
Growth Strategy of Rhizomatous and Non-Rhizomatous Tall Fescue Populations in Response to Defoliation

Abstract
:1. Introduction
2. Experimental Section
2.1. Plant Preparation and Management
2.2. Measurements
2.3. Statistical Analysis
3. Results
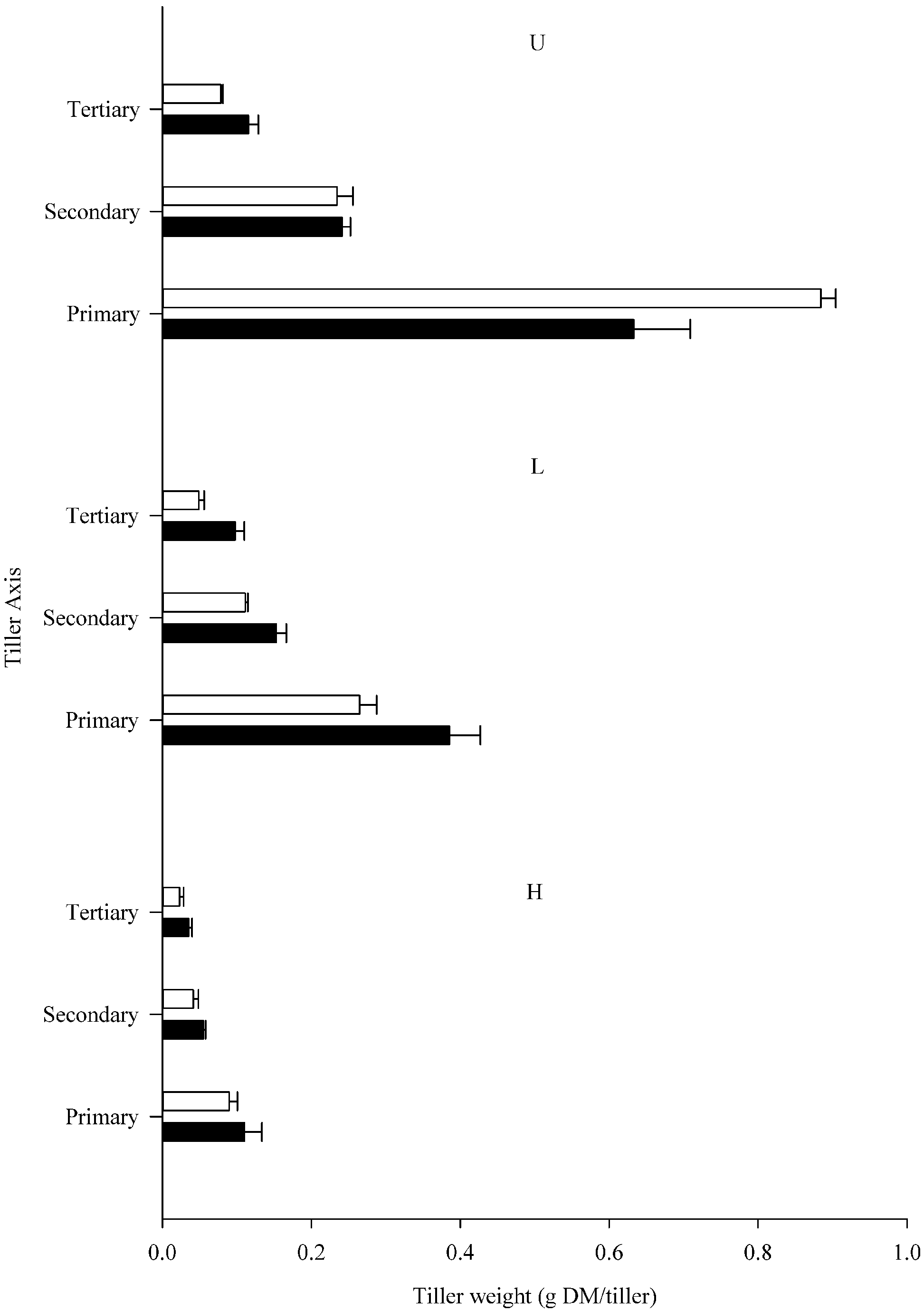
3.1. Plant Yield and Tiller Production

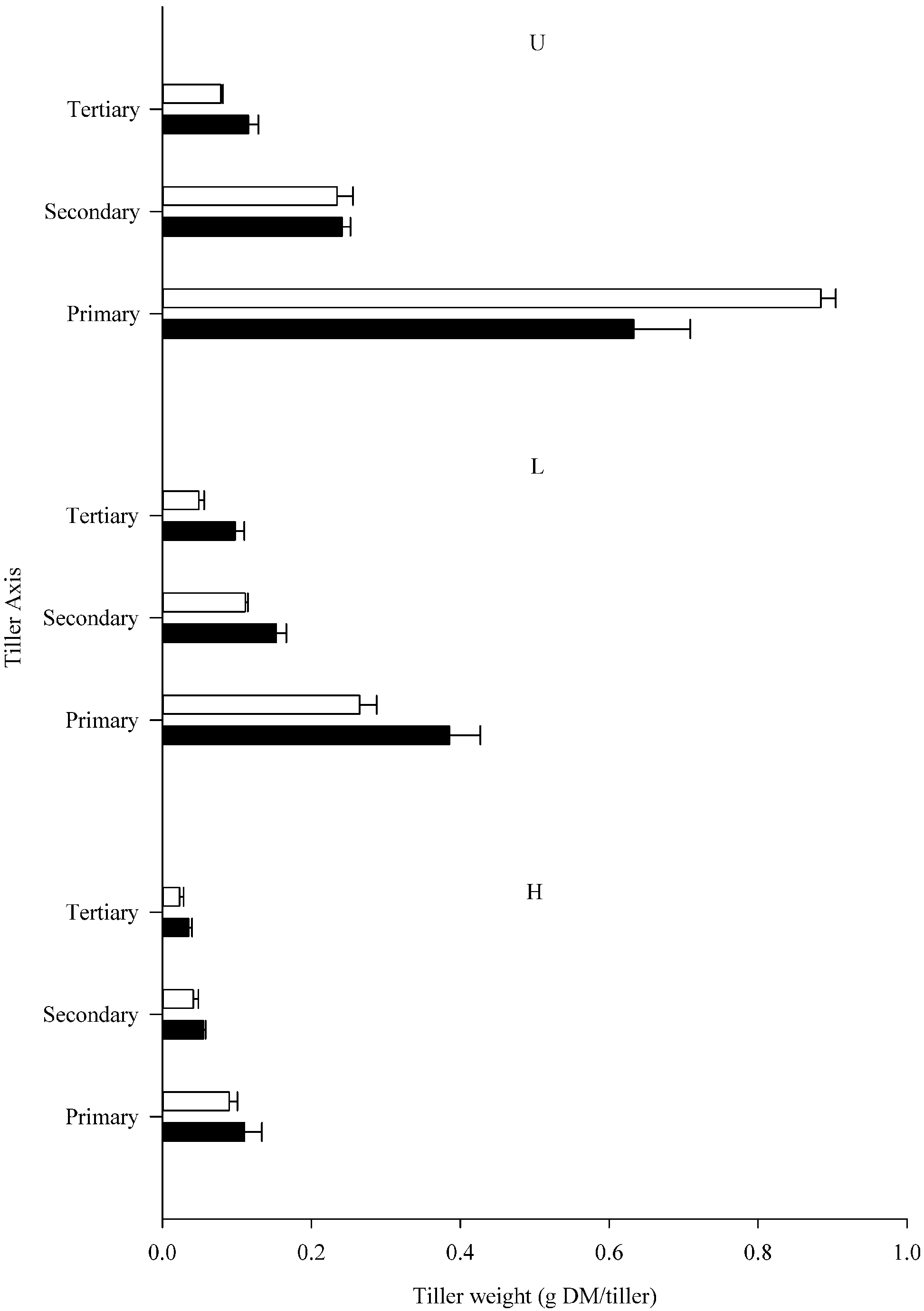{kind=link}
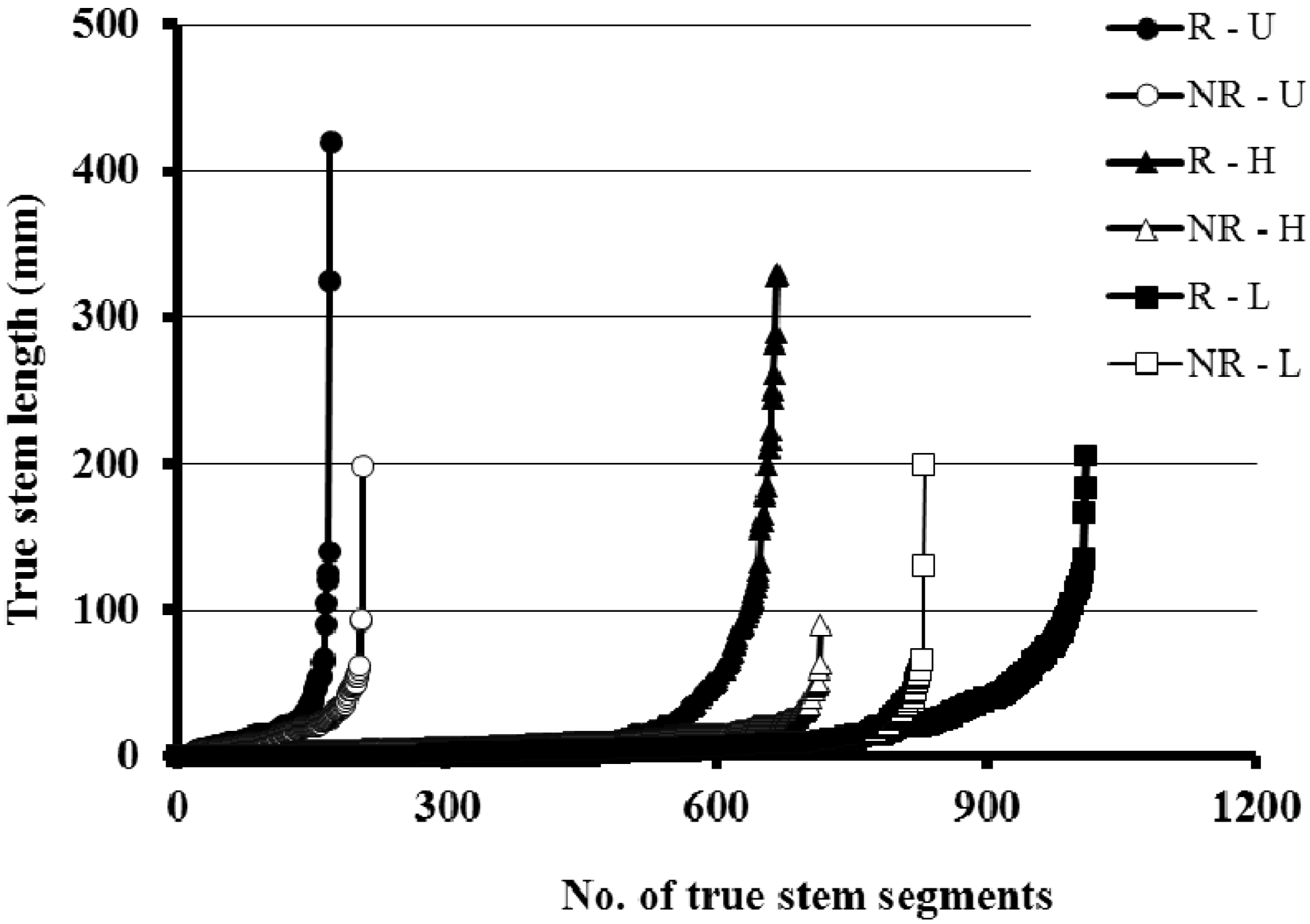{kind=link}
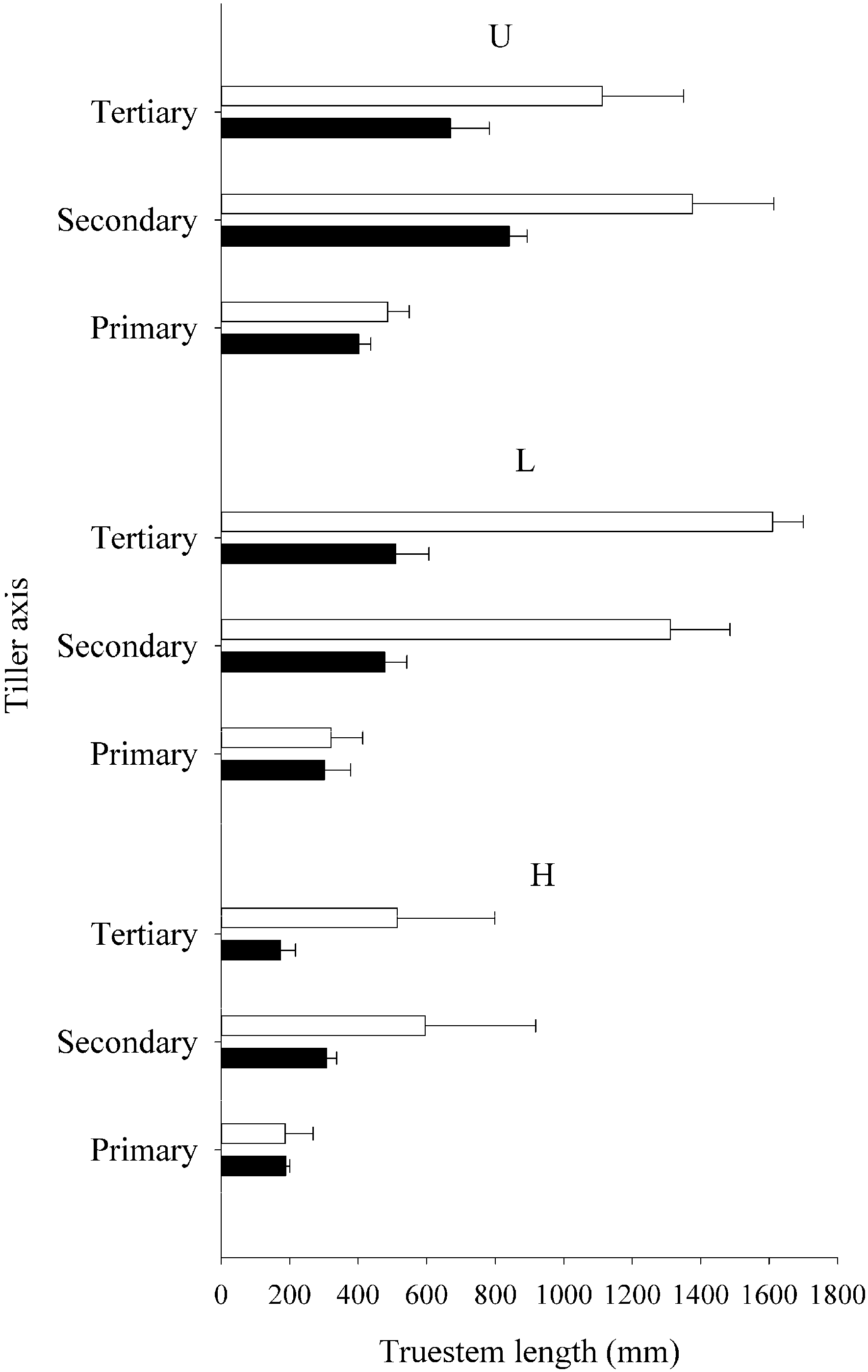{kind=link}
| Leaf | Pseudostem | True Stem | Root | Root:Shoot | Total | Tiller Weight | ||
|---|---|---|---|---|---|---|---|---|
| NR | U | 17.9 (43) | 11.4 (27) | 3.6 (8) | 9.2 (22) | 0.28 | 41.9 | 0.22 |
| L | 8.8 (41) | 5.9 (28) | 2.4 (11) | 4.7 (20) | 0.26 | 21.7 | 0.16 | |
| H | 2.7 (43) | 1.6 (27) | 0.7 (11) | 1.3 (21) | 0.27 | 6.4 | 0.06 | |
| Mean | 9.77 | 6.31 | 2.22 | 5.07 | 0.27 | 23.4 | 0.15 | |
| R | U | 15.2 (38) | 8.1 (20) | 4.3 (11) | 12.7 (32) | 0.47 | 40.4 | 0.21 |
| L | 6.2 (41) | 3.3 (22) | 1.9 (13) | 3.8 (25) | 0.33 | 15.3 | 0.09 | |
| H | 1.7 (42) | 1.0 (23) | 0.6 (15) | 0.8 (20) | 0.25 | 4.1 | 0.04 | |
| Mean | 7.73 | 4.13 | 2.29 | 5.77 | 0.35 | 19.9 | 0.12 | |
| SE | 0.80 | 0.47 | 0.32 | 0.77 | 0.032 | 2.01 | 0.0092 | |
| Signif | Popln | ** | *** | NS | NS | ** | * | *** |
| Cut | *** | *** | *** | *** | ** | *** | *** | |
| P × C | NS | * | NS | * | * | *,LT | ** |
| 12 Weeks | 18 Weeks | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Primary | Secondary | Tertiary | Total | Primary | Secondary | Tertiary | Total | ||
| NR | U | 16 | 45 | 44 | 104 | 19 | 71 | 102 | 192 |
| L | 15 | 42 | 20 | 77 | 16 | 57 | 57 | 130 | |
| H | 11 | 26 | 13 | 50 | 17 | 51 | 49 | 117 | |
| Mean | 14 | 38 | 25 | 77 | 17 | 60 | 69 | 146 | |
| R | U | 13 | 35 | 31 | 79 | 17 | 72 | 100 | 189 |
| L | 9 | 30 | 26 | 64 | 15 | 57 | 97 | 169 | |
| H | 8 | 23 | 19 | 49 | 11 | 38 | 56 | 105 | |
| Mean | 10 | 29 | 25 | 64 | 14 | 56 | 84 | 154 | |
| SEM | 7.8 | 1.3 | 4.0 | 4.1 | 1.6 | 5.4 | 9.8 | 12.8 | |
| Sig. | Popln | *** | * | NS | NS | * | NS | NS | NS |
| Cut | ** | ** | *** | *** | * | *** | *** | *** | |
| P × C | NS | NS | NS | NS | NS | NS | NS | NS | |
| Relative Tiller Appearance (No. obs) (Tillers/Tiller/Day) | Site Filling 1 | Site Usage 2 | ||||
|---|---|---|---|---|---|---|
| Overall | Initial | Final | ||||
| NR | U | 0.022 ± 0.0011 (17) | 0.030 ± 0.0011 (11) | 0.016 ± 0.0008 (6) | 0.16 | 0.17 |
| L | 0.021 ± 0.0005 (17) | N/A | 0.014 ± 0.0009 (6) | 0.14 | 0.15 | |
| H | 0.017 ± 0.0006 (17) | N/A | 0.020 ± 0.0011 (6) | 0.18 | 0.20 | |
| R | U | 0.025 ± 0.0007 (17) | 0.028 ± 0.006 (11) | 0.018 ± 0.0012 (6) | 0.30 | 0.35 |
| L | 0.024 ± 0.0004 (17) | N/A | 0.024 ± 0.0008 (6) | 0.30 | 0.35 | |
| H | 0.021 ± 0.0007 (17) | N/A | 0.027 ± 0.0013 (6) | 0.30 | 0.35 | |
3.2. Leaf Production Strategies
| Leaf area (cm2/Plant) | LER (mm/tlr/d) | SR (mm/tlr/d) | LAR (Leaves/d) | NL (Leaves/Tiller) | TiLL (mm) | LoLL (mm) | ||
|---|---|---|---|---|---|---|---|---|
| NR | U | 1805 | 16.2 | 16.6 | 0.10 | 5.8 | 846 | 191 |
| L | 954 | 12.6 | 9.1 | 0.10 | 6.5 | 463 | 147 | |
| H | 375 | 7.1 | 4.2 | 0.11 | 5.5 | 203 | 71 | |
| R | U | 1617 | 20.1 | 13.4 | 0.06 | 4.5 | 1247 | 364 |
| L | 543 | 18.0 | 10.7 | 0.08 | 4.8 | 461 | 232 | |
| H | 166 | 7.9 | 4.2 | 0.09 | 4.0 | 198 | 113 | |
| Sig. | SEM | 184.6 | 1.22 | 1.18 | 0.008 | 0.38 | 83.9 | 21.6 |
| Popln | NS | ** | NS | *** | *** | NS | *** | |
| Defol. | *** | *** | *** | NS | NS | *** | *** | |
| P × D | NS | NS | NS | NS | NS | * | * |
3.3. Rhizome Formation
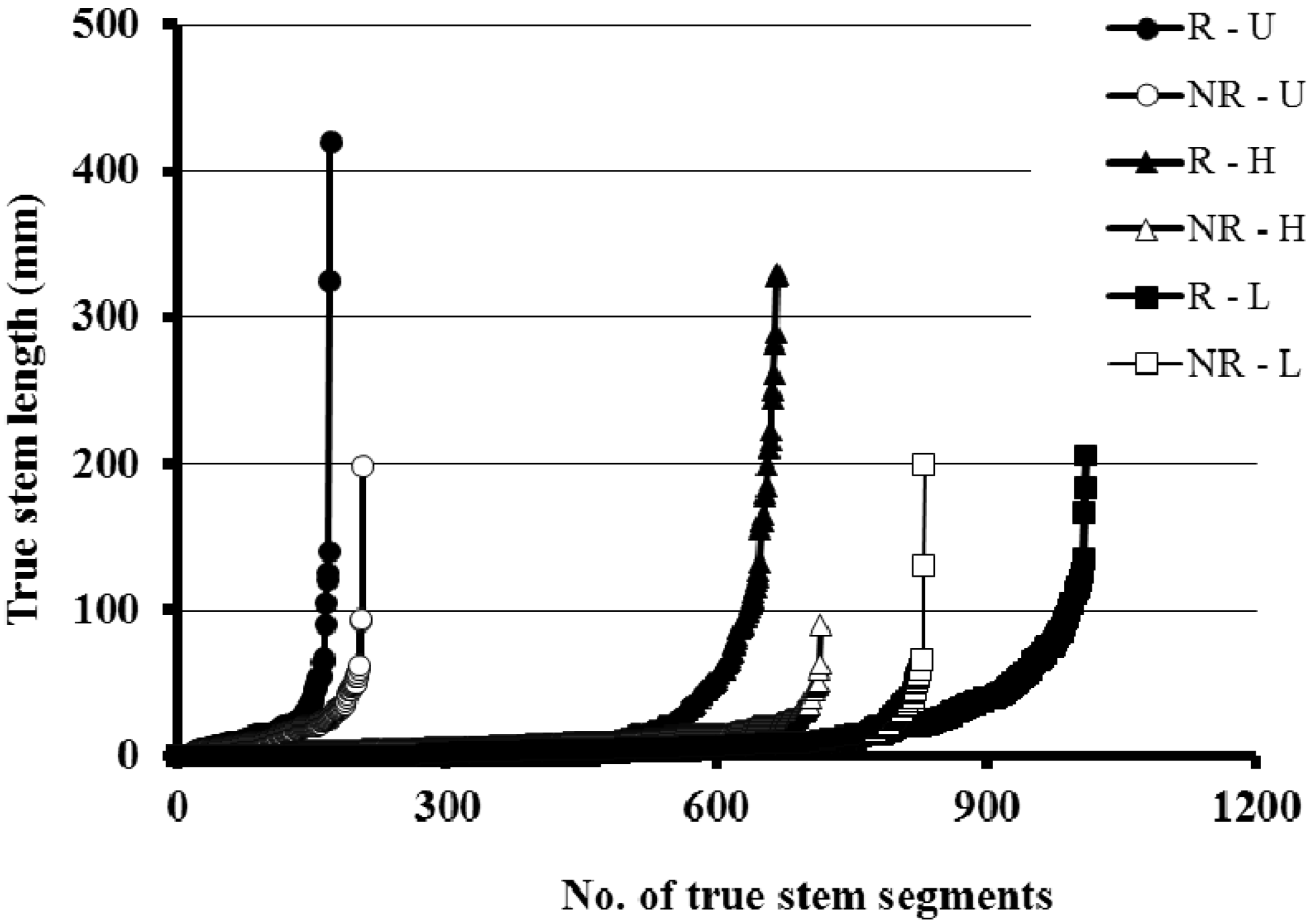
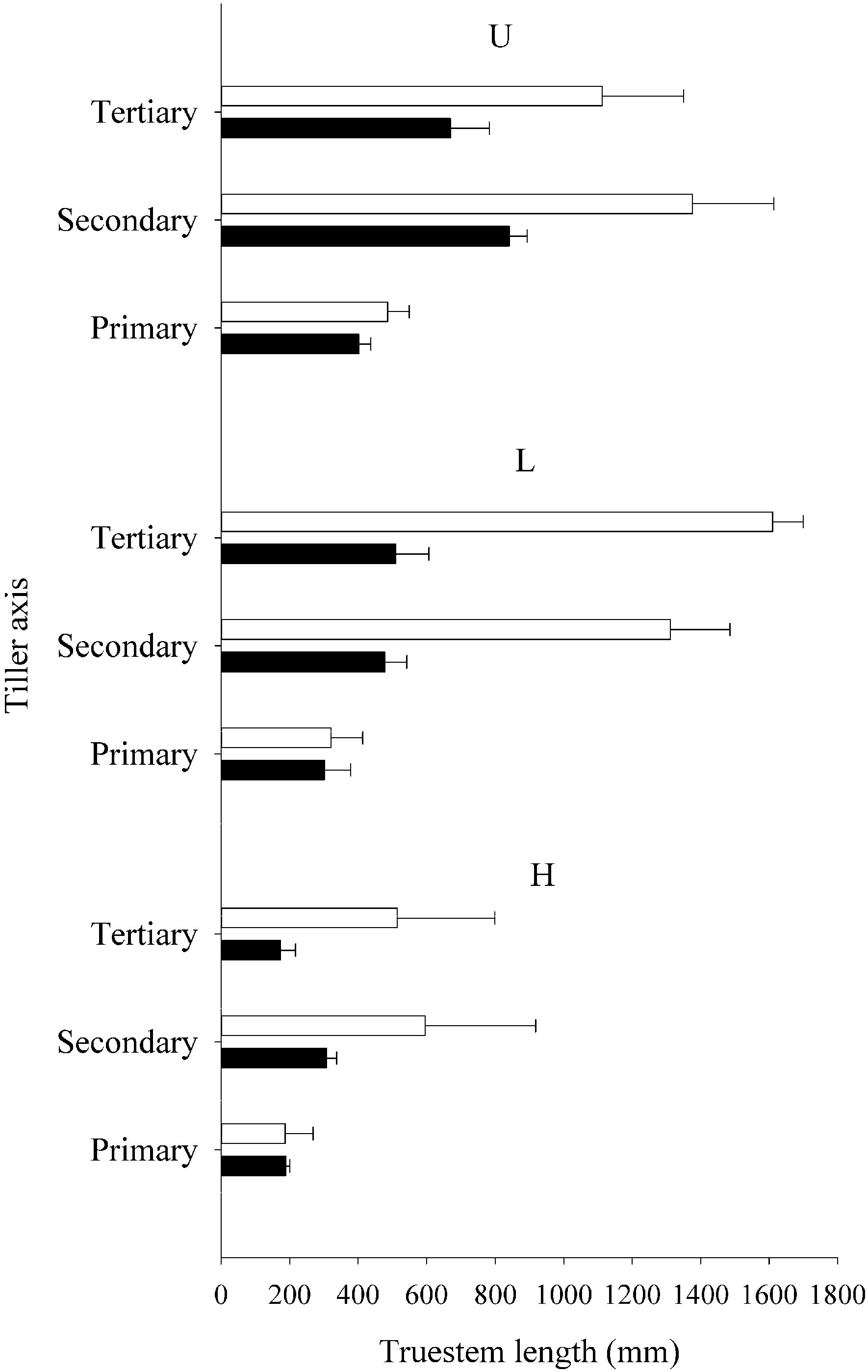
4. Discussion
4.1. Morphology of NR and R Tall Fescue Populations When Undefoliated
4.2. Response to Defoliation
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Belesky, D.P.; Fedders, J.M. Does endophyte influence the regrowth of tall fescue? Ann. Bot. 1996, 78, 499–505. [Google Scholar]
- Volenec, J.J.; Nelson, C.J. Responses of tall fescue leaf meristems to N fertilization and harvest frequency. Crop Sci. 1983, 23, 720–724. [Google Scholar] [CrossRef]
- Zarrough, K.M.; Nelson, C.J.; Sleper, D.A. Interrelationships between rates of leaf appearance and tillering in selected tall fescue populations. Crop Sci. 1984, 24, 565–569. [Google Scholar] [CrossRef]
- Brock, J.L.; Albrecht, K.A.; Hume, D.E. Stolons and rhizomes in tall fescue under grazing. Proc. N. Z. Grassland Assoc. 1997, 59, 93–98. [Google Scholar]
- Neuteboom, J.H.; Lantinga, E.A. Tillering potential and relationship between leaf and tiller production in perennial ryegrass. Ann. Bot. 1989, 63, 265–270. [Google Scholar]
- Richards, J.H. Physiology of plants recovering from defoliation. In Proceedings of the XVII International Grassland Congress, Palmerston North, New Zealand, 8–21 February 1993; pp. 85–94.
- Jernstedt, J.A.; Bouton, J.H. Anatomy, morphology, and growth of tall fescue rhizomes. Crop Sci. 1996, 36, 407–411. [Google Scholar]
- Matthew, C.; Assuero, S.G.; Black, C.K.; Sackville-Hamilton, N.R. Tiller dynamics of grazed swards. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., Moraes, A., Carvalho, P.C.F., Nabinger, C., Eds.; CAB International: New York, NY, USA, 2000; pp. 127–150. [Google Scholar]
- Hume, D.E.; Brock, J.L. Morphology of tall fescue (Festuca arundinacea) and perennial ryegrass (Lolium perenne) plants in pastures under sheep and cattle grazing. J. Agric. Sci. 1997, 129, 19–31. [Google Scholar] [CrossRef]
- De Battista, J.P.; Bouton, J.H. Greenhouse evaluation of tall fescue genotypes for rhizome production. Crop Sci. 1990, 30, 536–540. [Google Scholar] [CrossRef]
- Matthew, C.; Yang, J.Z.; Potter, J.F. Determination of tiller and root appearance in perennial ryegrass (Lolium perenne) swards by observation of the tiller axis, and potential application in mechanistic modelling. N. Z. J. Agric. Res. 1998, 41, 1–11. [Google Scholar] [CrossRef]
- Porter, H.L., Jr. Rhizomes in tall fescue. Agron. J. 1958, 50, 493–494. [Google Scholar] [CrossRef]
- Lopez, R.R.; Matches, A.G.; Baldridge, J.D. Vegetative development and organic reserves of tall fescue under conditions of accumulated growth. Crop Sci. 1967, 7, 409–412. [Google Scholar] [CrossRef]
- Bouton, J.H.; Smith, S.R., Jr.; Battista, J.P. Field screening for rhizome number in tall fescue. Crop Sci. 1992, 32, 686–689. [Google Scholar] [CrossRef]
- Ye, X.H.; Yu, F.H.; Dong, M. A trade-off between guerrilla and phalanx growth forms in Leymus secalinus under different nutrient supplies. Ann. Bot. 2006, 98, 187–191. [Google Scholar] [CrossRef] [PubMed]
- Herben, T.; Hara, T. Competition and spatial dynamics in clonal plants. In The Ecology and Evolution of Clonal Plants; Kroon, H., de van Groenendael, H., Eds.; Backhuys Publishers: Leiden, The Netherland, 1997; pp. 3331–3357. [Google Scholar]
- Thom, E.R.; Sheath, G.W.; Bryant, A.M. Seasonal-variations in total nonstructural carbohydrate and major element levels in perennial ryegrass and paspalum in a mixed pasture. N. Z. J. Agric. Res. 1989, 32, 157–165. [Google Scholar]
- Robson, M.J.; Ryle, G.J.A.; Woledge, J.; Nelson, C.J. The grass plant—Its form and function. In The Grass Crop. The Physiological Basis of Production; Jones, M.B., Lazenby, A., Eds.; Chapman and Hall: London, UK, 1988; pp. 25–83. [Google Scholar]
- Chapman, D.F.; Lemaire, G. Morphogenetic and structural determinants of plant regrowth following defoliation. In Proceedings of the XVII International Grassland Congress, Palmerston North, New Zealand, 8–21 February 1993; pp. 95–104.
- Nelson, C.J. Shoot morphological plasticity of grasses: Leaf growth versus tillering. In Grassland Ecophysiology and Grazing Ecology; Lemaire, G., Hodgson, J., Moraes, A., Carvalho, P.C.F., Nabinger, C., Eds.; CAB International: New York, NY, USA, 2000; pp. 101–126. [Google Scholar]
- Van Loo, E.N. Tillering, leaf expansion, and growth of plants of 2 cultivars of perennial ryegrass grown using hydroponics at 2 water potentials. Ann. Bot. 1992, 70, 511–518. [Google Scholar]
- Bahmani, I.; Hazard, L.; Varlet-Grancher, C.; Betin, M.; Lemaire, G.; Matthew, C.; Thom, E.R. Differences in tillering of long- and short-leaved perennial ryegrass genetic lines under full light and shade treatments. Crop Sci. 2000, 40, 1095–1102. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bryant, R.H.; Matthew, C.; Hodgson, J. Growth Strategy of Rhizomatous and Non-Rhizomatous Tall Fescue Populations in Response to Defoliation. Agriculture 2015, 5, 791-805. https://doi.org/10.3390/agriculture5030791
Bryant RH, Matthew C, Hodgson J. Growth Strategy of Rhizomatous and Non-Rhizomatous Tall Fescue Populations in Response to Defoliation. Agriculture. 2015; 5(3):791-805. https://doi.org/10.3390/agriculture5030791
Chicago/Turabian StyleBryant, Racheal H., Cory Matthew, and John Hodgson. 2015. "Growth Strategy of Rhizomatous and Non-Rhizomatous Tall Fescue Populations in Response to Defoliation" Agriculture 5, no. 3: 791-805. https://doi.org/10.3390/agriculture5030791
APA StyleBryant, R. H., Matthew, C., & Hodgson, J. (2015). Growth Strategy of Rhizomatous and Non-Rhizomatous Tall Fescue Populations in Response to Defoliation. Agriculture, 5(3), 791-805. https://doi.org/10.3390/agriculture5030791