
Cytotoxic Effects of Vicicitus globosus (Class Dictyochophyceae) and Chattonella marina (Class Raphidophyceae) on Rotifers and Other Microalgae

Abstract
:1. Introduction
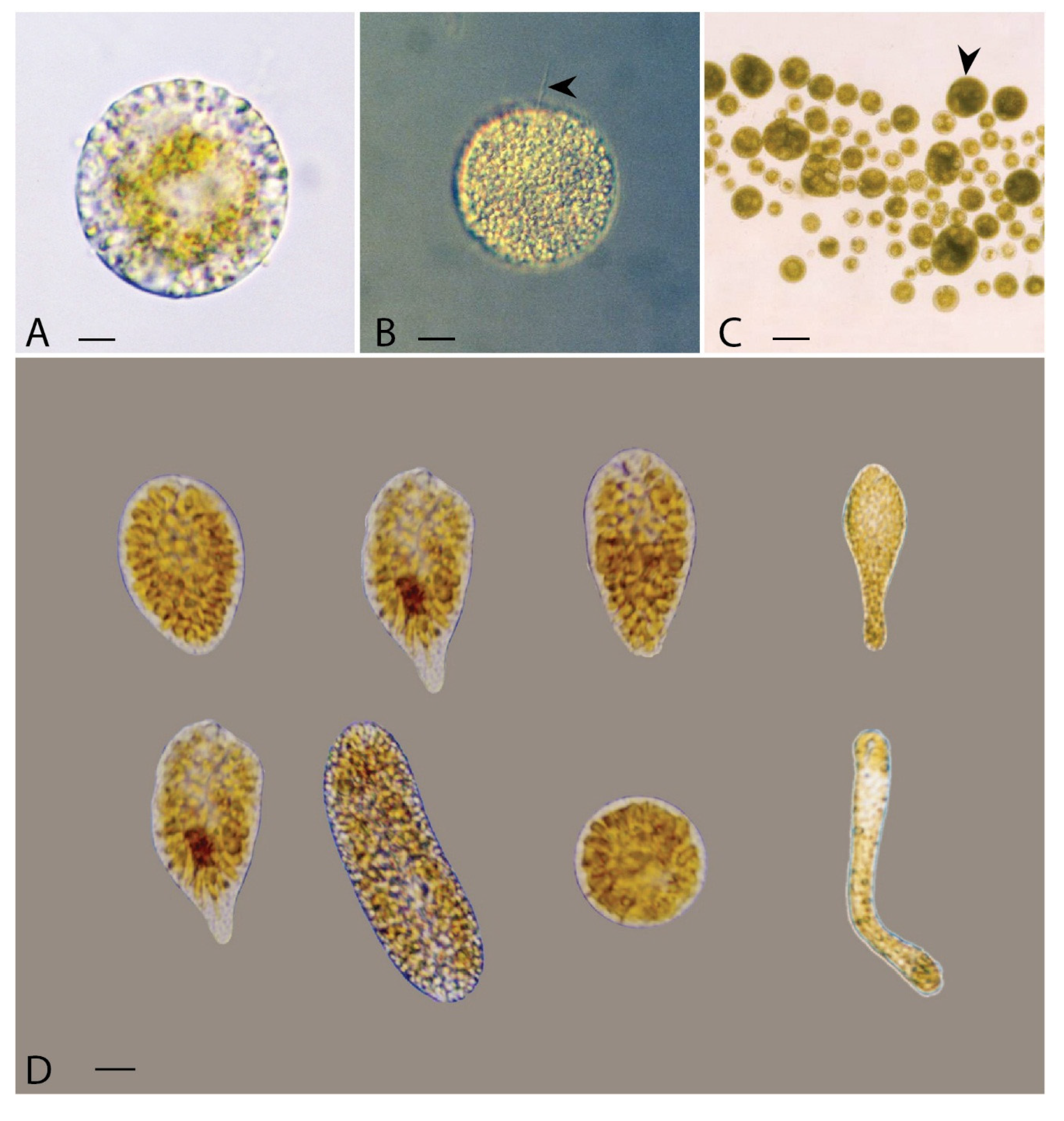
2. Materials and Methods
2.1. Cultures and Growth Conditions
2.2. Cell Extraction and Cytotoxicity Tests

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Lt50 | ||
|---|---|---|
| Taxa | VgTx | CmTx |
| Class Dinophyceae | ||
| Alexandrium catenella | 11 min | 25 min |
| Karenia concordia | 10 min | 20 min |
| Karenia mikimotoi | 12 min | 22 min |
| Class Dictyochophyceae | ||
| Vicicitus globosus | 7 min | 30 min |
| Class Raphidophyceae | ||
| Chattonella marina | 10 min | 25 min |
| Class Cryptophyceae | ||
| Rhodomonas sp. | 5 min | 16 min |
| Class Bacillariophyceae | ||
| Ditylum brightwellii | 9 min | 18 min |
| Lauderia annulata | 12 min | 22 min |
| Chaetoceros sp. | 11 min | 20 min |
| Rotifers | 80 min | 20 h |
3. Results
3.1. Effects of Lipophilic Extracts of Vicicitus globosus and Chattonella marina on Microalgal Cells
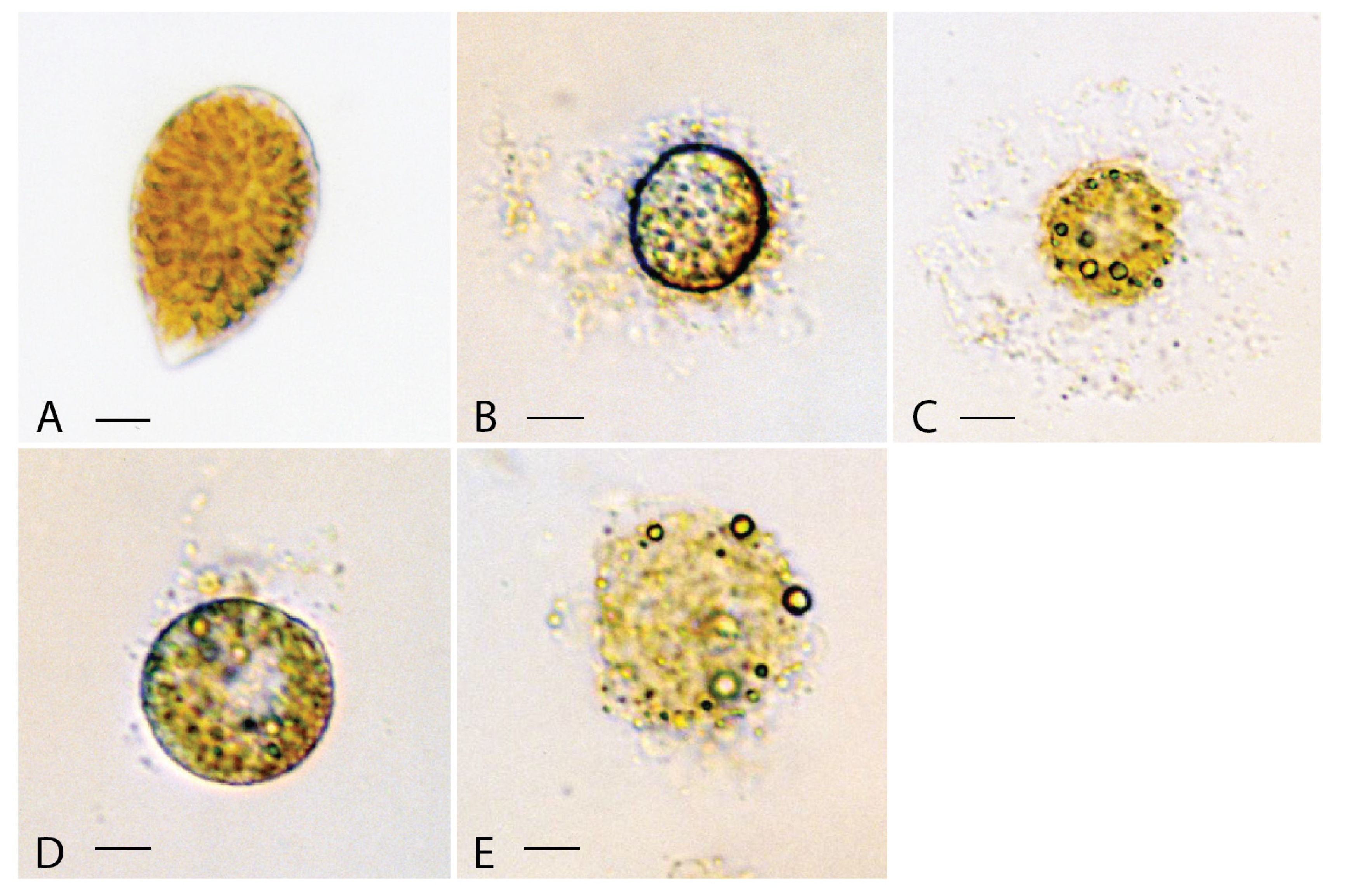
3.1.1. Alexandrium catenella
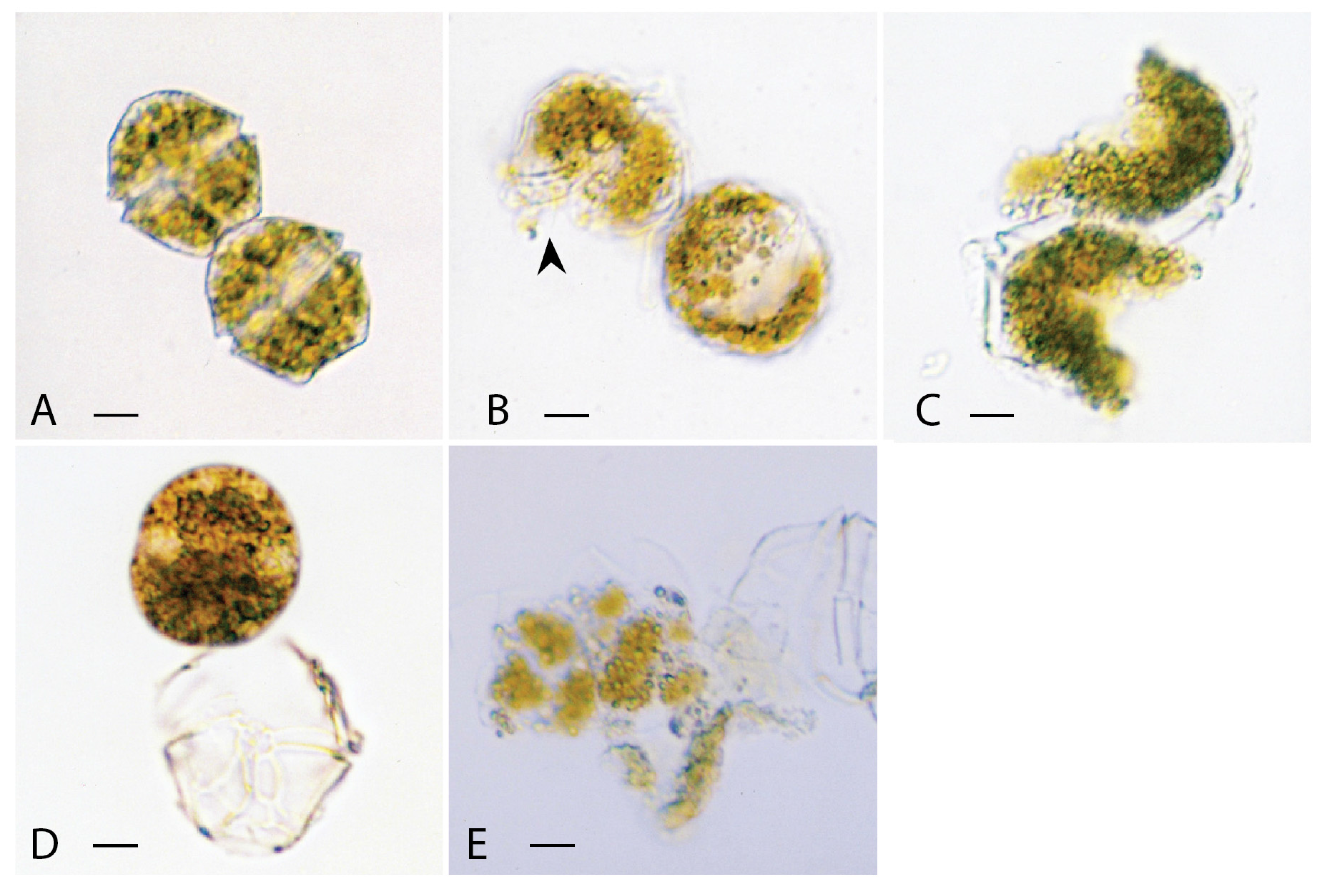
3.1.2. Chattonella marina
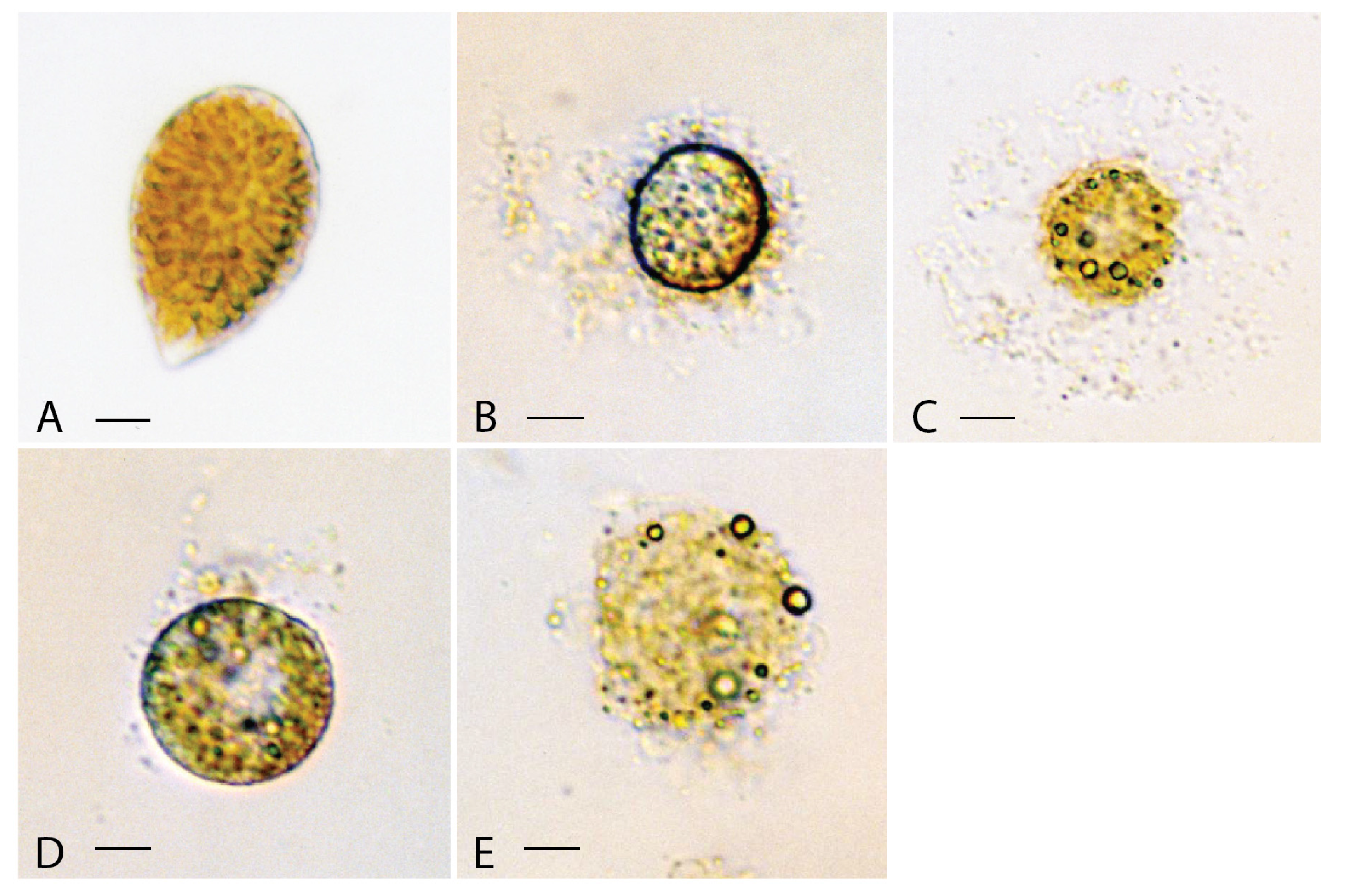
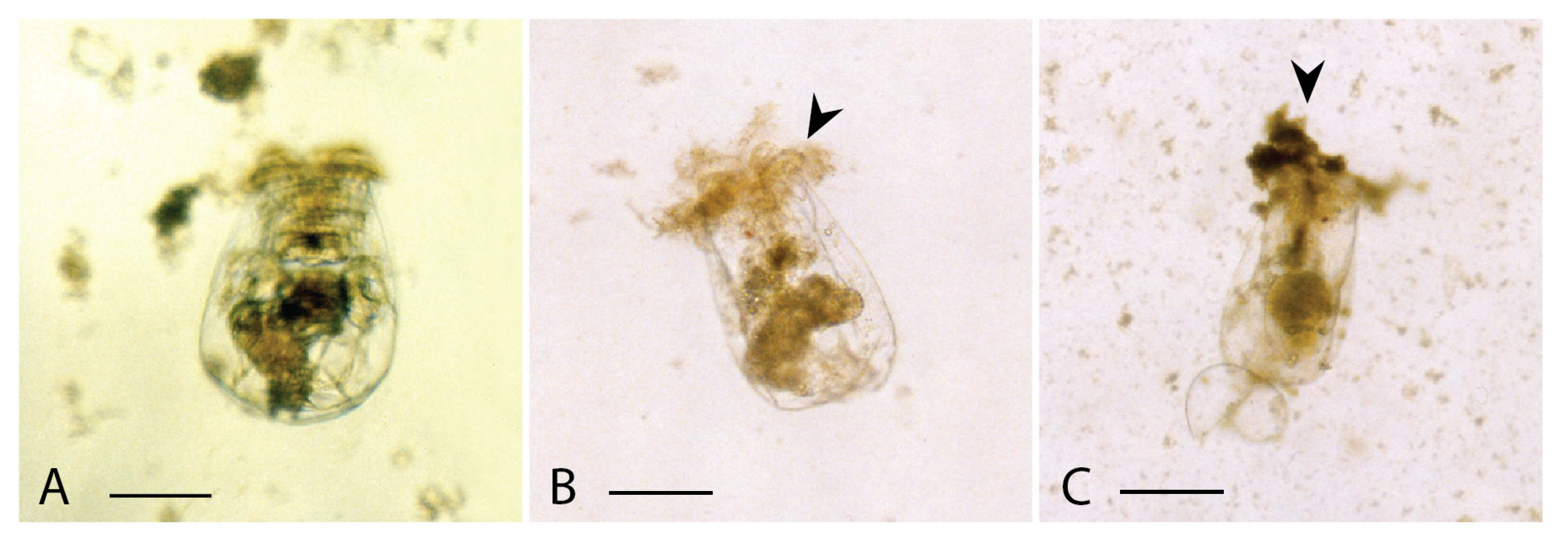
3.2. Cytotoxicity on Rotifers
4. Discussion
Acknowledgments
Conflicts of Interest
References
- Chang, F.H.; McVeagh, M.; Gall, M.; Smith, P. Chattonella globosus is a member of Dictyochophyceae: Reassignment of Vicicitus gen. nov., based on molecular phylogeny, pigment composition, morphology and life history. Phycologia 2012, 51, 403–420. [Google Scholar]
- Chang, F.H.; Mullan, A.B. Extended blooms of Karenia concordia and other harmful algae from 2009 to 2011 in Wellington Harbour, New Zealand. In Harmful Algae 2012, Proceedings of the 15th ICHA, 29 October–2 November 2012; Kim, H.G., Reguera, B., Hallegraeff, G.M., Lee, C.K., Han, M.S., Choi, J.K., Eds.; International Society for the Study of Harmful Algae: Changwon, Korea, 2014. [Google Scholar]
- Akizuki, Y.; Kitakado, I.; Sasaki, M. Globular type of flagellates occurring at the early stage of Hornellia blooms. In Report of Previous Conjecture and Research of Red Tides in 1979; Seto Inland Sea Block: Seto, Japan, 1981; pp. 201–204. [Google Scholar]
- Hosaka, M.; Takayama, N.; Hirai, S.; Hoda, M.; Hara, Y. The occurrence of raphidophycean alga Chattonella sp. (globular type) in Tokyo Bay, Japan. Bull. Plankton Soc. Jpn. 1991, 38, 1–8. [Google Scholar]
- Lu, S.H.; Hodgkiss, I.J. More raphidophyte blooms in South China waters. Harmful Algae News 2001, 22, 1–2. [Google Scholar]
- Hallegraeff, G.M.; Hara, Y. Taxonomy of harmful marine raphidophytes. In Manual of Harmful Marine Microalgae; Hallegraeff, G.M., Anderson, D.M., Cembella, A., Eds.; UNESCO Publishing: Paris, France, 2003; pp. 511–522. [Google Scholar]
- Ignatiades, L.; Gotsis-Skretas, O. A review on toxic and harmful algae in Greek coastal waters (E. Mediterranean Sea). Toxins 2010, 2, 1019–1037. [Google Scholar] [CrossRef] [PubMed]
- Simakova, N.V.; Orlova, T.Y.; Selina, M.S. Red tide caused by raphidophytes Chattonella sp. in Amurskii Bay, the Sea of Japan. Russ. J. Mar. Biol. 1990, 16, 77–79. [Google Scholar]
- Mafra Junior, L.L.; Fernades, L.F.; Proenca, L.A.O. Harmful algae and toxin in Paranaguá Bay, Brazil: Bases for monitoring. Braz. J. Oceanogr. 2006, 54, 1–49. [Google Scholar] [CrossRef]
- Tiffany, M.A.; Barlow, S.B.; Matey, V.E.; Hurlbert, S.H. Chattonella marina (Raphidophyceae), a potentially toxic alga in the Salton Sea, California. Hydrobiologia 2001, 466, 187–194. [Google Scholar] [CrossRef]
- Elbraechter, M. Exotic flagellates of coastal North Sea waters. Helgol. Meeresunters. 1998, 52, 235–242. [Google Scholar] [CrossRef]
- Sako, Y.; Otake, I.; Uchida, A. The harmful algae Chattonella antiqua, C. marina and C. ovata (Raphidophyceae). Phylogenetically the same species. In Proceedings of the HAB 2000, IX International Conference on Harmful Algal Blooms, Hobart, Tasmania, Australia, 7–11 September 2000; p. 213.
- Bowers, H.A.; Tomas, C.; Tengs, T.; Kepton, J.W.; Lewitus, A.J.; Oldach, D.W. Raphidophyceae (Chadefaud ex Silva) systematics and rapid identification: Sequence analyses and real time PCR assays. J. Phycol. 2006, 42, 1333–1348. [Google Scholar] [CrossRef] [PubMed]
- Hosoi-Tanabe, S.; Otake, I.; Sako, Y. Phylogenetic analysis of noxious red tide flagellates Chattonella antiqua, C. marina, C. ovata, and C. verruculosa (Raphidophyceae) based on the rRNA gene family. Fish. Sci. 2006, 72, 1200–1208. [Google Scholar]
- Demura, M.; Noel, M.-H.; Kasao, F.; Watanabe, M.M.; Kawachi, M. Taxonomic revision of Chattonella antiqua, C. marina and C. ovata (Raphidophyceae) based on their morphological characteristics and genetic diversity. Phycologia 2009, 48, 518–535. [Google Scholar]
- Hiroshi, S.; Okada, H.; Imai, I.; Yoshida, T. High toxicity of the novel bloom-forming species Chattonella ovata (Raphidophyceae) to cultured fish. Harmful Algae 2005, 4, 783–787. [Google Scholar] [CrossRef]
- Okaichi, T. Marine environmental studies on outbreaks of red tides in neritic waters. J. Oceanogr. Soc. Jpn. 1983, 39, 267–278. [Google Scholar] [CrossRef]
- Imai, I.; Itoh, K. Annual life cycle of Chattonella spp., causative flagellates of noxious red tides in the Inland Sea of Japan. Mar. Biol. 1987, 94, 287–292. [Google Scholar]
- Marshall, J.-A.; Nichols, P.D.; Hamilton, B.; Lewis, R.J.; Hallegraeff, G.M. Ichthyotoxicity of Chattonella marina (Raphidophyceae) to damselfish (Acanthochromis polycanthus): The synergistic role of reactive oxygen species and free fatty acids. Harmful Algae 2003, 2, 273–281. [Google Scholar] [CrossRef]
- Onoue, Y.; Nozawa, K. Separation of toxins from harmful red tides occuring alone the coast of Kagoshima Prefecture. In Red Tides: Biology, Environmental Science and Toxicology; Okaichi, T., Anderson, D.M., Nemoto, T., Eds.; Elseier: New York, NY, USA, 1989; pp. 371–374. [Google Scholar]
- Khan, S.; Ahmed, M.S.; Arakawa, O.; Onoue, Y. Properties of neurotoxins separated from harmful red tide organism Chattonella marina. Israeli J. Aquac.–Bamidgeh 1995, 47, 137–141. [Google Scholar]
- McNabb, P.; Rhodes, L.; Adamson, J.; Holland, P. Brevetoxin–An elusive toxin in New Zealand waters. Afr. J. Mar. Sci. 2006, 28, 375–377. [Google Scholar] [CrossRef]
- Endo, M.; Onoue, Y.; Kuroki, A. Neurotoxin-induced cardiac disorder and its role in the death of fish exposed to Chattonella marina. Mar. Biol. 1992, 112, 371–376. [Google Scholar] [CrossRef]
- Shimada, M.; Murakami, T.H.; Imahayashi, T.; Ozaki, H.S.; Toyoshima, T.; Okaichi, T. Effects of sea bloom, Chattonella antiqua, on gill primary lamellae of the young yellowtail, Seriola quinqueradiatta. Acta Histochem. Cytochem. 1983, 16, 232–244. [Google Scholar] [CrossRef]
- Oda, Y.; Nakamura, A.; Shimada, M.; Kawano, I.; Ishimatsu, A.; Muramatsu, T. Generation of reactive oxyen species by raphidophycean phytoplankton. Biosci. Biotechnol. Biochem. 1997, 61, 1658–1662. [Google Scholar] [CrossRef] [PubMed]
- Shen, M.; Xu, J.; Tsang, T.Y.; Au, D.W.T. Toxicity comparison between Chattonella marina and Karenia brevis using marine medaka (Oryzias melastigma): Evidence against the suspected ichthyotoxins of Chattonella marina. Chemosphere 2010, 80, 585–591. [Google Scholar] [CrossRef] [PubMed]
- Chang, F.H.; Sutherland, J.E.; McVeagh, M.; Gall, M. Molecular phylogeny, pigment composition, toxicology and life history of Pseudochattonella cf. verruculosa (Class Dictyochophyceae) from Wellington Harbour, New Zealand. Harmful Algae 2014, 34, 42–55. [Google Scholar]
- Blackburn, S.I.; Hallegraeff, G.M.; Bolch, C.J. Vegetative reproduction and sexual life cycle of the toxic dinoflagellate Gymnodinioum catenatum from Tasmania. Aust. J. Phycol. 1989, 25, 577–590. [Google Scholar] [CrossRef]
- Chang, F.H. Toxic effects of three closely-related dinoflagellates, Karenia concordia, K. brevisulcata and K. mikimotoi (Gymnodiniales, Dinophyceae) on other microalgal species. Harmful Algae 2010, 10, 81–187. [Google Scholar]
- Chang, F.H.; Gall, M. Pigment compositions and toxic effects of three harmful Karenia species, Karenia concordia, Karenia brevisulcata and Karenia mikimotoi (Gymnodiniales, Dinophyceae), on rotifers and brine shrimps. Harmful Algae 2013, 27, 113–120. [Google Scholar] [CrossRef]
- Dafni, Z.; Shilo, M. The cytotoxic principle of the phytoflagellate Prymnesium parvum. J. Cell Biol. 1966, 28, 464–471. [Google Scholar] [CrossRef]
- Gentien, P.; Arzul, G. Exotoxin production by Gyrodinium cf. aureolum (Dinophyceae). J. Mar. Biol. Assoc. UK 1990, 70, 571–581. [Google Scholar] [CrossRef]
- Deeds, J.R. Toxins and Toxicity from the Cosmopolitan, Bloom-Forming Dinoflagellate Karlodinium micrum. Ph.D. Thesis, University of Maryland, Centre of Marine Biotechnology, College Park, MD, USA, 2003; p. 247. [Google Scholar]
- Place, A.R.; Bowers, H.A.; Bachvaroff, T.R.; Adolf, J.E.; Deeds, J.R.; Sheng, J. Karlodinium veneficum—The little dinoflagellate with a big bite. Harmful Algae 2012, 14, 179–195. [Google Scholar] [CrossRef]
- Shilo, M. Toxins of Chrysophyceae. In Microbial Toxins; Kadis, S., Ciegler, A., Ajl, S.J., Eds.; Academic Press: New York, USA, 1980; Volume 7, pp. 67–103. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, F.H. Cytotoxic Effects of Vicicitus globosus (Class Dictyochophyceae) and Chattonella marina (Class Raphidophyceae) on Rotifers and Other Microalgae. J. Mar. Sci. Eng. 2015, 3, 401-411. https://doi.org/10.3390/jmse3020401
Chang FH. Cytotoxic Effects of Vicicitus globosus (Class Dictyochophyceae) and Chattonella marina (Class Raphidophyceae) on Rotifers and Other Microalgae. Journal of Marine Science and Engineering. 2015; 3(2):401-411. https://doi.org/10.3390/jmse3020401
Chicago/Turabian StyleChang, Fook Hoe. 2015. "Cytotoxic Effects of Vicicitus globosus (Class Dictyochophyceae) and Chattonella marina (Class Raphidophyceae) on Rotifers and Other Microalgae" Journal of Marine Science and Engineering 3, no. 2: 401-411. https://doi.org/10.3390/jmse3020401
APA StyleChang, F. H. (2015). Cytotoxic Effects of Vicicitus globosus (Class Dictyochophyceae) and Chattonella marina (Class Raphidophyceae) on Rotifers and Other Microalgae. Journal of Marine Science and Engineering, 3(2), 401-411. https://doi.org/10.3390/jmse3020401