
Differentiation of Crystal Cells, Gravity-Sensing Cells in the Placozoan Trichoplax adhaerens


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
- Calcofluor White (Sigma-Aldrich; St. Louis, MO, USA) diluted 1:1000 in PBST was applied for two hours; after washing with PBS, propidium iodide was added for 20 min.
- Samples were incubated in blocking buffer (3% normal goat serum, 2% horse serum, 1% BSA in PBS) for one hour and then Col-F dye (20 µM final concentration; ImmunoChemistry Technologies, Bloomington, MN, USA) was applied for 30 min at 37 °C. After washing with PBS, Hoechst was added for 20 min.
- A cocktail of seven fluorescein conjugated lectins (Concanavalin A, Dolichos biflorus agglutinin, peanut agglutinin, Ricinus communis agglutinin, soybean agglutinin, Ulex europaeus agglutinin, and wheat germ agglutinin, all from Vector Laboratories, USA) each diluted 1:200 in PBST and Hoechst was applied for two hours.
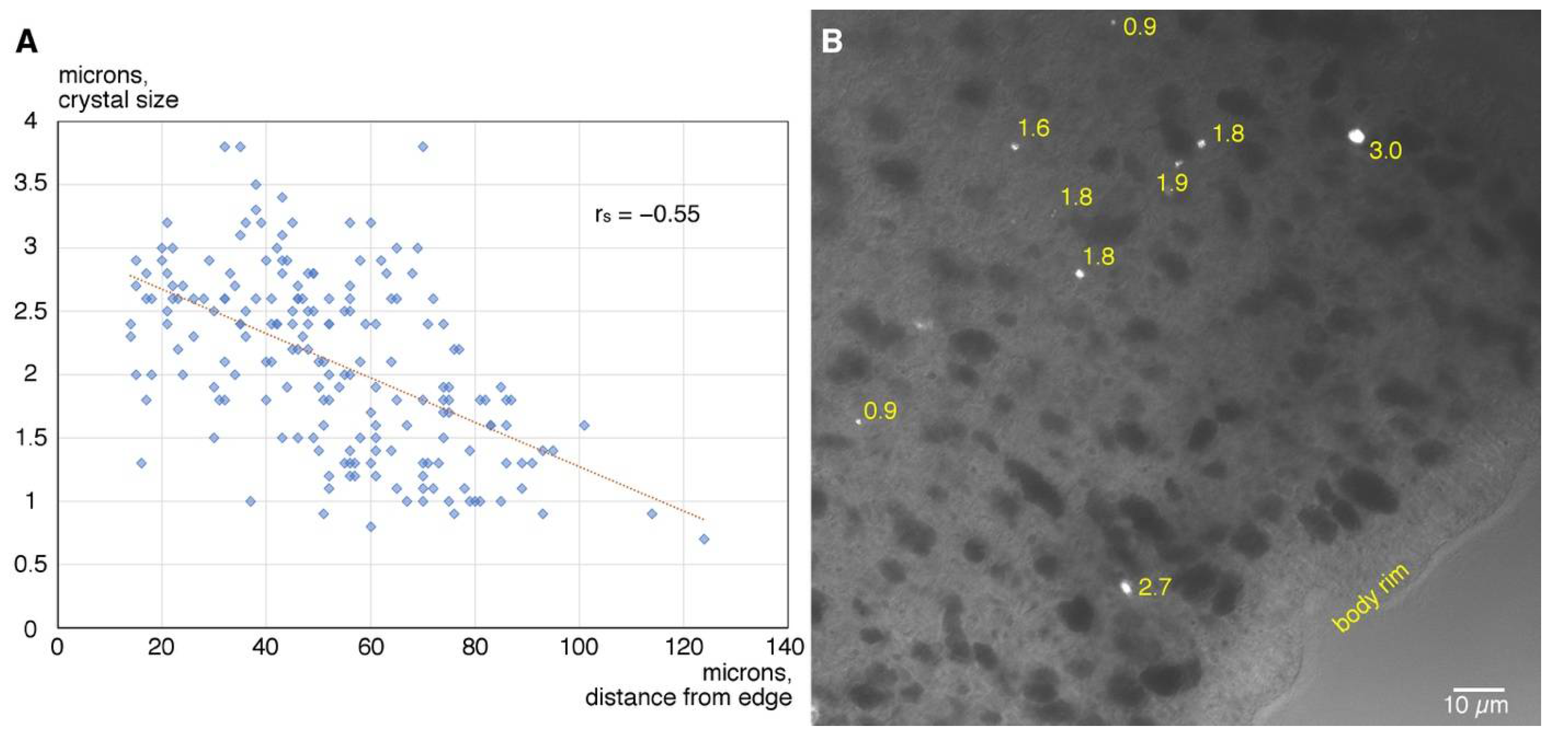
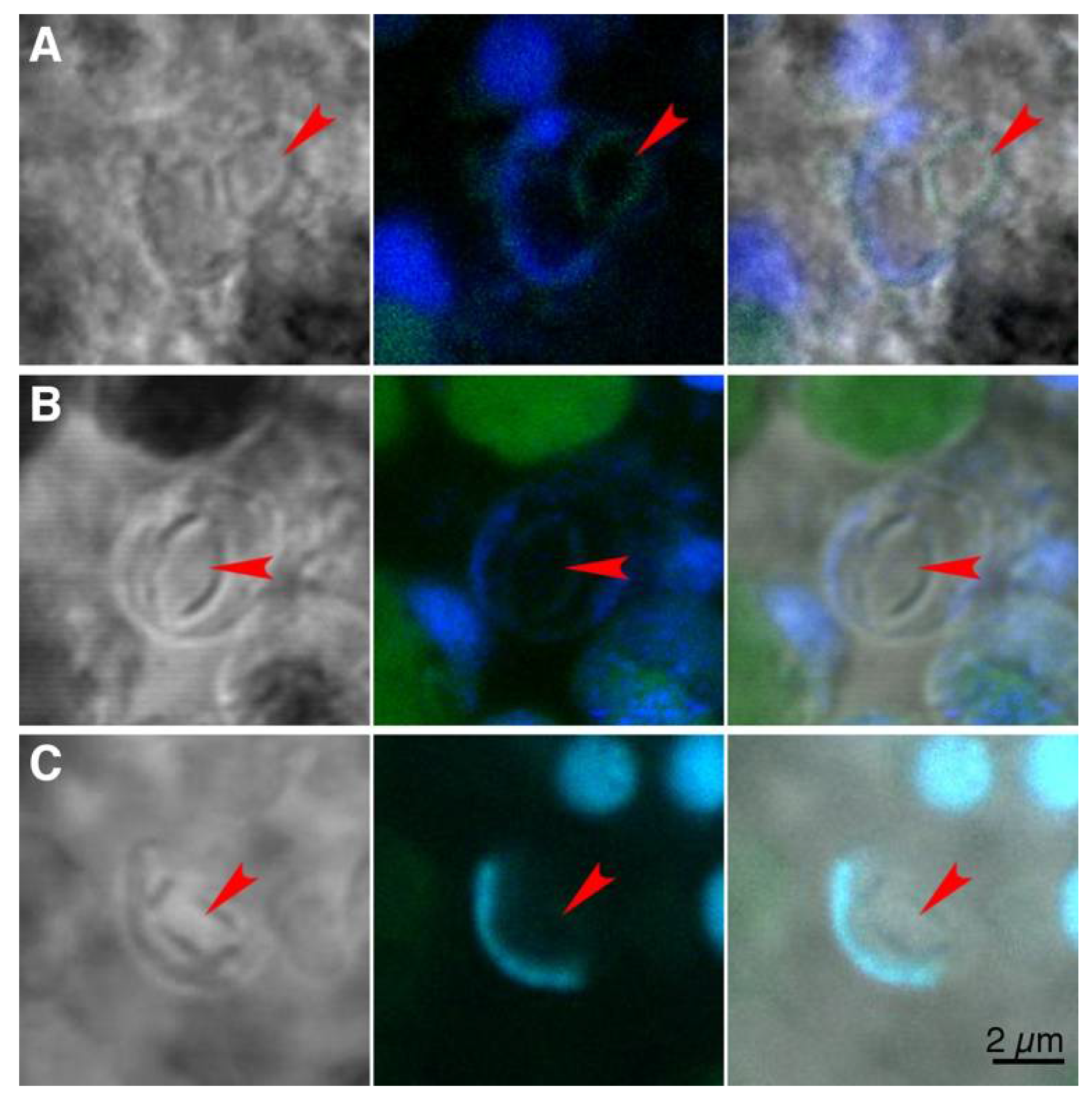
3. Results
4. Discussion
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Schulze, F.E. Trichoplax adhaerens, nov. gen., nov. spec. Zool. Anz. 1891, 6, 92–97. [Google Scholar]
- Grell, K.G.; Ruthmann, A. Placozoa. In Microscopic Anatomy of Invertebrates, Volume 2: Placozoa, Porifera, Cnidaria, and Ctenophora; Harrison, F.W., Westfall, J.A., Eds.; Wiley-Liss: New York, NY, USA, 1991; pp. 13–27. [Google Scholar]
- Smith, C.L.; Varoqueaux, F.; Kittelmann, M.; Azzam, R.N.; Cooper, B.; Winters, C.A.; Eitel, M.; Fasshauer, D.; Reese, T.S. Novel cell types, neurosecretory cells, and body plan of the early-diverging metazoan Trichoplax adhaerens. Curr. Biol. 2014, 24, 1565–1572. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.L.; Pivovarova, N.; Reese, T.S. Coordinated feeding behavior in Trichoplax, an animal without synapses. PLoS ONE 2015, 10, e00822-17. [Google Scholar] [CrossRef] [Green Version]
- Schierwater, B.; DeSalle, R. Placozoa. Curr. Biol. 2018, 28, R97–R98. [Google Scholar] [CrossRef] [Green Version]
- Smith, C.L.; Reese, T.S.; Govezensky, T.; Barrio, R.A. Coherent directed movement toward food modeled in Trichoplax, a ciliated animal lacking a nervous system. Proc. Natl. Acad. Sci. USA 2019, 116, 8901–8908. [Google Scholar] [CrossRef] [Green Version]
- Dunn, C.W.; Hejnol, A.; Matus, D.Q.; Pang, K.; Browne, W.E.; Smith, S.A.; Seaver, E.; Rouse, G.W.; Obst, M.; Edgecombe, G.D.; et al. Broad phylogenomic sampling improves resolution of the animal tree of life. Nature 2008, 452, 745–749. [Google Scholar] [CrossRef]
- Moroz, L.L.; Kocot, K.M.; Citarella, M.R.; Dosung, S.; Norekian, T.P.; Povolotskaya, I.S.; Grigorenko, A.P.; Dailey, C.; Berezikov, E.; Buckley, K.M.; et al. The ctenophore genome and the evolutionary origins of neural systems. Nature 2014, 510, 109–114. [Google Scholar] [CrossRef]
- Pett, W.; Adamski, M.; Adamska, M.; Francis, W.R.; Eitel, M.; Pisani, D.; Wörheide, G. The role of homology and orthology in the phylogenomic analysis of metazoan gene content. Mol. Biol. Evol. 2019, 36, 643–649. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simion, P.; Philippe, H.; Baurain, D.; Jager, M.; Richter, D.J.; Di Franco, A.; Roure, B.; Satoh, N.; Quéinnec, É.; Ereskovsky, A.; et al. A large and consistent phylogenomic dataset supports sponges as the sister group to all other animals. Curr. Biol. 2017, 27, 958–967. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ruthmann, A.; Behrendt, G.; Wahl, R. The ventral epithelium of Trichoplax adhaerens (Placozoa): Cytoskeletal structures, cell contacts and endocytosis. Zoomorphology 1986, 106, 115–122. [Google Scholar] [CrossRef]
- Smith, C.L.; Reese, T.S. Adherens junctions modulate diffusion between epithelial cells in Trichoplax adhaerens. Biol. Bull. 2016, 231, 216–224. [Google Scholar] [CrossRef]
- Smith, C.L.; Mayorova, T.D. Insights into the evolution of digestive systems from studies of Trichoplax adhaerens. Cell Tissue Res. 2019, 377, 353–367. [Google Scholar] [CrossRef] [Green Version]
- Mayorova, T.D.; Hammar, K.; Winters, C.A.; Reese, T.S.; Smith, C.L. The ventral epithelium of Trichoplax adhaerens deploys in distinct patterns cells that secrete digestive enzymes, mucus or diverse neuropeptides. Biol. Open 2019, 8, bio045674. [Google Scholar] [CrossRef] [Green Version]
- Varoqueaux, F.; Williams, E.A.; Grandemange, S.; Truscello, L.; Kamm, K.; Schierwater, B.; Jékely, G.; Fasshauer, D. High Cell Diversity and Complex Peptidergic Signaling Underlie Placozoan Behavior. Curr. Biol. 2018, 28, 3495–3501.e2. [Google Scholar] [CrossRef] [Green Version]
- Rassat, J.; Ruthmann, A. Trichoplax adhaerens F.E. Schulze (placozoa) in the scanning electron microscope. Zoomorphologie 1979, 93, 59–72. [Google Scholar] [CrossRef]
- Mayorova, T.D.; Hammar, K.; Jung, J.H.; Aronova, M.A.; Zhang, G.; Winters, C.A.; Reese, T.S.; Smith, C.L. Placozoan fiber cells: Mediators of innate immunity and participants in wound healing. Nat. Sci. Rep. 2021, Submitted. [Google Scholar]
- Mayorova, T.D.; Smith, C.L.; Hammar, K.; Winters, C.A.; Pivovarova, N.B.; Aronova, M.A.; Leapman, R.D.; Reese, T.S. Cells containing aragonite crystals mediate responses to gravity in Trichoplax adhaerens (Placozoa), an animal lacking neurons and synapses. PLoS ONE 2018, 13, e0190905. [Google Scholar] [CrossRef] [PubMed]
- Signorovitch, A.Y.; Dellaporta, S.L.; Buss, L.W. Molecular signatures for sex in the Placozoa. Proc. Natl. Acad. Sci. USA 2005, 102, 15518–15522. [Google Scholar] [CrossRef] [Green Version]
- Schierwater, B. My favorite animal, Trichoplax adhaerens. BioEssays 2005, 27, 1294–1302. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, M.; Begovic, E.; Chapman, J.; Putnam, N.H.; Hellsten, U.; Kawashima, T.; Kuo, A.; Mitros, T.; Salamov, A.; Carpenter, M.L.; et al. The Trichoplax genome and the nature of placozoans. Nature 2008, 454, 955–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zuccolotto-Arellano, J.; Cuervo-González, R. Binary fission in Trichoplax is orthogonal to the subsequent division plane. Mech. Dev. 2020, 162, 103608. [Google Scholar] [CrossRef]
- Schierwater, B.; Osigus, H.J.; Bergmann, T.; Blackstone, N.W.; Hadrys, H.; Hauslage, J.; Humbert, P.O.; Kamm, K.; Kvansakul, M.; Wysocki, K.; et al. The enigmatic Placozoa part 1: Exploring evolutionary controversies and poor ecological knowledge. BioEssays 2021, 43, 2100080. [Google Scholar] [CrossRef] [PubMed]
- Von Der Chevallerie, K.; Rolfes, S.; Schierwater, B. Inhibitors of the p53-Mdm2 interaction increase programmed cell death and produce abnormal phenotypes in the placozoon Trichoplax adhaerens (F.E. Schulze). Dev. Genes Evol. 2014, 224, 79–85. [Google Scholar] [CrossRef]
- Romanova, D.Y.; Smirnov, I.V.; Nikitin, M.A.; Kohn, A.B.; Borman, A.I.; Malyshev, A.Y.; Balaban, P.M.; Moroz, L.L. Sodium action potentials in placozoa: Insights into behavioral integration and evolution of nerveless animals. Biochem. Biophys. Res. Commun. 2020, 532, 120–126. [Google Scholar] [CrossRef]
- Schierwater, B.; Osigus, H.J.; Bergmann, T.; Blackstone, N.W.; Hadrys, H.; Hauslage, J.; Humbert, P.O.; Kamm, K.; Kvansakul, M.; Wysocki, K.; et al. The enigmatic Placozoa part 2: Exploring evolutionary controversies and promising questions on earth and in space. BioEssays 2021, 43, 2100083. [Google Scholar] [CrossRef] [PubMed]
- Ferrero, E. A Fine Structural Analysis of the Statocyst in Turbellaria Acsela. Zool. Scr. 1973, 2, 5–16. [Google Scholar] [CrossRef]
- Ehlers, U. Comparative morphology of statocysts in the Plathelminthes and the Xenoturbellida. Hydrobiologia 1991, 227, 263–271. [Google Scholar] [CrossRef]
- Jackson, A.M.; Buss, L.W. Shiny spheres of placozoans (Trichoplax) function in anti-predator defense. Invertebr. Biol. 2009, 128, 205–212. [Google Scholar] [CrossRef]
- Romanova, D.Y.; Varoqueaux, F.; Daraspe, J.; Nikitin, M.A.; Eitel, M.; Fasshauer, D.; Moroz, L.L. Hidden cell diversity in Placozoa: Ultrastructural insights from Hoilungia hongkongensis. Cell Tissue Res. 2021, 385, 623–637. [Google Scholar] [CrossRef]
- Watabe, N.; Kingsley, R.J. Calcification in Octocorals. In Hard Tissue Mineralization and Demineralization; Suga, S., Watabe, N., Eds.; Springer: Tokyo, Japan, 1992; pp. 127–147. [Google Scholar]
- Conci, N.; Vargas, S.; Wörheide, G. The Biology and Evolution of Calcite and Aragonite Mineralization in Octocorallia. Front. Ecol. Evol. 2021, 9, 623774. [Google Scholar] [CrossRef]
- Achatz, J.G.; Martinez, P. The nervous system of Isodiametra pulchra (Acoela) with a discussion on the neuroanatomy of the Xenacoelomorpha and its evolutionary implications. Front. Zool. 2012, 9, 27. [Google Scholar] [CrossRef] [Green Version]
- Sakagami, T.; Watanabe, K.; Ikeda, R.; Ando, M. Structural analysis of the statocyst and nervous system of Praesagittifera naikaiensis, an acoel flatworm, during development after hatching. Zoomorphology 2021, 140, 183–192. [Google Scholar] [CrossRef]
- Tamm, S.L. Functional consequences of the asymmetric architecture of the ctenophore statocyst. Biol. Bull. 2015, 229, 173–184. [Google Scholar] [CrossRef] [Green Version]
- Ehlers, U. Ultrastructure of the statocysts in the apodous sea cucumber Leptosynapta inhaerens (Holothuroidea, Echinodermata). Acta Zool. 1997, 78, 61–68. [Google Scholar] [CrossRef]
- Taylor, A.R.; Chrachri, A.; Wheeler, G.; Goddard, H.; Brownlee, C. A voltage-gated H+ channel underlying pH homeostasis in calcifying Coccolithophores. PLoS Biol. 2011, 9, e1001085. [Google Scholar] [CrossRef] [PubMed]
- Moroz, L.L. On the independent origins of complex brains and neurons. Brain. Behav. Evol. 2009, 74, 177–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrero, E.A.; Bedini, C. Ultrastructural aspects of nervous-system and statocyst morphogenesis during embryonic development of Convoluta psammophila (Turbellaria, Acoela). Hydrobiologia 1991, 227, 131–137. [Google Scholar] [CrossRef]
- Rohde, K.; Watson, N.; Faubel, A. Ultrastructure of the Statocyst in an Undescribed Species of Luridae (Platyhelminthes, Rhabdocoela, Luridae). Aust. J. Zool. 1993, 41, 215–224. [Google Scholar] [CrossRef]
- Voigt, O.; Adamski, M.; Sluzek, K.; Adamska, M. Calcareous sponge genomes reveal complex evolution of α-carbonic anhydrases and two key biomineralization enzymes. BMC Evol. Biol. 2014, 14, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Bhattacharya, D.; Agrawal, S.; Aranda, M.; Baumgarten, S.; Belcaid, M.; Drake, J.L.; Erwin, D.; Foret, S.; Gates, R.D.; Gruber, D.F.; et al. Comparative genomics explains the evolutionary success of reef-forming corals. Elife 2016, 5, e13288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simkiss, K.; Wilbur, K.M. The Control of Mineralization. In Biomineralization; Academic Press Inc.: San-Diego, CA, USA, 1989; pp. 42–57. [Google Scholar]
- Marin, F.; Luquet, G. Unusually Acidic Proteins in Biomineralization. In Handbook of Biomineralization: Biological Aspects and Structure Formation; Bäuerlein, E., Ed.; Wiley-VCH Verlag GmbH & Co. K: Hoboken, NJ, USA, 2007; pp. 273–290. ISBN 9783527316410. [Google Scholar]
- Arias, J.L.; Fernández, M.S. Polysaccharides and proteoglycans in calcium carbonate-based Biomineralization. Chem. Rev. 2008, 108, 4475–4482. [Google Scholar] [CrossRef]
- Ehrlich, H. Chitin and collagen as universal and alternative templates in biomineralization. Int. Geol. Rev. 2010, 52, 661–699. [Google Scholar] [CrossRef]
- Nudelman, F.; Lausch, A.J.; Sommerdijk, N.A.J.M.; Sone, E.D. In vitro models of collagen biomineralization. J. Struct. Biol. 2013, 183, 258–269. [Google Scholar] [CrossRef] [Green Version]
- Conci, N.; Wörheide, G.; Vargas, S. New non-bilaterian transcriptomes provide novel insights into the evolution of coral skeletomes. Genome Biol. Evol. 2019, 11, 3068–3081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinelli, C.; Spring, J. Distinct expression patterns of the two T-box homologues Brachyury and Tbx2/3 in the placozoan Trichoplax adhaerens. Dev. Genes Evol. 2003, 213, 492–499. [Google Scholar] [CrossRef] [Green Version]
- Hadrys, T.; DeSalle, R.; Sagasser, S.; Fischer, N.; Schierwater, B. The trichoplax PaxB gene: A putative proto-Paxa/B/C gene predating the origin of nerve and sensory cells. Mol. Biol. Evol. 2005, 22, 1569–1578. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubuc, T.Q.; Ryan, J.F.; Martindale, M.Q. “Dorsal-Ventral” Genes Are Part of an Ancient Axial Patterning System: Evidence from Trichoplax adhaerens (Placozoa). Mol. Biol. Evol. 2019, 36, 966–973. [Google Scholar] [CrossRef] [Green Version]
- Sebé-Pedrós, A.; Chomsky, E.; Pang, K.; Lara-Astiaso, D.; Gaiti, F.; Mukamel, Z.; Amit, I.; Hejnol, A.; Degnan, B.M.; Tanay, A. Early metazoan cell type diversity and the evolution of multicellular gene regulation. Nat. Ecol. Evol. 2018, 2, 1176–1188. [Google Scholar] [CrossRef] [PubMed]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mayorova, T.D. Differentiation of Crystal Cells, Gravity-Sensing Cells in the Placozoan Trichoplax adhaerens. J. Mar. Sci. Eng. 2021, 9, 1229. https://doi.org/10.3390/jmse9111229
Mayorova TD. Differentiation of Crystal Cells, Gravity-Sensing Cells in the Placozoan Trichoplax adhaerens. Journal of Marine Science and Engineering. 2021; 9(11):1229. https://doi.org/10.3390/jmse9111229
Chicago/Turabian StyleMayorova, Tatiana D. 2021. "Differentiation of Crystal Cells, Gravity-Sensing Cells in the Placozoan Trichoplax adhaerens" Journal of Marine Science and Engineering 9, no. 11: 1229. https://doi.org/10.3390/jmse9111229
APA StyleMayorova, T. D. (2021). Differentiation of Crystal Cells, Gravity-Sensing Cells in the Placozoan Trichoplax adhaerens. Journal of Marine Science and Engineering, 9(11), 1229. https://doi.org/10.3390/jmse9111229