
Dimethylsulphoniopropionate (DMSP) as an Indicator of Bleaching Tolerance in Scleractinian Corals

Abstract
:1. Introduction
2. Experimental Section
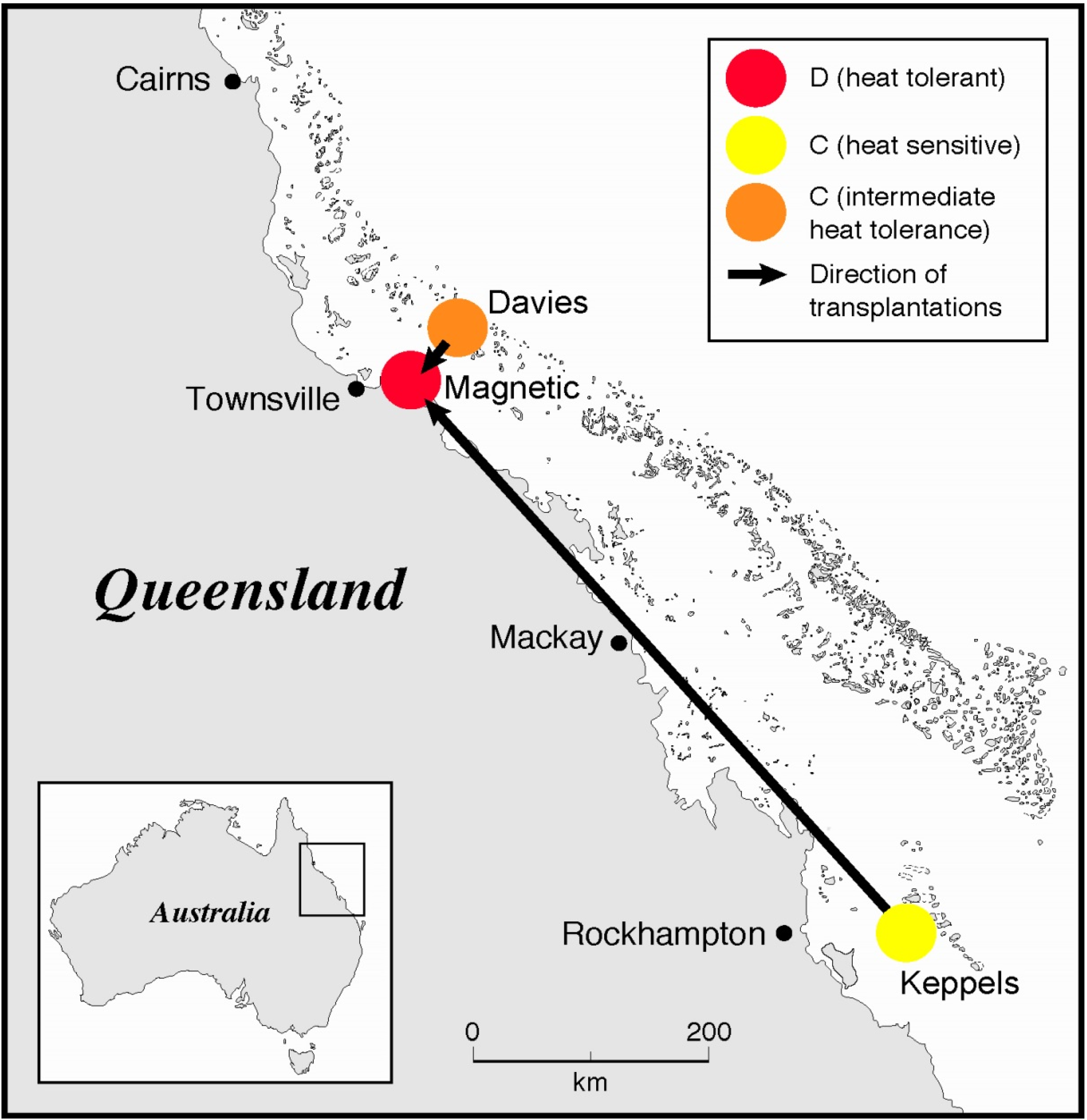
2.1. Coral Collection and Transplantation
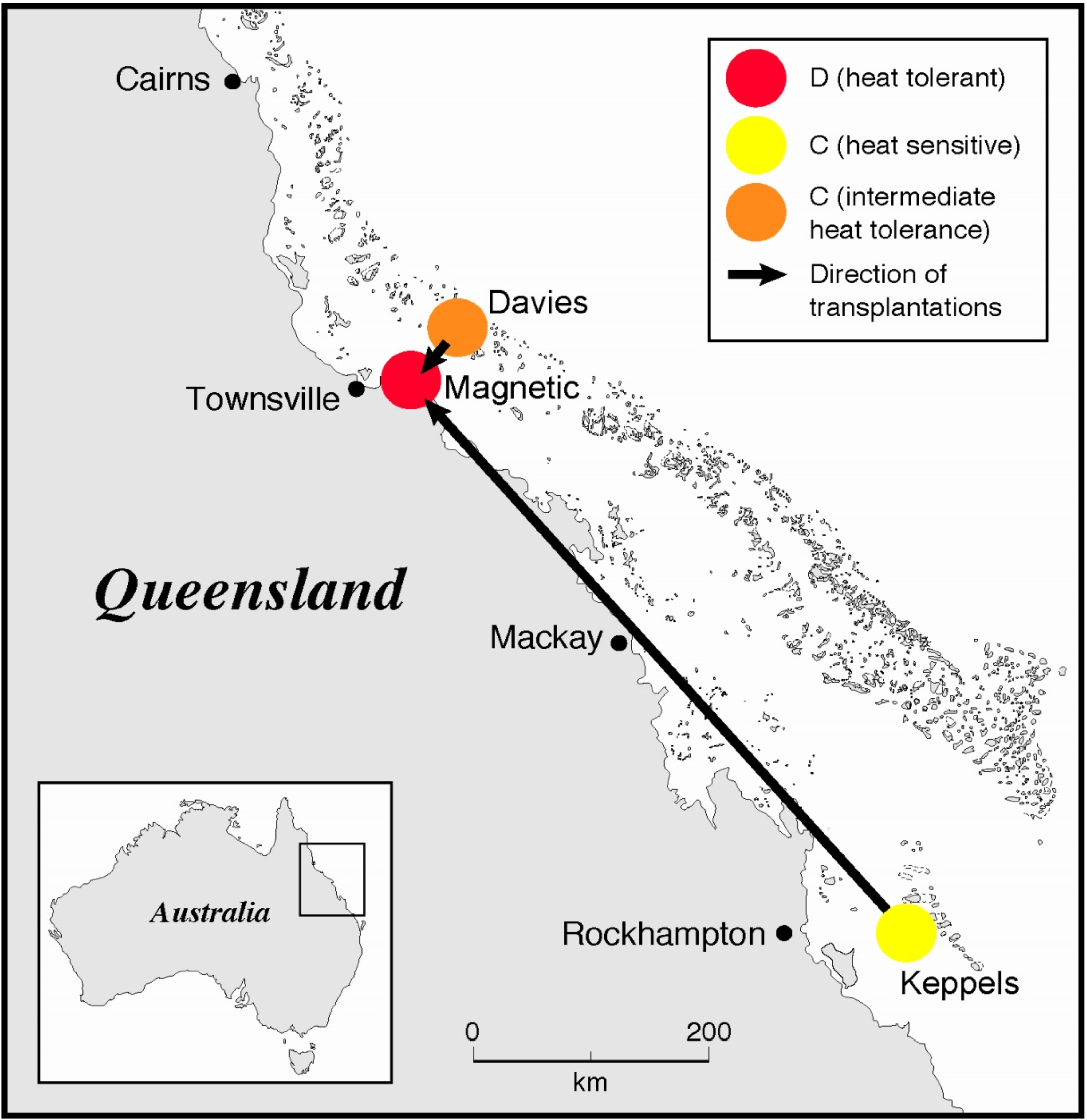
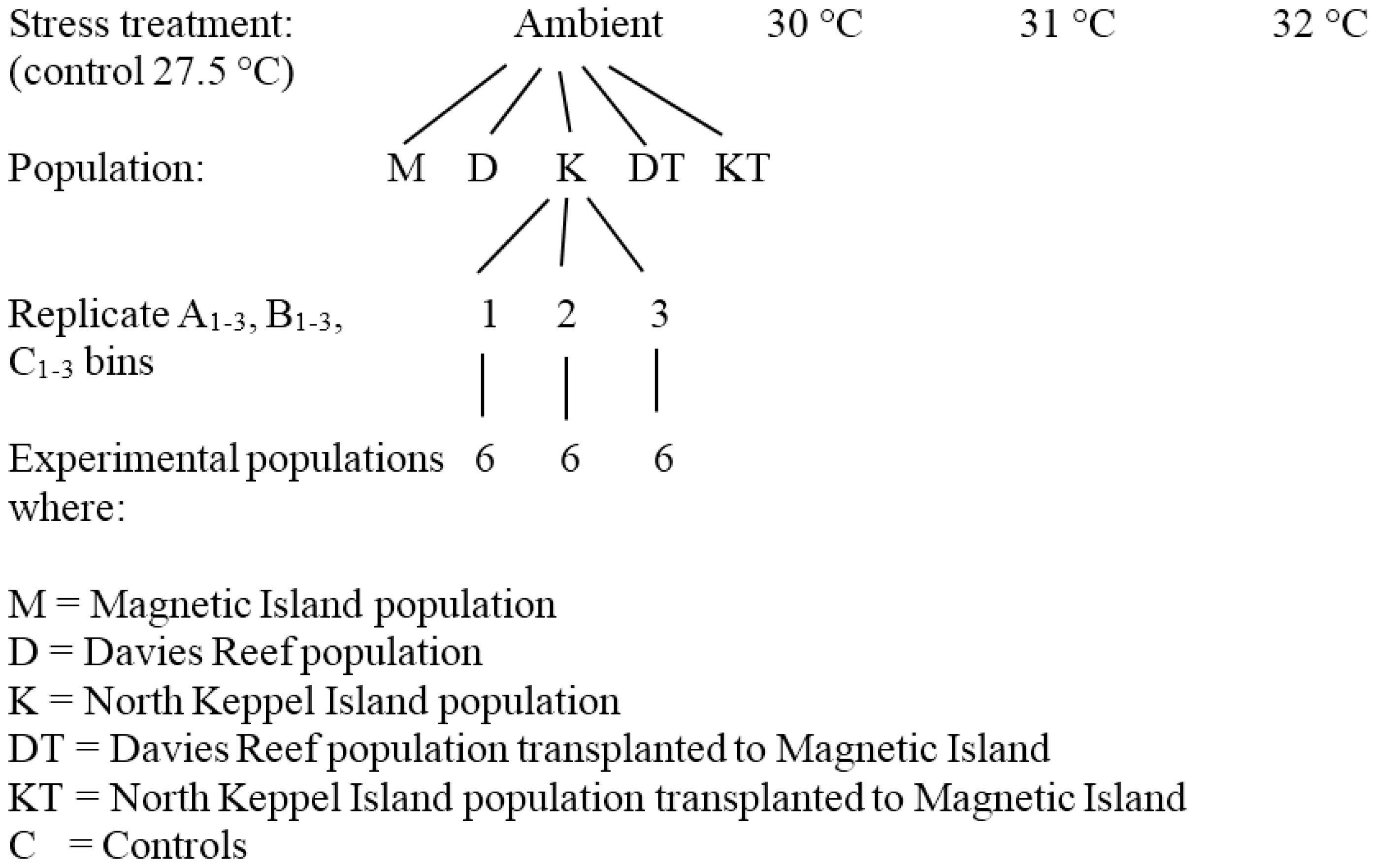
2.2. Temperature Experiment
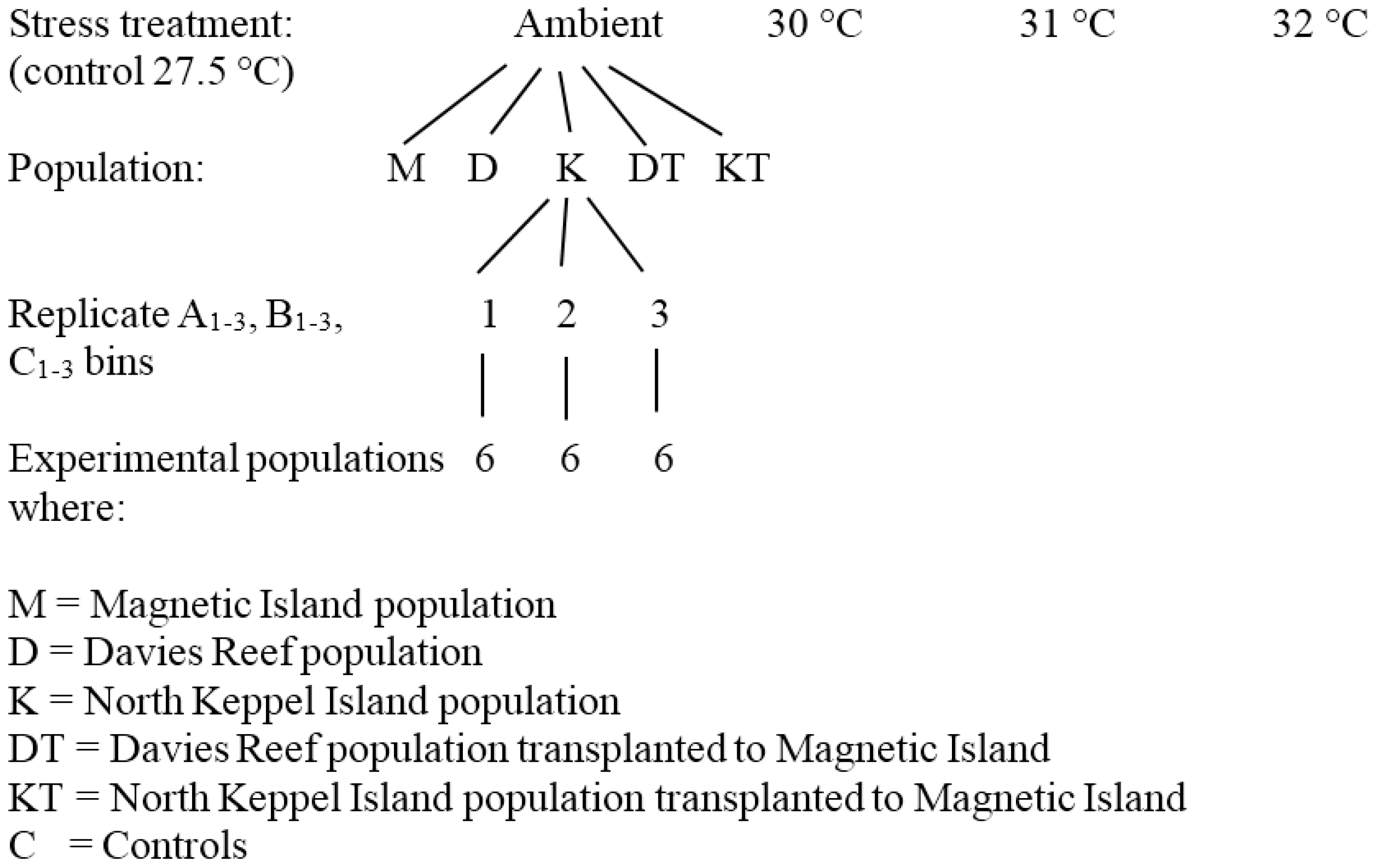
2.3. Zooxanthellae Genotyping
2.4. Zooxanthellae Counts and Coral Surface Area
2.5. Photosynthetic Fitness of the Zooxanthellae
2.6. Visual Observations of Bleaching

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dominant Zooxanthellae Clade Type | ||
|---|---|---|
| Coral Colony | During Transplant | After Transplant |
| Magnetic Island | D | D |
| Keppel Transplant | C2 (80%, sensu) | D |
| D (20%) | ||
| Davies Transplant | C2* | C2* |
2.7. DMSP Analysis
2.8. Statistical Analysis
3. Results
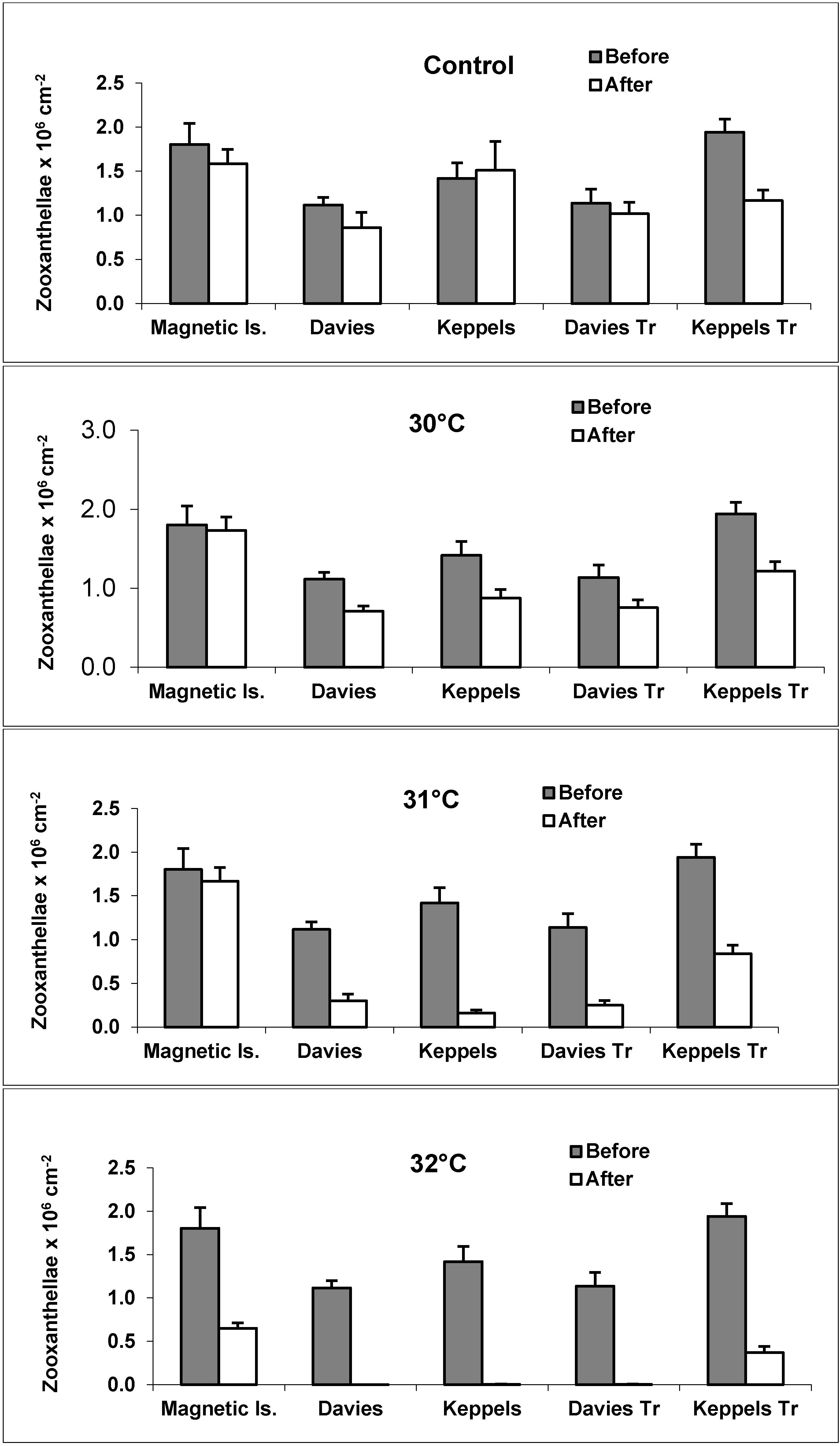
3.1. Zooxanthellae Densities
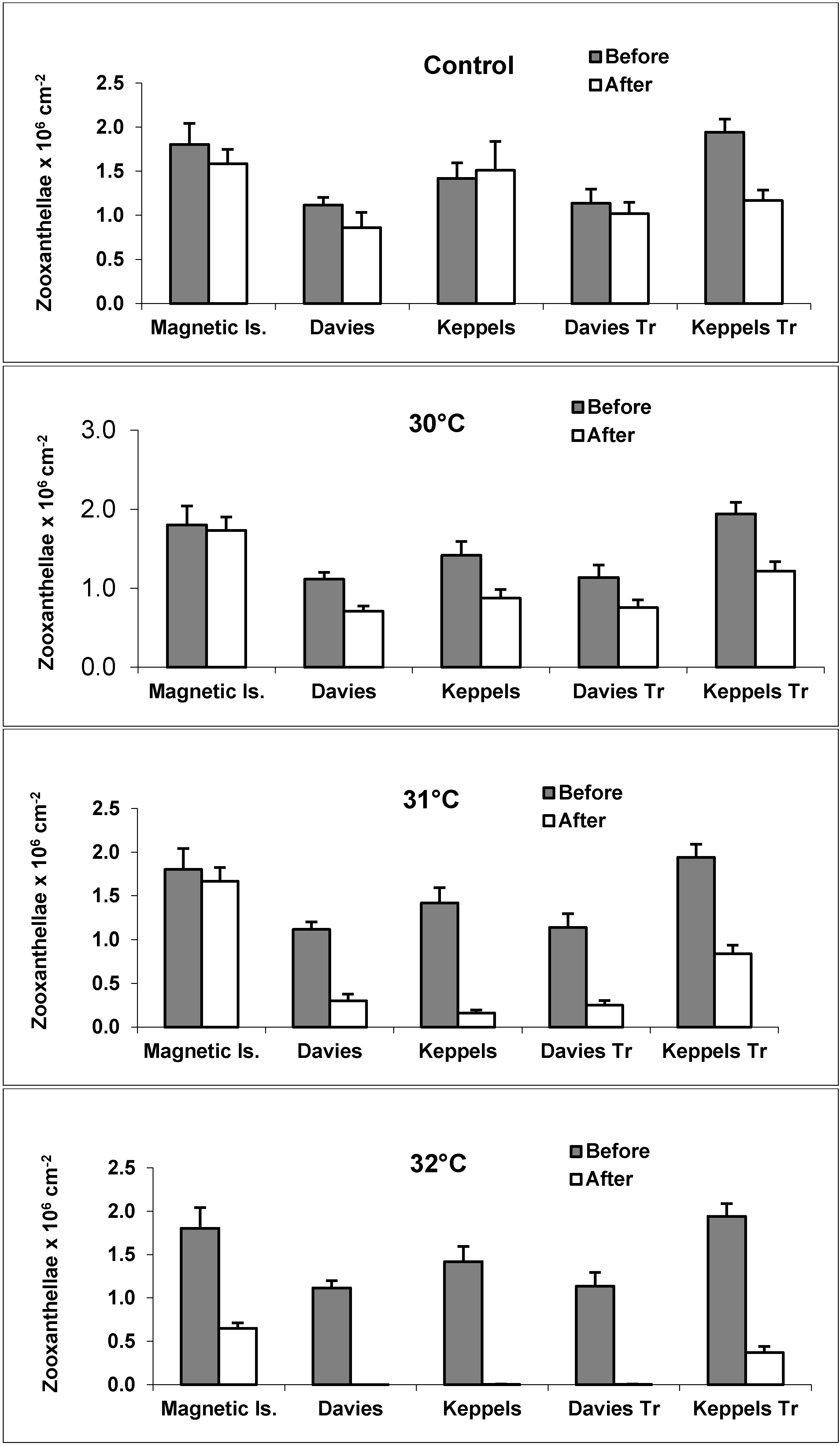
3.2. Bleaching Observations and Zooxanthellae Clade Changes
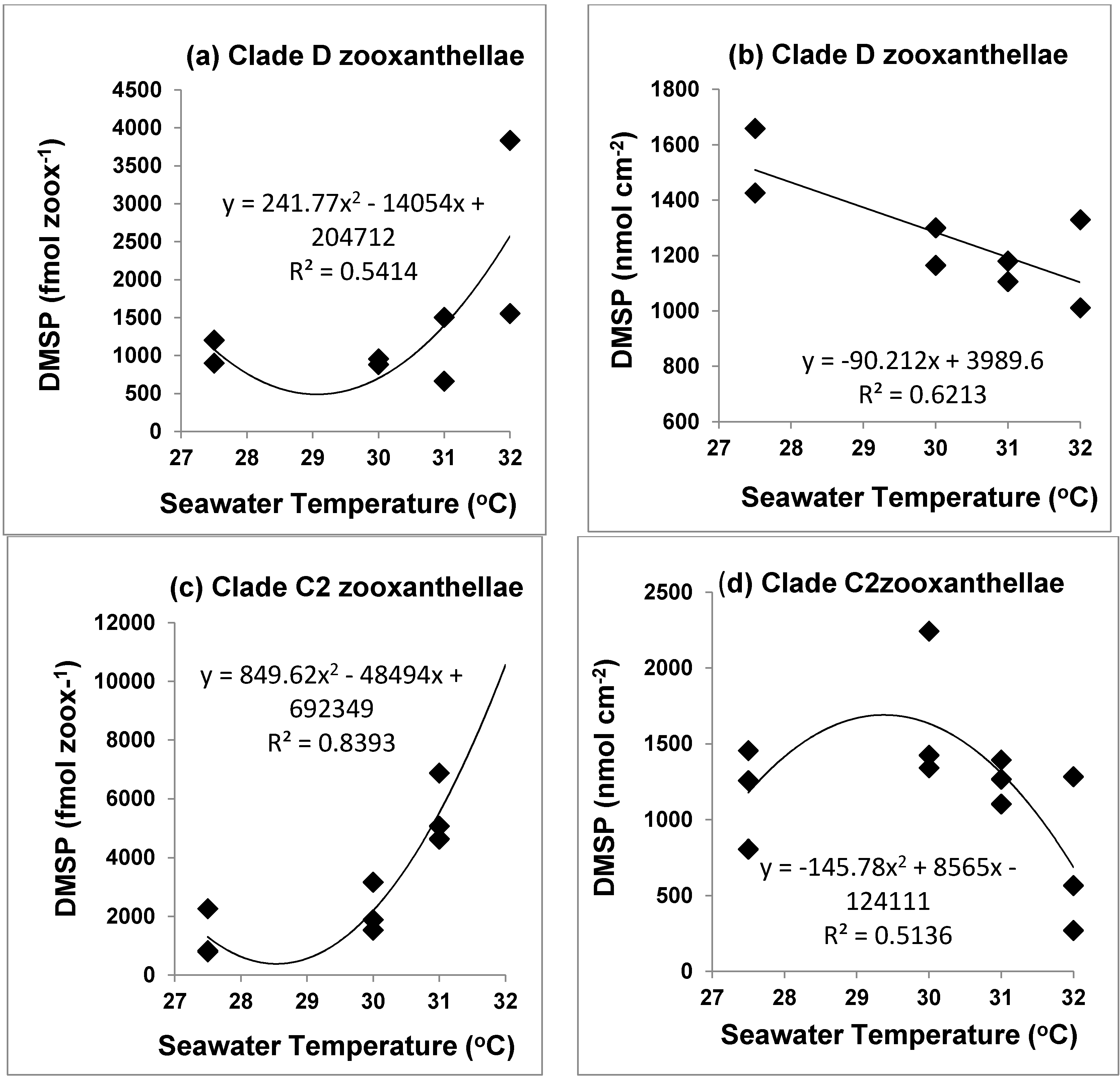
3.3. Tissue DMSP Concentrations
| Coral Colony | Before | Control | 30 °C | 31 °C | 32 °C |
|---|---|---|---|---|---|
| Magnetic Island | 449 ± 161 | 900 ± 99 | 882 ± 93 | 663 ± 70 | 1554 ± 263 # |
| Keppels Transplant | 326 ± 229 | 1204 ± 512 | 956 ± 217 | 1503 ± 504 | 3835 ± 1339 ** |
| Keppels | 520 ± 197 | 833 ± 404 | 1532 ± 323 | 6878 ± 322 | 521 ± 221 *# |
| Davies | 1553 ± 431 | 2262 ± 497 | 3163 ± 805 | 4634 ± 1876 | 1761 ± 2081 *# |
| Davies Transplant | 893 ± 218 | 791 ± 194 | 1886 ± 968 | 5068 ± 1657 | 80 ± 30 *# |
| Coral Colony | Before | Control | 30 °C | 31 °C | 32 °C |
|---|---|---|---|---|---|
| Magnetic Island | 809 ± 290 | 1426 ± 157 | 1529 ± 161 | 1106 ± 116 | 1011 ± 171 |
| Keppels Transplant | 634 ± 444 | 1404 ± 198 | 1165 ± 264 | 1264 ± 424 | 1418 ± 495 |
| Keppels | 738 ± 280 | 1258 ± 610 | 1342 ± 283 | 1103 ± 536 | 1283 ± 777 |
| Davies | 1734 ± 481 | 1455 ± 803 | 2244 ± 575 | 1393 ± 564 | 567 ± 67 * |
| Davies Transplant | 1016 ± 248 | 1100 ± 605 | 1424 ± 731 | 1266 ± 414 | 270 ± 99 * |
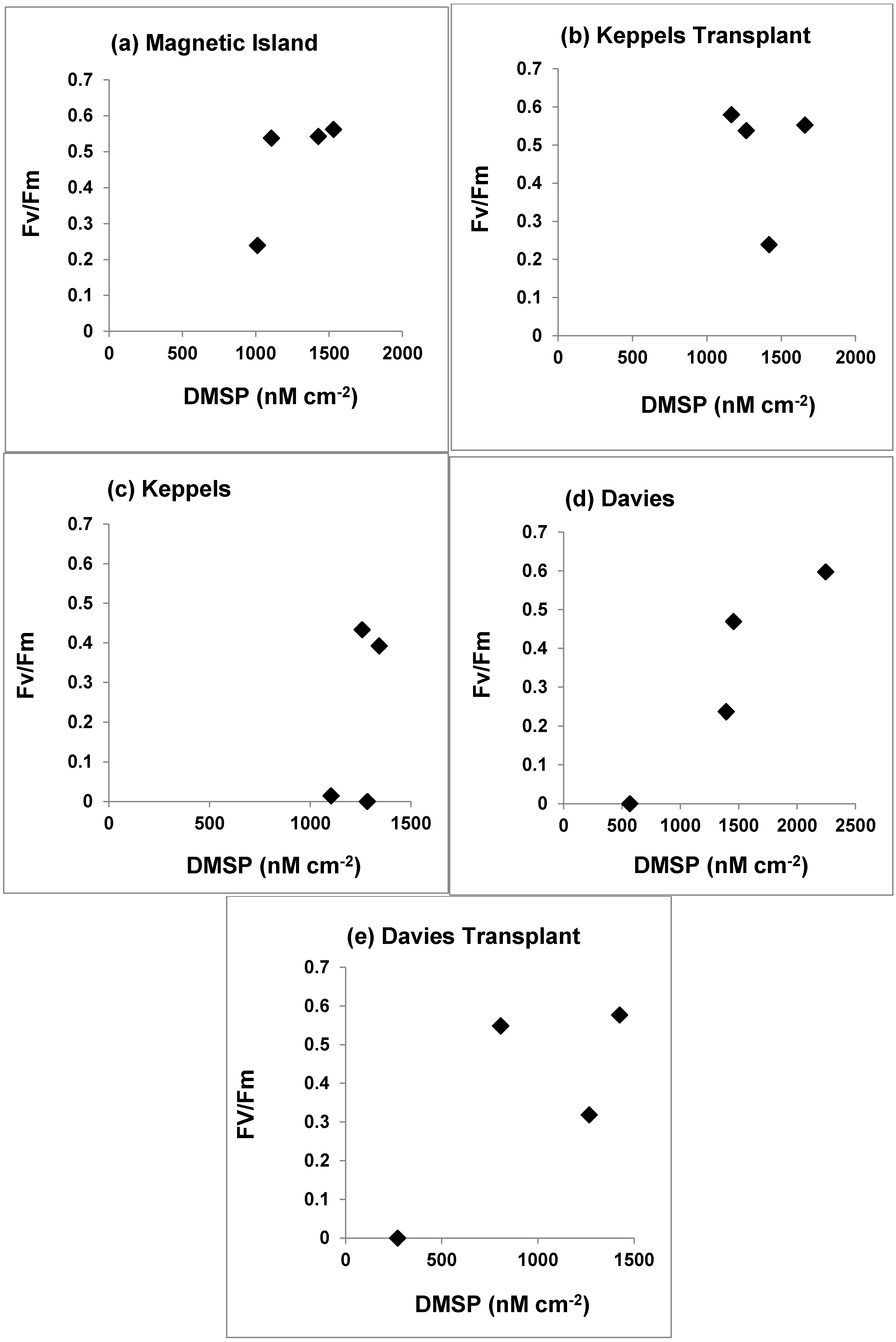
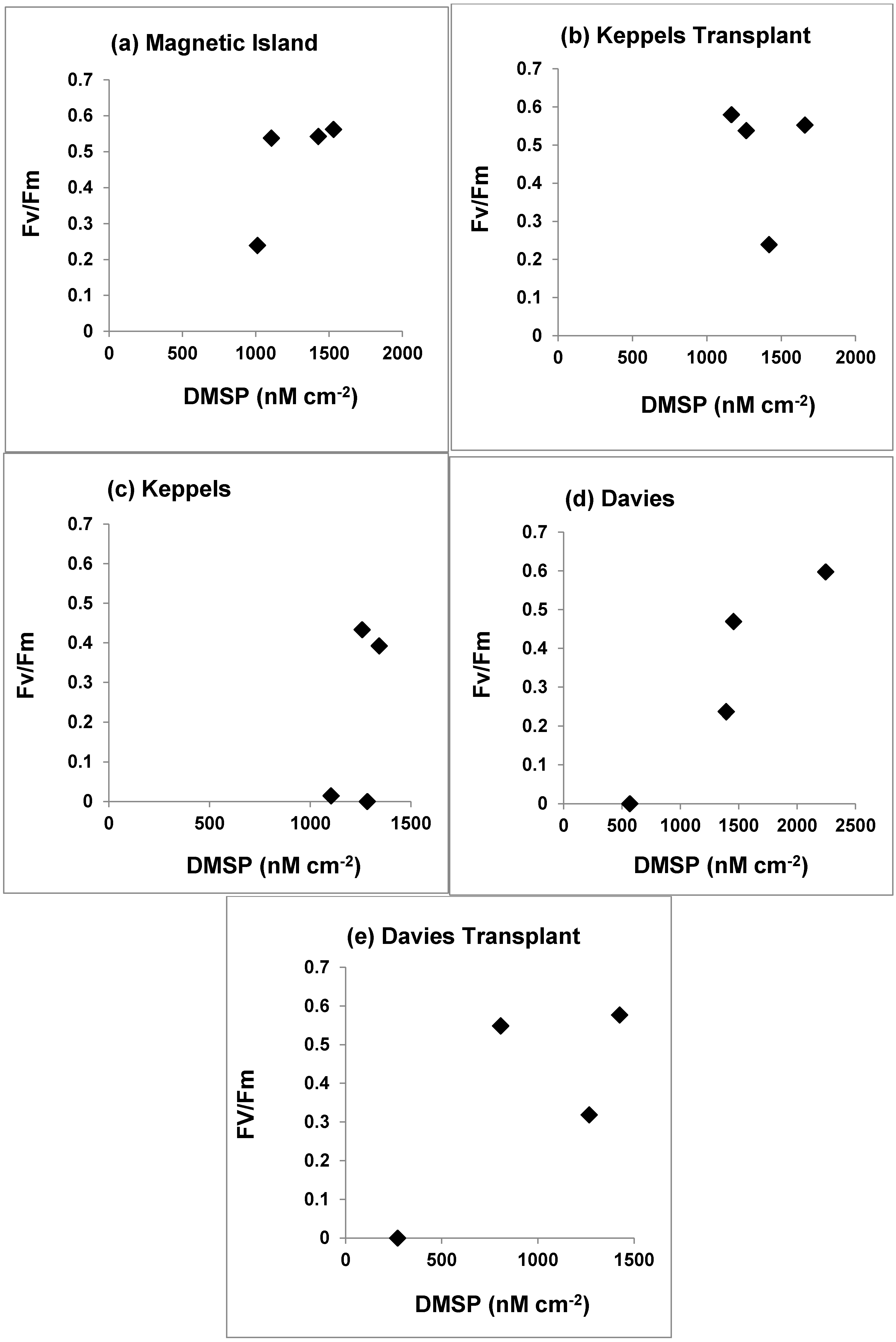
3.4. Photosynthetic Yield and DMSP Concentrations
| Region | Date Collected | Species | DMSP Concentration (nmol cm−2) | Author |
|---|---|---|---|---|
| Magnetic Is. Nelly Bay, GBR | 1992 | A. formosa (unbleached) | 237 | Broadbent et al. (2002) [27] |
| A. formosa (bleached) | 572 | |||
| Acropora palifera | 3842 ± 1237 | |||
| Heron Island GBR | 2012 | A. aspera (unbleached) | 97 ± 39 | Deschaseaux et al. (2014a) [31] |
| A. aspera * (bleached-heat stress) | 941 ± 121 | |||
| A. aspera ** (bleached-direct sunlight) | 1820 ± 168 | |||
| Davies Reef, GBR | A. millepora | 247 | Tapiolas et al. (2013) [47] | |
| Trunk Reef, GBR | A. millipora | 2720–4260 *** | Tapiolas et al. (2013) [47] | |
| Magnetic Island | 2011 | A. millipora | 51 ± 4.1 # | Deschaseaux et al. (2014b) [38] |
| Nelly Bay, GBR | A. tenuis (axenic cultures) | 63 ± 4.7 ## | ||
| Heron Island | 2001 | A. intermedia | 124 ± 21 | Jones et al. (2014) [39] |
| Heron Island | 2002 | A. intermedia ### | 544 ± 194 | Jones et al. (2014) [39] |
| Magnetic Island Geoffrey Bay GBR | 2002 | A. millipora (control) | 1426 ± 157 | This study |
4. Discussion
4.1. Zooxanthellae Density as a Thermal Stress Indicator
4.2. Photosynthetic Yield and DMSP Concentrations
4.3. DMSP and Different Symbiodinium Clades
4.4. DMSP as a Thermal Stress Indicator
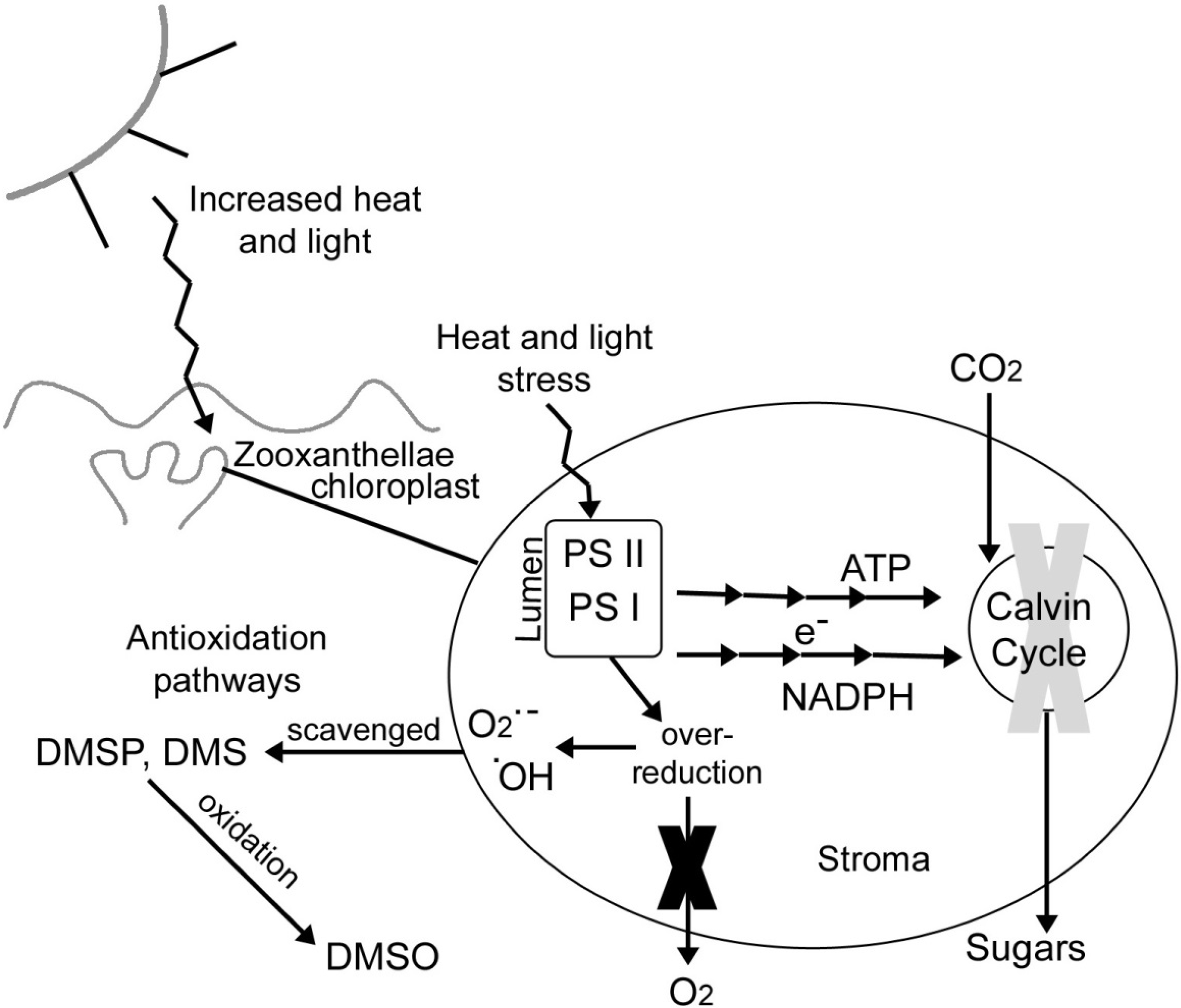
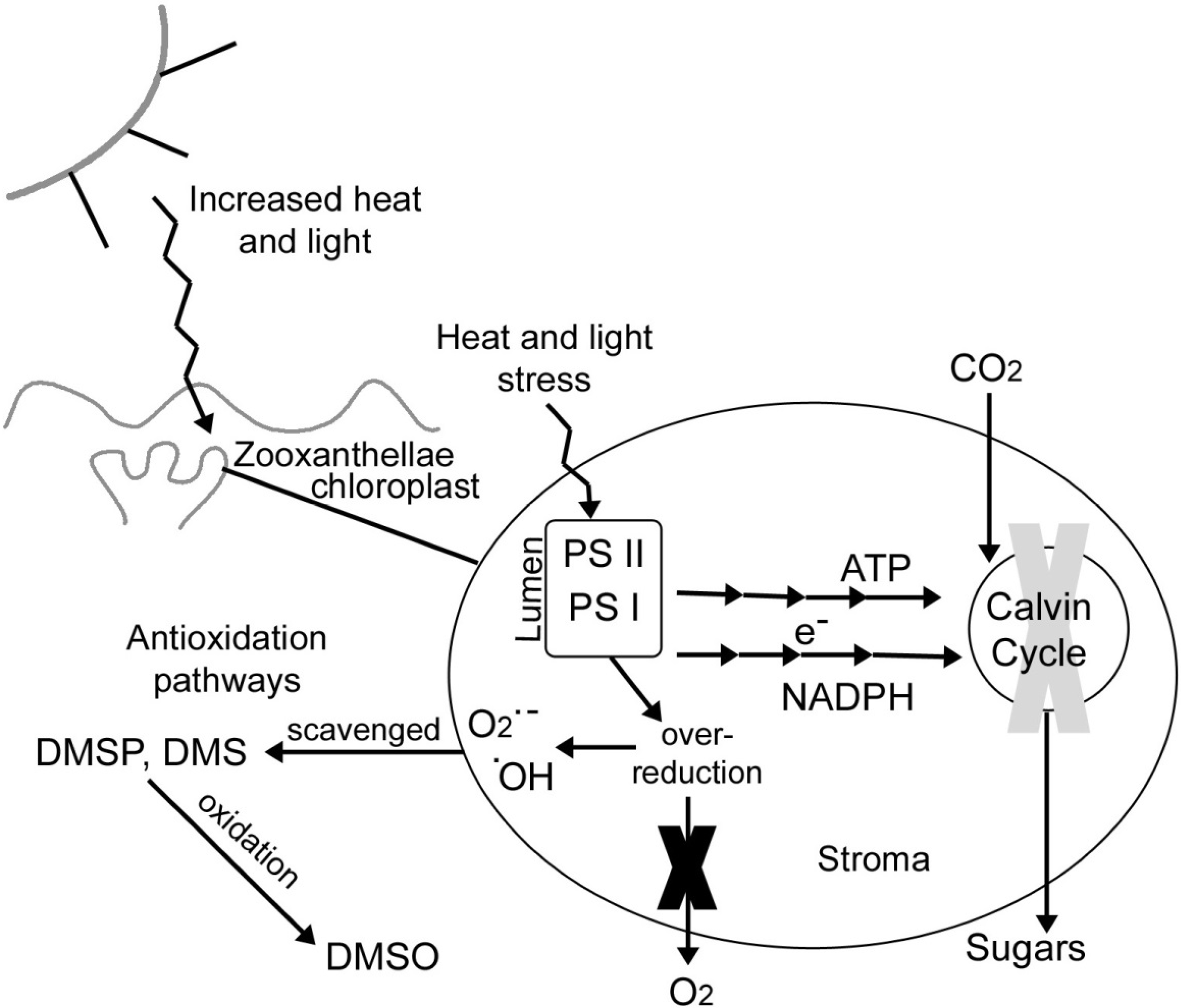
4.5. DMSP as an Antioxidant
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Berkelmans, R.; Oliver, J.K. Large-scale bleaching of corals on the Great Barrier Reef. Coral Reefs 1999, 18, 55–60. [Google Scholar] [CrossRef]
- Harriot, V.J. Mortality rates of scleractinian corals before and during a mass bleaching event. Mar. Ecol. Prog. Ser. 1985, 21, 81. [Google Scholar] [CrossRef]
- Jones, R.J. Changes in zooxanthellar densities and chlorophyll concentrations in corals during and after a bleaching event. Mar. Ecol. Prog. Ser. 1997, 158, 51–59. [Google Scholar] [CrossRef]
- Jones, R.J. Zooxanthellae loss as a bioassay for assessing stress in corals. Mar. Ecol. Prog. Ser. 1997, 149, 163–171. [Google Scholar] [CrossRef]
- Jones, R.J.; Berkelmans, R.; Oliver, J.K. Recurrent bleaching of corals at Magnetic Island (Australia) relative to air and seawater temperature. Mar. Ecol. Prog. Ser. 1997, 158, 289–292. [Google Scholar] [CrossRef]
- Berkelmans, R.; De’ath, G.; Kininmonth, S.; Skirving, W.J. A comparison of the 1998 and 2002 coral bleaching events on the Great Barrier Reef: Spatial correlation, patterns, and predictions. Coral Reefs 2004, 23, 74–83. [Google Scholar] [CrossRef]
- Hoegh-Guldberg, O. Climate change, coral bleaching and the future of the world's coral reefs. Mar. Freshw. Res. 1999, 50, 839–866. [Google Scholar] [CrossRef]
- Brown, B.E.; le Tissier, M.D.A.; Bythell, J.C. Mechanisms of bleaching deduced from histological studies of reef corals sampled during a natural bleaching event. Mar. Biol. 1995, 122, 655–663. [Google Scholar] [CrossRef]
- Douglas, A.E. Coral bleaching-how and why? Mar. Poll. Bull. 2003, 46, 385–392. [Google Scholar] [CrossRef]
- Ahern, H.; Ahmad, S.; Birnbaum, M.R.; Cadenas, E.; Chen, Y.; Cunningham, R.P.; Dalton, D.A.; Felton, G.W.; Grisham, M.B.; Kalyanaraman, B.; et al. Oxidative Stress and Antioxidant Defenses in Biology; Chapman & Hall: New York, NY, USA, 1995. [Google Scholar]
- Fridovich, I. The biology of oxygen radicals. Science 1978, 201, 875–880. [Google Scholar] [CrossRef] [PubMed]
- Dykens, J.A.; Shick, J.M. Oxygen production by endosymbiotic algae controls superoxide dismutase activity in their animal host. Nature 1982, 297, 579–580. [Google Scholar] [CrossRef]
- Franklin, D.J.; Hoegh-Guldberg, O.; Jones, R.J.; Berge’s, J.A. Cell death and degeneration in the symbiotic dinoflagellates of the coral Stylophora pistillata during bleaching. Mar. Ecol. Prog. Ser. 2004, 272, 117–130. [Google Scholar] [CrossRef]
- Lesser, M.P. Elevated temperatures and ultraviolet radiation cause oxidative stress and inhibit photosynthesis in symbiotic dinoflagellates. Limnol. Oceanogr. 1996, 41, 271–283. [Google Scholar] [CrossRef]
- Downs, C.A.; Fauth, J.E.; Halas, J.C.; Dustan, P.; Bemiss, J.; Woodley, C.M. Oxidative stress and seasonal coral bleaching. Free Radic. Biol. Med. 2002, 33, 533–543. [Google Scholar] [CrossRef]
- Jones, R.J.; Hoegh-Guldberg, O.; Larkum, A.W.D.; Schreiber, U. Temperature-induced bleaching of corals begins with impairment of the CO2 fixation mechanism in zooxanthellae. Plant Cell Environ. 1998, 21, 1219–1230. [Google Scholar] [CrossRef]
- Lesser, M.P.; Stochaj, W.R.; Tapley, D.W.; Shick, J.M. Bleaching in coral reef anthozoans: Effect of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 1990, 8, 225–232. [Google Scholar] [CrossRef]
- Rowan, R.; Powers, D.A. Molecular genetic identification of symbiotic dinoflagellates (zooxanthellae). Mar. Ecol. Prog. Ser. 1991, 71, 65–73. [Google Scholar] [CrossRef]
- Loh, W.; Carter, D.; Hoegh-Guldberg, O. Diversity of zooxanthellae from scleractinian corals of One Tree Island (The Great Barrier Reef). In Proceedings of the Australian Coral Reef Society 75th Anniversary Conference, School of Marine Science, University of Queensland, Brisbane, Heron Island, Austrilia, October 1997; Greenwood, J.G., Hall, N.J., Eds.; pp. 141–151.
- Baker, A. Reef corals bleach to survive change. Nature 2001, 411, 765–766. [Google Scholar] [CrossRef] [PubMed]
- Rowan, R.; Knowlton, N.; Baker, A.; Jara, J. Landscape ecology of algal symbionts creates variation in episodes of coral bleaching. Nature 1997, 388, 265–269. [Google Scholar] [CrossRef] [PubMed]
- Berkelmans, R.; van Oppen, M.J.H. The role of zooxanthellae in the thermal tolerance of corals: A “nugget of hope” for coral reefs in an era of climate change. Proc. R. Soc. B 2006, 273, 2305–2312. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.A.; Lam, K.K.; Nakano, Y.; Tsai, W.S. A stable association of the stress-tolerant zooxanthellae, Symbiodinium clade D, with the low-temperature-tolerant coral, Oulastrea crispate (Scleractinia: Faviidae) in subtropical non-reefal coral communities. Zool. Stud. 2003, 42, 540–550. [Google Scholar]
- Buddemeier, R.W.; Fautin, D.G. Coral bleaching as an adaptive mechanism: A testable hypothesis. Bioscience 1993, 43, 320–326. [Google Scholar] [CrossRef]
- Sunda, W.; Keiber, D.J.; Kiene, R.P.; Huntsman, S. An antioxidant function for DMSP and DMS in marine algae. Nature 2002, 418, 317–319. [Google Scholar] [CrossRef] [PubMed]
- Lee, P.A.; de Mora, S.J. Intracellular dimethylsulfoxide (DMSO) in unicellular marine algae: Speculations on its origin and possible biological role. J. Phycol. 1999, 35, 8–18. [Google Scholar] [CrossRef]
- Broadbent, A.D.; Jones, G.B.; Jones, R.J. DMSP in corals and benthic algae from the Great Barrier Reef. Estuar. Coast. Shelf Sci. 2002, 55, 547–555. [Google Scholar] [CrossRef]
- Hill, R.W.; Dacey, J.W.H.; Krupp, D.A. Dimethylsulfoniopropionate in Reef Corals. B Mar. Sci. 1995, 57, 489–494. [Google Scholar]
- Jones, G.B.; Curran, M.A.J.; Broadbent, A.D. Dimethylsulphide in the South Pacific. In Recent Advances in Marine Science and Technology; Bellwood, O., Choat, H., Saxena, N., Eds.; Pacon International and James Cook University of North Queensland: Townsville, Australia, 1994; pp. 183–190. [Google Scholar]
- Broadbent, A.D.; Jones, G.B. DMS and DMSP in mucus ropes, coral mucus, surface films and sediment pore waters from coral reefs in the Great Barrier Reef. Mar. Freshw. Res. 2004, 55, 849–855. [Google Scholar] [CrossRef]
- Deschaseaux, E.S.M.; Jones, G.B.; Deseo, M.A.; Shepherd, K.M.; Kiene, R.P.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Effects of environmental factors on dimethylated sulfur compounds and their potential role in the antioxidant system of the coral holobiont. Limnol. Oceanogr. 2014, 59, 758–768. [Google Scholar] [CrossRef]
- Fischer, E.; Jones, G.B. Atmospheric dimethysulphide production from corals in the Great Barrier Reef and links to solar radiation, climate and coral bleaching. Biogeochemistry 2012, 110, 31–46. [Google Scholar] [CrossRef]
- Jones, G.B. The ethnobiology of coral and coral reefs. In The Reef Sulphur Cycle: Influence on Climate and Ecosystem Services; Narchi, N., Price, L., Eds.; Springer: Berlin, Germany, 2015. (in press) [Google Scholar]
- Jones, G.B.; Trevena, A.J. The influence of coral reefs on atmospheric dimethylsulphide over the Great Barrier Reef, Coral Sea, Gulf of Papua, Solomon and Bismarck Seas. Mar. Freshw. Res. 2005, 56, 85–93. [Google Scholar] [CrossRef]
- Modini, R.L.; Ristovski, Z.D.; Johnson, G.R.; He, C.; Surawski, N.; Morawska, L.; Suni, T.; Kulmala, M. New particle formation and growth at a remote, sub-tropical coastal location. Atmos. Chem. Phys. Discuss. 2009, 9, 12101–12139. [Google Scholar] [CrossRef]
- Swan, H.; Jones, G.; Deschaseaux, E. Dimethylsulfide and coral reef ecosystems. In Proceedings of the 12th International Coral Reef Symposium, Cairns, Australia, 9–13 July 2012.
- Raina, J.B.; Tapiolas, D.M.; Forêt, S.; Lutz, A.; Abrego, D.; Ceh, J.; Seneca, F.O.; Clode, P.L.; Bourne, D.G.; Willis, B.L.; et al. DMSP biosynthesis by an animal and its role in coral thermal stress response. Nature 2013, 502, 677–680. [Google Scholar] [CrossRef] [PubMed]
- Deschaseaux, E.S.M.; Beltran, V.H.; Jones, G.B.; Deseo, M.A.; Swan, H.B.; Harrison, P.L.; Eyre, B.D. Comparative response of DMS and DMSP concentrations in Symbiodinium clades C1 and D1 under thermal stress. J. Exp. Mar. Biol. Ecol. 2014, 459, 181–189. [Google Scholar] [CrossRef]
- Jones, G.B.; Fischer, E.; Deschaseaux, E.S.M.; Harrison, P.L. The effect of coral bleaching on the cellular concentration of dimethylsulphoniopropionate in reef corals. J. Exp. Mar. Biol. Ecol. 2014, 460, 19–31. [Google Scholar] [CrossRef]
- Berkelmans, R. Time-integrated thermal bleaching thresholds of reefs and their variation on the Great Barrier Reef. Mar. Ecol. Prog. Ser. 2002, 229, 73–82. [Google Scholar] [CrossRef]
- Turner, P.; Berkelmans, R.; Brodie, M. Precise set-point control of temperature for coral bleaching experiments. Mar. Technol. Soc. J. 2002, 36, 70–75. [Google Scholar] [CrossRef]
- Ulstrup, K.E.; van Oppen, M.J.H. Geographic and habitat partitioning of genetically distinct zooxanthellae (Symbiodinium) in Acropora corals on the Great Barrier Reef. Mol. Ecol. 2003, 12, 3477–3484. [Google Scholar] [CrossRef] [PubMed]
- Van Oppen, M.J.H.; Palstra, F.P.; Piquet, A.M.-T.; Miller, D.J. Patterns of coral-dinoflagellate associations in Acropora: Significance of local availability and physiology of Symbiodinium strands and host-symbiont selectivity. Proc. R. Soc. Lond. B 2001, 268, 1759–1767. [Google Scholar] [CrossRef] [PubMed]
- Fabricus, K.E.; Mieog, J.C.; Colin, P.L.; Idip, D.; van Oppen, M.J.H. Identity and diversity of coral endosymbionts (zooxanthellae) from three Palauan reefs with contrasting bleaching, temperature and shading histories. Mol. Ecol. 2004, 13, 2445–2458. [Google Scholar] [CrossRef] [PubMed]
- Stimson, J.; Kinzie, R.A., III. The temporal pattern and rate of release of zooxanthellae from the reef coral Pocillopora damicornis (Linnaeus) under nitrogen enrichment and control conditions. J. Exp. Mar. Biol. Ecol. 1991, 153, 63–74. [Google Scholar] [CrossRef]
- Curran, M.A.J.; Jones, G.B.; Burton, H. Spatial distribution of DMS and DMSP in the Australasian sector of the Southern Ocean. In: Special Issue on the Southern Hemisphere Marine Aerosol Characterisation Experiment (ACE 1). J. Geophys. Res. 1998, 103, 16677–16689. [Google Scholar] [CrossRef]
- Tapiolas, D.M.; Raina, J.B.; Lutz, A.; Willis, B.L.; Motti, C.A. Direct measurement of dimethylsulphoniopropionate (DMSP) in reef-building corals using quantitative nuclear magnetic resonance (qNMR) spectroscopy. J. Exp. Mar. Biol. Ecol. 2013, 443, 85–89. [Google Scholar] [CrossRef]
- Stimson, J.; Sakai, K.; Sembali, H. Interspecific comparison of the symbiotic relationship in corals with high and low rates of bleaching-induced mortality. Coral Reefs 2002, 21, 409–421. [Google Scholar]
- Strychar, K.B.; Coates, M.; Sammarco, P.W. Loss of Symbiodinium from bleached Australian scleractinian corals (Acropora hyacinthus, Favites complanata and Porites solida). Mar. Freshw. Res. 2004, 55, 135–144. [Google Scholar] [CrossRef]
- Ralph, P.J.; Gademann, R.; Larkum, A.W.D. Zooxanthellae expelled from bleached corals at 33 degrees C are photosynthetically competent. Mar. Ecol. Prog. Ser. 2001, 220, 163–168. [Google Scholar] [CrossRef]
- Jones, R.J.; Hoegh-Guldberg, O. Diurnal changes in the photochemical efficiency of the symbiotic dinoflagellates (Dinophyceae) of corals: Photoprotection, photo-inactivation and the relationship to coral bleaching. Plant Cell Environ. 2001, 24, 89–99. [Google Scholar] [CrossRef]
- Andreae, M.O. Ocean-atmosphere interactions in the global biogeochemical sulphur cycle. Mar. Chem. 1990, 30, 1–29. [Google Scholar] [CrossRef]
- Heckathorn, S.A.; Downs, C.A.; Sharkey, T.D.; Coleman, J.S. The small, methionine-rich chloroplast heat-shock protein protects photosystem 11 electron transport during heat stress. Plant Physiol. 1998, 116, 439–444. [Google Scholar] [CrossRef] [PubMed]
- Muslim, I.; Jones, G.B. The seasonal variation of dissolved nutrients, chlorophyll a and suspended sediments at Nelly Bay, Magnetic Island. Estuar. Coast. Shelf Sci. 2003, 57, 445–455. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jones, G.B.; King, S. Dimethylsulphoniopropionate (DMSP) as an Indicator of Bleaching Tolerance in Scleractinian Corals. J. Mar. Sci. Eng. 2015, 3, 444-465. https://doi.org/10.3390/jmse3020444
Jones GB, King S. Dimethylsulphoniopropionate (DMSP) as an Indicator of Bleaching Tolerance in Scleractinian Corals. Journal of Marine Science and Engineering. 2015; 3(2):444-465. https://doi.org/10.3390/jmse3020444
Chicago/Turabian StyleJones, Graham B., and Stacey King. 2015. "Dimethylsulphoniopropionate (DMSP) as an Indicator of Bleaching Tolerance in Scleractinian Corals" Journal of Marine Science and Engineering 3, no. 2: 444-465. https://doi.org/10.3390/jmse3020444
APA StyleJones, G. B., & King, S. (2015). Dimethylsulphoniopropionate (DMSP) as an Indicator of Bleaching Tolerance in Scleractinian Corals. Journal of Marine Science and Engineering, 3(2), 444-465. https://doi.org/10.3390/jmse3020444