env. 2017, 23, 191–195. [Google Scholar]
- Crain, D.A.; Boltenu, A.B.; Bjorndal, K.A. Effects of beach nourishment on sea turtles: Review and research initiatives. Rest. Ecol. 1995, 3, 95–104. [Google Scholar]
- Lubinevsky, H. Ecological, Genetic and Morphological Aspects of Migrant Gastropod Strombus (Conomurex) Persicus. Ph.D. Thesis, Department of Geography and Environment Studies, University of Haifa, Haifa, Israel, 2011. (In Hebrew, English abstract). [Google Scholar]
- Çinar, M.E.; Bilecenoğlu, M.; Yokeş, M.B.; Öztürk, B.; Taşkin, E.; Bakir, K.; Doğan, A.; Açik, Ş. Current status (as of end of 2020) of marine alien species in Turkey. PLoS ONE 2021, 16, e0251086. [Google Scholar] [CrossRef] [PubMed]
- Parreira, F.; Martínez-Crego, B.; Afonso, C.M.L.; Machado, M.; Oliveira, F.; dos Santos Gonçalves, J.M.; Santos, R. Biodiversity consequences of Caulerpa prolifera takeover of a coastal lagoon. Estuar. Coast. Shelf Sci. 2021, 107344, 1–7. [Google Scholar]
- Galil, B.S.; Golani, D. Two new migrant decapods from the Eastern Mediterranean. Crustaceana 1990, 58, 229–236. [Google Scholar]
- Daneshgar, P.P.; Phillips, L.B.; James, D.P.; Mickley, M.G.; Bohackyj, A.M.; Rhoads, L.J.; Bastian, R.B.; Wootton, L.S. The role of beach nourishment on the success of invasive Asiatic Sand Sedge. Northeast. Nat. 2017, 24, 110–120. [Google Scholar] [CrossRef]
- Karachle, P.K.; Corsini Foka, M.; Crocetta, F.; Dulčić, J.; Dzhembekova, N. Setting-up a billboard of marine invasive species in the ESENIAS area: Current situation and future expectancies. Acta Adriatica 2017, 58, 429–458. [Google Scholar] [CrossRef][Green Version]
- Kleitou, P.; Doumpas, N. Cyprus: The alien moon crab Matuta victor is reported for the first time from the entire country (First record of the moon crab Matuta victor (Fabricius, 1781) (Crustacea; Decapoda; Matutidae) from Cyprus Periklis and Nikolaos Doumpas. Medit. Mar. Sci. 2019, 20, 244. [Google Scholar]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zviely, D.; Zurel, D.; Edelist, D.; Bitan, M.; Spanier, E. Does Sand Beach Nourishment Enhance the Dispersion of Non-Indigenous Species?—The Case of the Common Moon Crab, Matuta victor (Fabricius, 1781), in the Southeastern Mediterranean. J. Mar. Sci. Eng. 2021, 9, 911. https://doi.org/10.3390/jmse9080911
Zviely D, Zurel D, Edelist D, Bitan M, Spanier E. Does Sand Beach Nourishment Enhance the Dispersion of Non-Indigenous Species?—The Case of the Common Moon Crab, Matuta victor (Fabricius, 1781), in the Southeastern Mediterranean. Journal of Marine Science and Engineering. 2021; 9(8):911. https://doi.org/10.3390/jmse9080911
Chicago/Turabian StyleZviely, Dov, Dror Zurel, Dor Edelist, Menashe Bitan, and Ehud Spanier. 2021. "Does Sand Beach Nourishment Enhance the Dispersion of Non-Indigenous Species?—The Case of the Common Moon Crab, Matuta victor (Fabricius, 1781), in the Southeastern Mediterranean" Journal of Marine Science and Engineering 9, no. 8: 911. https://doi.org/10.3390/jmse9080911
APA StyleZviely, D., Zurel, D., Edelist, D., Bitan, M., & Spanier, E. (2021). Does Sand Beach Nourishment Enhance the Dispersion of Non-Indigenous Species?—The Case of the Common Moon Crab, Matuta victor (Fabricius, 1781), in the Southeastern Mediterranean. Journal of Marine Science and Engineering, 9(8), 911. https://doi.org/10.3390/jmse9080911