
Topological and Morphological Controls on Morphodynamics of Salt Marsh Interiors



Abstract
1. Introduction
2. Materials and Methods
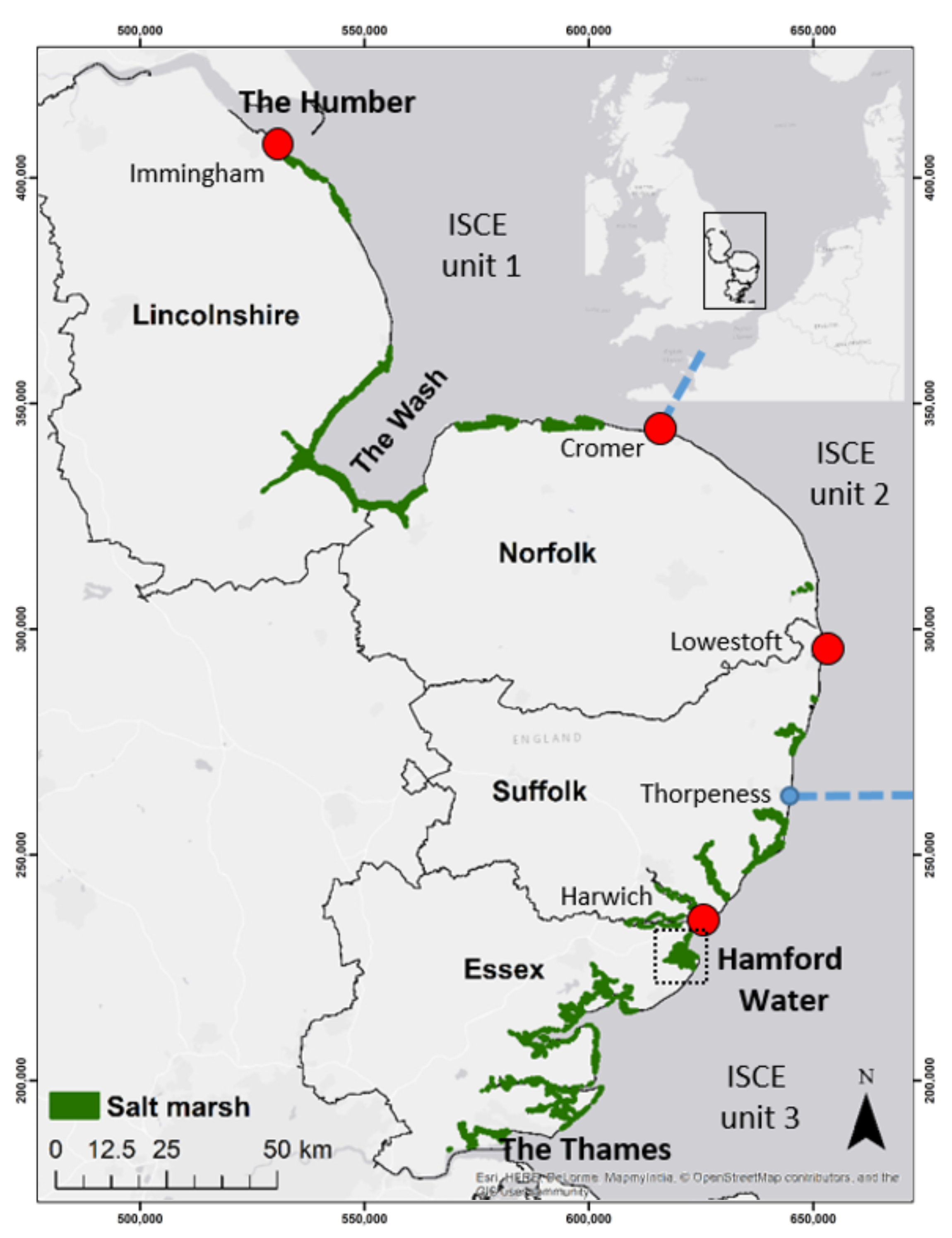
2.1. Study Area
2.2. Morphological Change
2.3. Topological and Morphological Metrics
2.3.1. Geomorphic Setting
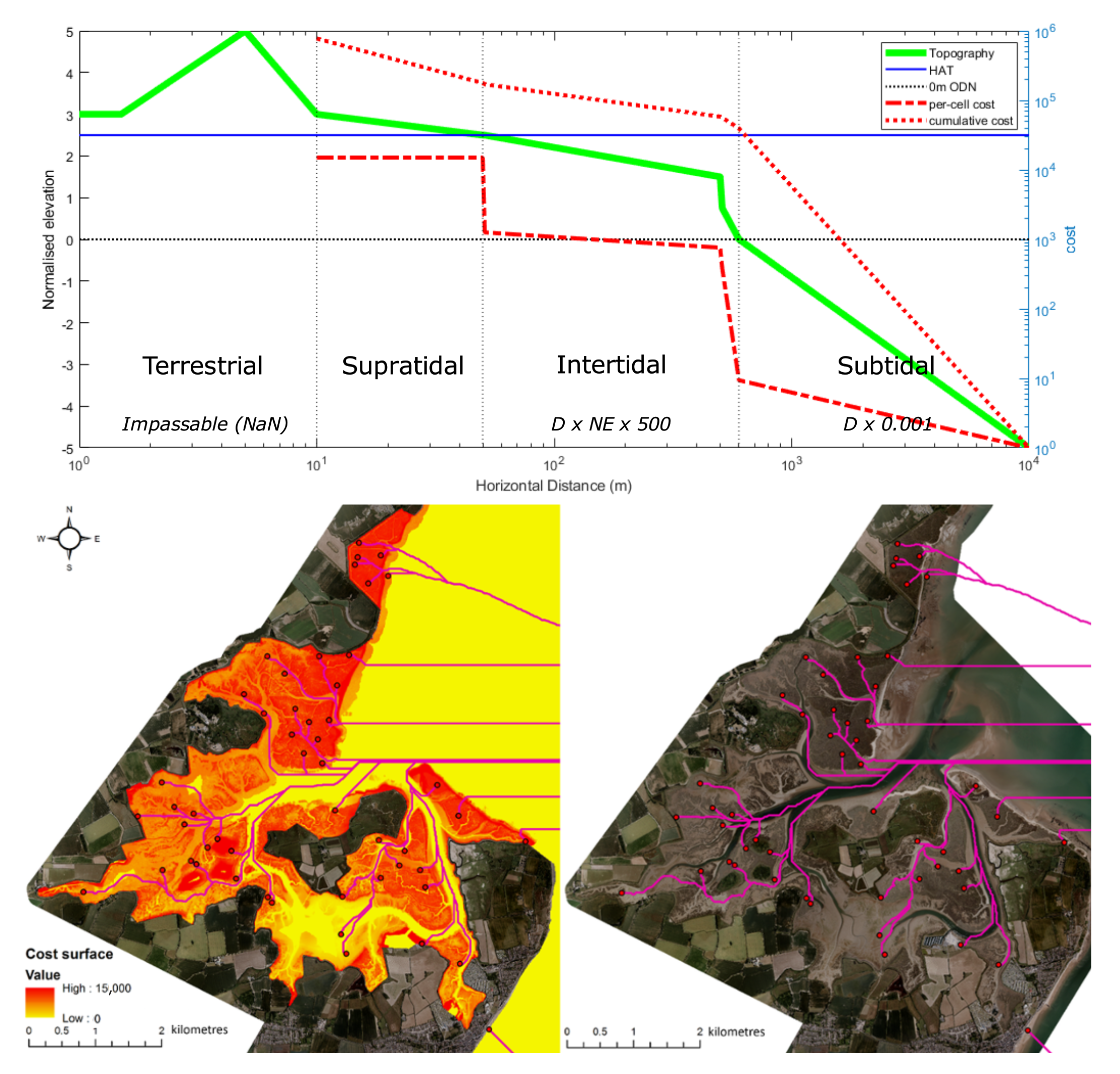
2.3.2. Distance from Creek/Edge of Marsh Parcel
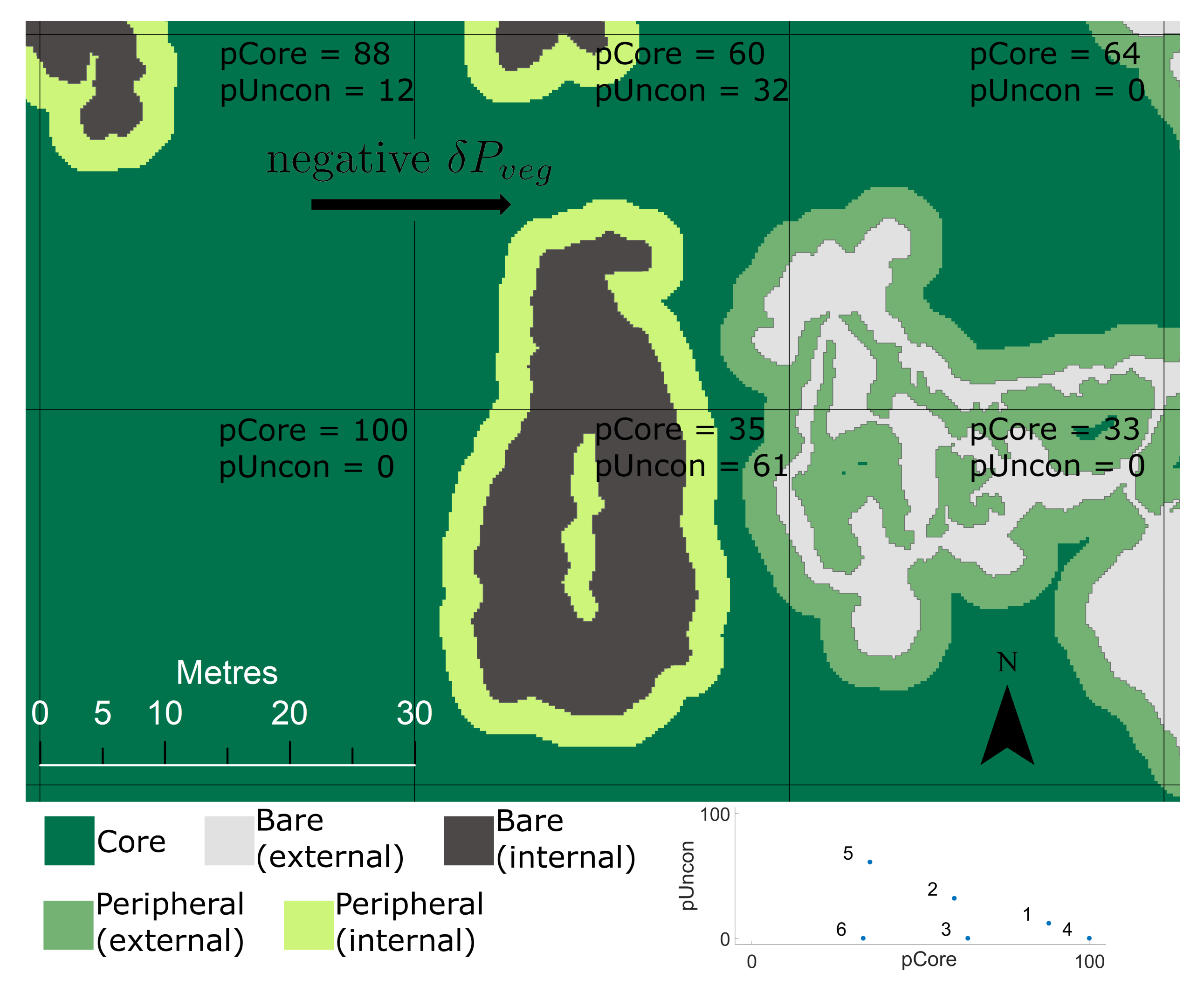
2.3.3. Integrity of Marsh Platform
2.3.4. Limitations to Tidal Connectivity
2.4. Probability of Observing Marsh Degradation
3. Results
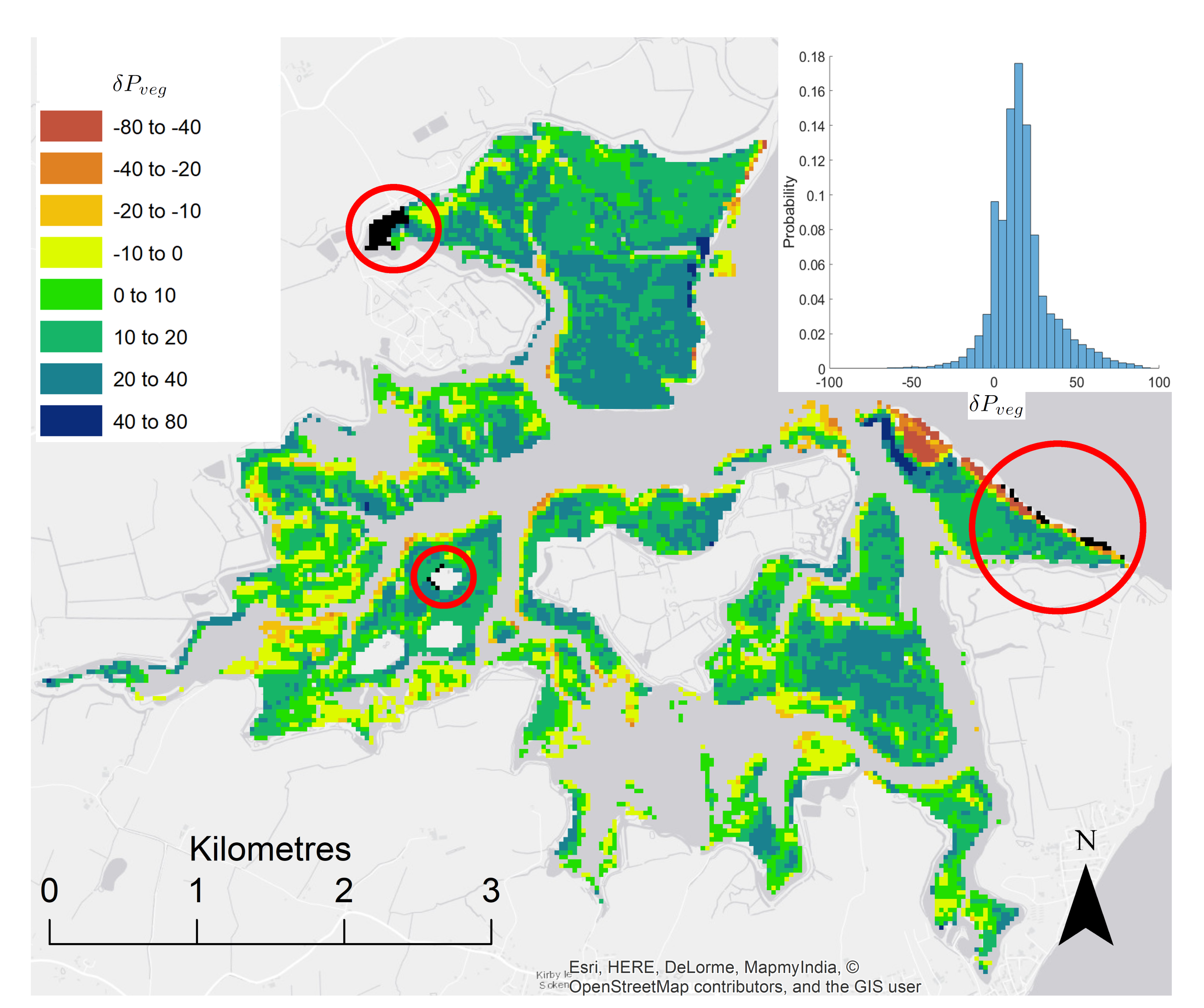
3.1. Morphodynamic Change
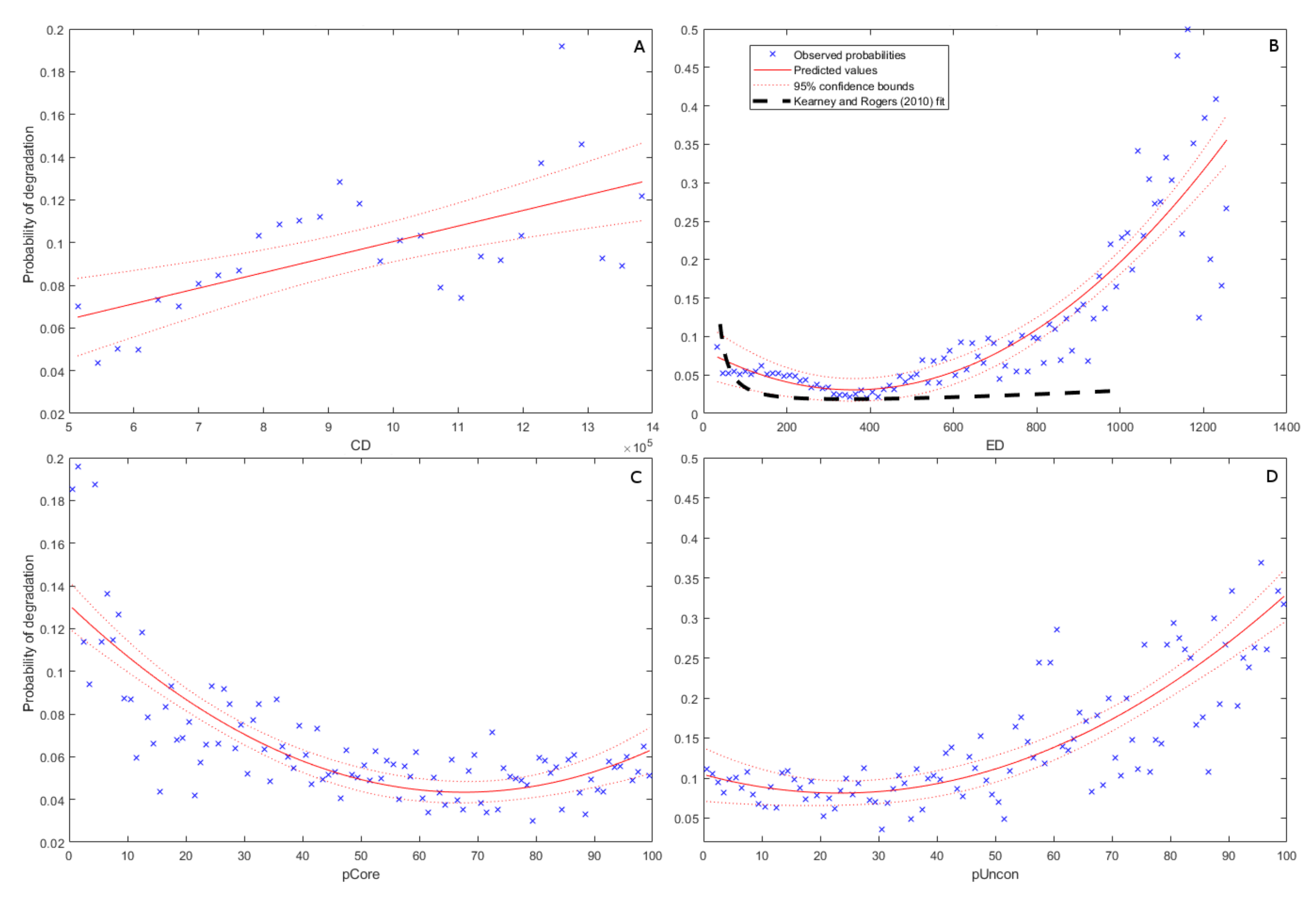
3.2. Topological and Morphological Relationships
4. Discussion
4.1. Morphodynamic Change
4.2. Geomorphic Setting (CD)
4.3. Distance from Creek/Edge of Marsh Parcel (ED)
4.4. Percentage Core Areas (pCore)
4.5. Percentage of Unconnected Areas (pUncon)
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CD | Cost distance |
| ED | Euclidean distance |
| HAT | Highest astronomical tide |
| ISCE | Integrated scale coastal evolution |
| Landsat 5 | |
| Landsat 7 | |
| Landsat 8 | |
| MSCI | Marsh surface condition index |
| MSPA | Morphological spatial pattern analysis |
| MSTR | Mean spring tidal range |
| NDVI | Normalised difference vegetation index |
| NE | Normalised elevation |
| ODN | Ordanace datum Newlyn |
| pCore | Percentage ‘core’ areas |
| pUncon | Percentage ‘unconnected’ areas |
| RMSE | Root-mean-squared error |
| SMP2 | Second-generation shoreline management plan |
| UVVR | Unvegetated–vegetated ratio |
| Change in percentage vegetation cover within pixel |
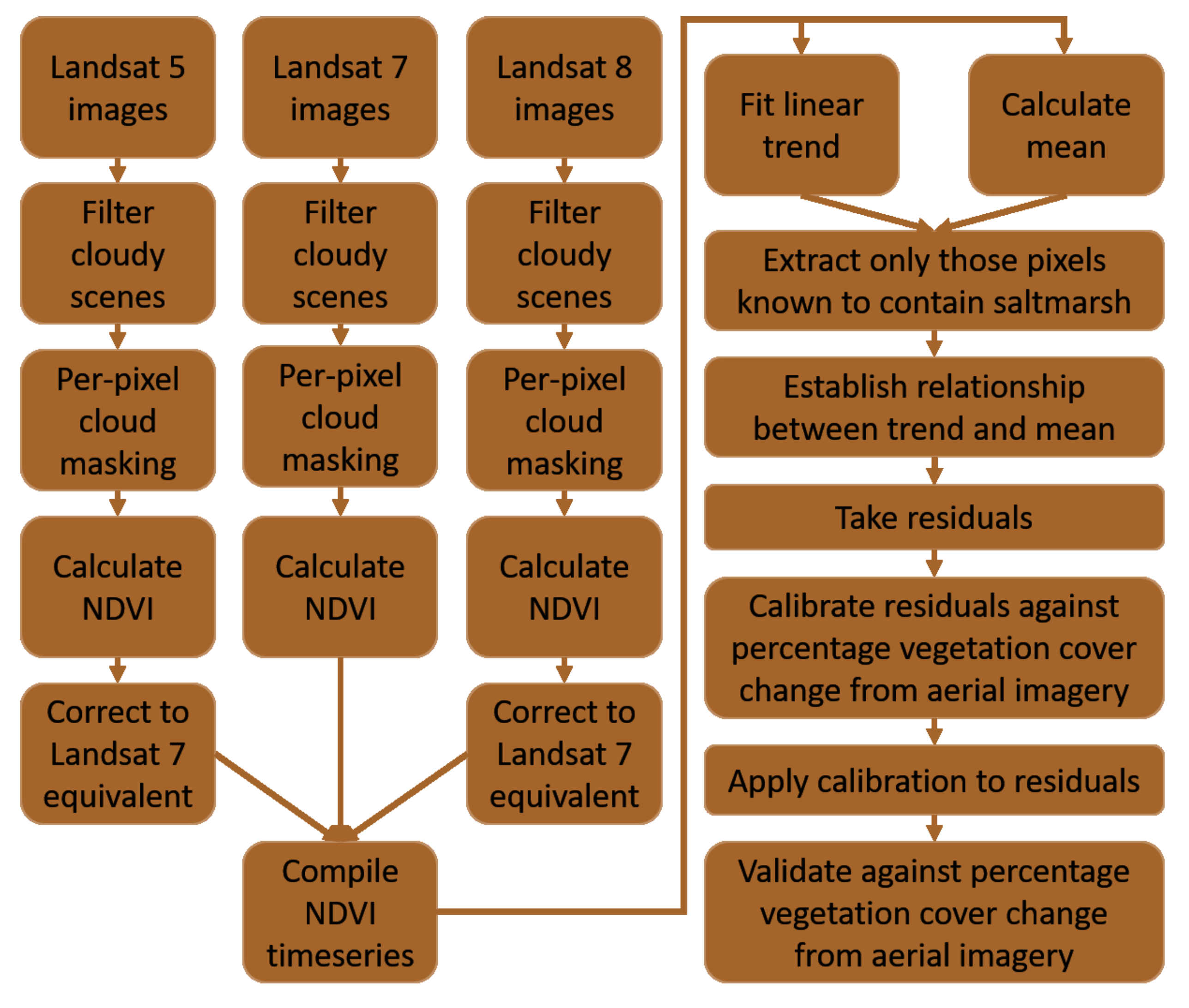
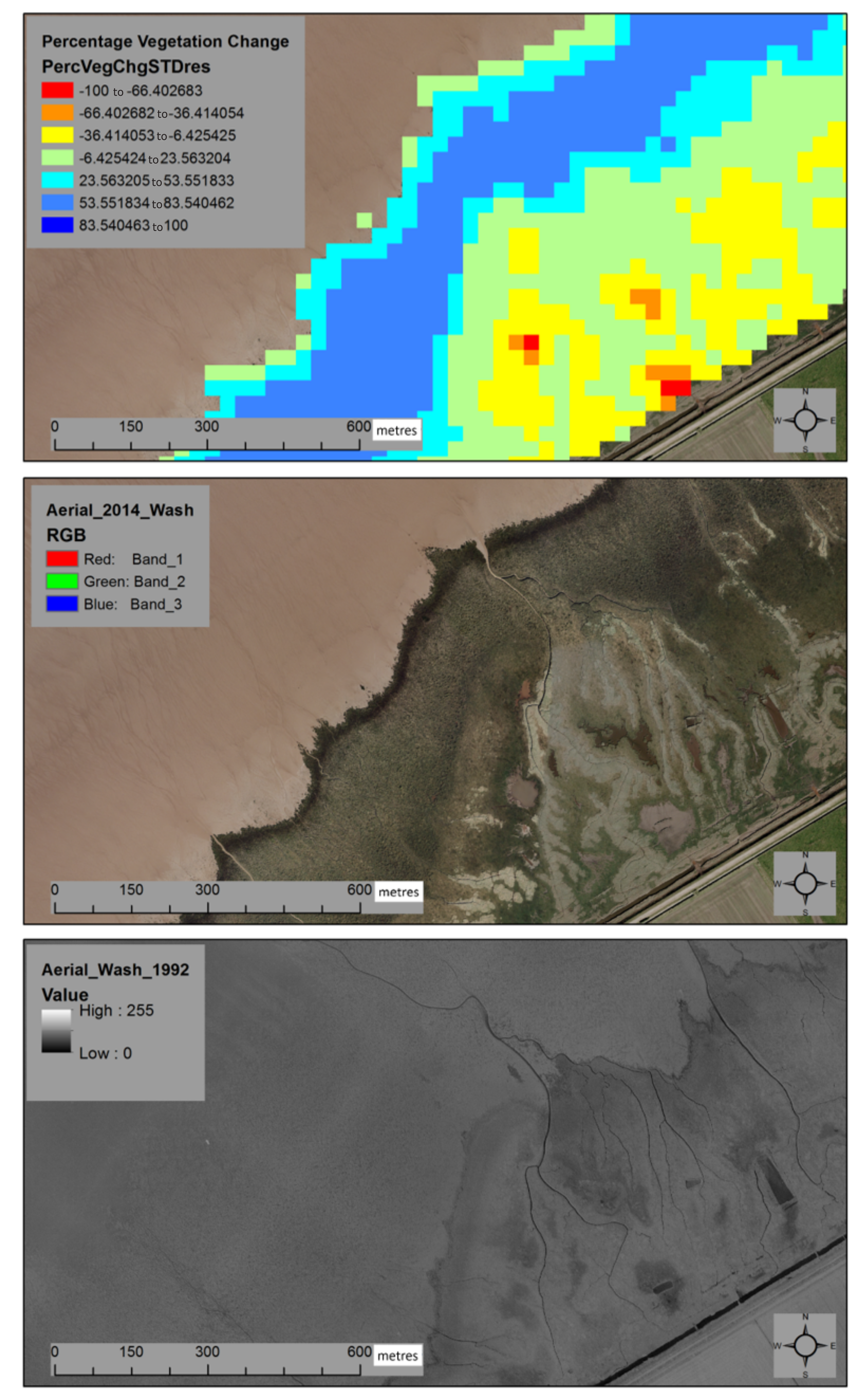
Appendix A. Methodology for Estimation of Marsh Platform Integrity Changes from Satellite Observations

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Transformation | Function | |
|---|---|---|
| to | 0.756 | |
| to | 0.906 |

References
- McOwen, C.; Weatherdon, L.; Bochove, J.W.; Sullivan, E.; Blyth, S.; Zockler, C.; Stanwell-Smith, D.; Kingston, N.; Martin, C.; Spalding, M.; et al. A global map of saltmarshes. Biodivers. Data J. 2017, 5, e11764. [Google Scholar] [CrossRef]
- Friess, D.A.; Yando, E.S.; Alemu I, J.B.; Wong, L.W.; Soto, S.D.; Bhatia, N. Ecosystem Serices and Dissserices of Mangrove Forests and Salt Marshes. In Oceanography and Marine Biology: An Annual Review; CRC Press: Boca Raton, FL, USA, 2020; pp. 58:107–58:142. [Google Scholar]
- Mitsch, W.J.; Gosselink, J.G. Wetlands, 3rd ed.; Wiley: New York, NY, USA, 2000; p. 920. [Google Scholar]
- McLeod, E.; Chmura, G.L.; Bouillon, S.; Salm, R.; Björk, M.; Duarte, C.M.; Lovelock, C.E.; Schlesinger, W.H.; Silliman, B.R. A blueprint for blue carbon: Toward an improved understanding of the role of vegetated coastal habitats in sequestering CO2. Front. Ecol. Environ. 2011, 9, 552–560. [Google Scholar] [CrossRef]
- Möller, I.; Spencer, T.; French, J.R.; Leggett, D.J.; Dixon, M. Wave transformation over salt marshes: A field and numerical modelling study from North Norfolk, England. Estuarine Coast. Shelf Sci. 1999, 49, 411–426. [Google Scholar] [CrossRef]
- Möller, I. Quantifying saltmarsh vegetation and its effect on wave height dissipation: Results from a UK East coast saltmarsh. Estuarine Coast. Shelf Sci. 2006, 69, 337–351. [Google Scholar] [CrossRef]
- Burden, A.; Garbutt, R.; Evans, C.; Jones, D.; Cooper, D. Carbon sequestration and biogeochemical cycling in a saltmarsh subject to coastal managed realignment. Estuarine Coast. Shelf Sci. 2013, 120, 12–20. [Google Scholar] [CrossRef]
- Chmura, G.L. What do we need to assess the sustainability of the tidal salt marsh carbon sink? Ocean. Coast. Manag. 2013, 83, 25–31. [Google Scholar] [CrossRef]
- Roner, M.; Alpaos, A.D.; Ghinassi, M.; Marani, M.; Silvestri, S.; Franceschinis, E. Spatial variation of salt-marsh organic and inorganic deposition and organic carbon accumulation: Inferences from the Venice lagoon, Italy. Adv. Water Resour. 2016, 93, 276–287. [Google Scholar] [CrossRef]
- Boesch, D.F.; Turner, R.E. Dependence of fishery species on salt marshes: The role of food and refuge. Estuaries 1984, 7, 460. [Google Scholar] [CrossRef]
- Costa, M.J.; Costa, J.; de Almeida, P.R.; Assis, C.A. Do eel grass beds and salt marsh borders act as preferential nurseries and spawning grounds for fish? An example of the Mira estuary in Portugal. Ecol. Eng. 1994, 3, 187–195. [Google Scholar] [CrossRef]
- Ladd, C.J.; Duggan-Edwards, M.F.; Bouma, T.J.; Pagès, J.F.; Skov, M.W. Sediment Supply Explains Long-Term and Large-Scale Patterns in Salt Marsh Lateral Expansion and Erosion. Geophys. Res. Lett. 2019, 46, 11178–11187. [Google Scholar] [CrossRef]
- Hughes, R.G.; Paramor, O.A.L. On the loss of saltmarshes in south-east England and. J. Appl. Ecol. 2004, 41, 440–448. [Google Scholar] [CrossRef]
- van der Wal, D.; Pye, K. Patterns, rates and possible causes of saltmarsh erosion in the Greater Thames area (UK). Geomorphology 2004, 61, 373–391. [Google Scholar] [CrossRef]
- Burd, F. Erosion and Vegetation Change on the Salt Marshes of Essex and North Kent between 1973 and 1988; Technical Report; Nature Conservancy Council: Peterborough, UK, 1992. [Google Scholar]
- Cooper, N.J.; Cooper, T.; Burd, F. 25 years of salt marsh erosion in Essex: Implications for coastal defence and nature conservation. J. Coast. Conserv. 2001, 7, 31–40. [Google Scholar] [CrossRef]
- Phelan, N.; Shaw, A.; Baylis, A. The Extent of Saltmarsh in England and Wales: 2006–2009; Environment Agency: Bristol, UK, 2011. [Google Scholar]
- Vandenbruwaene, W.; Meire, P.; Temmerman, S. Formation and evolution of a tidal channel network within a constructed tidal marsh. Geomorphology 2012, 151–152, 114–125. [Google Scholar] [CrossRef]
- French, J.R.; Spencer, T. Dynamics of sedimentation in a tide-dominated backbarrier salt marsh, Norfolk, UK. Mar. Geol. 1993, 110, 315–331. [Google Scholar] [CrossRef]
- Kearney, M.S.; Rogers, A.S. Forecasting sites of future coastal marsh loss using topographical relationships and logistic regression. Wetl. Ecol. Manag. 2010, 18, 449–461. [Google Scholar] [CrossRef]
- Allen, J. Simulation models of salt-marsh morphodynamics: Some implications for high-intertidal sediment couplets related to sea-level change. Sediment. Geol. 1997, 113, 211–223. [Google Scholar] [CrossRef]
- Da Lio, C.; D’Alpaos, A.; Marani, M. The secret gardener: Vegetation and the emergence of biogeomorphic patterns in tidal environments. Philos. Trans. Ser. Math. Phys. Eng. Sci. 2013, 371. [Google Scholar] [CrossRef]
- D’Alpaos, A.; Lanzoni, S.; Marani, M.; Rinaldo, A. Landscape evolution in tidal embayments: Modeling the interplay of erosion, sedimentation, and vegetation dynamics. J. Geophys. Res. Earth Surf. 2007, 112, 1–17. [Google Scholar] [CrossRef]
- Evans, B.R.; Möller, I.; Spencer, T.; Smith, G. Dynamics of salt marsh margins are related to their three-dimensional functional form. Earth Surf. Process. Landforms 2019, 44, esp.4614. [Google Scholar] [CrossRef]
- Pethick, J.; Leggett, D. The morphology of the Anglian coast. In Coastlines of the Southern North Sea; Hillen, R., Verhagen, H., Eds.; American Society of Civil Engineers (ASCE): Reston, VA, USA, 1993; pp. 52–64. [Google Scholar]
- Pye, K.; French, P. Erosion and Accretion Processes on British Saltmarshes: Volume Three; Technical Report; Cambridge Environmental Research Consultants: Cambridge, UK, 1993. [Google Scholar]
- Allen, J. Morphodynamics of Holocene salt marshes: A review sketch from the Atlantic and Southern North Sea coasts of Europe. Quat. Sci. Rev. 2000, 19, 1155–1231. [Google Scholar] [CrossRef]
- Kestner, F. The Old Coastline of the Wash. T Geogr. J. 1962, 128, 457–471. [Google Scholar] [CrossRef]
- Balke, T.; Stock, M.; Jensen, K.; Bouma, T.J.; Kleyer, M. A global analysis of the seaward salt marsh extent: The importance of tidal range. Water Resour. Res. 2016, 52, 3775–3786. [Google Scholar] [CrossRef]
- Suchrow, S.; Jensen, K. Plant species responses to an elevational gradient in German North Sea salt marshes. Wetlands 2010, 30, 735–746. [Google Scholar] [CrossRef]
- Ganju, N.K.; Defne, Z.; Kirwan, M.L.; Fagherazzi, S.; D’Alpaos, A.; Carniello, L. Spatially integrative metrics reveal hidden vulnerability of microtidal salt marshes. Nat. Commun. 2017, 8, 14156. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Lopes, C.L.; Mendes, R.; Caçador, I.; Dias, J.M. Assessing salt marsh extent and condition changes with 35 years of Landsat imagery: Tagus Estuary case study. Remote Sens. Environ. 2020, 247, 111939. [Google Scholar] [CrossRef]
- Hargrove, W.W.; Hoffman, F.M.; Hessburg, P.F. Mapcurves: A quantitative method for comparing categorical maps. J. Geogr. Syst. 2006, 8, 187–208. [Google Scholar] [CrossRef]
- ABPMer. Atlas of Marine Energy Resources. 2008. Available online: http://www.renewables-atlas.info/ (accessed on 5 May 2015).
- French, J. Tidal marsh sedimentation and resilience to environmental change: Exploratory modelling of tidal, sea-level and sediment supply forcing in predominantly allochthonous systems. Mar. Geol. 2006, 235, 119–136. [Google Scholar] [CrossRef]
- French, J.R.; Spencer, T.; Murray, A.L.; Arnold, N.S. Geostatistical analysis of sediment deposition in two small tidal wetlands, Norfolk, UK. J. Coast. Res. 1985, 11, 308–321. [Google Scholar]
- Reed, D.J.; Spencer, T.; Murray, A.L.; French, J.R.; Leonard, L. Marsh surface sediment deposition and the role of tidal creeks: Implications for created and managed coastal marshes. J. Coast. Conserv. 1999, 5, 81–90. [Google Scholar] [CrossRef]
- Soille, P.; Vogt, P. Morphological segmentation of binary patterns. Pattern Recognit. Lett. 2009, 30, 456–459. [Google Scholar] [CrossRef]
- Vogt, P. GUIDOS: Tools for the assessment of pattern, connectivity, and fragmentation. In Proceedings of the EGU General Assembly 2013, Vienna, Austria, 7–12 April 2013; Volume 15, p. 13526. [Google Scholar]
- Ostapowicz, K.; Vogt, P.; Riitters, K.H.; Kozak, J.; Estreguil, C. Impact of scale on morphological spatial pattern of forest. Landsc. Ecol. 2008, 23, 1107–1117. [Google Scholar] [CrossRef]
- Saura, S.; Vogt, P.; Velázquez, J.; Hernando, A.; Tejera, R. Key structural forest connectors can be identified by combining landscape spatial pattern and network analyses. For. Ecol. Manag. 2011, 262, 150–160. [Google Scholar] [CrossRef]
- Chang, Q.; Liu, X.; Wu, J.; He, P. MSPA-Based Urban Green Infrastructure Planning and Management Approach for Urban Sustainability: Case Study of Longgang in China. J. Urban Plan. Dev. 2015, 141, A5014006. [Google Scholar] [CrossRef]
- French, J.R.; Stoddart, D.R. Hydrodynamics of saltmarsh creek systems: Implications for marsh morphological development and material exchange. Earth Surf. Process. Landforms 1992, 17, 235–252. [Google Scholar] [CrossRef]
- Friedrichs, C.T.; Perry, J.E. Tidal salt marsh morphodynamics: A synthesis. J. Coast. Res. 2001, 27, 7–37. [Google Scholar]
- Adam, P. Saltmarsh Ecology; Cambridge University Press: Cambridge, UK, 1990; p. 461. [Google Scholar]
- Pethick, J. The distribution of salt pans on tidal salt marshes. J. Biogeogr. 1974, 1, 57–62. [Google Scholar] [CrossRef]
- Yapp, R.H.; Johns, D.; Jones, O.T. The salt marshes of the Dovey Estuary. J. Ecol. 1917, 5, 65–103. [Google Scholar] [CrossRef]
- Goudie, A. Characterising the distribution and morphology of creeks and pans on salt marshes in England and Wales using Google Earth. Estuarine Coast. Shelf Sci. 2013, 129, 112–123. [Google Scholar] [CrossRef]
- Murray, A.B.; Knaapen, M.A.; Tal, M.; Kirwan, M.L. Biomorphodynamics: Physical-biological feedbacks that shape landscapes. Water Resour. Res. 2008, 44. [Google Scholar] [CrossRef]
- Spencer, T.; Möller, I.; Rupprecht, F.; Bouma, T.J.; van Wesenbeeck, B.K.; Kudella, M.; Paul, M.; Jensen, K.; Wolters, G.; Miranda-Lange, M.; et al. Salt marsh surface survives true-to-scale simulated storm surges. Earth Surf. Process. Landforms 2016, 41. [Google Scholar] [CrossRef]
- Kennedy, R.E.; Yang, Z.; Cohen, W.B. Detecting trends in forest disturbance and recovery using yearly Landsat time series: 1. LandTrendr—Temporal segmentation algorithms. Remote Sens. Environ. 2010, 114, 2897–2910. [Google Scholar] [CrossRef]
- Kennedy, R.; Yang, Z.; Gorelick, N.; Braaten, J.; Cavalcante, L.; Cohen, W.; Healey, S. Implementation of the LandTrendr Algorithm on Google Earth Engine. Remote Sens. 2018, 10, 691. [Google Scholar] [CrossRef]
- Reed, D.J. Sediment dynamics and deposition in a retreating coastal salt marsh. Estuarine Coast. Shelf Sci. 1988, 26, 67–79. [Google Scholar] [CrossRef]
- Kirwan, M.L.; Langley, J.A.; Guntenspergen, G.R.; Megonigal, J.P. The impact of sea-level rise on organic matter decay rates in Chesapeake Bay brackish tidal marshes. Biogeosciences 2013, 10, 1869–1876. [Google Scholar] [CrossRef]
- Reef, R.; Schuerch, M.; Christie, E.K.; Möller, I.; Spencer, T. The effect of vegetation height and biomass on the sediment budget of a European saltmarsh. Estuarine Coast. Shelf Sci. 2018, 202, 125–133. [Google Scholar] [CrossRef]
- Temmerman, S.; Govers, G.; Wartel, S.; Meire, P. Spatial and temporal factors controlling short-term sedimentation in a salt and freshwater tidal marsh, scheldt estuary, Belgium, SW Netherlands. Earth Surf. Process. Landforms 2003, 28, 739–755. [Google Scholar] [CrossRef]
- Reed, D.J. The impact of sea-level rise on coastal salt marshes. Prog. Phys. Geogr. 1990, 14, 465–481. [Google Scholar] [CrossRef]
- Ursino, N.; Silvestri, S.; Marani, M. Subsurface flow and vegetation patterns in tidal environments. Water Resour. Res. 2004, 40. [Google Scholar] [CrossRef]
- Shen, C.; Zhang, C.; Xin, P.; Kong, J.; Li, L. Salt Dynamics in Coastal Marshes: Formation of Hypersaline Zones. Water Resour. Res. 2018, 54, 3259–3276. [Google Scholar] [CrossRef]
- Neumeier, U.; Poulin, P.; Roge, M.; Morisette, A.; Huard, A.M. Morphology and evolution of salt marsh pans in the lower St. Lawrence Estuary. In Proceedings of the Coastal Dynamics, Arcachon, France, 24–28 June 2013; pp. 1275–1286. [Google Scholar]
- French, J.; Spencer, T.; Stoddart, D.R. Backbarrier Salt Marshes of the North Norfolk Coast: Geomorphic Developments and Response to Rising Sea-Levels; Discussion Papers in Conservation; Ecology and Conservation Unit, University College London: London, UK, 1990; Volume 54, pp. 1–3. [Google Scholar]
- Ford, H.; Garbutt, A.; Ladd, C.; Malarkey, J.; Skov, M.W. Soil stabilization linked to plant diversity and environmental context in coastal wetlands. J. Veg. Sci. 2016, 27, 259–268. [Google Scholar] [CrossRef]
- Bernik, B.; Pardue, J.; Blum, M. Soil erodibility differs according to heritable trait variation and nutrient-induced plasticity in the salt marsh engineer Spartina alterniflora. Mar. Ecol. Prog. Ser. 2018, 601, 1–14. [Google Scholar] [CrossRef]
- Rouse, J.; Haas, R.; Schell, J.; Deering, D. Monitoring The vernal Advancement and Retrogradation (Green Wave Effect) of Natural Vegetation; Progress Report RSC 1978-1; Remote Sensing Center, Texas A&M Univ: College Station, TX, USA, 1973; p. 93. [Google Scholar]
- Ke, Y.; Im, J.; Lee, J.; Gong, H.; Ryu, Y. Characteristics of Landsat 8 OLI-derived NDVI by comparison with multiple satellite sensors and in-situ observations. Remote Sens. Environ. 2015, 164, 298–313. [Google Scholar] [CrossRef]
- Sulla-Menashe, D.; Friedl, M.A.; Woodcock, C.E. Sources of bias and variability in long-term Landsat time series over Canadian boreal forests. Remote Sens. Environ. 2016, 177, 206–219. [Google Scholar] [CrossRef]
- Roy, D.; Zhang, H.; Ju, J.; Gomez-Dans, J.; Lewis, P.; Schaaf, C.; Sun, Q.; Li, J.; Huang, H.; Kovalskyy, V. A general method to normalize Landsat reflectance data to nadir BRDF adjusted reflectance. Remote Sens. Environ. 2016, 176, 255–271. [Google Scholar] [CrossRef]
- Evans, B. Processes Governing Saltmarsh Morphodynamics Methodological Challenges and Spatial Variability. Master’s Thesis, Department of Geography, University of Cambridge, Cambridge, UK, 2011. [Google Scholar]






| Feature | Classification Rule | Interpretation |
|---|---|---|
| Background | Background pixels | Salt pans or pools if internal, channel or mudflat if external |
| Core | Areas greater than the edge distance from nearest background pixel | Large, coherent marsh areas |
| Edge | Areas within the edge distance of background pixels | Margins of core areas with channel connectivity |
| Perforation | Edge pixels entirely enclosing an area of background pixels | Margins of salt pans or pools without channel connectivity |
| Branch | Strip less than twice the edge distance wide that joins to a core area at one end | Narrow extensions from core areas |
| Islet | Foreground area too small to contain any core area | Small, isolated marsh fragments |
| Bridge | Strip less than twice the edge distance wide that joins two core areas | Narrow causeway between larger coherent marsh areas |
| Bridge in Edge | Bridge joining two areas of edge class | As above but for edge areas |
| Bridge in Perforation | Bridge joining two areas of perforation class | Narrow strip of marsh separating marsh areas that are distinct but entirely contained within a salt pan or pool |
| Loop | Strip less than twice the edge distance wide that joins to the same core area at both ends | Narrow strip of marsh enclosing a salt pan or pool |
| Loop in Edge | Strip less than twice the edge distance wide that joins to the same edge area at both ends | Narrow strip of marsh enclosing a salt pan or pool |
| Loop in Perforation | Strip less than twice the edge distance wide that joins to the same perforation area at both ends | Narrow strip of marsh enclosing a smaller salt pan or pool within a larger one |
| Adjusted | x | f-Stat | p-Value | ||
|---|---|---|---|---|---|
| CD | 0.37 | 7.3 | N/A | 17.4 | ≤0.001 |
| ED | 0.76 | −2.9 | 4.0 | 147 | ≤0.001 |
| pCore | 0.63 | −2.6 | 1.9 | 85.7 | ≤0.001 |
| pUncon | 0.63 | −2.0 | −4.2 | 83.9 | ≤0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Evans, B.R.; Möller, I.; Spencer, T. Topological and Morphological Controls on Morphodynamics of Salt Marsh Interiors. J. Mar. Sci. Eng. 2021, 9, 311. https://doi.org/10.3390/jmse9030311
Evans BR, Möller I, Spencer T. Topological and Morphological Controls on Morphodynamics of Salt Marsh Interiors. Journal of Marine Science and Engineering. 2021; 9(3):311. https://doi.org/10.3390/jmse9030311
Chicago/Turabian StyleEvans, Ben R., Iris Möller, and Tom Spencer. 2021. "Topological and Morphological Controls on Morphodynamics of Salt Marsh Interiors" Journal of Marine Science and Engineering 9, no. 3: 311. https://doi.org/10.3390/jmse9030311
APA StyleEvans, B. R., Möller, I., & Spencer, T. (2021). Topological and Morphological Controls on Morphodynamics of Salt Marsh Interiors. Journal of Marine Science and Engineering, 9(3), 311. https://doi.org/10.3390/jmse9030311