


Effects of the Ionic Liquid [BMIM]Cl on the Baltic Microphytobenthic Communities



Abstract
:1. Introduction
2. Materials and Methods
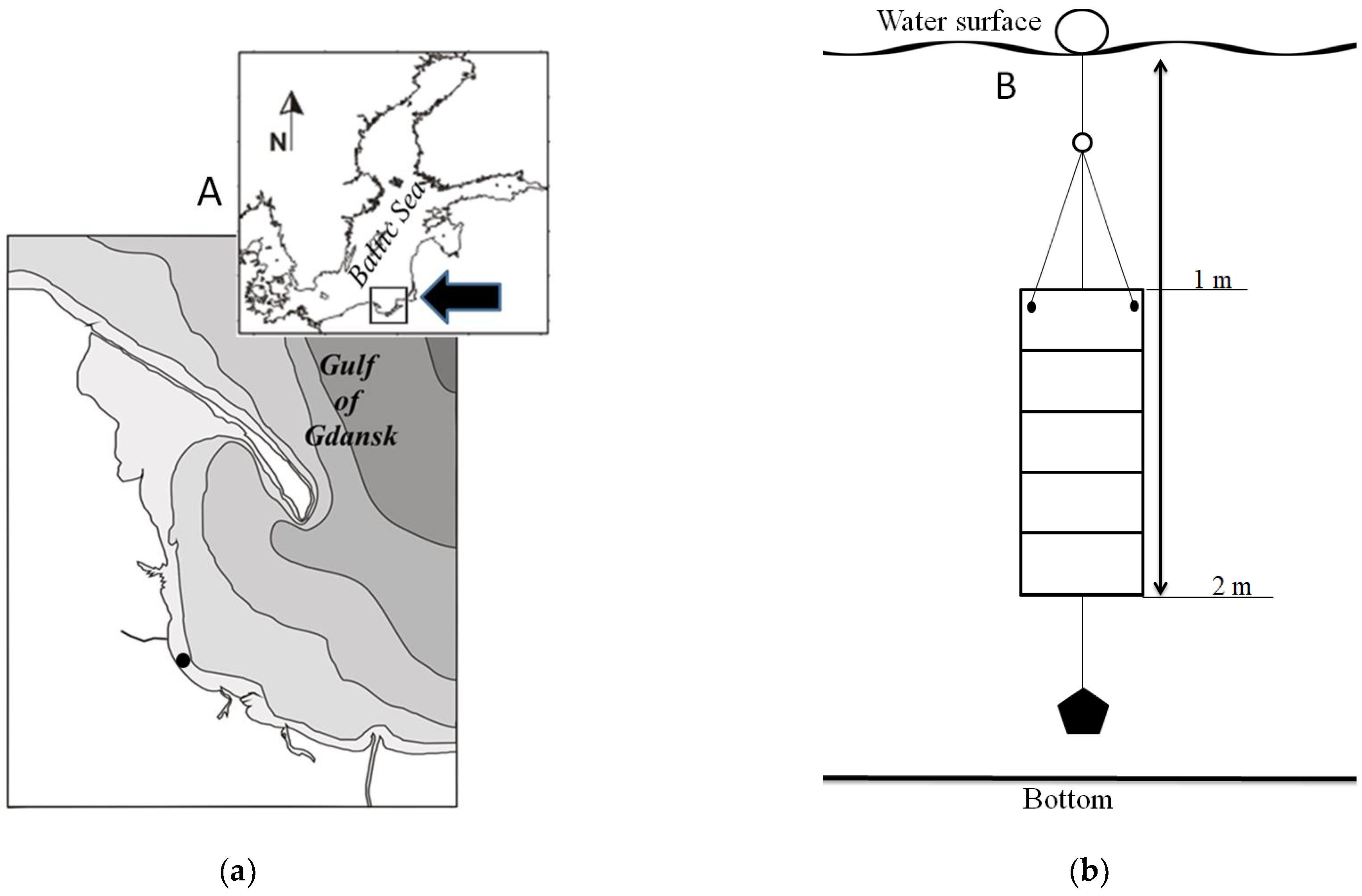
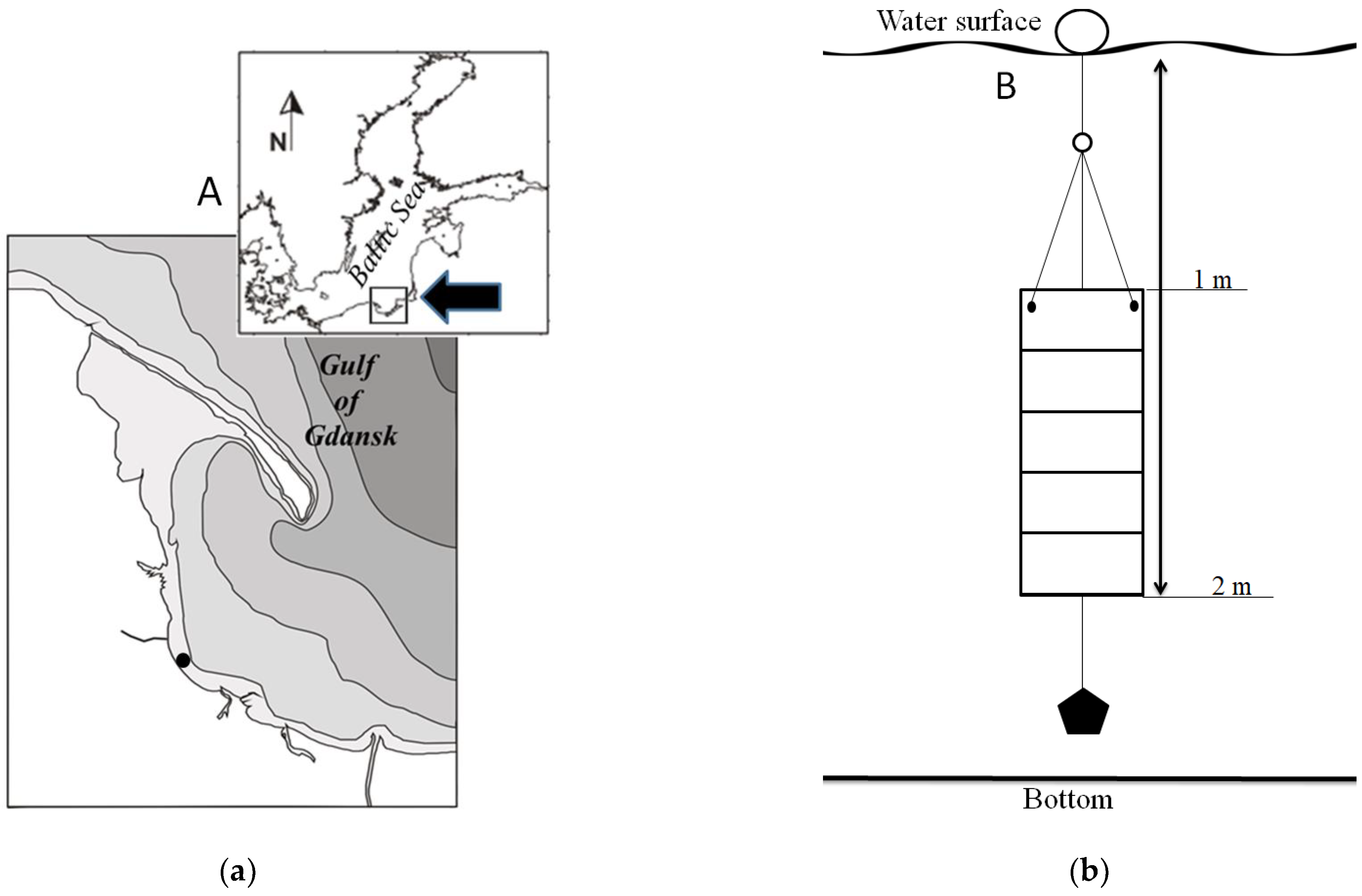
2.1. Study Area and Field Works
2.2. Microalgal Material Preparation Procedure and Experimental Design
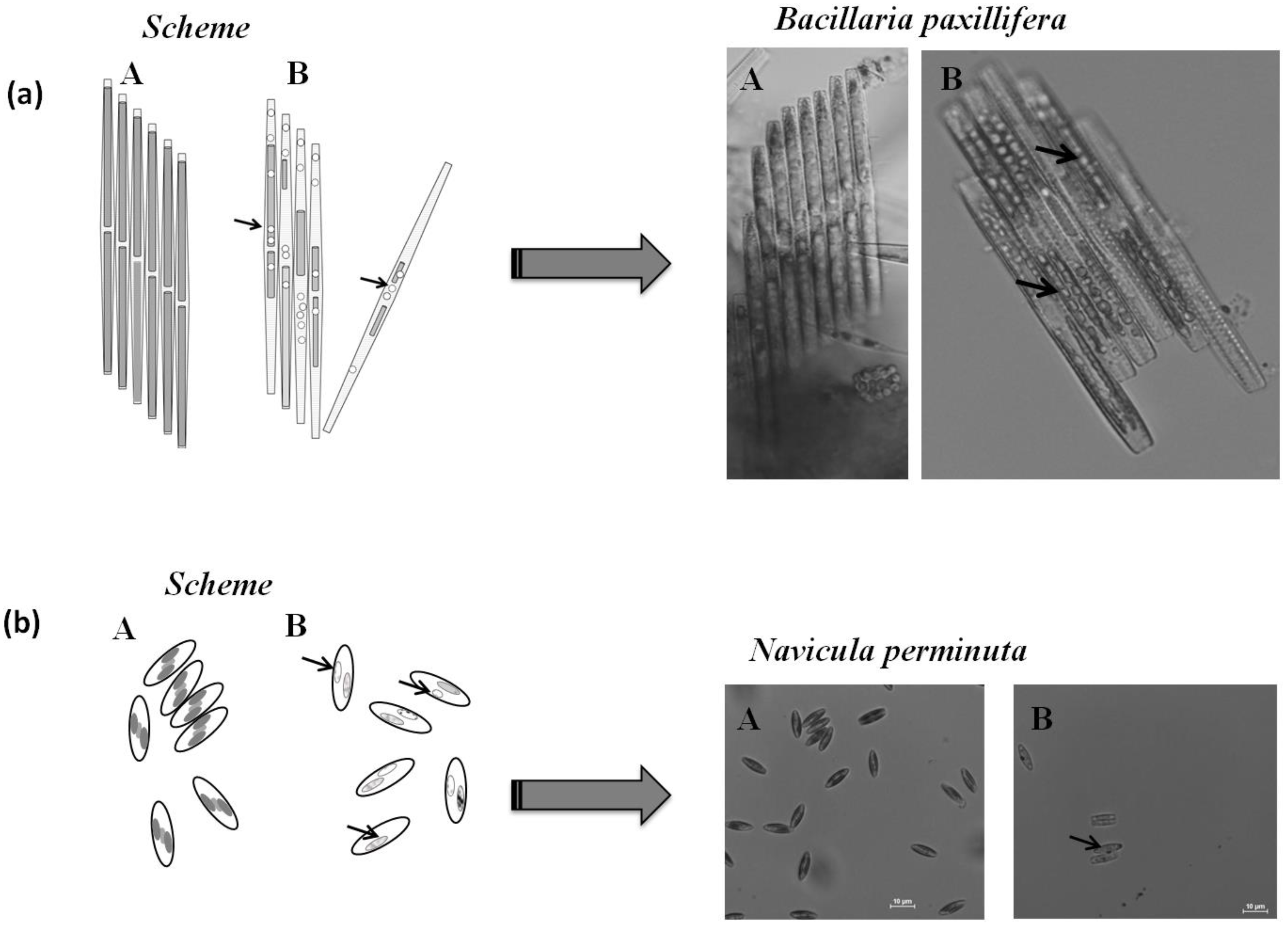
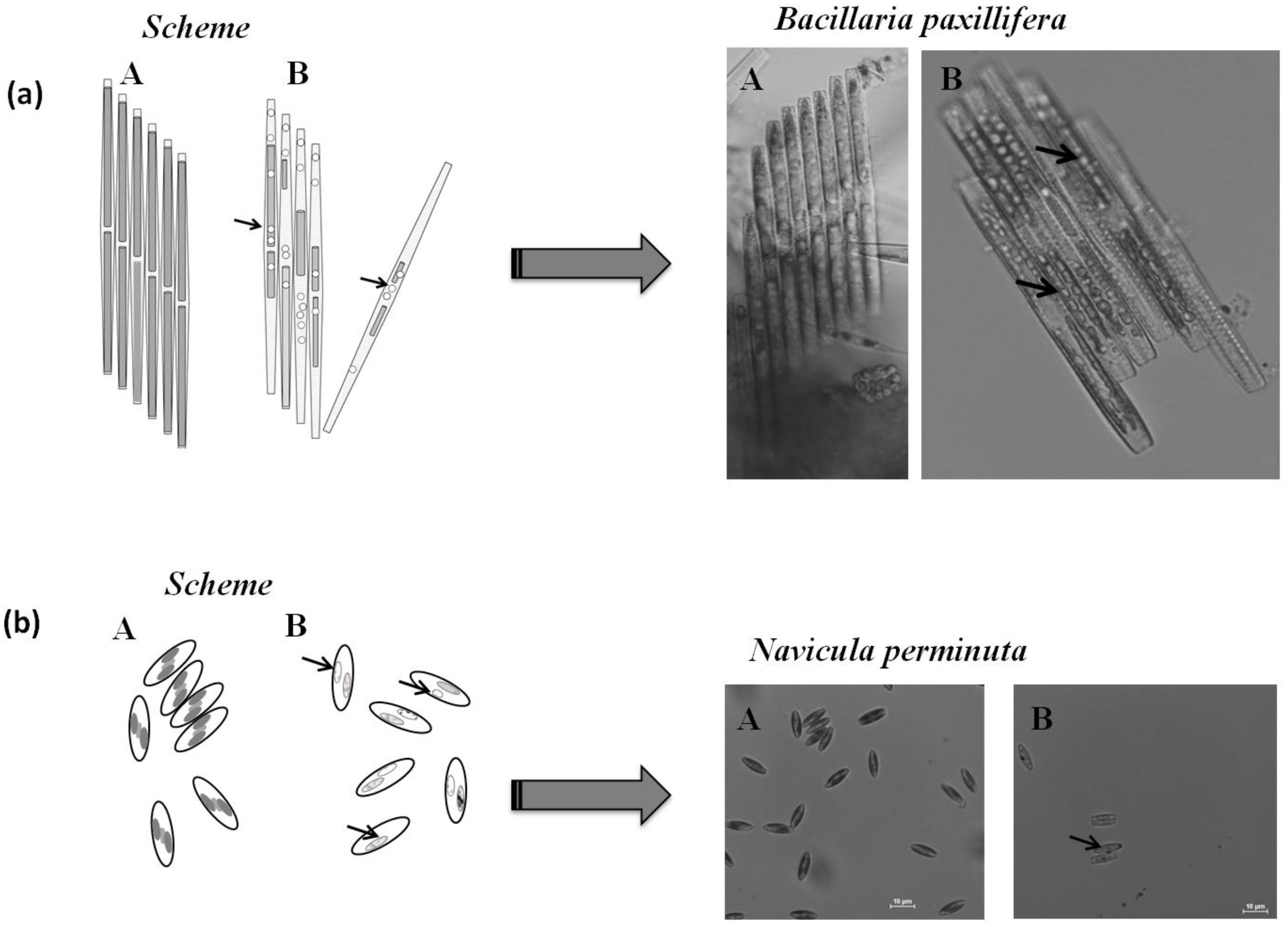
2.3. Microscopic Analysis
2.4. Statistical Analysis
3. Results
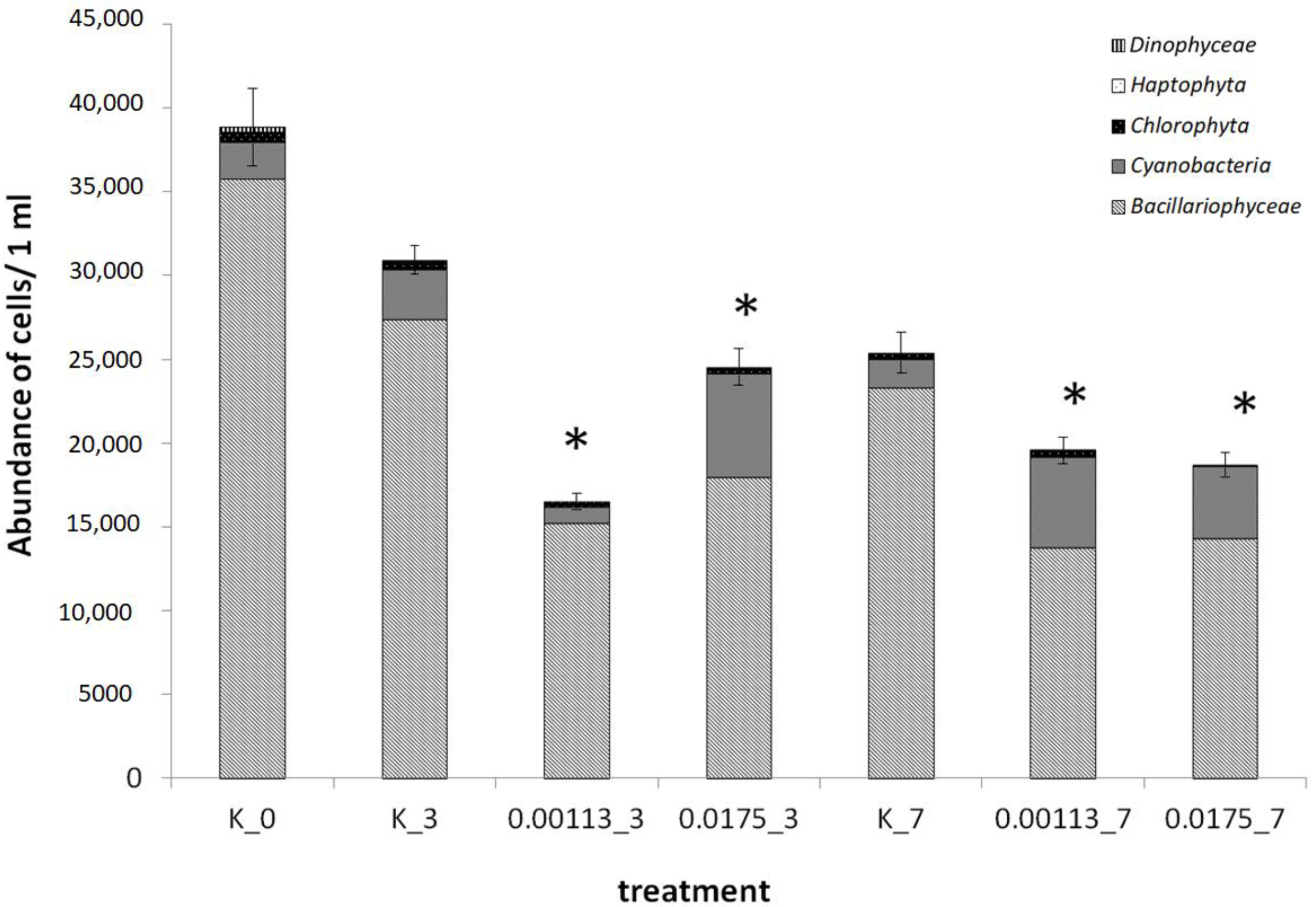
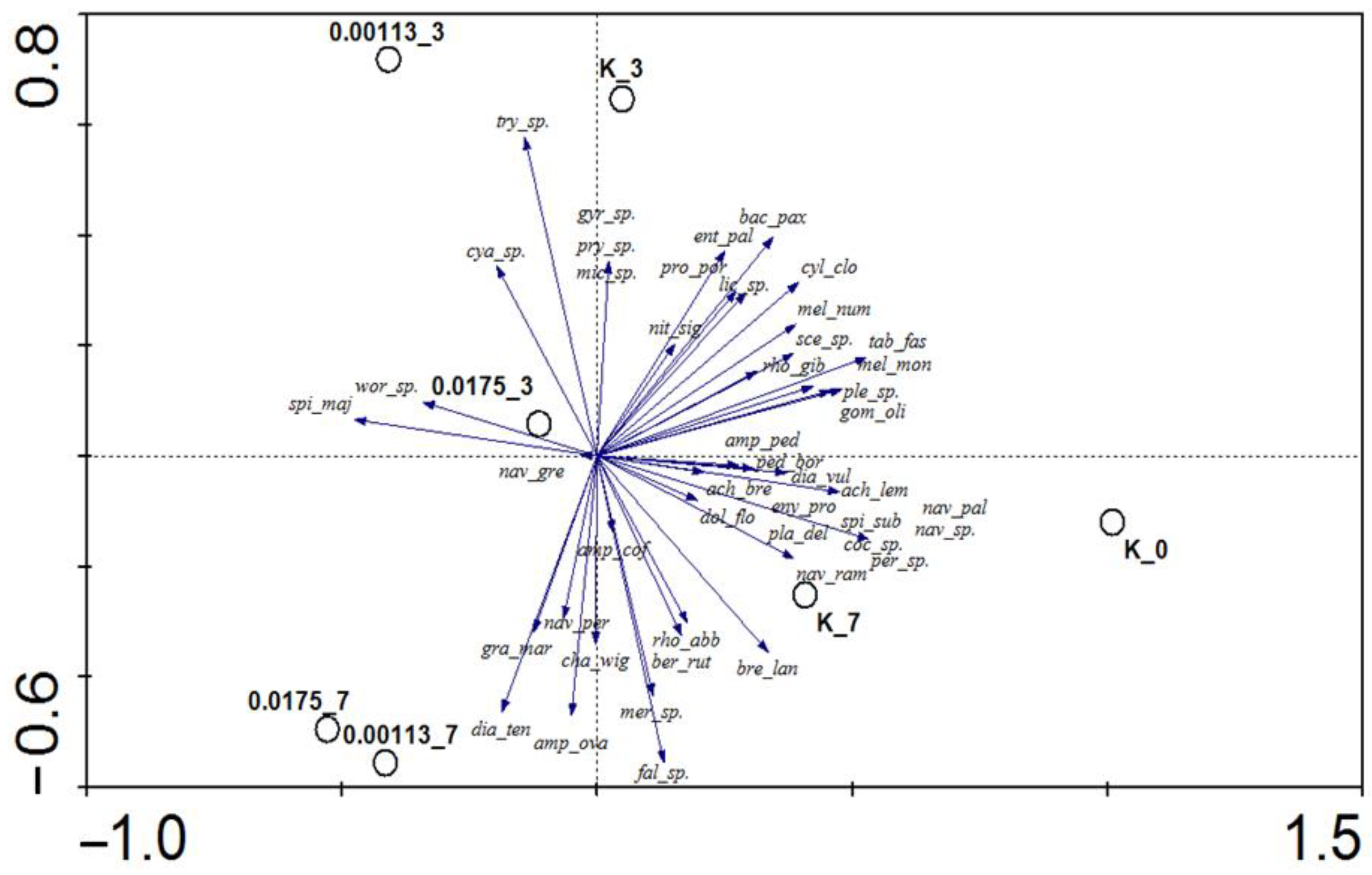
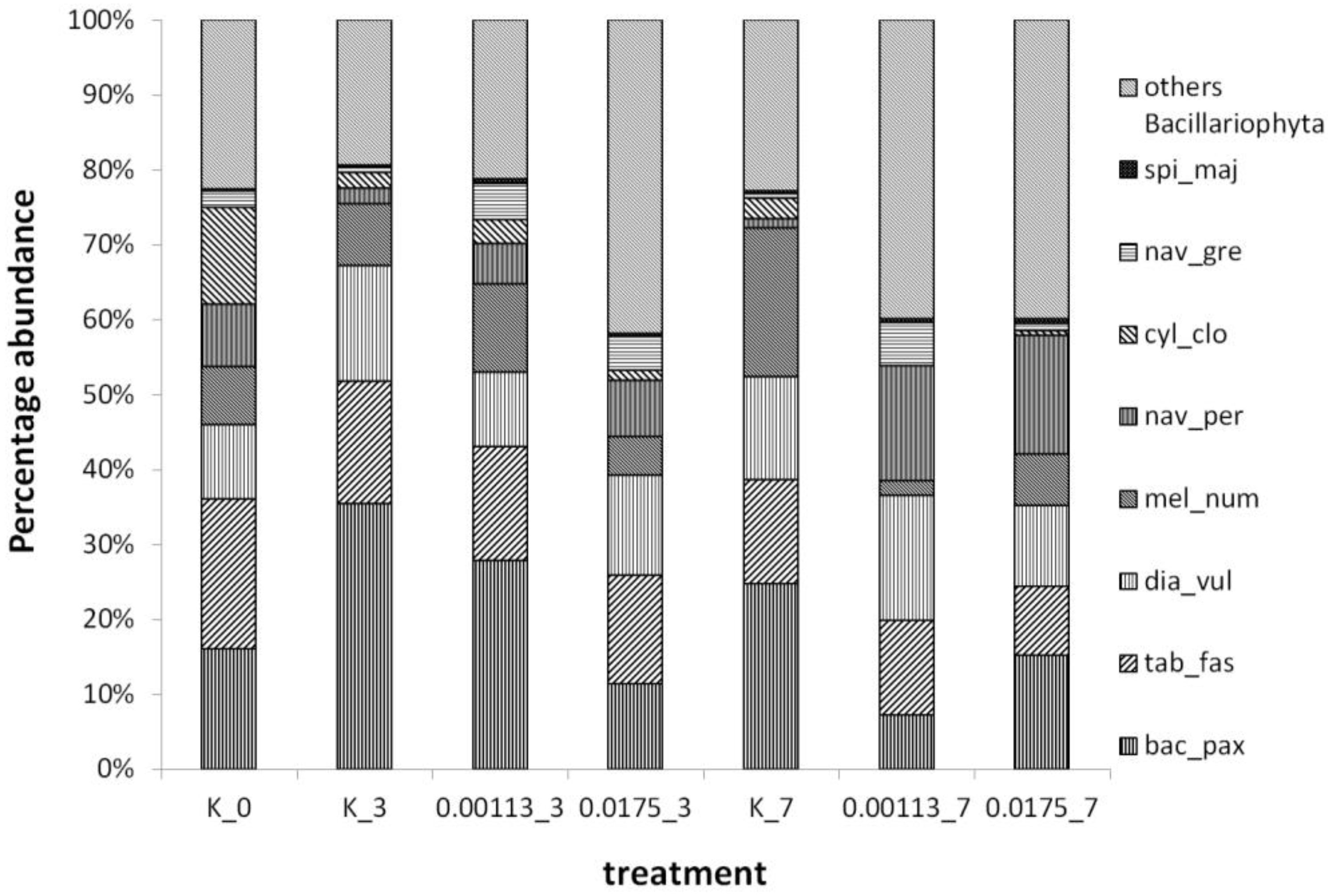
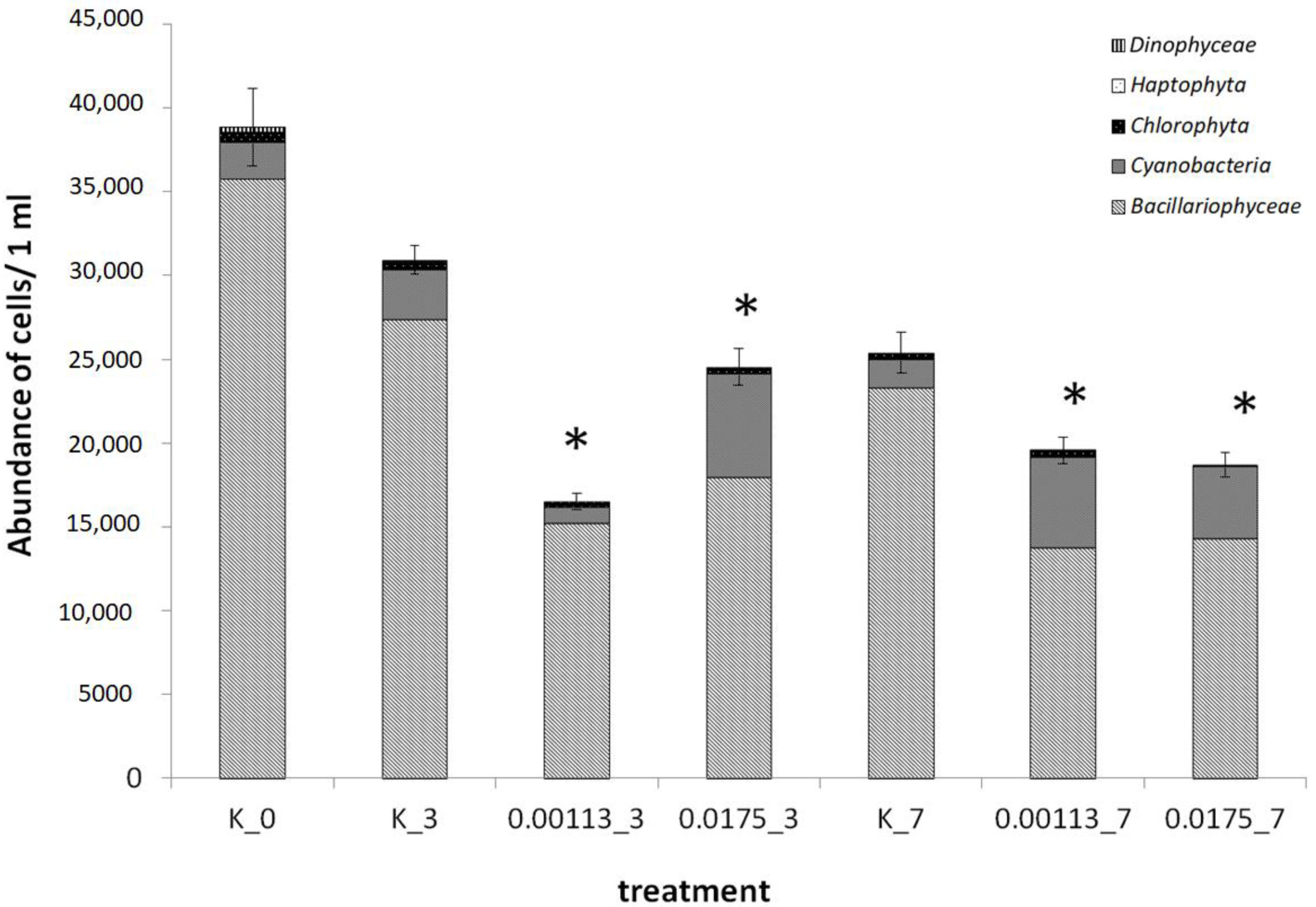
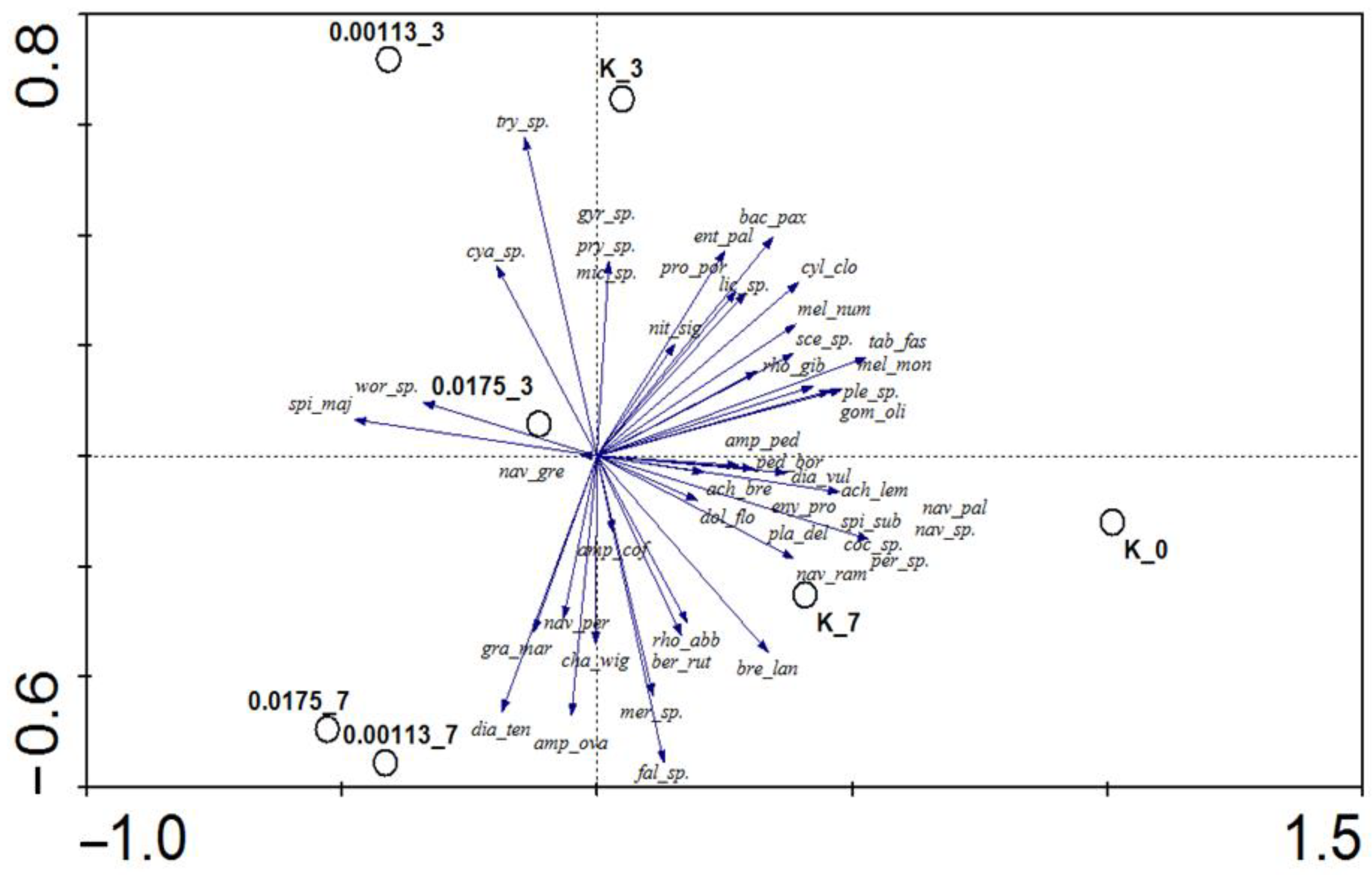
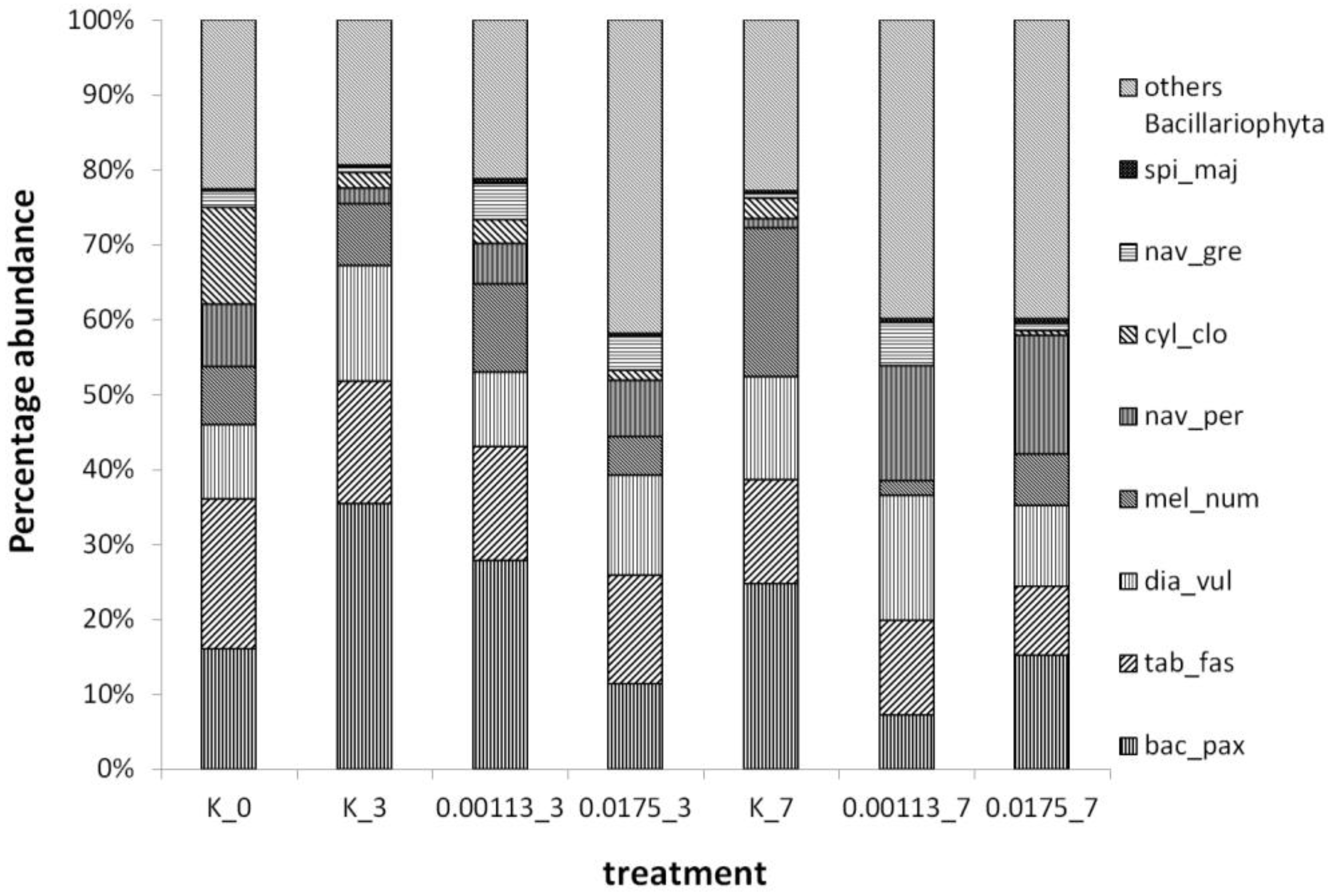
3.1. Analysis of Taxonomic Composition and Structure
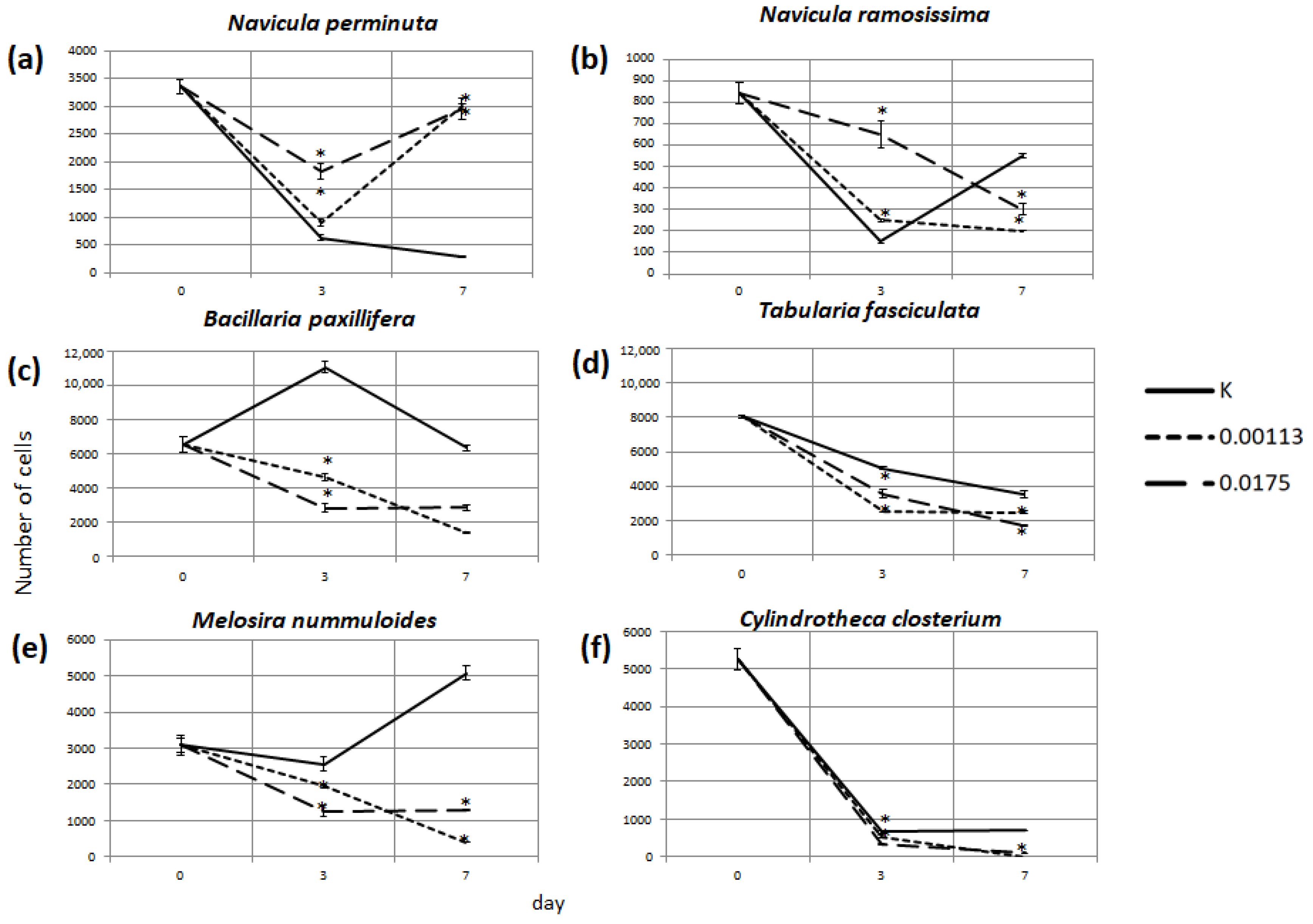
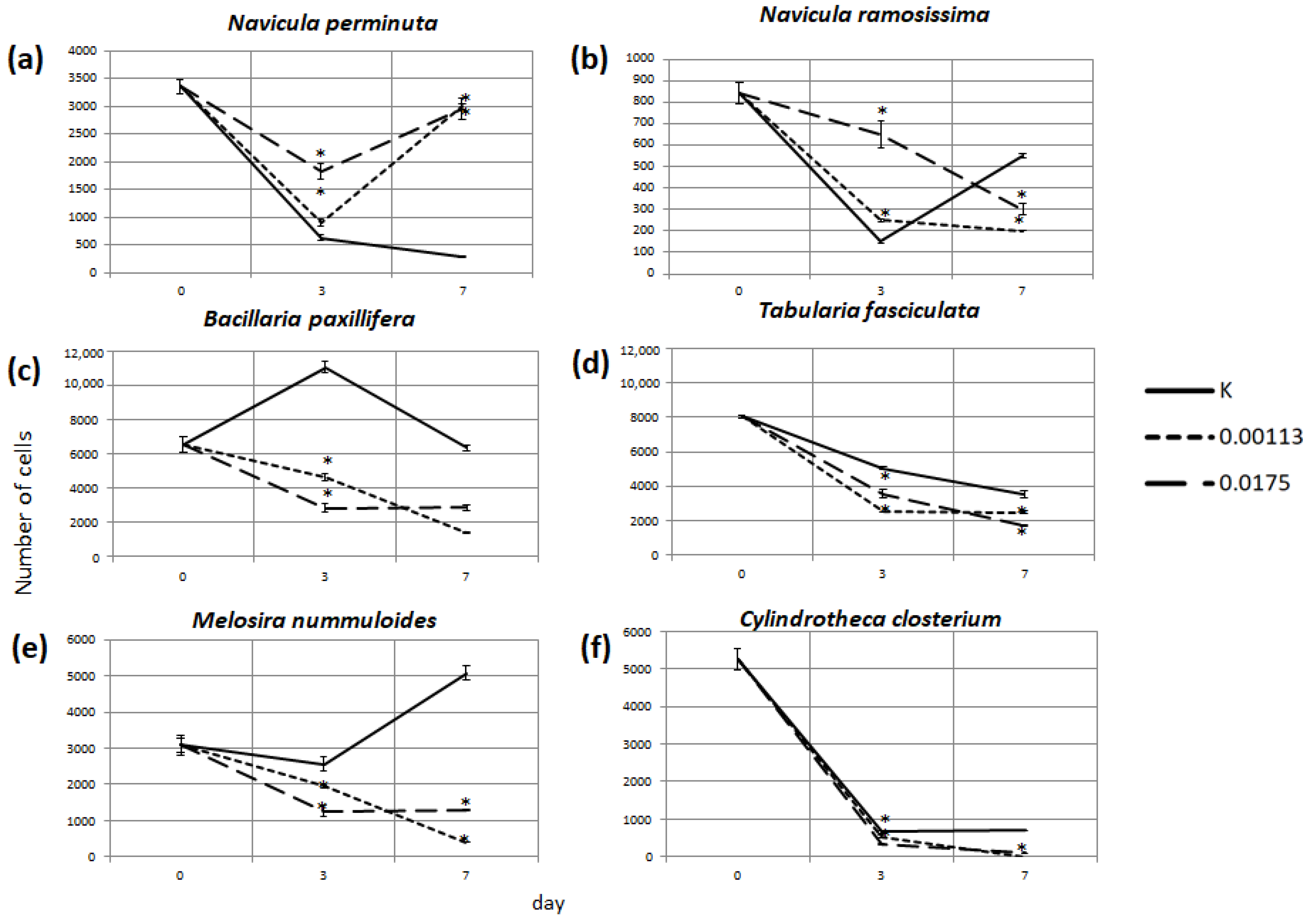
3.2. Abundance of Selected Taxa
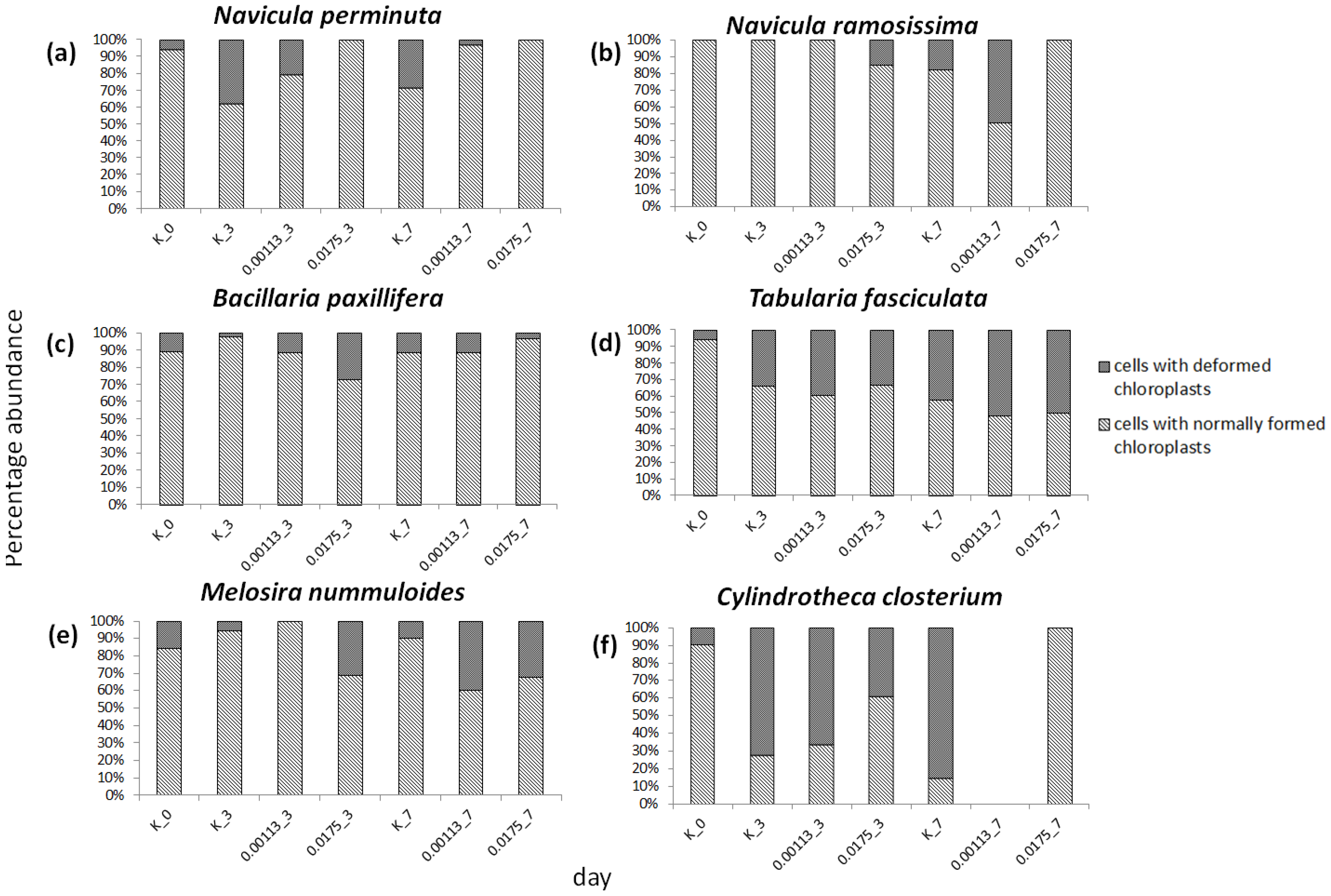
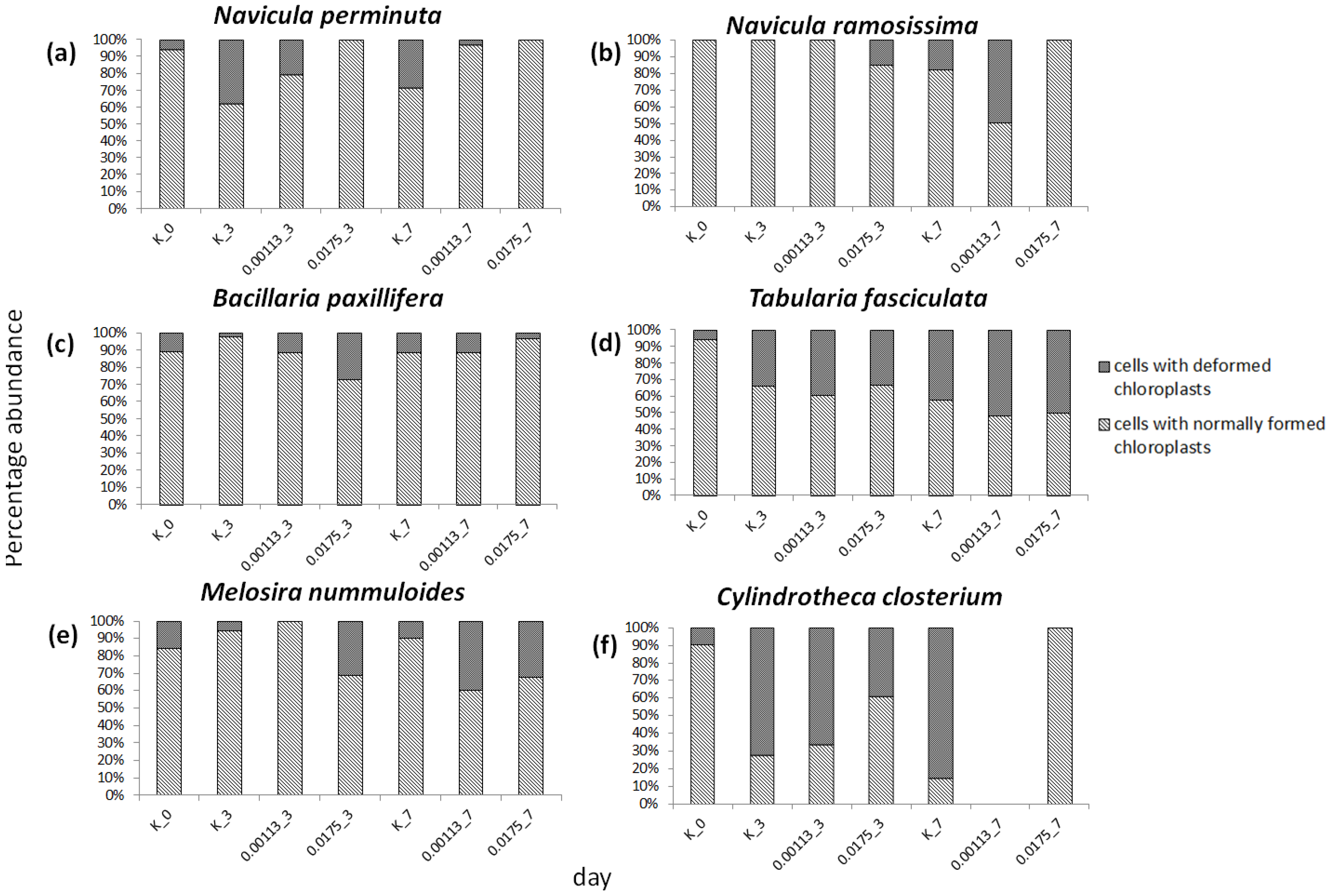
3.3. Cell Condition in Selected Taxa
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group of Organisms | Taxon Code | Taxon Name | Author |
|---|---|---|---|
| Bacillariophyta | ach_bre | Achnanthes adnata | Bory |
| ach_lem | Achnanthes lemmermannii | Hustedt | |
| amp_ova | Amphora ovalis | (Kützing) Kützing | |
| amp_ped | Amphora pediculus | (Kützing) Grunow | |
| bac_pax | Bacillaria paxillifera | (O.F. Müller) T. Marsson | |
| ber_rut | Berkeleya rutilans | (Trentepohl ex Roth) Grunow | |
| bre_lan | Brebissonia lanceolata | (C. Agardh) R.K. Mahoney & Reimer | |
| cha_wig | Chaetoceros wighamii | Brightwell | |
| coc_sp. | Cocconeis sp. | Ehrenberg | |
| cyl_clo | Cylindrotheca closterium | (Ehrenberg) Reimann & J.C. Lewin | |
| dia_ten | Diatoma tenuis | C. Agardh | |
| dia_vul | Diatoma vulgaris | Bory | |
| eny_pro | Encyonema leibleinii | (C. Agardh) W.J. Silva, R. Jahn, T.A.V. Ludwig, & M. Menezes | |
| ent_pal | Entomoneis paludosa | (W. Smith) Reimer | |
| fal_sp. | Fallacia sp. | Kützing | |
| gom_oli | Gomphonella olivacea | (Hornemann) Rabenhorst | |
| gram_mar | Grammatophora marina | (Lyngbye) Kützing | |
| gyr_sp. | Gyrosigma acuminatum | (Kützing) Rabenhorst | |
| amp_cof | Halamphora coffeiformis | (C. Agardh) Mereschkowsky | |
| lic_sp. | Licmophora gracilis | (Ehrenberg) Grunow | |
| mel_mon | Melosira moniliformis | C. Agardh | |
| mel_num | Melosira nummuloides | C. Agardh | |
| nav_gre | Navicula gregaria | Donkin | |
| nav_pal | Navicula palpebralis | Brébisson ex W. Smith | |
| nav_per | Navicula perminuta | Grunow | |
| nav_ram | Navicula ramossisima | (C. Agardh) Cleve | |
| nav_sp. | Navicula sp. | Bory | |
| nit_sig | Nitzschia sigma | (Kützing) W. Smith | |
| pla_del | Planothidium delicatulum | (Kützing) Round & Bukhtiyarova | |
| ple_sp. | Pleurosigma sp. | W. Smith | |
| pro_por | Proschkinia poretzkajae | (Koretkevich) D.G. Mann | |
| rho_abb | Rhoicosphenia abbreviata | (C. Agardh) Lange-Bertalot | |
| rho_gib | Rhopalodia gibba | (Ehrenberg) O. Müller | |
| tab_fas | Tabularia fasciculata | (C. Agardh) D.M. Williams & Round | |
| try_sp. | Tryblionella | W. Smith | |
| Cyanobacteria | dol_flo | Dolichospermum flosaquae | (Brébisson ex Bornet & Flahault) P. Wacklin, L. Hoffmann & J. Komárek |
| cya_sp. | Cyanobacteria | ||
| mer_sp. | Merismopedia sp. | (Turpin) Meneghini | |
| mic_sp. | Microcystis sp. | Lemmermann | |
| spi_maj | Spirulina major | Meyen | |
| spi_sub | Spirulina subsalsa | Oersted ex Gomont | |
| wor_sp. | Woronichinia sp. | A.A. Elenkin | |
| Chlorophyta | ped_bor | Pseudopediastrum boryanum | (Turpin) E. Hegewald |
| sce_sp. | Scenedesmus sp. | Meyen | |
| Dinophyceae | per_sp. | Peridinium sp. | Ehrenberg |
| Haptophyta | pry_sp. | Prymnesium sp. | N. Carter |
| Species | Av. Abundance | Av. Similarity | Sim/SD | Contribution% | Cumulative% |
|---|---|---|---|---|---|
| Bacillaria paxillifera | 6.06 | 6.13 | 8.84 | 8.52 | 8.52 |
| Tabularia fasciculata | 5.93 | 6.06 | 9.60 | 8.43 | 16.94 |
| Diatoma vulgaris | 5.73 | 5.89 | 9.08 | 8.19 | 25.13 |
| Melosira nummuloides | 5.09 | 5.02 | 7.19 | 6.98 | 32.12 |
| Navicula perminuta | 4.94 | 4.84 | 5.10 | 6.73 | 38.85 |
| Merismopedia sp. | 5.11 | 4.50 | 1.85 | 6.26 | 45.10 |
| Halamphora coffeiformis | 3.58 | 3.56 | 6.10 | 4.94 | 50.05 |
| Navicula gregaria | 3.73 | 3.52 | 4.99 | 4.90 | 54.94 |
| Navicula ramossisima | 3.54 | 3.40 | 6.27 | 4.72 | 59.67 |
| Cylindrotheca closterium | 3.99 | 3.17 | 1.78 | 4.40 | 64.07 |
| Grammatophora marina | 3.23 | 3.16 | 7.58 | 4.40 | 68.47 |
| Spirulina major | 2.38 | 2.55 | 7.16 | 3.55 | 72.01 |
References
- Chen, B.; Xue, C.; Amoah, P.K.; Li, D.; Gao, K.; Deng, X. Impacts of Four Ionic Liquids Exposure on a Marine Diatom Phaeodactylum Tricornutum at Physiological and Biochemical Levels. Sci. Total Environ. 2019, 665, 492–501. [Google Scholar] [CrossRef] [PubMed]
- Mai, N.L.; Ahn, K.; Koo, Y.-M. Methods for Recovery of Ionic Liquids—A Review. Process Biochem. 2014, 49, 872–881. [Google Scholar] [CrossRef]
- Liu, D.; Liu, H.; Wang, S.; Chen, J.; Xia, Y. The Toxicity of Ionic Liquid 1-Decylpyridinium Bromide to the Algae Scenedesmus Obliquus: Growth Inhibition, Phototoxicity, and Oxidative Stress. Sci. Total Environ. 2018, 622, 1572–1580. [Google Scholar] [CrossRef]
- Isosaari, P.; Srivastava, V.; Sillanpää, M. Ionic Liquid-Based Water Treatment Technologies for Organic Pollutants: Current Status and Future Prospects of Ionic Liquid Mediated Technologies. Sci. Total Environ. 2019, 690, 604–619. [Google Scholar] [CrossRef] [PubMed]
- Szepiński, E. Synteza, Właściwości i Zastosowanie Cieczy Jonowych, Pochodnych Naturalnych Związków Organicznych. Ph.D. Thesis, Department of Organic Chemistry, Faculty of Chemistry, Gdansk University of Technology, Gdansk, Poland, 2019. [Google Scholar]
- Cvjetko Bubalo, M.; Radošević, K.; Radojčić Redovniković, I.; Halambek, J.; Gaurina Srček, V. A Brief Overview of the Potential Environmental Hazards of Ionic Liquids. Ecotoxicol. Environ. Saf. 2014, 99, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Flieger, J.; Flieger, M. Ionic Liquids Toxicity—Benefits and Threats. Int. J. Mol. Sci. 2020, 21, 6267. [Google Scholar] [CrossRef]
- Nikitenko, S.I.; Berthon, C.; Moisy, P. Instability of Actinide (IV) Hexachloro Complexes in Room-Temperature Ionic Liquid [BuMeIm] PF6 Due to Hydrolysis of the Hexafluorophosphate Anion. Comptes Rendus Chim. 2007, 10, 1122–1127. [Google Scholar] [CrossRef]
- Freire, M.G.; Neves, C.M.S.S.; Marrucho, I.M.; Coutinho, J.A.P.; Fernandes, A.M. Hydrolysis of Tetrafluoroborate and Hexafluorophosphate Counter Ions in Imidazolium-Based Ionic Liquids. J. Phys. Chem. A 2010, 114, 3744–3749. [Google Scholar] [CrossRef]
- Maculewicz, J.; Świacka, K.; Stepnowski, P.; Dołżonek, J.; Białk-Bielińska, A. Ionic Liquids as Potentially Hazardous Pollutants: Evidences of Their Presence in the Environment and Recent Analytical Developments. J. Hazard. Mater. 2022, 437, 129353. [Google Scholar] [CrossRef]
- Kowalska, D.; Stolte, S.; Wyrzykowski, D.; Stepnowski, P.; Dołżonek, J. Interaction of Ionic Liquids with Human Serum Albumin in the View of Bioconcentration: A Preliminary Study. Chem. Pap. 2022, 76, 2405–2417. [Google Scholar] [CrossRef]
- Frade, R.F.; Afonso, C.A. Impact of Ionic Liquids in Environment and Humans: An Overview. Hum. Exp. Toxicol. 2010, 29, 1038–1054. [Google Scholar] [CrossRef] [PubMed]
- Costa, S.P.F.; Azevedo, A.M.O.; Pinto, P.; Saraiva, L. Environmental Impact of Ionic Liquids: An Overview of Recent (Eco) Toxicological and (Bio) Degradability Literature. ChemSusChem 2017, 10, 2321–2347. [Google Scholar] [CrossRef] [PubMed]
- Singh, S.K.; Savoy, A.W. Ionic Liquids Synthesis and Applications: An Overview. J. Mol. Liq. 2020, 297, 112038. [Google Scholar] [CrossRef]
- Armitage, J.M.; Erickson, R.J.; Luckenbach, T.; Ng, C.A.; Prosser, R.S.; Arnot, J.A.; Schirmer, K.; Nichols, J.W. Assessing the Bioaccumulation Potential of Ionizable Organic Compounds: Current Knowledge and Research Priorities. Environ. Toxicol. Chem. 2017, 36, 882–897. [Google Scholar] [CrossRef] [PubMed]
- Latała, A.; Stepnowski, P.; Nędzi, M.; Mrozik, W. Marine Toxicity Assessment of Imidazolium Ionic Liquids: Acute Effects on the Baltic Algae Oocystis Submarina and Cyclotella Meneghiniana. Aquat. Toxicol. 2005, 73, 91–98. [Google Scholar] [CrossRef] [PubMed]
- Latała, A.; Nędzi, M.; Stepnowski, P. Toxicity of Imidazolium Ionic Liquids towards Algae. Influence of Salinity Variations. Green Chem. 2010, 12, 60–64. [Google Scholar] [CrossRef]
- Kulacki, K.J.; Lamberti, G.A. Toxicity of Imidazolium Ionic Liquids to Freshwater Algae. Green Chem. 2008, 10, 104–110. [Google Scholar] [CrossRef]
- Samorì, C.; Sciutto, G.; Pezzolesi, L.; Galletti, P.; Guerrini, F.; Mazzeo, R.; Pistocchi, R.; Prati, S.; Tagliavini, E. Effects of Imidazolium Ionic Liquids on Growth, Photosynthetic Efficiency, and Cellular Components of the Diatoms Skeletonema Marinoi and Phaeodactylum Tricornutum. Chem. Res. Toxicol. 2011, 24, 392–401. [Google Scholar] [CrossRef]
- Fan, H.; Liu, H.; Dong, Y.; Chen, C.; Wang, Z.; Guo, J.; Du, S. Growth Inhibition and Oxidative Stress Caused by Four Ionic Liquids in Scenedesmus Obliquus: Role of Cations and Anions. Sci. Total Environ. 2019, 651, 570–579. [Google Scholar] [CrossRef]
- Zhu, Y.; Zhong, X.; Wang, Y.; Zhao, Q.; Huang, H. Growth Performance and Antioxidative Response of Chlorella Pyrenoidesa, Dunaliella Salina, and Anabaena Cylindrica to Four Kinds of Ionic Liquids. Appl. Biochem. Biotechnol. 2021, 193, 1945–1966. [Google Scholar] [CrossRef]
- Liu, H.; Zhang, S.; Zhang, X.; Chen, C. Growth Inhibition and Effect on Photosystem by Three Imidazolium Chloride Ionic Liquids in Rice Seedlings. J. Hazard. Mater. 2015, 286, 440–448. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Zhang, X.; Chen, C.; Du, S.; Dong, Y. Effects of Imidazolium Chloride Ionic Liquids and Their Toxicity to Scenedesmus Obliquus. Ecotoxicol. Environ. Saf. 2015, 122, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Dahl, B.; Blanck, H. Pollution-Induced Community Tolerance (PICT) in Periphyton Communities Established under Tri-n-Butyltin (TBT) Stress in Marine Microcosms. Aquat. Toxicol. 1996, 34, 305–325. [Google Scholar] [CrossRef]
- Porsbring, T.; Arrhenius, Å.; Backhaus, T.; Kuylenstierna, M.; Scholze, M.; Blanck, H. The SWIFT Periphyton Test for High-Capacity Assessments of Toxicant Effects on Microalgal Community Development. J. Exp. Mar. Biol. Ecol. 2007, 349, 299–312. [Google Scholar] [CrossRef]
- Sylwestrzak, Z.; Zgrundo, A.; Pniewski, F. Ecotoxicological Studies on the Effect of Roundup® (Glyphosate Formulation) on Marine Benthic Microalgae. Int. J. Environ. Res. Public. Health 2021, 18, 884. [Google Scholar] [CrossRef]
- Sylwestrzak, Z.; Zgrundo, A.; Pniewski, F. Copper Chloride (II) Effect on the Composition and Structure of Marine Microphytobenthic Communities. Environ. Monit. Assess. 2022, 194, 1–15. [Google Scholar] [CrossRef]
- Telesh, I.; Schubert, H.; Skarlato, S. Life in the Salinity Gradient: Discovering Mechanisms behind a New Biodiversity Pattern. Estuar. Coast. Shelf Sci. 2013, 135, 317–327. [Google Scholar] [CrossRef]
- Pham, T.P.T.; Cho, C.-W.; Yun, Y.-S. Environmental Fate and Toxicity of Ionic Liquids: A Review. Water Res. 2010, 44, 352–372. [Google Scholar] [CrossRef]
- Rosenberg, M.; Kulkarni, G.V.; Bosy, A.; McCulloch, C.A.G. Reproducibility and Sensitivity of Oral Malodor Measurements with a Portable Sulphide Monitor. J. Dent. Res. 1991, 70, 1436–1440. [Google Scholar] [CrossRef] [PubMed]
- Connolly, N.M.; Crossland, M.R.; Pearson, R.G. Effect of Low Dissolved Oxygen on Survival, Emergence, and Drift of Tropical Stream Macroinvertebrates. J. N. Am. Benthol. Soc. 2004, 23, 251–270. [Google Scholar] [CrossRef]
- Sylwestrzak, Z.; Zgrundo, A. Preliminary Results of Laboratory Tests Using Microphytobenthic Communities in the Gulf of Gdańsk for Toxicity Testing of Ionic Liquids. In Metodologia Badań Wykorzystywana Przez Młodych Naukowców; CREATIVETIME: Kraków, Poland, 2014; pp. 74–81. (In Polish) [Google Scholar]
- Sylwestrzak, Z.; Zgrundo, A.; Latała, A. Effect of the Ionic Liquid [BMIM] Cl on the Baltic Diatom Navicula ramosissma (C. Agardh) Cleve in a Laboratory Experiment on Natural Microphytobenthic Communities of the Gulf of Gdansk. In Zagadnienia Aktualnie Poruszane przez Młodych Naukowców 3; CREATIVETIME: Kraków, Poland, 2015; pp. 227–231. (In Polish) [Google Scholar]
- Utermöhl, H. Zur Vervollkommnung der Quantitativen Phytoplankton-MethodikMitteilungen Internationale Vereinigung Theoretische und Angewandte. Limnologie 1958, 9, 1–38. [Google Scholar]
- HELCOM. Manual for Marine Monitoring in the COMBINE Programme of HELCOM; HELCOM: Helsinki, Finland, 2014. [Google Scholar]
- Snoeijs, P.; Potapova, M. Intercalibration and Distribution of Diatom Species in the Baltic Sea; Opulus Press: Uppsala, Sweden, 1993; Volume 1. [Google Scholar]
- Snoeijs, P.; Vilbaste, S. Intercalibration and Distribution of Diatom Species in the Baltic Sea; Opulus Press: Uppsala, Sweden, 1994; Volume 2. [Google Scholar]
- Snoeijs, P.; Potapova, M. Intercalibration and Distribution of Diatom Species in the Baltic Sea; Opulus Press: Uppsala, Sweden, 1995; Volume 3. [Google Scholar]
- Snoeijs, P.; Kasperovičiene, J. Intercalibration and Distribution of Diatom Species in the Baltic Sea; Opulus Press: Uppsala, Sweden, 1996; Volume 4. [Google Scholar]
- Snoeijs, P.; Balashova, N. Intercalibration and Distribution of Diatom Species in the Baltic Sea; Opulus Press: Uppsala, Sweden, 1998; Volume 5. [Google Scholar]
- Witkowski, A.; Lange-Bertalot, H.; Metzeltin, D. Diatom Flora of Marine Coasts; A.R.G. Gantner Verlag K.G.: Königstein, Germany, 2000; Volume I. [Google Scholar]
- Pliński, M.; Hindák, F. Zielenice-Chlorophyta (Green Algae):(With the English Key for the Identification to the Genus), Cz. 1, Zielenice Nienitkowate (Prasinophyceae & Chlorophyceae) = Pt. 1, Non-Filamentous Green Algae; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2010. [Google Scholar]
- Pliński, M.; Komárek, J. Sinice – Cyanobakterie (Cyanoprokaryota):(With the English Key for the Identification to the Genus) Vol. 1; Wydawnictwo Uniwersytetu Gdańskiego: Gdańsk, Poland, 2007; p. 172. [Google Scholar]
- Lepš, J.; Šmilauer, P. Multivariate Analysis of Ecological Data Using CANOCO; Cambridge University Press: Cambridge, UK, 2003. [Google Scholar]
- Ter Braak, C.J.F.; Šmilauer, P. Program CANOCO, Version 4.52. Biometris: Quantitative Methods in the Life and Earth Sciences; Plant Research International, Wageningen University and Research Centre: Wageningen, The Netherlands, 2003. [Google Scholar]
- Zajac, A.; Kukawka, R.; Pawlowska-Zygarowicz, A.; Stolarska, O.; Smiglak, M. Ionic Liquids as Bioactive Chemical Tools for Use in Agriculture and the Preservation of Agricultural Products. Green Chem. 2018, 20, 4764–4789. [Google Scholar] [CrossRef]
- Kowalska, D.; Maculewicz, J.; Stepnowski, P.; Dołżonek, J. Ionic Liquids as Environmental Hazards–Crucial Data in View of Future PBT and PMT Assessment. J. Hazard. Mater. 2021, 403, 123896. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, L. Polyfluoroalkyl Compounds in the Aquatic Environment: A Review of Their Occurrence and Fate. J. Environ. Monit. 2011, 13, 20–31. [Google Scholar] [CrossRef]
- Li, Y.; Si-Cong, L.; Shi, Y.; Yu-Ping, Q. Effects of Soil Characteristics on Sorption-Desorption of 1-Butyl-3-Methyl-Imidazolium-Based Ionic Liquids. J. Agric. Resour. Environ. 2017, 34, 30. [Google Scholar]
- Neuwald, I.; Muschket, M.; Zahn, D.; Berger, U.; Seiwert, B.; Meier, T.; Kuckelkorn, J.; Strobel, C.; Knepper, T.P.; Reemtsma, T. Filling the Knowledge Gap: A Suspect Screening Study for 1310 Potentially Persistent and Mobile Chemicals with SFC-and HILIC-HRMS in Two German River Systems. Water Res. 2021, 204, 117645. [Google Scholar] [CrossRef]
- Pati, S.G.; Arnold, W.A. Comprehensive Screening of Quaternary Ammonium Surfactants and Ionic Liquids in Wastewater Effluents and Lake Sediments. Environ. Sci. Process. Impacts 2020, 22, 430–441. [Google Scholar] [CrossRef]
- Probert, P.M.; Leitch, A.C.; Dunn, M.P.; Meyer, S.K.; Palmer, J.M.; Abdelghany, T.M.; Lakey, A.F.; Cooke, M.P.; Talbot, H.; Wills, C. Identification of a Xenobiotic as a Potential Environmental Trigger in Primary Biliary Cholangitis. J. Hepatol. 2018, 69, 1123–1135. [Google Scholar] [CrossRef]
- Leitch, A.C.; Ibrahim, I.; Abdelghany, T.M.; Charlton, A.; Roper, C.; Vidler, D.; Palmer, J.M.; Wilson, C.; Jones, D.E.; Blain, P.G. The Methylimidazolium Ionic Liquid M8OI Is Detectable in Human Sera and Is Subject to Biliary Excretion in Perfused Human Liver. Toxicology 2021, 459, 152854. [Google Scholar] [CrossRef]
- Gathergood, N.; Scammells, P.J. Design and Preparation of Room-Temperature Ionic Liquids Containing Biodegradable Side Chains. Aust. J. Chem. 2002, 55, 557–560. [Google Scholar] [CrossRef]
- Stepnowski, P.; Zaleska, A. Comparison of Different Advanced Oxidation Processes for the Degradation of Room Temperature Ionic Liquids. J. Photochem. Photobiol. Chem. 2005, 170, 45–50. [Google Scholar] [CrossRef]
- Coleman, D.; Gathergood, N. Biodegradation Studies of Ionic Liquids. Chem. Soc. Rev. 2010, 39, 600. [Google Scholar] [CrossRef] [PubMed]
- Hagiwara, R.; Lee, J.S. Ionic Liquids for Electrochemical Devices. Electrochemistry 2007, 75, 23–34. [Google Scholar] [CrossRef]
- Amde, M.; Liu, J.-F.; Pang, L. Environmental Application, Fate, Effects, and Concerns of Ionic Liquids: A Review. Environ. Sci. Technol. 2015, 49, 12611–12627. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.I.; Mubashir, M.; Zaini, D.; Mahnashi, M.H.; Alyami, B.A.; Alqarni, A.O.; Show, P.L. Cumulative Impact Assessment of Hazardous Ionic Liquids towards Aquatic Species Using Risk Assessment Methods. J. Hazard. Mater. 2021, 415, 125364. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Zhang, X.; Deng, J.; Cheng, Y.; Chen, Z.; Qin, B.; Tefsen, B.; Wells, M. A New Perspective of Copper-Iron Effects on Bloom-Forming Algae in a Highly Impacted Environment. Water Res. 2021, 195, 116889. [Google Scholar] [CrossRef]
- Śliwińska-Wilczewska, S.; Sylwestrzak, Z.; Maculewicz, J.; Zgrundo, A.; Pniewski, F.; Latała, A. The Effects of Allelochemicals and Selected Anthropogenic Substances on the Diatom Bacillaria Paxillifera. Eduk. Biol. Sr. 2016, 1, 21–27. [Google Scholar]
- Zhang, C.; Zhu, L.; Wang, J.; Wang, J.; Zhou, T.; Xu, Y.; Cheng, C. The Acute Toxic Effects of Imidazolium-Based Ionic Liquids with Different Alkyl-Chain Lengths and Anions on Zebrafish (Danio Rerio). Ecotoxicol. Environ. Saf. 2017, 140, 235–240. [Google Scholar] [CrossRef]
- Kim, Y.; Ponomarev, A.V. Low-Dose Electron Beam Treatment of Red Tide Blooms Microalgae. Radiat. Phys. Chem. 2021, 179, 109201. [Google Scholar] [CrossRef]
- Jungnickel, C.; Mrozik, W.; Markiewicz, M.; Luczak, J. Fate of Ionic Liquids in Soils and Sediments. Curr. Org. Chem. 2011, 15, 1928–1945. [Google Scholar] [CrossRef] [Green Version]
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sylwestrzak, Z.; Zgrundo, A.; Pniewski, F. Effects of the Ionic Liquid [BMIM]Cl on the Baltic Microphytobenthic Communities. J. Mar. Sci. Eng. 2022, 10, 1223. https://doi.org/10.3390/jmse10091223
Sylwestrzak Z, Zgrundo A, Pniewski F. Effects of the Ionic Liquid [BMIM]Cl on the Baltic Microphytobenthic Communities. Journal of Marine Science and Engineering. 2022; 10(9):1223. https://doi.org/10.3390/jmse10091223
Chicago/Turabian StyleSylwestrzak, Zuzanna, Aleksandra Zgrundo, and Filip Pniewski. 2022. "Effects of the Ionic Liquid [BMIM]Cl on the Baltic Microphytobenthic Communities" Journal of Marine Science and Engineering 10, no. 9: 1223. https://doi.org/10.3390/jmse10091223
APA StyleSylwestrzak, Z., Zgrundo, A., & Pniewski, F. (2022). Effects of the Ionic Liquid [BMIM]Cl on the Baltic Microphytobenthic Communities. Journal of Marine Science and Engineering, 10(9), 1223. https://doi.org/10.3390/jmse10091223