
Promoting the Growth of Haematococcus lacustris under High Light Intensity through the Combination of Light/Dark Cycle and Light Color



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Strain and Cultural Conditions
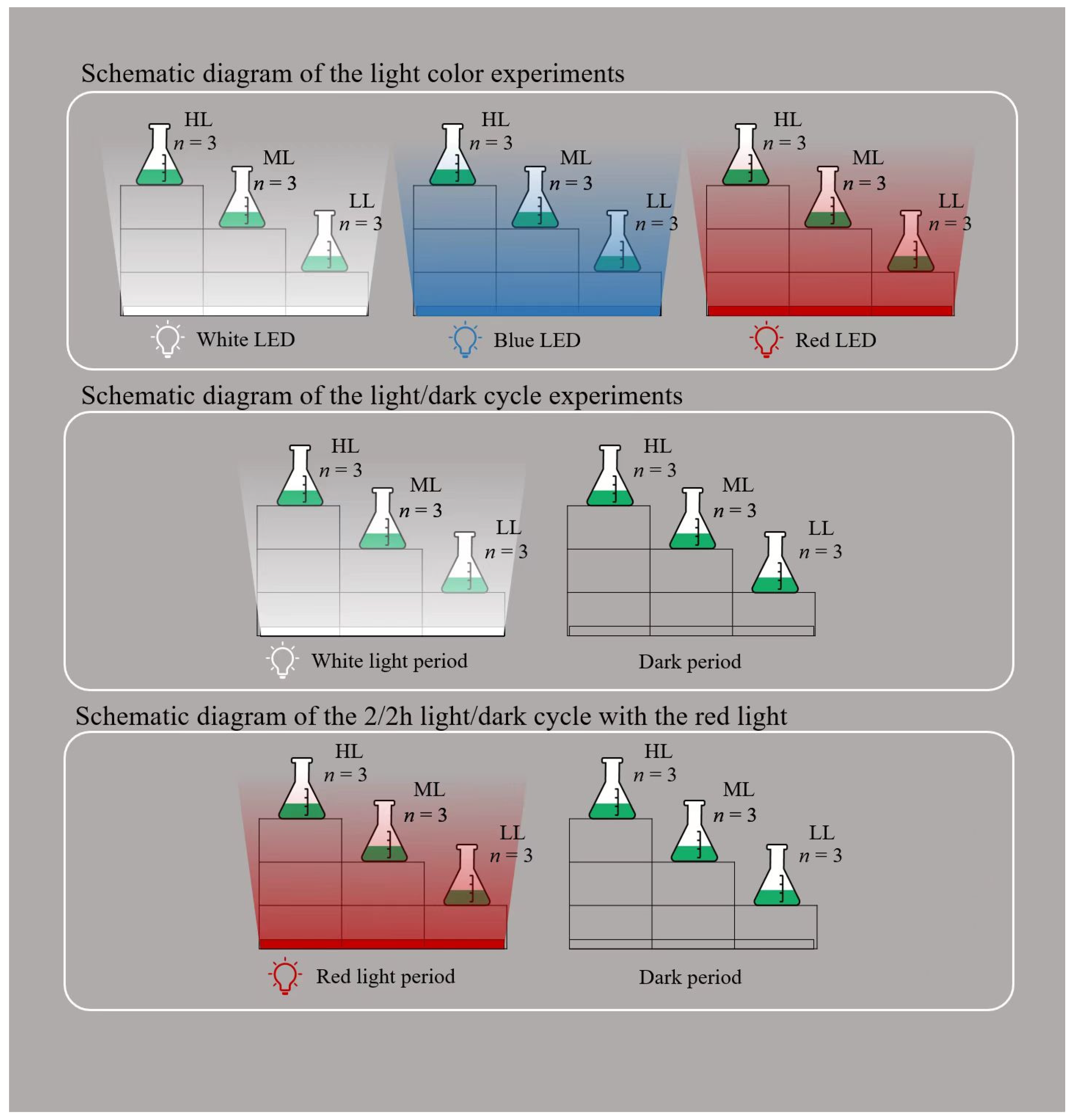
2.2. Experimental Setup
2.3. Analytical Methods
2.4. Statistical Analysis
3. Results
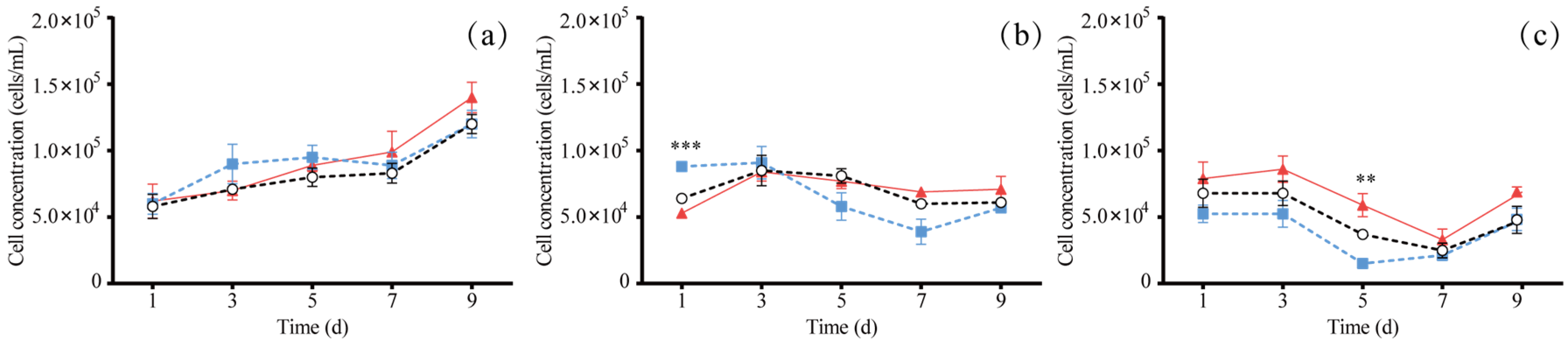
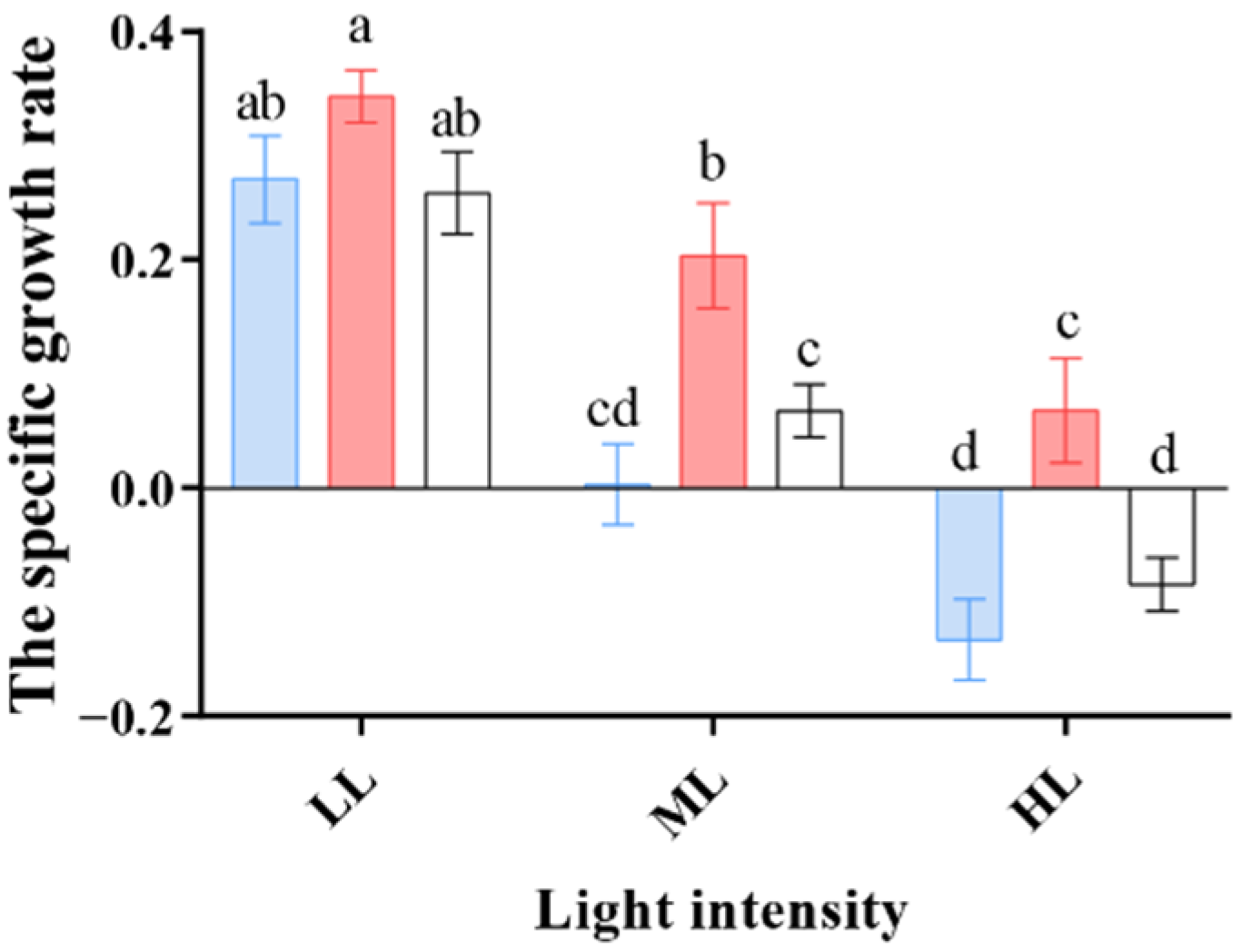
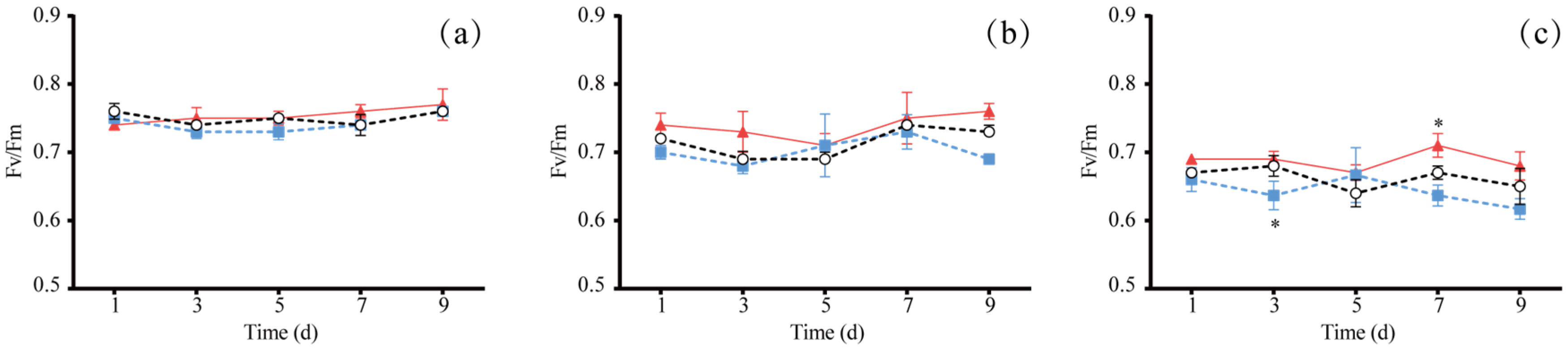
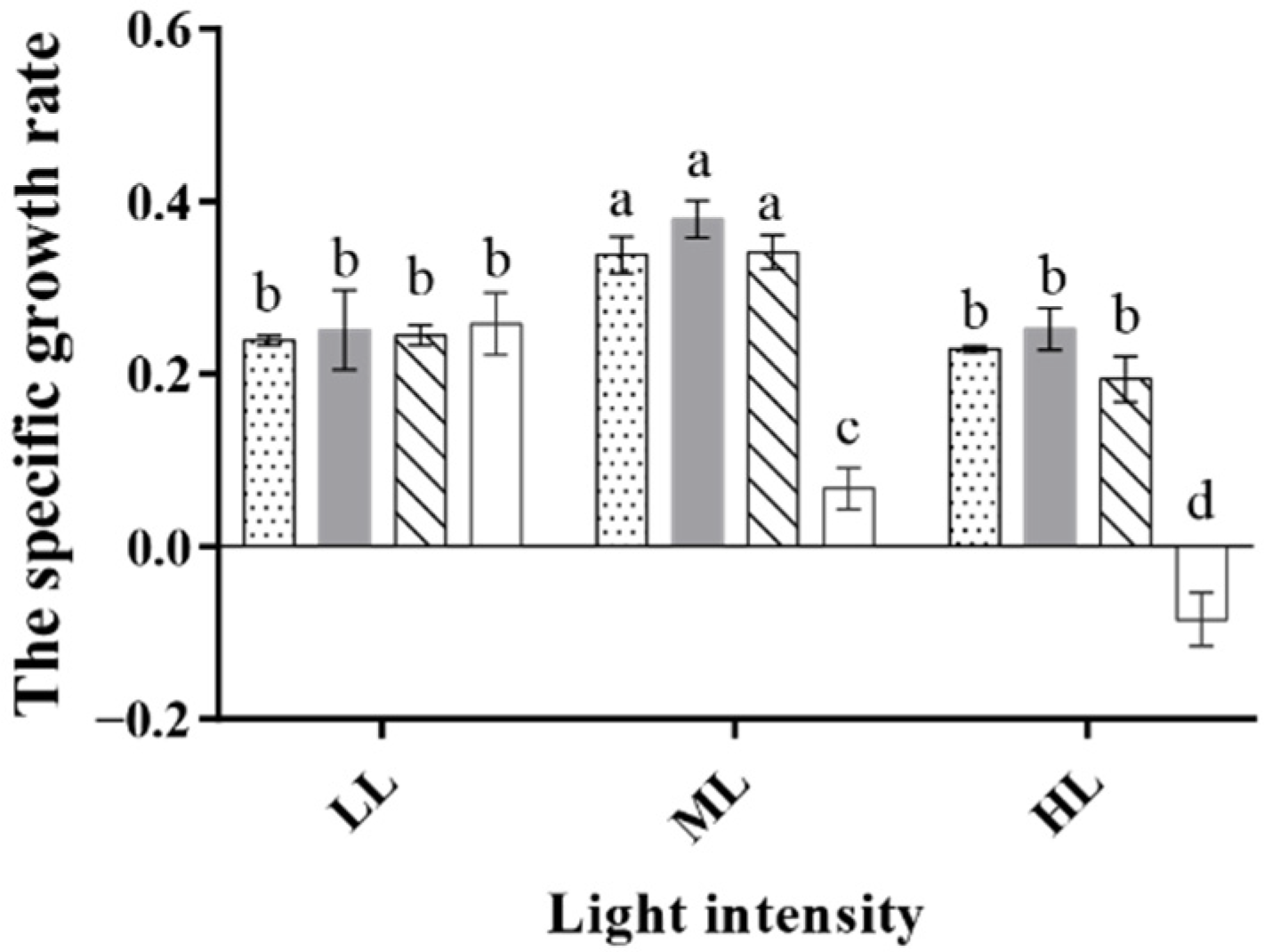
3.1. The Effect of Different Light Colors on Algal Growth
3.2. The Effect of Light/Dark Cycles on Algal Growth
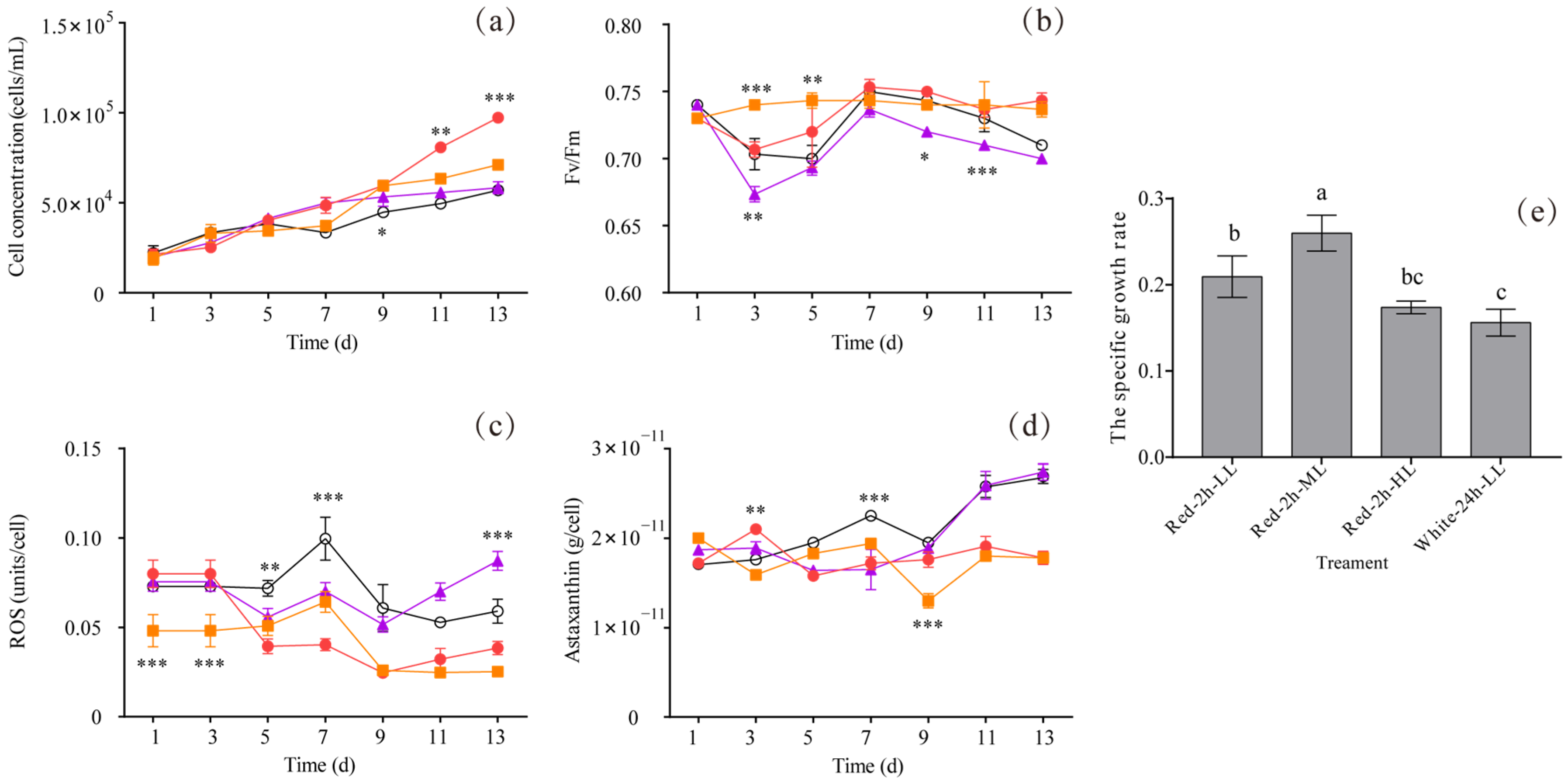
3.3. The Effect of 2/2 h Light/Dark Cycles with Red Light on Algal Growth
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brown, D.R.; Gough, L.A.; Deb, S.K.; Sparks, S.A.; McNaughton, L.R. Astaxanthin in Exercise Metabolism, Performance and Recovery: A Review. Front. Nutr. 2018, 4, 76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakajima, Y.; Inokuchi, Y.; Shimazawa, M.; Otsubo, K.; Ishibashi, T.; Hara, H. Astaxanthin, a Dietary Carotenoid, Protects Retinal Cells against Oxidative Stress in-Vitro and in Mice In-Vivo. J. Pharm. Pharmacol. 2008, 60, 1365–1374. [Google Scholar] [CrossRef] [PubMed]
- Oslan, S.N.H.; Shoparwe, N.F.; Yusoff, A.H.; Rahim, A.A.; Chang, C.S.; Tan, J.S.; Oslan, S.N.; Arumugam, K.; Ariff, A.B.; Sulaiman, A.Z.; et al. A Review on Haematococcus Pluvialis Bioprocess Optimization of Green and Red Stage Culture Conditions for the Production of Natural Astaxanthin. Biomolecules 2021, 11, 256. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Liu, J.; Zhang, L. Cell Cycles and Proliferation Patterns in Haematococcus Pluvialis. Chin. J. Ocean. Limnol. 2017, 35, 1205–1211. [Google Scholar] [CrossRef]
- Jeon, Y.-C.; Cho, C.-W.; Yun, Y.-S. Combined Effects of Light Intensity and Acetate Concentration on the Growth of Unicellular Microalga Haematococcus Pluvialis. Enzym. Microb. Technol. 2006, 39, 490–495. [Google Scholar] [CrossRef]
- Kang, C.D.; Han, S.J.; Choi, S.P.; Sim, S.J. Fed-Batch Culture of Astaxanthin-Rich Haematococcus Pluvialis by Exponential Nutrient Feeding and Stepwise Light Supplementation. Bioprocess Biosyst. Eng. 2010, 33, 133–139. [Google Scholar] [CrossRef]
- Carvalho, A.P.; Silva, S.O.; Baptista, J.M.; Malcata, F.X. Light Requirements in Microalgal Photobioreactors: An Overview of Biophotonic Aspects. Appl. Microbiol. Biotechnol. 2011, 89, 1275–1288. [Google Scholar] [CrossRef]
- Bustos-Garza, C.; Yáñez-Fernández, J.; Barragán-Huerta, B.E. Thermal and PH Stability of Spray-Dried Encapsulated Astaxanthin Oleoresin from Haematococcus Pluvialis Using Several Encapsulation Wall Materials. Food Res. Int. 2013, 54, 641–649. [Google Scholar] [CrossRef]
- Cui, S.; Cai, X.; Zhang, Y.; Wu, X.; He, L. Research on PH Value Control System of Haematococcus Pluvialis Algae Based on Fuzzy PID Algorithm. J. Phys. Conf. Ser. 2020, 1631, 012051. [Google Scholar] [CrossRef]
- Hong, M.-E.; Choi, Y.Y.; Sim, S.J. Effect of Red Cyst Cell Inoculation and Iron(II) Supplementation on Autotrophic Astaxanthin Production by Haematococcus Pluvialis under Outdoor Summer Conditions. J. Biotechnol. 2016, 218, 25–33. [Google Scholar] [CrossRef]
- Pereira, S.; Otero, A. Haematococcus Pluvialis Bioprocess Optimization: Effect of Light Quality, Temperature and Irradiance on Growth, Pigment Content and Photosynthetic Response. Algal Res. 2020, 51, 102027. [Google Scholar] [CrossRef]
- Gong, W.; Zhao, L.; Hu, B.; Chen, X.; Zhang, F.; Zhu, Z.; Chen, D. Identifying Novel Salt-Tolerant Genes from Dunaliella Salina Using a Haematococcus Pluvialis Expression System. Plant Cell Tiss Organ Cult. 2014, 117, 113–124. [Google Scholar] [CrossRef]
- Fábregas, J.; Domínguez, A.; Regueiro, M.; Maseda, A.; Otero, A. Optimization of Culture Medium for the Continuous Cultivation of the Microalga Haematococcus Pluvialis. Appl. Microbiol. Biotechnol. 2000, 53, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Wang, H.-P.; Yu, C.; Ding, W.; Han, B.; Geng, S.; Ning, D.; Ma, T.; Yu, X. Integration of Physiological and Metabolomic Profiles to Elucidate the Regulatory Mechanisms Underlying the Stimulatory Effect of Melatonin on Astaxanthin and Lipids Coproduction in Haematococcus Pluvialis under Inductive Stress Conditions. Bioresour. Technol. 2021, 319, 124150. [Google Scholar] [CrossRef] [PubMed]
- García-Malea, M.C.; Brindley, C.; Río, E.D.; Acién, F.G.; Fernández, J.M.; Molina, E. Modelling of Growth and Accumulation of Carotenoids in Haematococcus Pluvialis as a Function of Irradiance and Nutrients Supply. Biochem. Eng. J. 2005, 26, 107–114. [Google Scholar] [CrossRef] [Green Version]
- Kaewpintong, K.; Shotipruk, A.; Powtongsook, S.; Pavasant, P. Photoautotrophic High-Density Cultivation of Vegetative Cells of Haematococcus Pluvialis in Airlift Bioreactor. Bioresour. Technol. 2007, 98, 288–295. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]
- Muramatsu, M.; Hihara, Y. Acclimation to High-Light Conditions in Cyanobacteria: From Gene Expression to Physiological Responses. J. Plant Res. 2012, 125, 11–39. [Google Scholar] [CrossRef]
- Choi, S.-L.; Suh, I.S.; Lee, C.-G. Lumostatic Operation of Bubble Column Photobioreactors for Haematococcus Pluvialis Cultures Using a Specific Light Uptake Rate as a Control Parameter. Enzym. Microb. Technol. 2003, 33, 403–409. [Google Scholar] [CrossRef]
- Lee, H.-S.; Seo, M.-W.; Kim, Z.-H.; Lee, C.-G. Determining the Best Specific Light Uptake Rates for the Lumostatic Cultures in Bubble Column Photobioreactors. Enzym. Microb. Technol. 2006, 39, 447–452. [Google Scholar] [CrossRef]
- Abiusi, F.; Sampietro, G.; Marturano, G.; Biondi, N.; Rodolfi, L.; D’Ottavio, M.; Tredici, M.R. Growth, Photosynthetic Efficiency, and Biochemical Composition of Tetraselmis Suecica F&M-M33 Grown with LEDs of Different Colors: Growth of Tetraselmis With Different LED Colors. Biotechnol. Bioeng. 2014, 111, 956–964. [Google Scholar] [CrossRef] [PubMed]
- Baba, M.; Kikuta, F.; Suzuki, I.; Watanabe, M.M.; Shiraiwa, Y. Wavelength Specificity of Growth, Photosynthesis, and Hydrocarbon Production in the Oil-Producing Green Alga Botryococcus Braunii. Bioresour. Technol. 2012, 109, 266–270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsuda, T.; Lababpour, A.; Shimahara, K.; Katoh, S. Astaxanthin Production by Haematococcus Pluvialis under Illumination with LEDs. Enzym. Microb. Technol. 2004, 35, 81–86. [Google Scholar] [CrossRef] [Green Version]
- Kim, T.-H.; Lee, Y.; Han, S.-H.; Hwang, S.-J. The Effects of Wavelength and Wavelength Mixing Ratios on Microalgae Growth and Nitrogen, Phosphorus Removal Using Scenedesmus Sp. for Wastewater Treatment. Bioresour. Technol. 2013, 130, 75–80. [Google Scholar] [CrossRef]
- Schulze, P.S.C.; Barreira, L.A.; Pereira, H.G.C.; Perales, J.A.; Varela, J.C.S. Light Emitting Diodes (LEDs) Applied to Microalgal Production. Trends Biotechnol. 2014, 32, 422–430. [Google Scholar] [CrossRef]
- Das, P.; Lei, W.; Aziz, S.S.; Obbard, J.P. Enhanced Algae Growth in Both Phototrophic and Mixotrophic Culture under Blue Light. Bioresour. Technol. 2011, 102, 3883–3887. [Google Scholar] [CrossRef]
- Ma, R.; Thomas-Hall, S.R.; Chua, E.T.; Eltanahy, E.; Netzel, M.E.; Netzel, G.; Lu, Y.; Schenk, P.M. Blue Light Enhances Astaxanthin Biosynthesis Metabolism and Extraction Efficiency in Haematococcus Pluvialis by Inducing Haematocyst Germination. Algal Res. 2018, 35, 215–222. [Google Scholar] [CrossRef]
- Torres-Carvajal, L.K.; Gonzalez-Delgado, A.D.; Barajas-Solano, A.F.; Suarez-Gelvez, J.H.; Urbina-Suarez, N.A. Astaxanthin Production from Haematococcus Pluvialis: Effects of Light Wavelength and Salinity. Contemp. Eng. Sci. 2017, 10, 1739–1746. [Google Scholar] [CrossRef]
- Xiaojuan, Z.; Jie, L.; Chengxu, Z.; Jianyuan, W.; Ran, M.; Jilin, X.; Juanjuan, C.; Qijun, L.; Xiaojun, Y. Changes of Pigments and Lipids Composition in Haematococcus Pluvialis Vegetative Cell as Affected by Monochromatic Red Light Compared with White Light. J. Oceanol. Limnol. 2018, 36, 11. [Google Scholar] [CrossRef]
- Grobbelaar, J.U.; Nedbal, L.; Tichý, V. Influence of High Frequency Light/Dark Fluctuations on Photosynthetic Characteristics of Microalgae Photoacclimated to Different Light Intensities and Implications for Mass Algal Cultivation. J. Appl. Phycol. 1996, 8, 335–343. [Google Scholar] [CrossRef]
- Janssen, M.; Slenders, P.; Tramper, J.; Mur, L.R.; Wijffels, R. Photosynthetic Efficiency of Dunaliella Tertiolecta under Short Light/Dark Cycles. Enzym. Microb. Technol. 2001, 29, 298–305. [Google Scholar] [CrossRef]
- Sforza, E.; Simionato, D.; Giacometti, G.M.; Bertucco, A.; Morosinotto, T. Adjusted Light and Dark Cycles Can Optimize Photosynthetic Efficiency in Algae Growing in Photobioreactors. PLoS ONE 2012, 7, e38975. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, A.; Pereira, S.; Otero, A. Does Haematococcus Pluvialis Need to Sleep? Algal Res. 2019, 44, 101722. [Google Scholar] [CrossRef]
- Cheng, J.; Li, K.; Yang, Z.; Zhou, J.; Cen, K. Enhancing the Growth Rate and Astaxanthin Yield of Haematococcus Pluvialis by Nuclear Irradiation and High Concentration of Carbon Dioxide Stress. Bioresour. Technol. 2016, 204, 49–54. [Google Scholar] [CrossRef] [PubMed]
- Nahidian, B.; Ghanati, F.; Shahbazi, M.; Soltani, N. Effect of Nutrients on the Growth and Physiological Features of Newly Isolated Haematococcus Pluvialis TMU1. Bioresour. Technol. 2018, 255, 229–237. [Google Scholar] [CrossRef]
- Wang, J.; Sommerfeld, M.R.; Lu, C.; Hu, Q. Combined Effect of Initial Biomass Density and Nitrogen Concentration on Growth and Astaxanthin Production of Haematococcus Pluvialis (Chlorophyta) in Outdoor Cultivation. ALGAE 2013, 28, 193–202. [Google Scholar] [CrossRef]
- Krause, G.H.; Weis, E. Chlorophyll Fluorescence and Photosynthesis: The Basics. Annu. Rev. Plant. Physiol. Plant. Mol. Biol. 1991, 42, 313–349. [Google Scholar] [CrossRef]
- Tan, L.; Xu, W.; He, X.; Wang, J. The Feasibility of Fv/Fm on Judging Nutrient Limitation of Marine Algae through Indoor Simulation and in Situ Experiment. Estuar. Coast. Shelf Sci. 2019, 229, 106411. [Google Scholar] [CrossRef]
- Sun, H.; Liu, B.; Lu, X.; Cheng, K.-W.; Chen, F. Staged Cultivation Enhances Biomass Accumulation in the Green Growth Phase of Haematococcus Pluvialis. Bioresour. Technol. 2017, 233, 326–331. [Google Scholar] [CrossRef]
- Karadag, A.; Ozcelik, B.; Saner, S. Review of Methods to Determine Antioxidant Capacities. Food Anal. Methods 2009, 2, 41–60. [Google Scholar] [CrossRef]
- Shahidi, F.; Zhong, Y. Measurement of Antioxidant Activity. J. Funct. Foods 2015, 18, 757–781. [Google Scholar] [CrossRef]
- Zhang, H.; An, X.; Zhou, Y.; Zhang, B.; Zhang, S.; Li, D.; Chen, Z.; Li, Y.; Bai, S.; Lv, J.; et al. Effect of Oxidative Stress Induced by Brevibacterium Sp. BS01 on a HAB Causing Species-Alexandrium Tamarense. PLoS ONE 2013, 8, 9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kobayashi, M.; Kakizono, T.; Nishio, N.; Nagai, S.; Kurimura, Y.; Tsuji, Y. Antioxidant Role of Astaxanthin in the Green Alga Haematococcus Pluvialis. Appl. Microbiol. Biotechnol. 1997, 48, 351–356. [Google Scholar] [CrossRef]
- Mallick, N.; Mohn, F.H. Reactive Oxygen Species: Response of Algal Cells. J. Plant Physiol. 2000, 157, 183–193. [Google Scholar] [CrossRef]
- Masojídek, J.; Torzillo, G.; Koblížek, M. Photosynthesis in Microalgae. In Handbook of Microalgal Culture; Richmond, A., Hu, Q., Eds.; John Wiley & Sons, Ltd.: Oxford, UK, 2013; pp. 21–36. ISBN 978-1-118-56716-6. [Google Scholar]
- Powles, S.B. Photoinhibition of Photosynthesis Induced by Visible Light. Annu. Rev. Plant. Physiol. 1984, 35, 15–44. [Google Scholar] [CrossRef]
- Masojídek, J.; Ranglová, K.; Lakatos, G.E.; Silva Benavides, A.M.; Torzillo, G. Variables Governing Photosynthesis and Growth in Microalgae Mass Cultures. Processes 2021, 9, 820. [Google Scholar] [CrossRef]
- Falkowski, P.G.; Owens, T.G. Effects of Light Intensity on Photosynthesis and Dark Respiration in Six Species of Marine Phytoplankton. Mar. Biol. 1978, 45, 289–295. [Google Scholar] [CrossRef]
- Goss, R.; Lepetit, B. Biodiversity of NPQ. J. Plant Physiol. 2015, 172, 13–32. [Google Scholar] [CrossRef]
- Kim, Z.-H.; Kim, S.-H.; Lee, H.-S.; Lee, C.-G. Enhanced Production of Astaxanthin by Flashing Light Using Haematococcus Pluvialis. Enzym. Microb. Technol. 2006, 39, 414–419. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, L.; Ying, K.; Wu, K.; Tang, S.; Zhou, J.; Cai, Z. Promoting the Growth of Haematococcus lacustris under High Light Intensity through the Combination of Light/Dark Cycle and Light Color. J. Mar. Sci. Eng. 2022, 10, 839. https://doi.org/10.3390/jmse10070839
Liu L, Ying K, Wu K, Tang S, Zhou J, Cai Z. Promoting the Growth of Haematococcus lacustris under High Light Intensity through the Combination of Light/Dark Cycle and Light Color. Journal of Marine Science and Engineering. 2022; 10(7):839. https://doi.org/10.3390/jmse10070839
Chicago/Turabian StyleLiu, Lu, Kezhen Ying, Kebi Wu, Si Tang, Jin Zhou, and Zhonghua Cai. 2022. "Promoting the Growth of Haematococcus lacustris under High Light Intensity through the Combination of Light/Dark Cycle and Light Color" Journal of Marine Science and Engineering 10, no. 7: 839. https://doi.org/10.3390/jmse10070839
APA StyleLiu, L., Ying, K., Wu, K., Tang, S., Zhou, J., & Cai, Z. (2022). Promoting the Growth of Haematococcus lacustris under High Light Intensity through the Combination of Light/Dark Cycle and Light Color. Journal of Marine Science and Engineering, 10(7), 839. https://doi.org/10.3390/jmse10070839