
Unveiling the Impact of Soil Prebiotics on Rhizospheric Microbial Functionality in Zea mays L.

 ,
,  ,
,  , , ,
, , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Harvesting
2.2. Total Soil DNA Extraction and Quantification of Microbial Abundance
2.3. Enzymatic Activities Analysis
- Soil fertility index (Mw) reflects the soil fertility state where the higher the Mw, the more fertile the soil [50]. It was calculated according to this formula:
- 2.
- Geometric mean (Gmea) presents the soil functional diversity, with elevated Gmea corresponding to high functional diversity [51]. It was calculated according to this formula:
2.4. Metabolic Profiling of Soil Microbial Communities
2.5. Statistical Analysis
3. Results
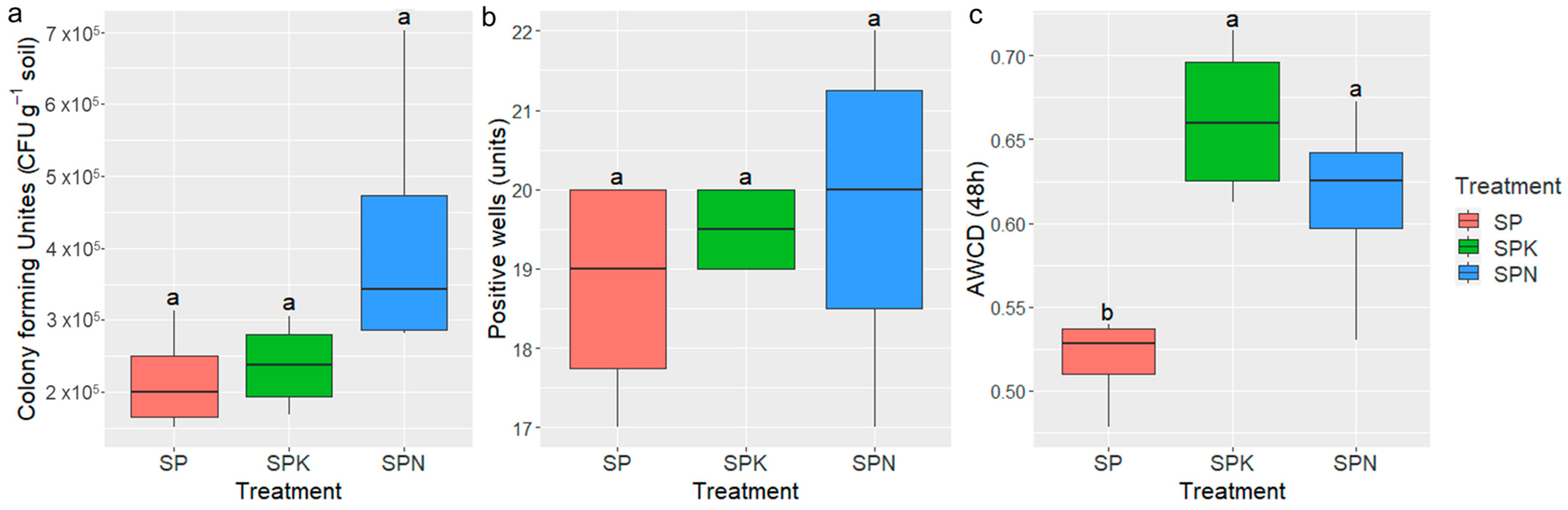
3.1. Effect of Prebiotics on Soil Microbial Abundance
3.2. Effect of Prebiotics on Enzyme Activities in Soil
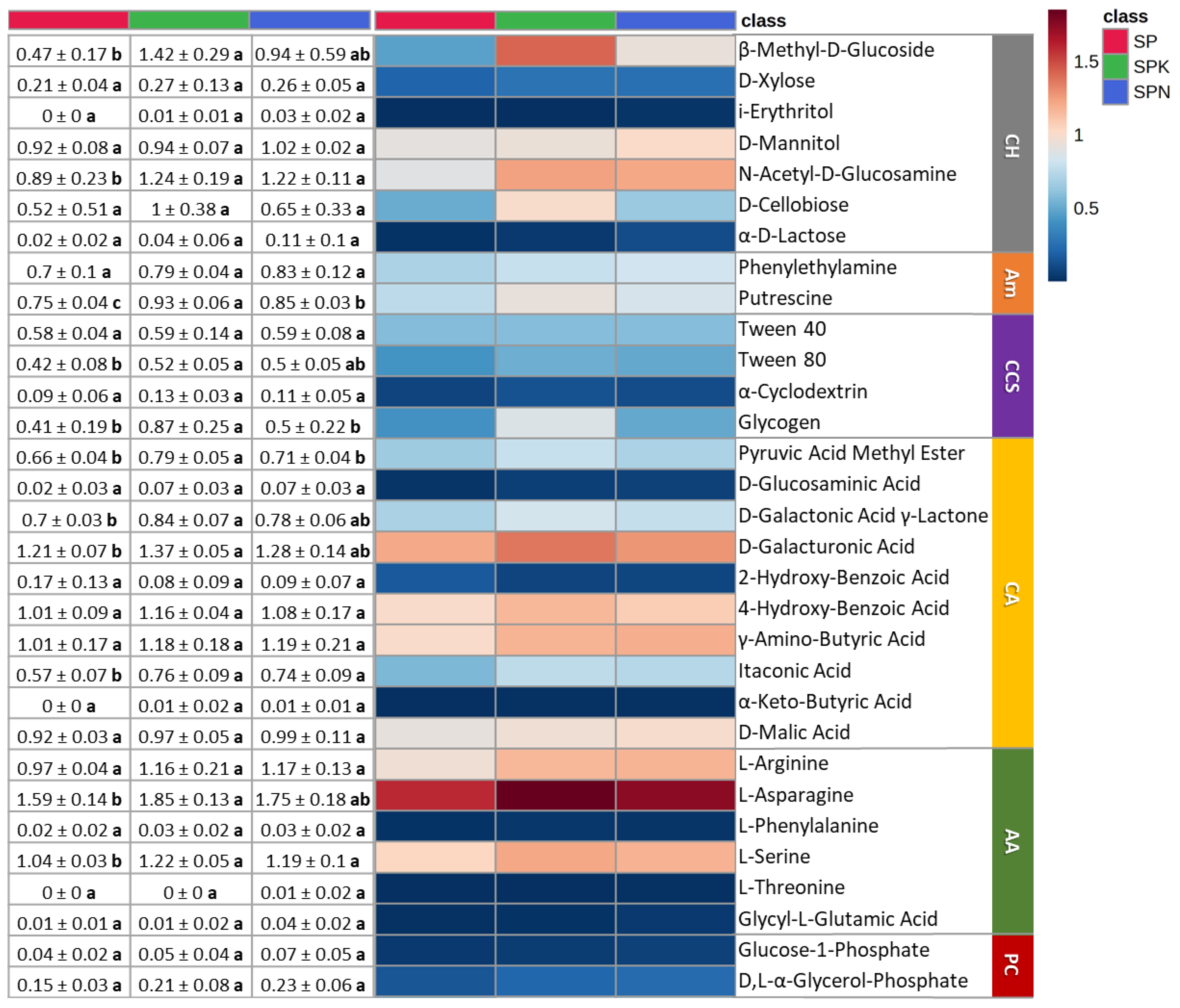
3.3. Effect of Prebiotics on the Metabolic Pattern in Soil
4. Discussion
4.1. Prebiotics’ Application Increase Soil Enzymatic Activity
4.2. Prebiotics’ Application Increases the Abundance of Soil Fungi and the Metabolic Activity of Heterotrophic Culturable Soil Bacteria
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Caradonia, F.; Battaglia, V.; Righi, L.; Pascali, G.; La Torre, A. Plant Biostimulant Regulatory Framework: Prospects in Europe and Current Situation at International Level. J. Plant Growth Regul. 2019, 38, 438–448. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Editorial: Biostimulants in Agriculture. Front. Plant Sci. 2020, 11, 40. [Google Scholar] [CrossRef] [PubMed]
- Bulgari, R.; Cocetta, G.; Trivellini, A.; Vernieri, P.; Ferrante, A. Biostimulants and Crop Responses: A Review. Biol. Agric. Hortic. 2015, 31, 1–17. [Google Scholar] [CrossRef]
- Therond, O.; Debril, T.; Duru, M.; Magrini, M.-B.; Plumecocq, G.; Sarthou, J.-P. Socio-Economic Characterisation of Agriculture Models. In Agroecological Transitions: From Theory to Practice in Local Participatory Design; Bergez, J.-E., Audouin, E., Therond, O., Eds.; Springer International Publishing: Cham, Switzerland, 2019; pp. 21–43. ISBN 978-3-030-01953-2. [Google Scholar]
- Calvo, P.; Nelson, L.; Kloepper, J.W. Agricultural Uses of Plant Biostimulants. Plant Soil 2014, 383, 3–41. [Google Scholar] [CrossRef]
- Albrecht, U. Plant Biostimulants: Definition and Overview of Categories and Effects. EDIS 2019, 2019-3. [Google Scholar] [CrossRef]
- Yakhin, O.I.; Lubyanov, A.A.; Yakhin, I.A.; Brown, P.H. Biostimulants in Plant Science: A Global Perspective. Front. Plant Sci. 2017, 7, 2049. [Google Scholar] [CrossRef] [PubMed]
- Ricci, M.; Tilbury, L.; Daridon, B.; Sukalac, K. General Principles to Justify Plant Biostimulant Claims. Front. Plant Sci. 2019, 10, 494. [Google Scholar] [CrossRef] [PubMed]
- Vassileva, M.; Flor-Peregrin, E.; Malusá, E.; Vassilev, N. Towards Better Understanding of the Interactions and Efficient Application of Plant Beneficial Prebiotics, Probiotics, Postbiotics and Synbiotics. Front. Plant Sci. 2020, 11, 1068. [Google Scholar] [CrossRef] [PubMed]
- Berg, S.; Dennis, P.G.; Paungfoo-Lonhienne, C.; Anderson, J.; Robinson, N.; Brackin, R.; Royle, A.; DiBella, L.; Schmidt, S. Effects of Commercial Microbial Biostimulants on Soil and Root Microbial Communities and Sugarcane Yield. Biol. Fertil. Soils 2020, 56, 565–580. [Google Scholar] [CrossRef]
- Colla, G.; Hoagland, L.; Ruzzi, M.; Cardarelli, M.; Bonini, P.; Canaguier, R.; Rouphael, Y. Biostimulant Action of Protein Hydrolysates: Unraveling Their Effects on Plant Physiology and Microbiome. Front. Plant Sci. 2017, 8, 2202. [Google Scholar] [CrossRef]
- Nosheen, S.; Ajmal, I.; Song, Y. Microbes as Biofertilizers, a Potential Approach for Sustainable Crop Production. Sustainability 2021, 13, 1868. [Google Scholar] [CrossRef]
- du Jardin, P. Plant Biostimulants: Definition, Concept, Main Categories and Regulation. Sci. Hortic. 2015, 196, 3–14. [Google Scholar] [CrossRef]
- Wadduwage, J.; Liu, H.; Egidi, E.; Singh, B.K.; Macdonald, C.A. Effects of Biostimulant Application on Soil Biological and Physicochemical Properties: A Field Study. J. Sustain. Agric. Environ. 2023, 2, 285–300. [Google Scholar] [CrossRef]
- Hellequin, E.; Monard, C.; Chorin, M.; Le Bris, N.; Daburon, V.; Klarzynski, O.; Binet, F. Responses of Active Soil Microorganisms Facing to a Soil Biostimulant Input Compared to Plant Legacy Effects. Sci. Rep. 2020, 10, 13727. [Google Scholar] [CrossRef] [PubMed]
- Hellequin, E.; Monard, C.; Quaiser, A.; Henriot, M.; Klarzynski, O.; Binet, F. Specific Recruitment of Soil Bacteria and Fungi Decomposers Following a Biostimulant Application Increased Crop Residues Mineralization. PLoS ONE 2018, 13, e0209089. [Google Scholar] [CrossRef] [PubMed]
- Arif, I.; Batool, M.; Schenk, P.M. Plant Microbiome Engineering: Expected Benefits for Improved Crop Growth and Resilience. Trends Biotechnol. 2020, 38, 1385–1396. [Google Scholar] [CrossRef] [PubMed]
- Palese, A.M.; Pane, C.; Villecco, D.; Zaccardelli, M.; Altieri, G.; Celano, G. Effects of Organic Additives on Chemical, Microbiological and Plant Pathogen Suppressive Properties of Aerated Municipal Waste Compost Teas. Appl. Sci. 2021, 11, 7402. [Google Scholar] [CrossRef]
- Mattarozzi, M.; Di Zinno, J.; Montanini, B.; Manfredi, M.; Marengo, E.; Fornasier, F.; Ferrarini, A.; Careri, M.; Visioli, G. Biostimulants Applied to Maize Seeds Modulate the Enzymatic Activity and Metaproteome of the Rhizosphere. Appl. Soil Ecol. 2020, 148, 103480. [Google Scholar] [CrossRef]
- Pizzeghello, D.; Schiavon, M.; Francioso, O.; Dalla Vecchia, F.; Ertani, A.; Nardi, S. Bioactivity of Size-Fractionated and Unfractionated Humic Substances from Two Forest Soils and Comparative Effects on N and S Metabolism, Nutrition, and Root Anatomy of Allium sativum L. Front. Plant Sci. 2020, 11, 1203. [Google Scholar] [CrossRef]
- Rashid, N.; Khan, S.; Wahid, A.; Ibrar, D.; Hasnain, Z.; Irshad, S.; Bashir, S.; Al-Hashimi, A.; Elshikh, M.S.; Kamran, M.; et al. Exogenous Application of Biostimulants and Synthetic Growth Promoters Improved the Productivity and Grain Quality of Quinoa Linked with Enhanced Photosynthetic Pigments and Metabolomics. Agronomy 2021, 11, 2302. [Google Scholar] [CrossRef]
- Rezgui, C.; Trinsoutrot-gattin, I.; Benoit, M.; Laval, K.; Riah-anglet, W. Linking Changes in the Soil Microbial Community to C and N Dynamics during Crop Residue Decomposition. J. Integr. Agric. 2021, 20, 3039–3059. [Google Scholar] [CrossRef]
- Antón-Herrero, R.; Vega-Jara, L.; García-Delgado, C.; Mayans, B.; Camacho-Arévalo, R.; Moreno-Jiménez, E.; Plaza, C.; Eymar, E. Synergistic Effects of Biochar and Biostimulants on Nutrient and Toxic Element Uptake by Pepper in Contaminated Soils. J. Sci. Food Agric. 2022, 102, 167–174. [Google Scholar] [CrossRef] [PubMed]
- Bello, A.S.; Saadaoui, I.; Ben-Hamadou, R. “Beyond the Source of Bioenergy”: Microalgae in Modern Agriculture as a Biostimulant, Biofertilizer, and Anti-Abiotic Stress. Agronomy 2021, 11, 1610. [Google Scholar] [CrossRef]
- Ertani, A.; Nardi, S.; Francioso, O.; Pizzeghello, D.; Tinti, A.; Schiavon, M. Metabolite-Targeted Analysis and Physiological Traits of Zea mays L. in Response to Application of a Leonardite-Humate and Lignosulfonate-Based Products for Their Evaluation as Potential Biostimulants. Agronomy 2019, 9, 445. [Google Scholar] [CrossRef]
- Medina-Herrera, M.d.R.; Negrete-Rodríguez, M.d.l.L.X.; Álvarez-Trejo, J.L.; Samaniego-Hernández, M.; González-Cruz, L.; Bernardino-Nicanor, A.; Conde-Barajas, E. Evaluation of Non-Conventional Biological and Molecular Parameters as Potential Indicators of Quality and Functionality of Urban Biosolids Used as Organic Amendments of Agricultural Soils. Appl. Sci. 2020, 10, 517. [Google Scholar] [CrossRef]
- Song, Z.; Bi, Y.; Zhang, J.; Gong, Y.; Yang, H. Arbuscular Mycorrhizal Fungi Promote the Growth of Plants in the Mining Associated Clay. Sci. Rep. 2020, 10, 2663. [Google Scholar] [CrossRef] [PubMed]
- Ferraz-Almeida, R.; Naves, E.; Mota, R. Soil Quality: Enzymatic Activity of Soil β-Glucosidase. Glob. J. Agric. Res. Rev. 2015, 3, 146–450. [Google Scholar]
- Sinsabaugh, R.L.; Manzoni, S.; Moorhead, D.L.; Richter, A. Carbon Use Efficiency of Microbial Communities: Stoichiometry, Methodology and Modelling. Ecol. Lett. 2013, 16, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Utobo, E.B.; Tewari, L. Soil Enzymes as Bioindicators of Soil Ecosystem Status. Appl. Ecol. Environ. Res. 2015, 13, 147–169. [Google Scholar] [CrossRef]
- Weber, K.P.; Legge, R.L. Community-Level Physiological Profiling. In Bioremediation: Methods and Protocols; Cummings, S.P., Ed.; Methods in Molecular Biology; Humana Press: Totowa, NJ, USA, 2010; pp. 263–281. ISBN 978-1-60761-439-5. [Google Scholar]
- Adedayo, A.A.; Fadiji, A.E.; Babalola, O.O. Quantifying the Respiratory Pattern of Rhizosphere Microbial Communities in Healthy and Diseased Tomato Plants Using Carbon Substrates. J. Soil Sci. Plant Nutr. 2023, 23, 6485–6496. [Google Scholar] [CrossRef]
- Frąc, M.; Oszust, K.; Lipiec, J.; Jezierska-Tys, S.; Nwaichi, E.O. Soil Microbial Functional and Fungal Diversity as Influenced by Municipal Sewage Sludge Accumulation. Int. J. Environ. Res. Public Health 2014, 11, 8891–8908. [Google Scholar] [CrossRef] [PubMed]
- Jacobs-Hoffman, I.; Hills, P.N. Effects of the Commercial Biostimulant BC204 on the Rhizosphere Microbial Community of Solanum lycopersicum L. S. Afr. J. Bot. 2021, 143, 52–60. [Google Scholar] [CrossRef]
- Gałązka, A.; Grządziel, J. Fungal Genetics and Functional Diversity of Microbial Communities in the Soil under Long-Term Monoculture of Maize Using Different Cultivation Techniques. Front. Microbiol. 2018, 9, 76. [Google Scholar] [CrossRef] [PubMed]
- Classen, A.T.; Boyle, S.I.; Haskins, K.E.; Overby, S.T.; Hart, S.C. Community-Level Physiological Profiles of Bacteria and Fungi: Plate Type and Incubation Temperature Influences on Contrasting Soils. FEMS Microbiol. Ecol. 2003, 44, 319–328. [Google Scholar] [CrossRef] [PubMed]
- Paul, K.; Sorrentino, M.; Lucini, L.; Rouphael, Y.; Cardarelli, M.; Bonini, P.; Reynaud, H.; Canaguier, R.; Trtílek, M.; Panzarová, K.; et al. Understanding the Biostimulant Action of Vegetal-Derived Protein Hydrolysates by High-Throughput Plant Phenotyping and Metabolomics: A Case Study on Tomato. Front. Plant Sci. 2019, 10, 47. [Google Scholar] [CrossRef] [PubMed]
- Vidak, M.; Lazarević, B.; Petek, M.; Gunjača, J.; Šatović, Z.; Budor, I.; Carović-Stanko, K. Multispectral Assessment of Sweet Pepper (Capsicum annuum L.) Fruit Quality Affected by Calcite Nanoparticles. Biomolecules 2021, 11, 832. [Google Scholar] [CrossRef]
- Alahmad, A.; Edelman, L.; Castel, L.; Bernardon-Mery, A.; Laval, K.; Trinsoutrot-Gattin, I.; Thioye, B. Prebiotics: A Solution for Improving Plant Growth, Soil Health, and Carbon Sequestration? J. Soil Sci. Plant Nutr. 2023, 23, 6647–6669. [Google Scholar] [CrossRef]
- Shahrajabian, M.H.; Chaski, C.; Polyzos, N.; Petropoulos, S.A. Biostimulants Application: A Low Input Cropping Management Tool for Sustainable Farming of Vegetables. Biomolecules 2021, 11, 698. [Google Scholar] [CrossRef]
- Gangneux, C.C.; Akpa-Vinceslas, M.M.; Sauvage, H.H.; Desaire, S.S.; Houot, S.S.; Laval, K.K. Fungal, Bacterial and Plant dsDNA Contributions to Soil Total DNA Extracted from Silty Soils under Different Farming Practices: Relationships with Chloroform-Labile Carbon. Soil Biol. Biochem. 2011, 43, 431–437. [Google Scholar] [CrossRef]
- Marchesi, J.R.; Sato, T.; Weightman, A.J.; Martin, T.A.; Fry, J.C.; Hiom, S.J.; Dymock, D.; Wade, W.G. Design and Evaluation of Useful Bacterium-Specific PCR Primers That Amplify Genes Coding for Bacterial 16S rRNA. Appl. Environ. Microbiol. 1998, 64, 795–799. [Google Scholar] [CrossRef]
- Muyzer, G.; de Waal, E.C.; Uitterlinden, A.G. Profiling of Complex Microbial Populations by Denaturing Gradient Gel Electrophoresis Analysis of Polymerase Chain Reaction-Amplified Genes Coding for 16S rRNA. Appl. Environ. Microbiol. 1993, 59, 695–700. [Google Scholar] [CrossRef] [PubMed]
- Borneman, J.; Hartin, R.J. PCR Primers That Amplify Fungal rRNA Genes from Environmental Samples. Appl. Environ. Microbiol. 2000, 66, 4356–4360. [Google Scholar] [CrossRef] [PubMed]
- ISO 20130:2018; Soil Quality—Measurement of Enzyme Activity Patterns in Soil Samples Using Colorimetric Substrates in Micro-Well Plates. Association Française de Normalisation (AFNOR): La Plaine Saint-Denis, France, 2018.
- ISO/TS 22939:2019; Soil Quality—Measurement of Enzyme Activity Patterns in Soil Samples Using Fluorogenic Substrates in Micro-Well Plates. Association Française de Normalisation (AFNOR): La Plaine Saint-Denis, France, 2019.
- Kanté, M.; Riah-Anglet, W.; Cliquet, J.-B.; Trinsoutrot-Gattin, I. Soil Enzyme Activity and Stoichiometry: Linking Soil Microorganism Resource Requirement and Legume Carbon Rhizodeposition. Agronomy 2021, 11, 2131. [Google Scholar] [CrossRef]
- Riah-Anglet, W.; Cusset, E.; Chaussod, R.; Criquet, S.; Norini, M.-P.; Cheviron, N.; Mougin, C.; Laval, K.; Latour, X.; Legras, M.; et al. Introducing Grasslands into Crop Rotations, a Way to Restore Microbiodiversity and Soil Functions. Agriculture 2021, 11, 909. [Google Scholar] [CrossRef]
- Puglisi, E.; Del Re, A.A.M.; Rao, M.A.; Gianfreda, L. Development and Validation of Numerical Indexes Integrating Enzyme Activities of Soils. Soil Biol. Biochem. 2006, 38, 1673–1681. [Google Scholar] [CrossRef]
- Kompała-Bąba, A.; Bierza, W.; Sierka, E.; Błońska, A.; Besenyei, L.; Woźniak, G. The Role of Plants and Soil Properties in the Enzyme Activities of Substrates on Hard Coal Mine Spoil Heaps. Sci. Rep. 2021, 11, 5155. [Google Scholar] [CrossRef] [PubMed]
- Hinojosa, M.B.; Carreira, J.A.; García-Ruíz, R.; Dick, R.P. Soil Moisture Pre-Treatment Effects on Enzyme Activities as Indicators of Heavy Metal-Contaminated and Reclaimed Soils. Soil Biol. Biochem. 2004, 36, 1559–1568. [Google Scholar] [CrossRef]
- Garland, J.L. Analysis and Interpretation of Community-Level Physiological Profiles in Microbial Ecology. FEMS Microbiol. Ecol. 1997, 24, 289–300. [Google Scholar] [CrossRef]
- Calbrix, R.; Laval, K.; Barray, S. Analysis of the Potential Functional Diversity of the Bacterial Community in Soil: A Reproducible Procedure Using Sole-Carbon-Source Utilization Profiles. Eur. J. Soil Biol. 2005, 41, 11–20. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- de Mendiburu, F.; Yaseen, M. Agricolae: Statistical Procedures for Agricultural Research; R Package Version 1.3-3. 2020. Available online: https://www.cran.r-project.org/web/packages/agricolae/index.html (accessed on 28 May 2024).
- Pang, Z.; Chong, J.; Zhou, G.; de Lima Morais, D.A.; Chang, L.; Barrette, M.; Gauthier, C.; Jacques, P.-É.; Li, S.; Xia, J. MetaboAnalyst 5.0: Narrowing the Gap between Raw Spectra and Functional Insights. Nucleic Acids Res. 2021, 49, W388–W396. [Google Scholar] [CrossRef]
- Xia, J.; Psychogios, N.; Young, N.; Wishart, D.S. MetaboAnalyst: A Web Server for Metabolomic Data Analysis and Interpretation. Nucleic Acids Res. 2009, 37, W652–W660. [Google Scholar] [CrossRef]
- Xia, J.; Wishart, D.S. Metabolomic Data Processing, Analysis, and Interpretation Using MetaboAnalyst. Curr. Protoc. Bioinform. 2011, 34, 14.10.1–14.10.48. [Google Scholar] [CrossRef] [PubMed]
- Pang, Z.; Zhou, G.; Ewald, J.; Chang, L.; Hacariz, O.; Basu, N.; Xia, J. Using MetaboAnalyst 5.0 for LC–HRMS Spectra Processing, Multi-Omics Integration and Covariate Adjustment of Global Metabolomics Data. Nat. Protoc. 2022, 17, 1735–1761. [Google Scholar] [CrossRef]
- Chong, J.; Xia, J. MetaboAnalystR: An R Package for Flexible and Reproducible Analysis of Metabolomics Data. Bioinformatics 2018, 34, 4313–4314. [Google Scholar] [CrossRef]
- Elbl, J.; Maková, J.; Javoreková, S.; Medo, J.; Kintl, A.; Lošák, T.; Lukas, V. Response of Microbial Activities in Soil to Various Organic and Mineral Amendments as an Indicator of Soil Quality. Agronomy 2019, 9, 485. [Google Scholar] [CrossRef]
- Macias-Benitez, S.; Garcia-Martinez, A.M.; Caballero Jimenez, P.; Gonzalez, J.M.; Tejada Moral, M.; Parrado Rubio, J. Rhizospheric Organic Acids as Biostimulants: Monitoring Feedbacks on Soil Microorganisms and Biochemical Properties. Front. Plant Sci. 2020, 11, 633. [Google Scholar] [CrossRef]
- Tejada, M.; Benítez, C.; Gómez, I.; Parrado, J. Use of Biostimulants on Soil Restoration: Effects on Soil Biochemical Properties and Microbial Community. Appl. Soil Ecol. 2011, 49, 11–17. [Google Scholar] [CrossRef]
- Baćmaga, M.; Wyszkowska, J.; Kucharski, J. Biostimulation as a Process Aiding Tebuconazole Degradation in Soil. J. Soils Sediments 2019, 19, 3728–3741. [Google Scholar] [CrossRef]
- Halpern, M.; Bar-Tal, A.; Ofek, M.; Minz, D.; Muller, T.; Yermiyahu, U. Chapter Two—The Use of Biostimulants for Enhancing Nutrient Uptake. In Advances in Agronomy; Sparks, D.L., Ed.; Academic Press: Cambridge, MA, USA, 2015; Volume 130, pp. 141–174. [Google Scholar] [CrossRef]
- Nath, D.; Ozah, B.; Baruah, R.; Barooah, R.C.; —Borah, D.K. Effect of Integrated Nutrient Management on Soil Enzymes, Microbial Biomass Carbon and Bacterial Populations under Rice (Oryza sativa)-Wheat (Triticum aestivum) Sequence. Indian J. Agric. Sci. 2011, 81, 1143–1148. [Google Scholar]
- García-Martínez, A.M.; Díaz, A.; Tejada, M.; Bautista, J.; Rodríguez, B.; Santa María, C.; Revilla, E.; Parrado, J. Enzymatic Production of an Organic Soil Biostimulant from Wheat-Condensed Distiller Solubles: Effects on Soil Biochemistry and Biodiversity. Process Biochem. 2010, 45, 1127–1133. [Google Scholar] [CrossRef]
- Mueller, S.R.; Kussow, W. Biostimulant Influences on Turfgrass Microbial Communities and Creeping Bentgrass Putting Green Quality. HortScience 2005, 40, 1904–1910. [Google Scholar] [CrossRef]
- Onet, A.; Dincă, L.C.; Grenni, P.; Laslo, V.; Teusdea, A.C.; Vasile, D.L.; Enescu, R.E.; Crisan, V.E. Biological Indicators for Evaluating Soil Quality Improvement in a Soil Degraded by Erosion Processes. J. Soils Sediments 2019, 19, 2393–2404. [Google Scholar] [CrossRef]
- Lauber, C.L.; Strickland, M.S.; Bradford, M.A.; Fierer, N. The Influence of Soil Properties on the Structure of Bacterial and Fungal Communities across Land-Use Types. Soil Biol. Biochem. 2008, 40, 2407–2415. [Google Scholar] [CrossRef]
- Philippot, L.; Raaijmakers, J.M.; Lemanceau, P.; van der Putten, W.H. Going Back to the Roots: The Microbial Ecology of the Rhizosphere. Nat. Rev. Microbiol. 2013, 11, 789–799. [Google Scholar] [CrossRef] [PubMed]
- Adedayo, A.A.; Babalola, O.O. The Potential of Biostimulants on Soil Microbial Community: A Review. Front. Ind. Microbiol. 2023, 1, 1308641. [Google Scholar] [CrossRef]
- Siwik-Ziomek, A.; Szczepanek, M. Soil Extracellular Enzyme Activities and Uptake of N by Oilseed Rape Depending on Fertilization and Seaweed Biostimulant Application. Agronomy 2019, 9, 480. [Google Scholar] [CrossRef]
- Tejada, M.; Rodríguez-Morgado, B.; Gómez, I.; Parrado, J. Degradation of Chlorpyrifos Using Different Biostimulants/Biofertilizers: Effects on Soil Biochemical Properties and Microbial Community. Appl. Soil Ecol. 2014, 84, 158–165. [Google Scholar] [CrossRef]
- Tejada, M.; García-Martínez, A.M.; Gómez, I.; Parrado, J. Application of MCPA Herbicide on Soils Amended with Biostimulants: Short-Time Effects on Soil Biological Properties. Chemosphere 2010, 80, 1088–1094. [Google Scholar] [CrossRef]
- Abbott, L.K.; Macdonald, L.M.; Wong, M.T.F.; Webb, M.J.; Jenkins, S.N.; Farrell, M. Potential Roles of Biological Amendments for Profitable Grain Production—A Review. Agric. Ecosyst. Environ. 2018, 256, 34–50. [Google Scholar] [CrossRef]
- Rehim, A.; Amjad Bashir, M.; Raza, Q.-U.-A.; Gallagher, K.; Berlyn, G.P. Yield Enhancement of Biostimulants, Vitamin B12, and CoQ10 Compared to Inorganic Fertilizer in Radish. Agronomy 2021, 11, 697. [Google Scholar] [CrossRef]
- Grammenou, A.; Petropoulos, S.A.; Thalassinos, G.; Rinklebe, J.; Shaheen, S.M.; Antoniadis, V. Biostimulants in the Soil–Plant Interface: Agro-Environmental Implications—A Review. Earth Syst. Environ. 2023, 7, 583–600. [Google Scholar] [CrossRef]
- Mandal, S.; Anand, U.; López-Bucio, J.; Radha; Kumar, M.; Lal, M.K.; Tiwari, R.K.; Dey, A. Biostimulants and Environmental Stress Mitigation in Crops: A Novel and Emerging Approach for Agricultural Sustainability under Climate Change. Environ. Res. 2023, 233, 116357. [Google Scholar] [CrossRef] [PubMed]
- Yousfi, S.; Marín, J.; Parra, L.; Lloret, J.; Mauri, P.V. A Rhizogenic Biostimulant Effect on Soil Fertility and Roots Growth of Turfgrass. Agronomy 2021, 11, 573. [Google Scholar] [CrossRef]
- Chen, Q.; Li, Z.; Qu, Z.; Zhou, H.; Qi, Y.; Liu, Z.; Zhang, M. Maize Yield and Root Morphological Characteristics Affected by Controlled-Release Diammonium Phosphate and Paecilomyces Variotii Extracts. Field Crops Res. 2020, 255, 107862. [Google Scholar] [CrossRef]
- Ji, R.; Dong, G.; Shi, W.; Min, J. Effects of Liquid Organic Fertilizers on Plant Growth and Rhizosphere Soil Characteristics of Chrysanthemum. Sustainability 2017, 9, 841. [Google Scholar] [CrossRef]
- Wu, S.C.; Cao, Z.H.; Li, Z.G.; Cheung, K.C.; Wong, M.H. Effects of Biofertilizer Containing N-Fixer, P and K Solubilizers and AM Fungi on Maize Growth: A Greenhouse Trial. Geoderma 2005, 125, 155–166. [Google Scholar] [CrossRef]
- Kim, H.-J.; Ku, K.-M.; Choi, S.; Cardarelli, M. Vegetal-Derived Biostimulant Enhances Adventitious Rooting in Cuttings of Basil, Tomato, and Chrysanthemum via Brassinosteroid-Mediated Processes. Agronomy 2019, 9, 74. [Google Scholar] [CrossRef]
- Monda, H.; Cozzolino, V.; Vinci, G.; Drosos, M.; Savy, D.; Piccolo, A. Molecular Composition of the Humeome Extracted from Different Green Composts and Their Biostimulation on Early Growth of Maize. Plant Soil 2018, 429, 407–424. [Google Scholar] [CrossRef]
- Niewiadomska, A.; Sulewska, H.; Wolna-Maruwka, A.; Ratajczak, K.; Waraczewska, Z.; Budka, A. The Influence of Bio-Stimulants and Foliar Fertilizers on Yield, Plant Features, and the Level of Soil Biochemical Activity in White Lupine (Lupinus albus L.) Cultivation. Agronomy 2020, 10, 150. [Google Scholar] [CrossRef]
- Yildiztekin, M.; Tuna, A.L.; Kaya, C. Physiological Effects of the Brown Seaweed (Ascophyllum nodosum) and Humic Substances on Plant Growth, Enzyme Activities of Certain Pepper Plants Grown under Salt Stress. Biol. Futur. 2018, 69, 325–335. [Google Scholar] [CrossRef]
- Francesca, S.; Arena, C.; Hay Mele, B.; Schettini, C.; Ambrosino, P.; Barone, A.; Rigano, M.M. The Use of a Plant-Based Biostimulant Improves Plant Performances and Fruit Quality in Tomato Plants Grown at Elevated Temperatures. Agronomy 2020, 10, 363. [Google Scholar] [CrossRef]
- Xu, L.; Geelen, D. Developing Biostimulants from Agro-Food and Industrial By-Products. Front. Plant Sci. 2018, 9, 1567. [Google Scholar] [CrossRef] [PubMed]
- Sible, C.N.; Seebauer, J.R.; Below, F.E. Plant Biostimulants: A Categorical Review, Their Implications for Row Crop Production, and Relation to Soil Health Indicators. Agronomy 2021, 11, 1297. [Google Scholar] [CrossRef]
- Canellas, N.O.A.; Olivares, F.L.; Canellas, L.P. Metabolite Fingerprints of Maize and Sugarcane Seedlings: Searching for Markers after Inoculation with Plant Growth-Promoting Bacteria in Humic Acids. Chem. Biol. Technol. Agric. 2019, 6, 14. [Google Scholar] [CrossRef]
- Barone, V.; Puglisi, I.; Fragalà, F.; Stevanato, P.; Baglieri, A. Effect of Living Cells of Microalgae or Their Extracts on Soil Enzyme Activities. Arch. Agron. Soil Sci. 2019, 65, 712–726. [Google Scholar] [CrossRef]
- Paz-Ferreiro, J.; Gascó, G.; Gutiérrez, B.; Méndez, A. Soil Biochemical Activities and the Geometric Mean of Enzyme Activities after Application of Sewage Sludge and Sewage Sludge Biochar to Soil. Biol. Fertil. Soils 2012, 48, 511–517. [Google Scholar] [CrossRef]
- Jezierska-Tys, S.; Wesołowska, S.; Gałązka, A.; Joniec, J.; Bednarz, J.; Cierpiała, R. Biological Activity and Functional Diversity in Soil in Different Cultivation Systems. Int. J. Environ. Sci. Technol. 2020, 17, 4189–4204. [Google Scholar] [CrossRef]
- Mattana, S.; Petrovičová, B.; Landi, L.; Gelsomino, A.; Cortés, P.; Ortiz, O.; Renella, G. Sewage Sludge Processing Determines Its Impact on Soil Microbial Community Structure and Function. Appl. Soil Ecol. 2014, 75, 150–161. [Google Scholar] [CrossRef]
- Wolińska, A.; Włodarczyk, K.; Kuźniar, A.; Marzec-Grządziel, A.; Grządziel, J.; Gałązka, A.; Uzarowicz, Ł. Soil Microbial Community Profiling and Bacterial Metabolic Activity of Technosols as an Effect of Soil Properties Following Land Reclamation: A Case Study from the Abandoned Iron Sulphide and Uranium Mine in Rudki (South-Central Poland). Agronomy 2020, 10, 1795. [Google Scholar] [CrossRef]
- Ma, Y.; Freitas, H.; Dias, M.C. Strategies and Prospects for Biostimulants to Alleviate Abiotic Stress in Plants. Front. Plant Sci. 2022, 13, 1024243. [Google Scholar] [CrossRef]
- Wang, J.Y.; Jamil, M.; Hossain, M.G.; Chen, G.-T.E.; Berqdar, L.; Ota, T.; Blilou, I.; Asami, T.; Al-Solimani, S.J.; Mousa, M.A.A.; et al. Evaluation of the Biostimulant Activity of Zaxinone Mimics (MiZax) in Crop Plants. Front. Plant Sci. 2022, 13, 874858. [Google Scholar] [CrossRef]
- Mazoyon, C.; Catterou, M.; Alahmad, A.; Mongelard, G.; Guénin, S.; Sarazin, V.; Dubois, F.; Duclercq, J. Sphingomonas Sediminicola Dae20 Is a Highly Promising Beneficial Bacteria for Crop Biostimulation Due to Its Positive Effects on Plant Growth and Development. Microorganisms 2023, 11, 2061. [Google Scholar] [CrossRef]
- Hartmann, M.; Frey, B.; Mayer, J.; Mäder, P.; Widmer, F. Distinct Soil Microbial Diversity under Long-Term Organic and Conventional Farming. ISME J. 2015, 9, 1177–1194. [Google Scholar] [CrossRef] [PubMed]
- Mattner, S.W.; Villalta, O.N.; Wite, D.; Porter, I.J.; Arioli, T. In Vitro Suppression of Sclerotinia Minor by a Seaweed Extract from Durvillaea Potatorum and Ascophyllum Nodosum. Australas. Plant Dis. Notes 2014, 9, 137. [Google Scholar] [CrossRef]
- Wang, S.; Wu, Q.; He, X. Exogenous Easily Extractable Glomalin-Related Soil Protein Promotes Soil Aggregation, Relevant Soil Enzyme Activities and Plant Growth in Trifoliate Orange. Plant Soil Environ. 2015, 61, 66–71. [Google Scholar] [CrossRef]
- Sher, Y.; Baker, N.R.; Herman, D.; Fossum, C.; Hale, L.; Zhang, X.; Nuccio, E.; Saha, M.; Zhou, J.; Pett-Ridge, J.; et al. Microbial Extracellular Polysaccharide Production and Aggregate Stability Controlled by Switchgrass (Panicum virgatum) Root Biomass and Soil Water Potential. Soil Biol. Biochem. 2020, 143, 107742. [Google Scholar] [CrossRef]
- Sae-Tun, O.; Bodner, G.; Rosinger, C.; Zechmeister-Boltenstern, S.; Mentler, A.; Keiblinger, K. Fungal Biomass and Microbial Necromass Facilitate Soil Carbon Sequestration and Aggregate Stability under Different Soil Tillage Intensities. Appl. Soil Ecol. 2022, 179, 104599. [Google Scholar] [CrossRef]
- Rutgers, M.; Wouterse, M.; Drost, S.; Breure, A.; Mulder, C.; Stone, D.; Creamer, R.; Winding, A.; Bloem, J. Monitoring Soil Bacteria with Community-Level Physiological Profiles Using BiologTM ECO-Plates in the Netherlands and Europe. Appl. Soil Ecol. 2016, 97, 23–35. [Google Scholar] [CrossRef]
- Chen, S.-K.; Edwards, C.A.; Subler, S. The Influence of Two Agricultural Biostimulants on Nitrogen Transformations, Microbial Activity, and Plant Growth in Soil Microcosms. Soil Biol. Biochem. 2003, 35, 9–19. [Google Scholar] [CrossRef]
- Jezierska-Tys, S.; Joniec, J.; Mocek-Płóciniak, A.; Gałązka, A.; Bednarz, J.; Furtak, K. Microbial Activity and Community Level Physiological Profiles (CLPP) of Soil under the Cultivation of Spring Rape with the Roundup 360 SL Herbicide. J. Environ. Health Sci. Eng. 2021, 19, 2013–2026. [Google Scholar] [CrossRef]
- Houlden, A.; Timms-Wilson, T.M.; Day, M.J.; Bailey, M.J. Influence of Plant Developmental Stage on Microbial Community Structure and Activity in the Rhizosphere of Three Field Crops. FEMS Microbiol. Ecol. 2008, 65, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Zielewicz, W.; Swędrzyński, A.; Dobrzyński, J.; Swędrzyńska, D.; Kulkova, I.; Wierzchowski, P.S.; Wróbel, B. Effect of Forage Plant Mixture and Biostimulants Application on the Yield, Changes of Botanical Composition, and Microbiological Soil Activity. Agronomy 2021, 11, 1786. [Google Scholar] [CrossRef]
- Chen, Y.; Wang, L.; Dai, F.; Tao, M.; Li, X.; Tan, Z. Biostimulants Application for Bacterial Metabolic Activity Promotion and Sodium Dodecyl Sulfate Degradation under Copper Stress. Chemosphere 2019, 226, 736–743. [Google Scholar] [CrossRef] [PubMed]
- Almendras, K.; Leiva, D.; Carú, M.; Orlando, J. Carbon Consumption Patterns of Microbial Communities Associated with Peltigera Lichens from a Chilean Temperate Forest. Molecules 2018, 23, 2746. [Google Scholar] [CrossRef] [PubMed]
- Gryta, A.; Frąc, M.; Oszust, K. Genetic and Metabolic Diversity of Soil Microbiome in Response to Exogenous Organic Matter Amendments. Agronomy 2020, 10, 546. [Google Scholar] [CrossRef]
- Visconti, D.; Ventorino, V.; Fagnano, M.; Woo, S.L.; Pepe, O.; Adamo, P.; Caporale, A.G.; Carrino, L.; Fiorentino, N. Compost and Microbial Biostimulant Applications Improve Plant Growth and Soil Biological Fertility of a Grass-Based Phytostabilization System. Environ. Geochem. Health 2023, 45, 787–807. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Zheng, Y.; Hu, J.; Du, N.; Chen, F. Functional Diversity of the Microbial Community in Healthy Subjects and Periodontitis Patients Based on Sole Carbon Source Utilization. PLoS ONE 2014, 9, e91977. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.-K.; Subler, S.; Edwards, C.A. Effects of Agricultural Biostimulants on Soil Microbial Activity and Nitrogen Dynamics. Appl. Soil Ecol. 2002, 19, 249–259. [Google Scholar] [CrossRef]
- Li, J.; Van Gerrewey, T.; Geelen, D. A Meta-Analysis of Biostimulant Yield Effectiveness in Field Trials. Front. Plant Sci. 2022, 13, 836702. [Google Scholar] [CrossRef]
- Rouphael, Y.; Colla, G. Synergistic Biostimulatory Action: Designing the Next Generation of Plant Biostimulants for Sustainable Agriculture. Front. Plant Sci. 2018, 9, 1655. [Google Scholar] [CrossRef]
- Singh, V.K.; Singh, R.; Kumar, A.; Bhadouria, R. Chapter 13—Harnessing the Potential of Biostimulants and Biocontrol Agents for Sustainable Management of Agricultural Productivity. In Food Security and Plant Disease Management; Kumar, A., Droby, S., Eds.; Woodhead Publishing: Sawston, UK, 2021; pp. 257–277. ISBN 978-0-12-821843-3. [Google Scholar]
- Morales, S.E.; Holben, W.E. Linking Bacterial Identities and Ecosystem Processes: Can “omic” Analyses Be More than the Sum of Their Parts? FEMS Microbiol. Ecol. 2011, 75, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Aguiar-Pulido, V.; Huang, W.; Suarez-Ulloa, V.; Cickovski, T.; Mathee, K.; Narasimhan, G. Metagenomics, Metatranscriptomics, and Metabolomics Approaches for Microbiome Analysis: Supplementary Issue: Bioinformatics Methods and Applications for Big Metagenomics Data. Evol. Bioinform. Online 2016, 12, EBO.S36436. [Google Scholar] [CrossRef] [PubMed]
- Alahmad, A.; Harir, M.; Fochesato, S.; Tulumello, J.; Walker, A.; Barakat, M.; Ndour, P.M.S.; Schmitt-Kopplin, P.; Cournac, L.; Laplaze, L.; et al. Unraveling the Interplay between Root Exudates, Microbiota, and Rhizosheath Formation in Pearl Millet. Microbiome 2024, 12, 1. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Enzymes | Abbreviations | E.C. Numbers | Substrates (Buffer pH and Concentration) |
|---|---|---|---|
| C cycle enzymatic activities | |||
| α-Glucosidase | aGLU | 3.2.1.20 | p-NP-β-D-glucopyranoside (pH 6; 25 mM) |
| β-Glucosidase | bGLU | 3.2.1.21 | p-NP-β-D-glucopyranoside (pH 6; 50 mM) |
| β-Galactosidase | bGAL | 3.2.1.23 | p-NP-β-D-galactopyranoside (pH 7; 0.02 M) |
| Cellulase | CEL | 3.2.1.4 | p-NP-β-D-cellobioside (pH 6; 10 mM) |
| Laccase | LAC | 1.10.3.2 | 2,2′-azino-bis-(3 ethylbenzothiazoline-6-sulfonate) (pH 5; 0.5 mM) |
| N cycle enzymatic activities | |||
| N-acetylglucosaminidase | NAG | 3.2.1.30 | p-NP-N-acetylglucosaminide (pH 6; 10 mM) |
| Urease | URE | 3.5.1.5 | Urea (pH 7; 0.05 mM) |
| P cycle enzymatic activities | |||
| Phosphatase | PHOS | 3.1.4.1 | p-NP-phosphate (pH 7.5; 50 mM) |
| Acid phosphatase | ACP | 3.1.3.2 | p-NP-phosphate (pH 5; 50 mM) |
| Alkaline phosphatase | AKP | 3.1.3.1 | p-NP-phosphate (pH 11; 50 mM) |
| S cycle enzymatic activities | |||
| Arylsulfatase | ARYLS | 3.1.6.1 | p-NP-sulfate (pH 7; 25 mM) |
| Global enzymatic activities | |||
| Dehydrogenase | DEH | 1.1.1.1 | 2,3,5 triphenyltetrazolium chloride (pH 7; 0.12 M) |
| Treatment | ||||
|---|---|---|---|---|
| Enzymes | Unit | SP | SPK | SPN |
| aGLU | nmol PNP min−1 g−1 dry soil | 4.09 ± 0.14 ab | 4.06 ± 0.16 b | 4.3 ± 0.12 a |
| bGLU | nmol PNP min−1 g−1 dry soil | 15.12 ± 0.47 b | 16.48 ± 0.4 a | 16.23 ± 0.17 a |
| bGAL | nmol PNP min−1 g−1 dry soil | 2.27 ± 0.17 b | 2.43 ± 0.03 ab | 2.52 ± 0.05 a |
| CEL | nmol PNP min−1 g−1 dry soil | 6.55 ± 0.96 a | 10.65 ± 5.51 a | 9.68 ± 0.88 a |
| LAC | nmol ATBS ox min−1 g−1 dry soil | 26.68 ± 4.42 b | 33.19 ± 1.37 a | 33.66 ± 4.99 a |
| PHOS | nmol PNP min−1 g−1 dry soil | 50.41 ± 2.52 b | 54.02 ± 2.07 a | 53.56 ± 1.35 ab |
| ACP | nmol PNP min−1 g−1 dry soil | 78.33 ± 1.79 b | 81.2 ± 1.53 a | 83.13 ± 1.36 a |
| AKP | nmol PNP min−1 g−1 dry soil | 104.59 ± 3.42 b | 109.83 ± 0.75 a | 108.04 ± 3.9 ab |
| NAG | nmol PNP min−1 g−1 dry soil | 4.12 ± 0.25 a | 4.09 ± 0.13 a | 4.35 ± 0.26 a |
| URE | nmol NH4Cl min−1 g−1 dry soil | 4.9 ± 0.14 b | 5.27 ± 0.18 a | 5.14 ± 0.24 ab |
| ARYLS | nmol PNP min−1 g−1 dry soil | 5.62 ± 0.02 b | 5.83 ± 0.04 a | 5.7 ± 0.18 ab |
| DEH | INTF nmol min−1 g−1 dry soil | 0.4 ± 0.02 a | 0.4 ± 0.04 a | 0.37 ± 0.02 a |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Alahmad, A.; Edelman, L.; Bouteiller, M.; Castel, L.; Riah-Anglet, W.; Bennegadi-Laurent, N.; Thioye, B.; Bernardon-Mery, A.; Laval, K.; Trinsoutrot-Gattin, I. Unveiling the Impact of Soil Prebiotics on Rhizospheric Microbial Functionality in Zea mays L. Agriculture 2024, 14, 1115. https://doi.org/10.3390/agriculture14071115
Alahmad A, Edelman L, Bouteiller M, Castel L, Riah-Anglet W, Bennegadi-Laurent N, Thioye B, Bernardon-Mery A, Laval K, Trinsoutrot-Gattin I. Unveiling the Impact of Soil Prebiotics on Rhizospheric Microbial Functionality in Zea mays L. Agriculture. 2024; 14(7):1115. https://doi.org/10.3390/agriculture14071115
Chicago/Turabian StyleAlahmad, Abdelrahman, Lucas Edelman, Mathilde Bouteiller, Lisa Castel, Wassila Riah-Anglet, Nadia Bennegadi-Laurent, Babacar Thioye, Aude Bernardon-Mery, Karine Laval, and Isabelle Trinsoutrot-Gattin. 2024. "Unveiling the Impact of Soil Prebiotics on Rhizospheric Microbial Functionality in Zea mays L." Agriculture 14, no. 7: 1115. https://doi.org/10.3390/agriculture14071115
APA StyleAlahmad, A., Edelman, L., Bouteiller, M., Castel, L., Riah-Anglet, W., Bennegadi-Laurent, N., Thioye, B., Bernardon-Mery, A., Laval, K., & Trinsoutrot-Gattin, I. (2024). Unveiling the Impact of Soil Prebiotics on Rhizospheric Microbial Functionality in Zea mays L. Agriculture, 14(7), 1115. https://doi.org/10.3390/agriculture14071115