
Histologic Examination of a Sea Pig (Scotoplanes sp.) Using Bright Field Light Microscopy

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
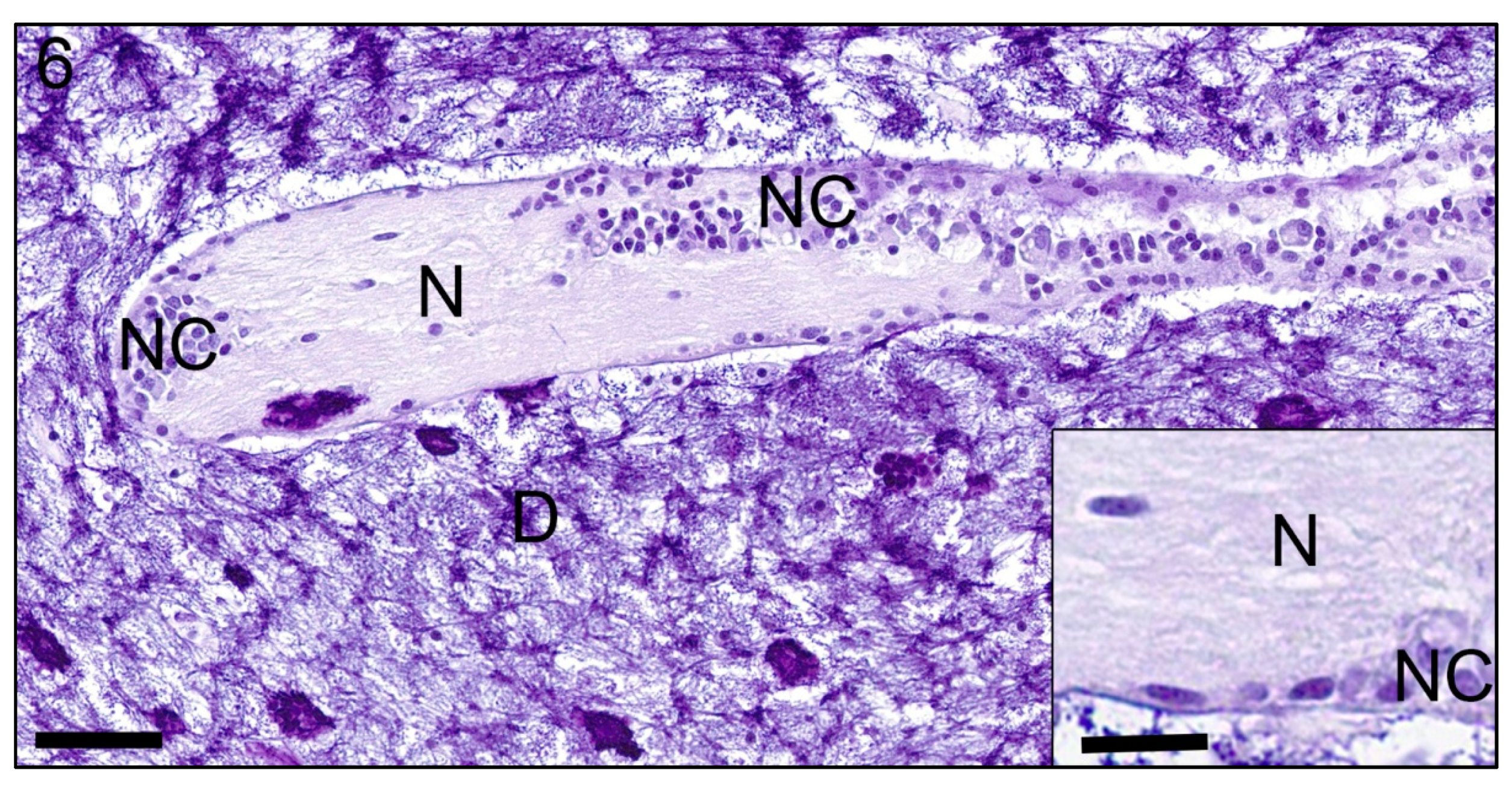{kind=link}
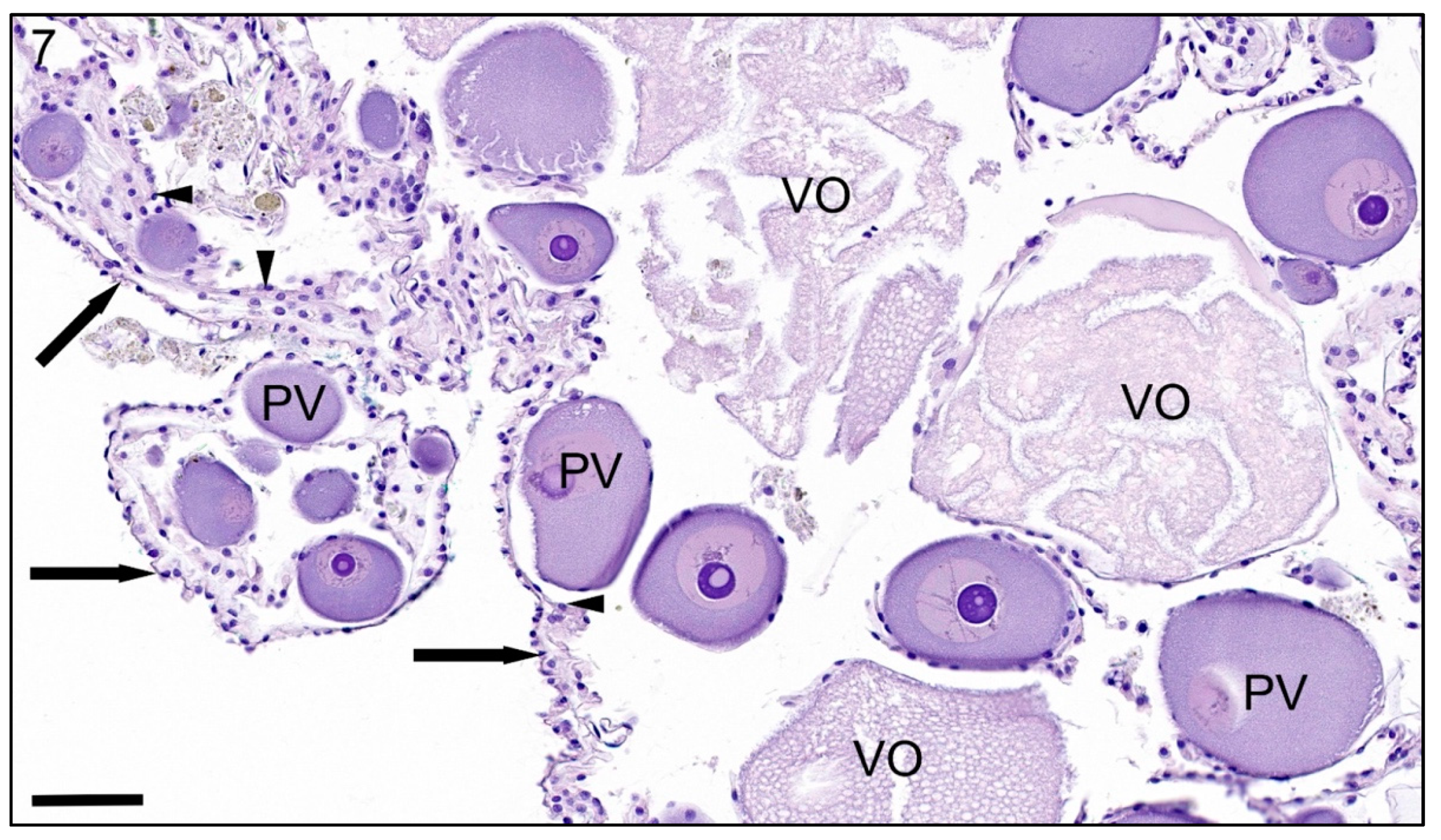{kind=link}
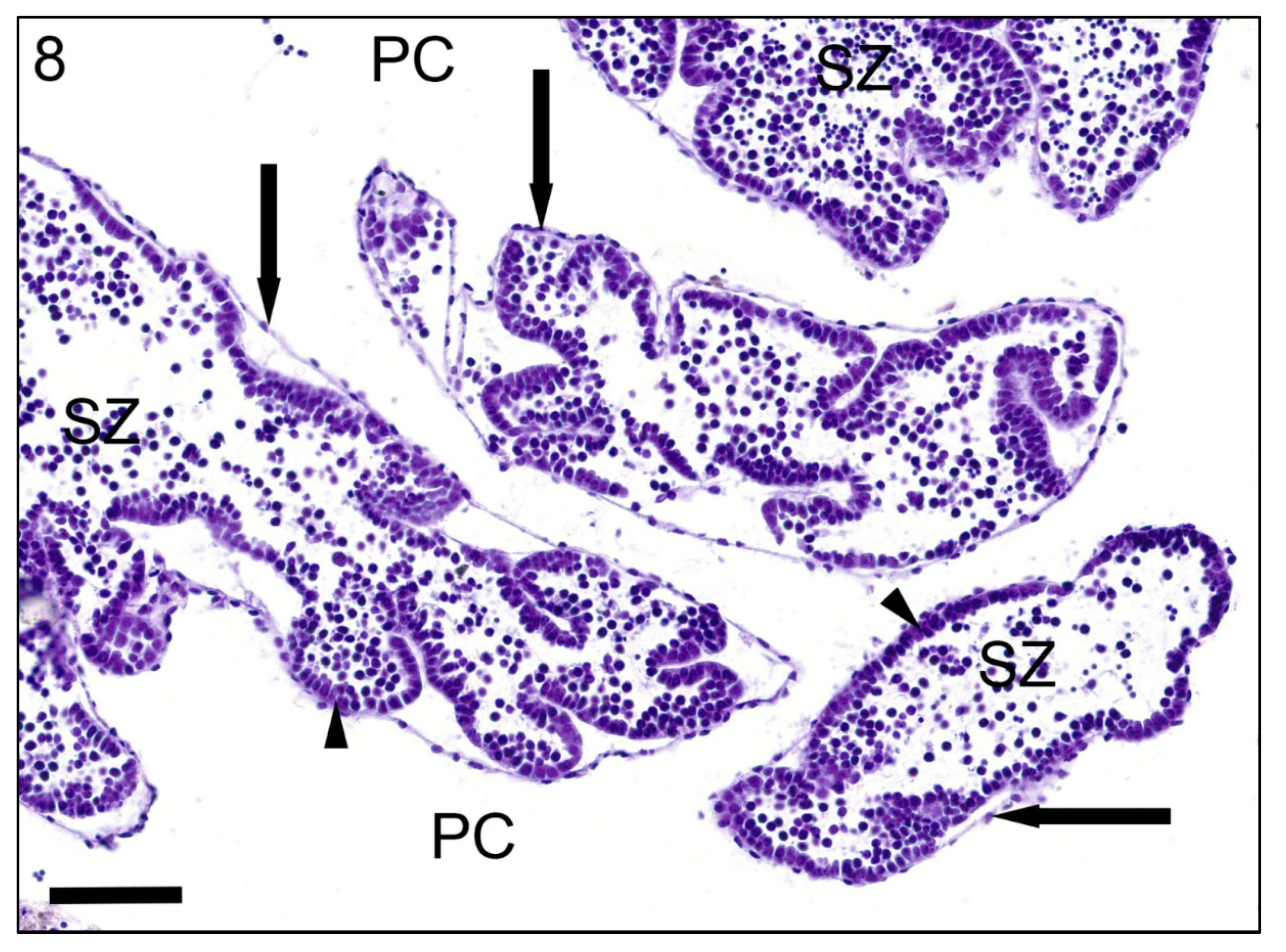{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
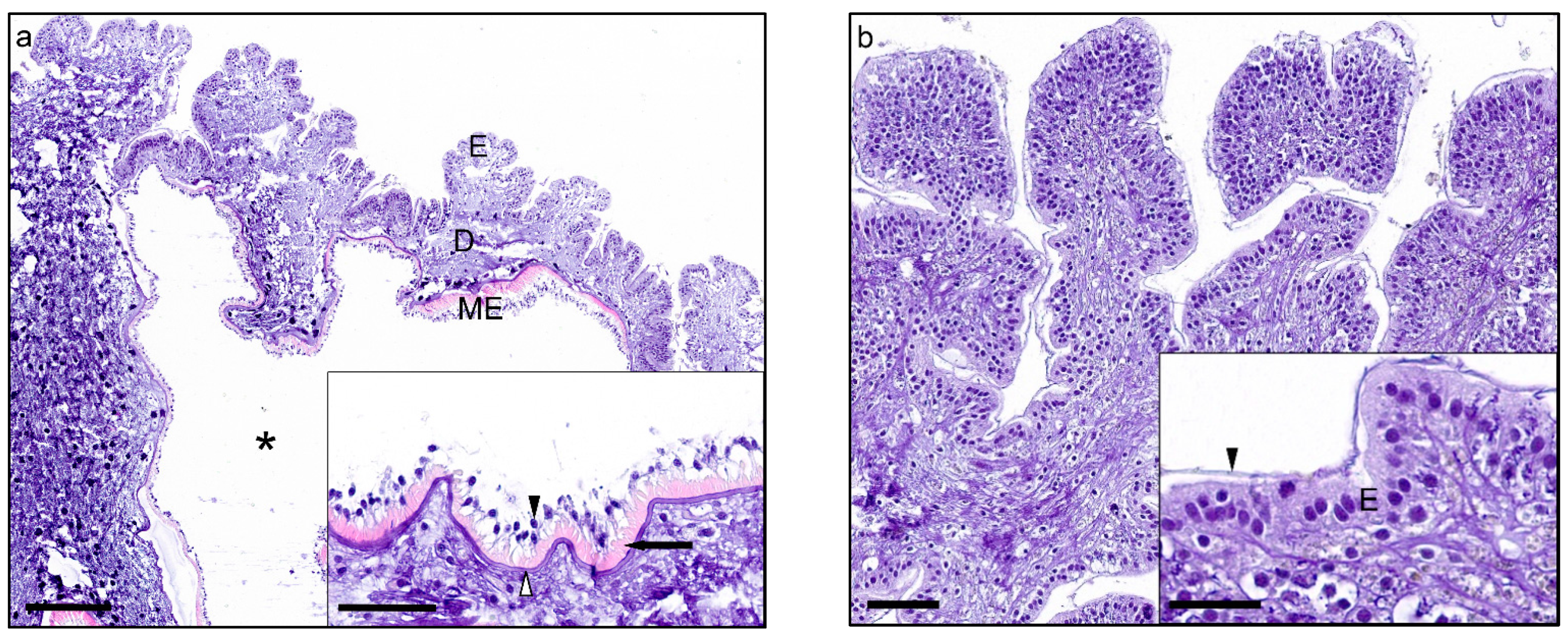
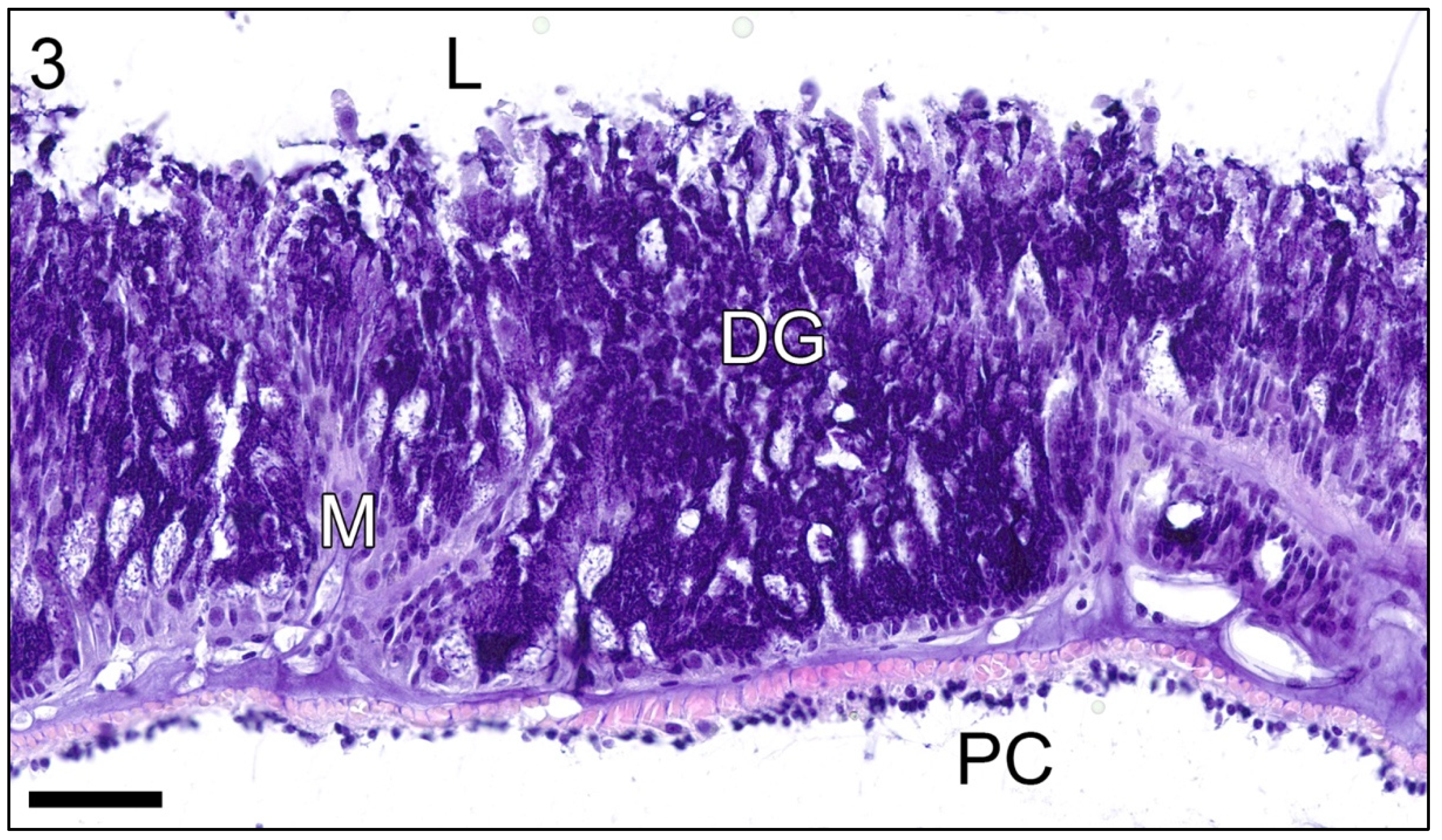
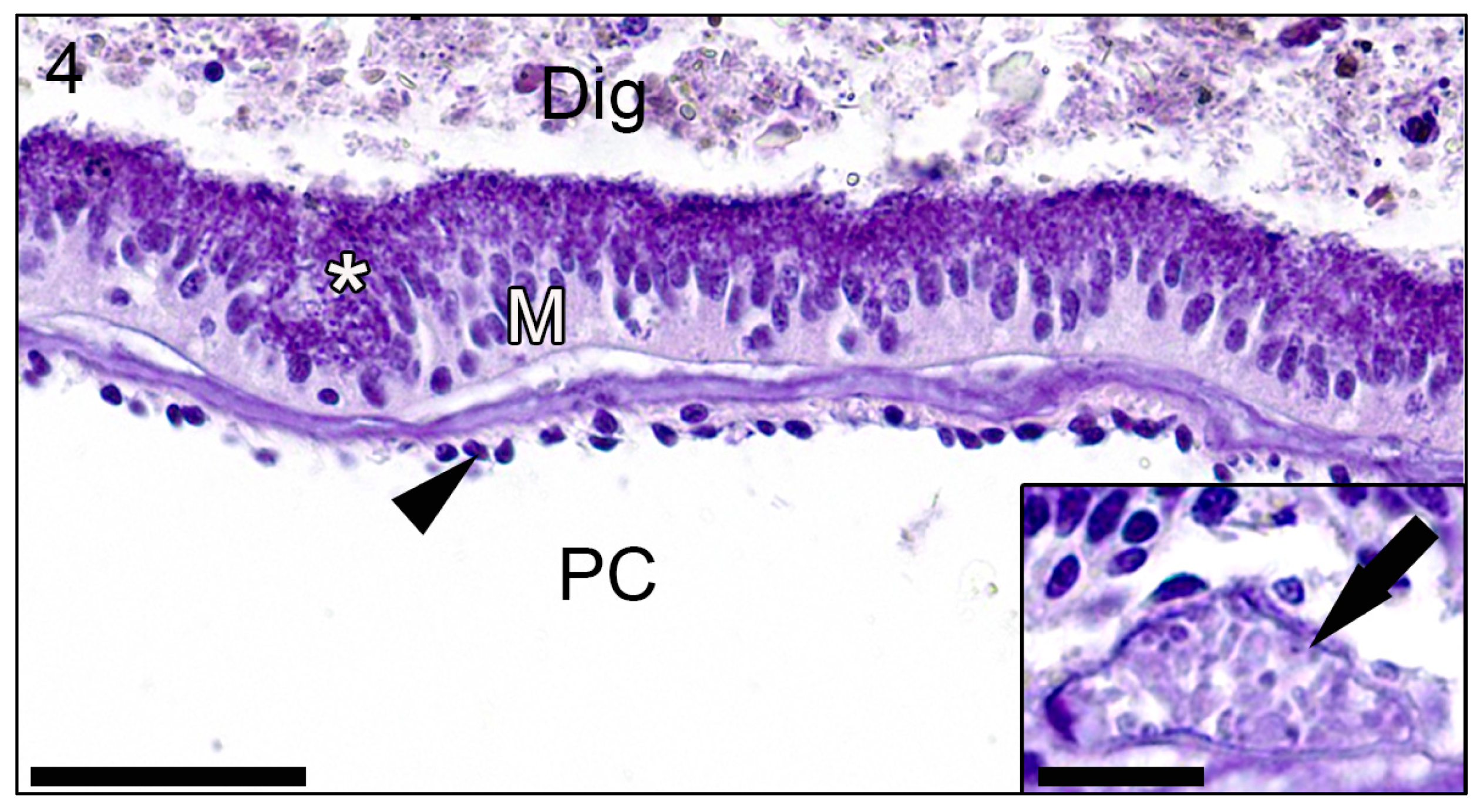
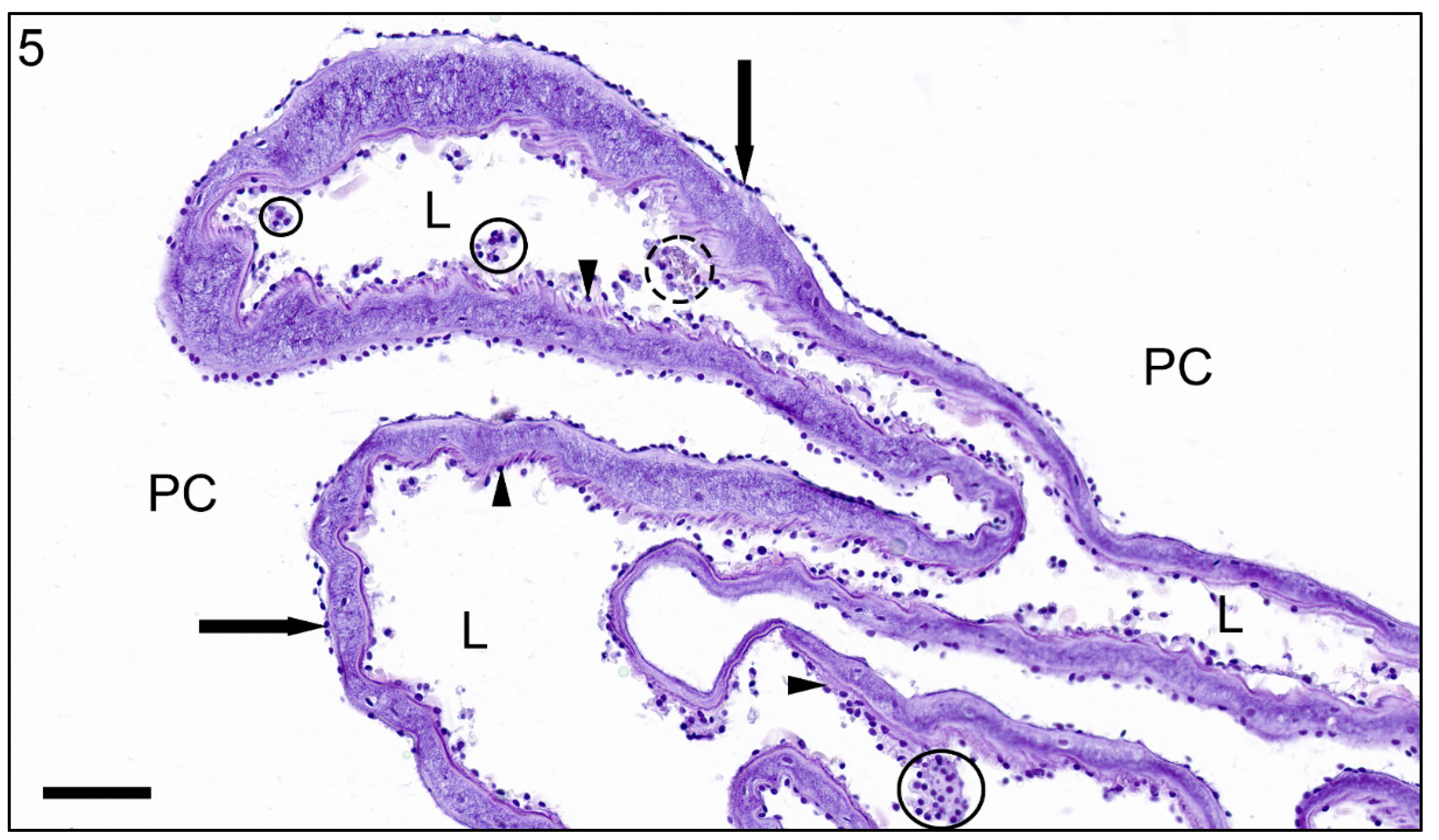
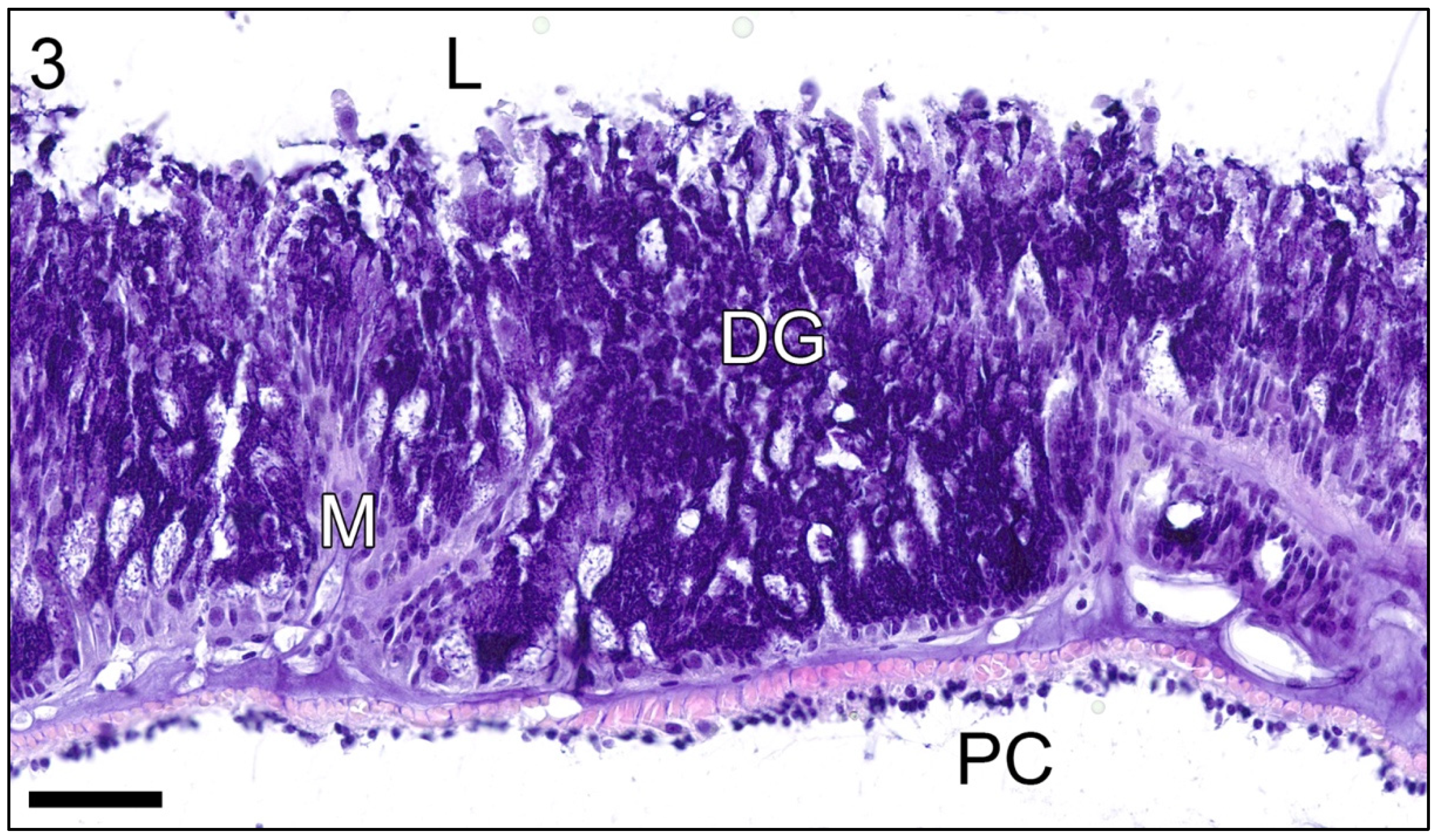
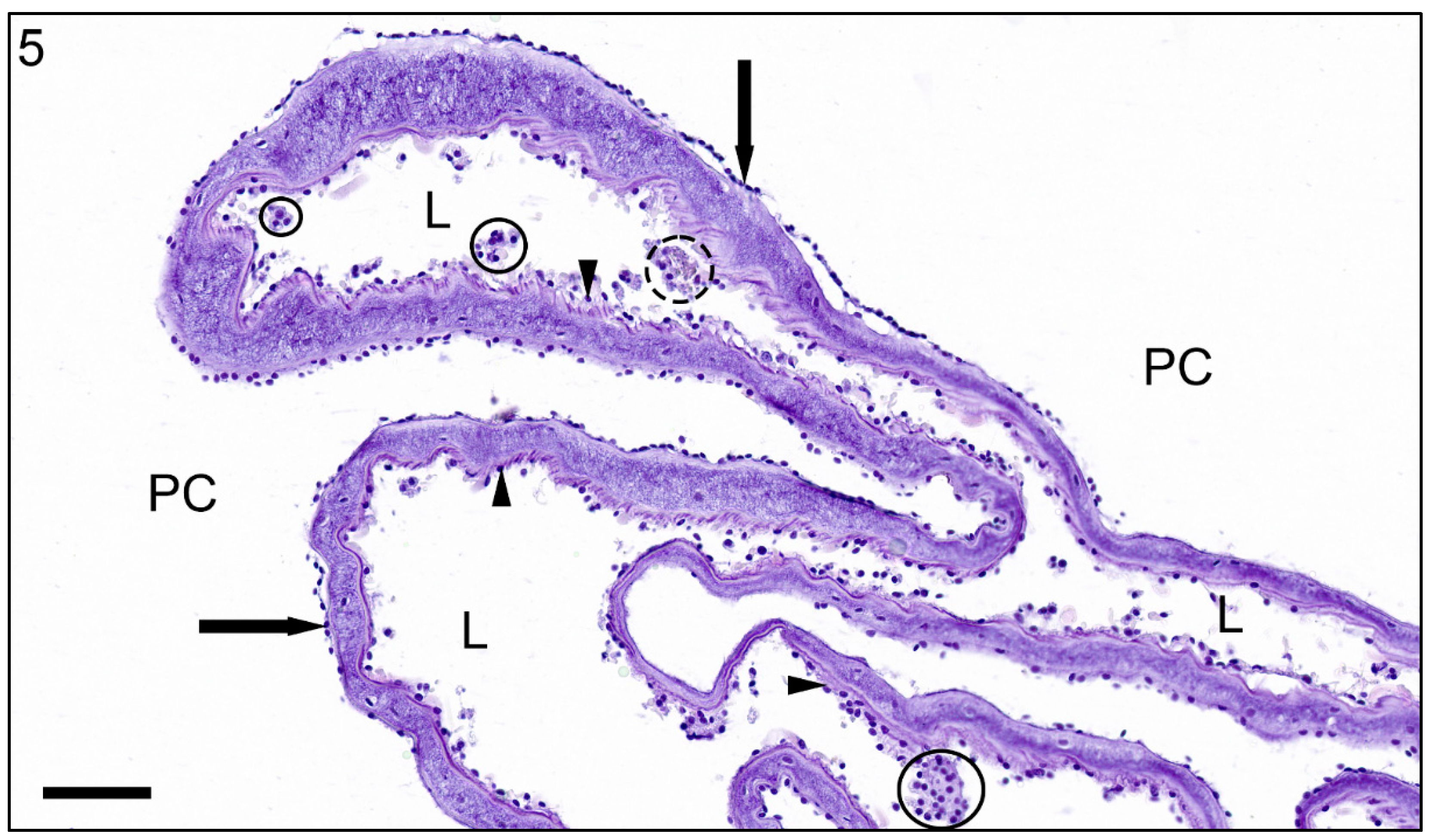
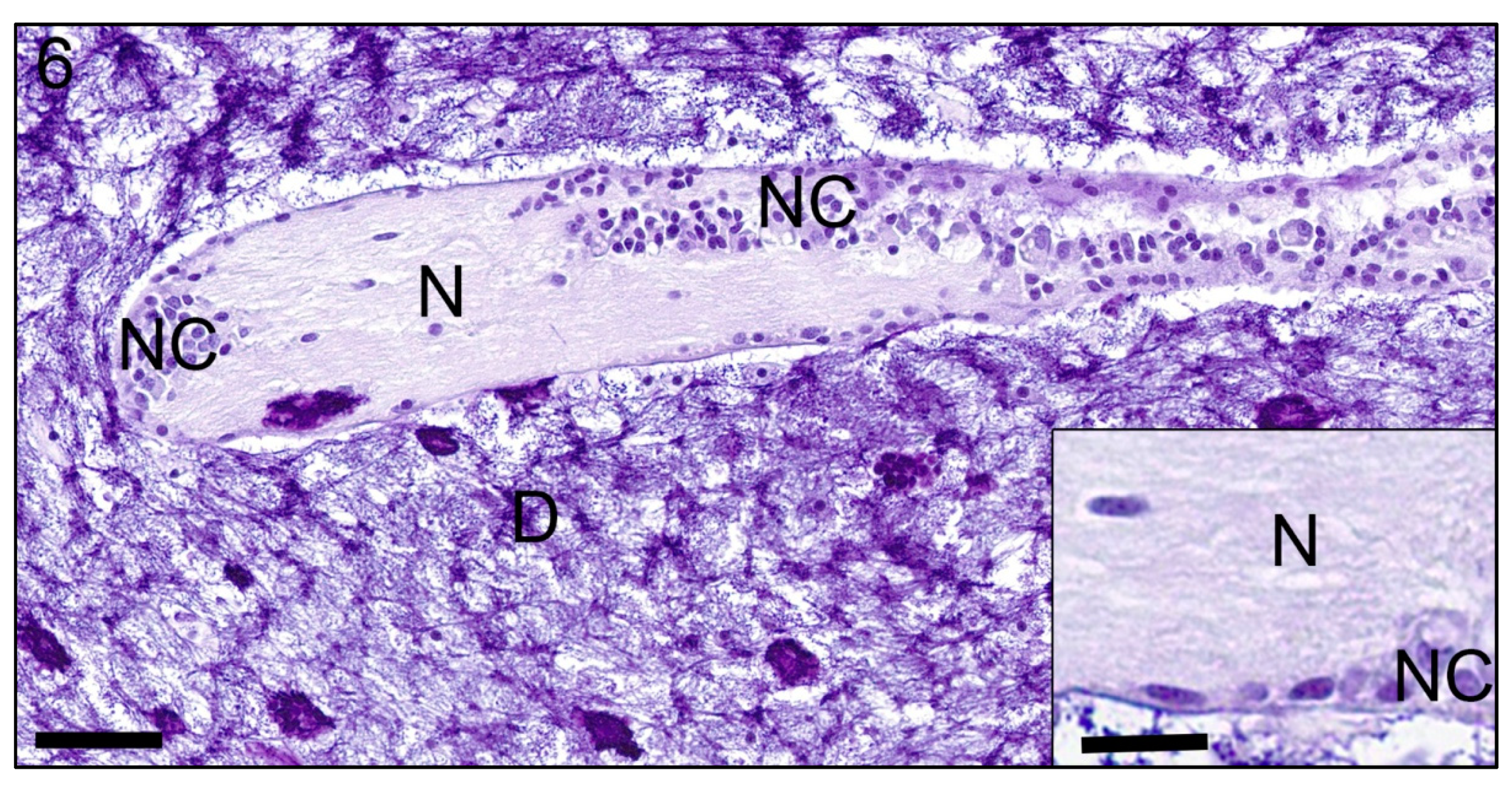
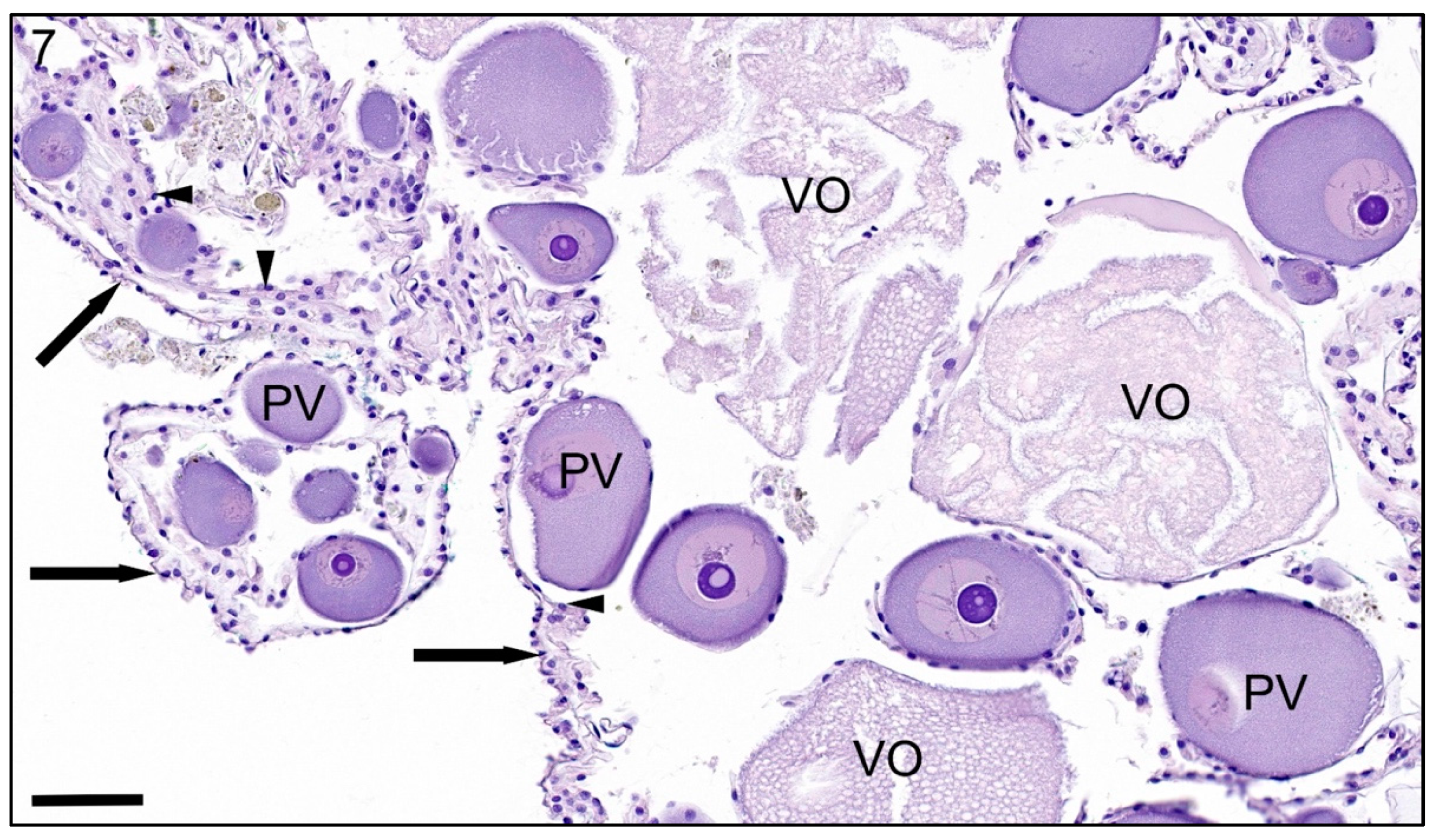
3. Results
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smirnov, A.V. Sea cucumbers symmetry (Echinodermata: Holothuroidea). Paleontol. J. 2014, 48, 1215–1236. [Google Scholar] [CrossRef]
- Ruppert, E.E.; Fox, R.S.; Barnes, R.D. Echinodermata. In Invertebrate Zoology, 7th ed.; Ruppert, E.E., Fox, R.S., Barnes, R.D., Eds.; Brooks/Cole: Pacific Grove, CA, USA, 2004; pp. 872–929. [Google Scholar]
- Théel, H. Report on Holothurioidea. Pt. I. Report of the Scientific Results of the Voyage of H.M.S. Challenger. Zoology 1882, 4, 1–176. [Google Scholar]
- Ramirez-Llodra, E.; Brandt, A.; Danovaro, R.; De Mol, B.; Escobar, E.; German, C.R.; Levin, L.A.; Martinez Arbizu, P.; Menot, L.; Buhl-Mortensen, P.; et al. Deep diverse and definitely different: Unique attributes of the world’s largest ecosystem. Biogeosciences 2010, 7, 2851–2899. [Google Scholar] [CrossRef] [Green Version]
- Woolley, S.N.; Tittensor, D.P.; Dunstan, P.K.; Guillera-Arroita, G.; Lahoz-Monfort, J.J.; Wintle, B.A. Deep-sea diversity patterns are shaped by energy availability. Nature 2016, 533, 393–396. [Google Scholar] [CrossRef] [PubMed]
- Roberts, D.; Gebruk, A.; Levin, V.; Manship, B.A.D. Feeding and digestive strategies in deposit-feeding holothurians. In Oceanography and Marine Biology; Barnes, M., Gibson, R.N., Eds.; Taylor and Francis: Oxford, UK, 2000; pp. 257–310. [Google Scholar]
- Barry, J.P.; Taylor, J.R.; Kuhnz, L.A.; De Vogelaere, A.P. Symbiosis between the holothurian Scotoplanes sp. and the lithodid crab Neolithodes diomedeae on a featureless bathyal sediment plain. Mar. Ecol. 2016, 38, 1–10. [Google Scholar]
- Jacobsen Stout, N.; Kuhnz, L.; Lundsten, B.; Schlining, K.; Schlining, B.; Von Thun, S. The Deep-Sea Guide (DSG); Monterey Bay Aquarium Research Institute: Monterey, CY, USA, 2021. [Google Scholar]
- Diaz-Balzac, C.A.; Abrea-Arbelo, J.E.; Garcia-Arraras, J.E. Neuroanatomy of the tube feet and tentacles in Holothuria glaberrima (Holothuroidea, Echinodermata). Zoomorphology 2010, 129, 33–43. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, U. Ultrastructue of the statocysts in the apodus sea cucumber Leptosynapta inhaerens. (Holothuroidea, Echinodermata). Acta Zool. 1997, 78, 61–68. [Google Scholar] [CrossRef]
- Ezhova, O.V.; Ershova, N.A.; Malakhov, V.V. Microscopic anatomy of the axial complex and associated structures in the sea cucumber Chiridota laevis Frabricius, 1780 (Echinodermata, Holothuroidea). Zoomorphology 2017, 136, 205–217. [Google Scholar] [CrossRef]
- Frick, J.E.; Ruppert, E.E.; Wourms, J.P. Morphology of the ovotestis of Synaptula hydriformis (Holothuroidea, Apoda): An evolutionary model of oogenesis and the origin of egg polarity in echinoderms. Invertebr. Biol. 1996, 115, 46–66. [Google Scholar] [CrossRef]
- Foster, G.G.; Hodgson, A.N. Feeding, tentacle and gut morphology in five species of southern African intertidal holothuroids (Echinodermata). S. Afr. J. Zool. 1996, 31, 70–79. [Google Scholar] [CrossRef]
- Kamenev, Y.O.; Dolmatov, I.Y.; Frolova, L.T.; Khang, N.A. The morphology of the digestive tract and respiratory organs of the holothurian Cladolabes schmeltzii (Holothuroidea, Dendrochirotida). Tissue Cell 2013, 45, 126–139. [Google Scholar] [CrossRef] [PubMed]
- Kremenetskaia, A.; Ezhova, O.; Drozdov, A.L.; Rybakova, E.; Gebruk, A. On the reproduction of two deep-sea Arctic holothurians, Elpidia heckeri and Kolga hyaline (Holothuroidea: Elpidiidae). Invertebr. Reprod. Dev. 2020, 64, 33–47. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Zueva, O.R.; Heinzeller, T.; Dolmatov, I.Y. Ultrastructure of the circumoral nerve ring and the radial nerve cords in holothurians (Echinodermata). Zoomorphology 2006, 125, 27–38. [Google Scholar] [CrossRef]
- Mashanov, V.S.; Frolova, L.T.; Dolmatov, I.Y. Structure of the digestive tube in the Holothurian Eupentacta fraudatrix (Holothuroidea: Dendrochirota). Russ. J. Mar. Biol. 2004, 30, 314–322. [Google Scholar] [CrossRef]
- Utzeri, V.J.; Ribani, A.; Bovo, S.; Taurisano, V.; Calassanzio, M.; Baldo, D.; Fontanesi, L. Microscopic ossicle analyses and the complete mitochondrial genome sequence of Holothuria (Roweothuria) polii (Echinodermata; Holothuroidea) provide new information to support the phylogenetic positioning of this sea cucumber species. Mar. Genom. 2020, 51, 100735. [Google Scholar] [CrossRef] [PubMed]
- Smiley, S. Holothuroidea. In Microscopic Anatomy of Invertebrates; Harrison, F.W., Chia, F., Eds.; Wiley-Liss: Hoboken, NJ, USA, 1994; Volume 14, pp. 401–471. [Google Scholar]
- Hedgpeth, J.W. Galathea Report. Scientific results of the Danish deep-sea expedition round the world 1950–52. Science 1960, 131, 1521. [Google Scholar] [CrossRef]
- Roberts, D.; Moore, H.M. Tentacular diversity in deep-sea deposit-feeding holothurians: Implications for biodiversity in the deep sea. Biodivers. Conserv. 1997, 6, 1487–1505. [Google Scholar] [CrossRef]
- Newton, A.L.; Dennis, M.M. Echinodermata. In Invertebrate Histology; LaDouceur, E.E.B., Ed.; Wiley: Hoboken, NJ, USA, 2021; pp. 1–18. [Google Scholar]
- Galley, E.A.; Tyler, P.A.; Smith, C.R.; Clarke, A. Reproductive biology of two species of holothurian from the deep-sea order Elasipoda, on the Antarctic continental shelf. Deep Sea Res. II 2008, 55, 2515–2526. [Google Scholar] [CrossRef]
- Hansen, B. Photographic Evidence of a unique type of walking in deep-sea Holothurians. Deep Sea Res. Oceanogr. Abstr. 1972, 19, 461–462. [Google Scholar] [CrossRef]
- Feral, J.P.; Massin, C. Digestive systems: Holothuroidea. In Echinoderm Nutrition; Jangoux, M., Lawrence, J.M., Eds.; CRC Press: Boca Raton, FL, USA, 1982; pp. 191–212. [Google Scholar]
- Billett, D.S.M. The Ecology of Deep Sea Holothurians. Ph.D. Thesis, University of Southhampton, Southampton, UK, 1988. [Google Scholar]
- Massin, C.; Jangoux, M.; Sibuet, M. Description d’ Ixoreis psychropotae nov. gen., nov. sp., coccidie parasite du tube digetif de I’Holothurie abyssal Psychropotes longicauda Theel. Protistologica 1978, 14, 253–259. [Google Scholar]
- Massin, C. Structures digestives d’holothuries Elasipoda (Echinodermata): Benthogone rosea Koehler, 1896 et Oneirophanta mutabilis Theel, 1879. Arch. Biol. 1984, 95, 153–185. [Google Scholar]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
LaDouceur, E.E.B.; Kuhnz, L.A.; Biggs, C.; Bitondo, A.; Olhasso, M.; Scott, K.L.; Murray, M. Histologic Examination of a Sea Pig (Scotoplanes sp.) Using Bright Field Light Microscopy. J. Mar. Sci. Eng. 2021, 9, 848. https://doi.org/10.3390/jmse9080848
LaDouceur EEB, Kuhnz LA, Biggs C, Bitondo A, Olhasso M, Scott KL, Murray M. Histologic Examination of a Sea Pig (Scotoplanes sp.) Using Bright Field Light Microscopy. Journal of Marine Science and Engineering. 2021; 9(8):848. https://doi.org/10.3390/jmse9080848
Chicago/Turabian StyleLaDouceur, Elise E. B., Linda A. Kuhnz, Christina Biggs, Alicia Bitondo, Megan Olhasso, Katherine L. Scott, and Michael Murray. 2021. "Histologic Examination of a Sea Pig (Scotoplanes sp.) Using Bright Field Light Microscopy" Journal of Marine Science and Engineering 9, no. 8: 848. https://doi.org/10.3390/jmse9080848
APA StyleLaDouceur, E. E. B., Kuhnz, L. A., Biggs, C., Bitondo, A., Olhasso, M., Scott, K. L., & Murray, M. (2021). Histologic Examination of a Sea Pig (Scotoplanes sp.) Using Bright Field Light Microscopy. Journal of Marine Science and Engineering, 9(8), 848. https://doi.org/10.3390/jmse9080848