
Unveiling Drought-Tolerant Corn Hybrids for Early-Season Drought Resilience Using Morpho-Physiological Traits

 , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
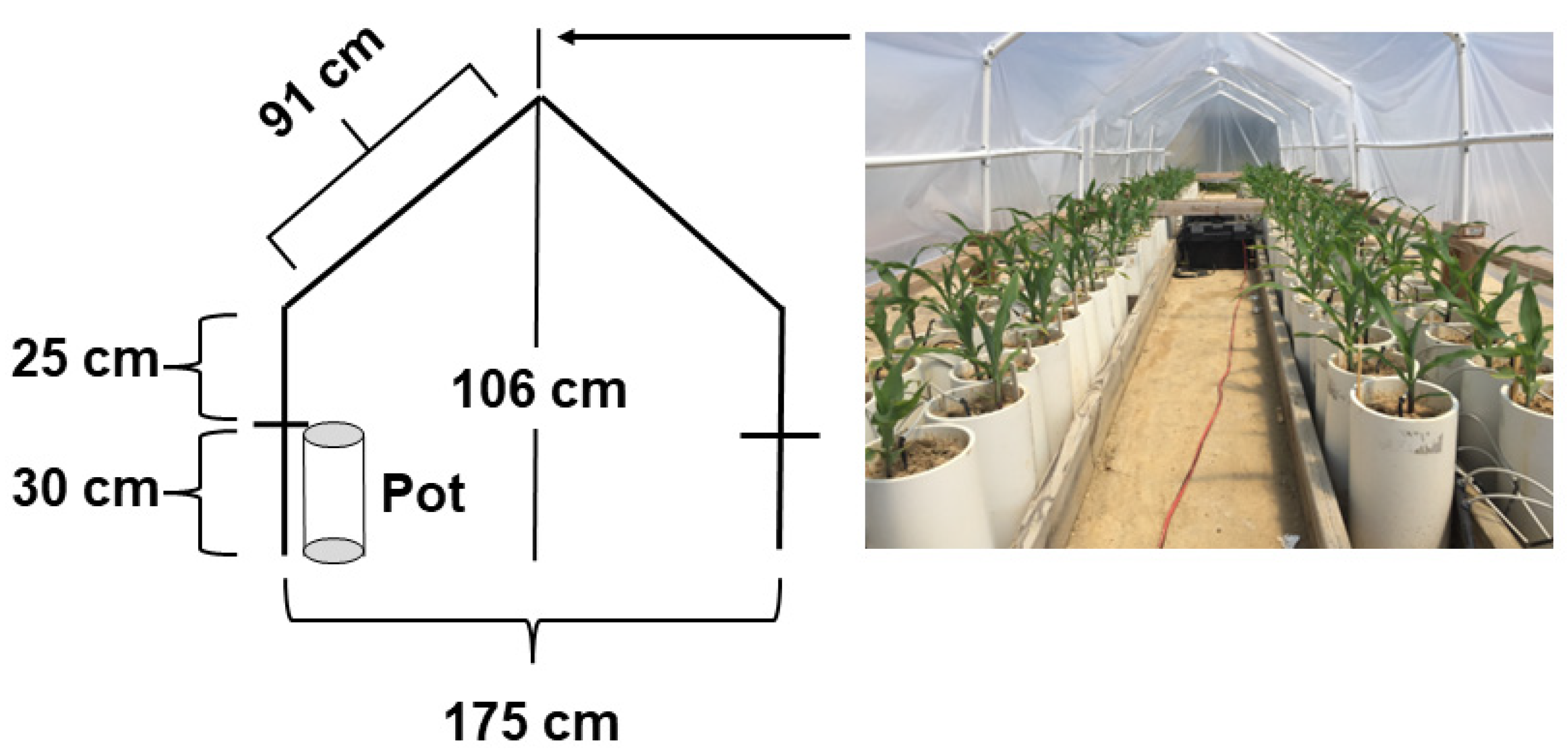
2.1. Experimental Details
2.2. Drought Treatment
2.3. Measurements
2.3.1. Physiological Parameters
2.3.2. Shoot Growth Measurements
2.3.3. Root Image Acquisition and Analysis
2.4. Data Analysis
2.4.1. Principal Component Analysis (PCA)
2.4.2. Drought Stress Response Index (DSRI)
2.4.3. Parameter Relationship
3. Results
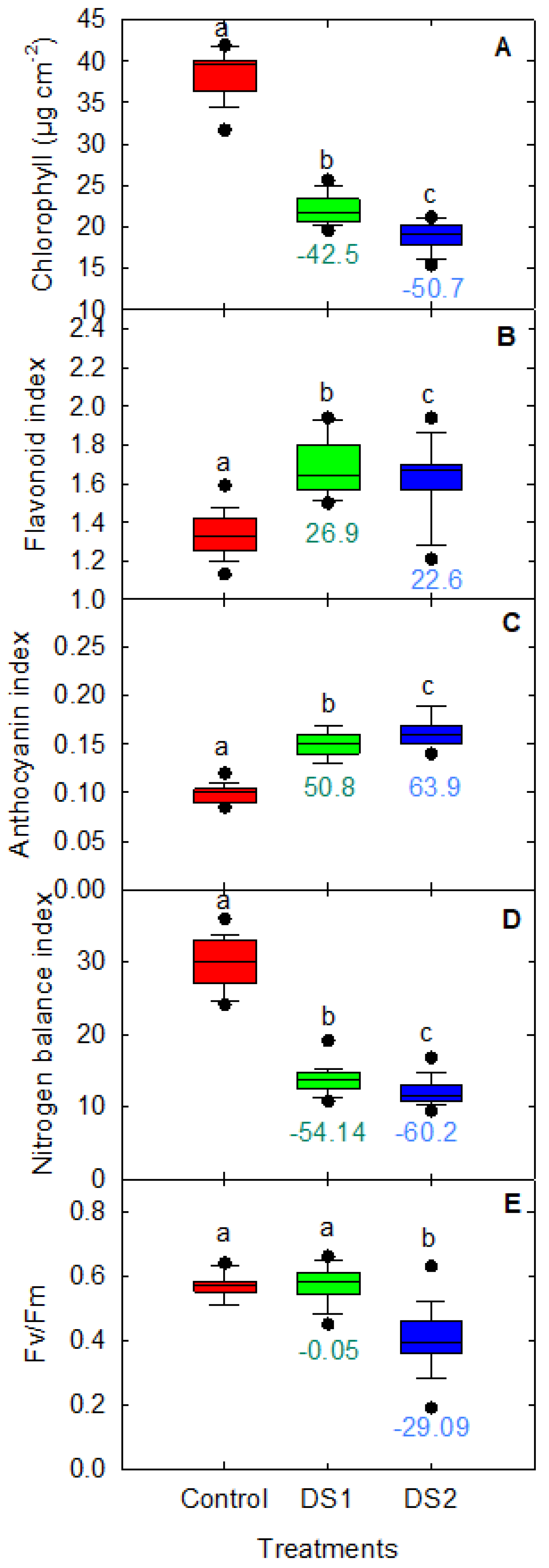
3.1. Physiological Traits in Response to Drought Stress
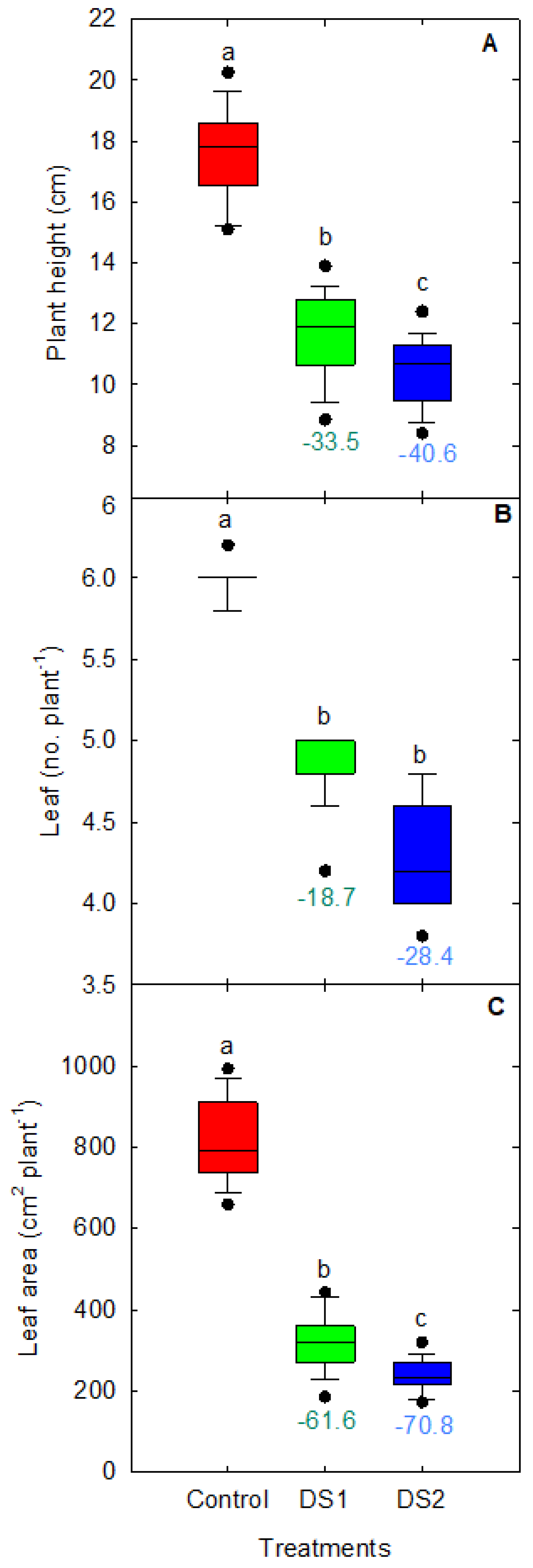
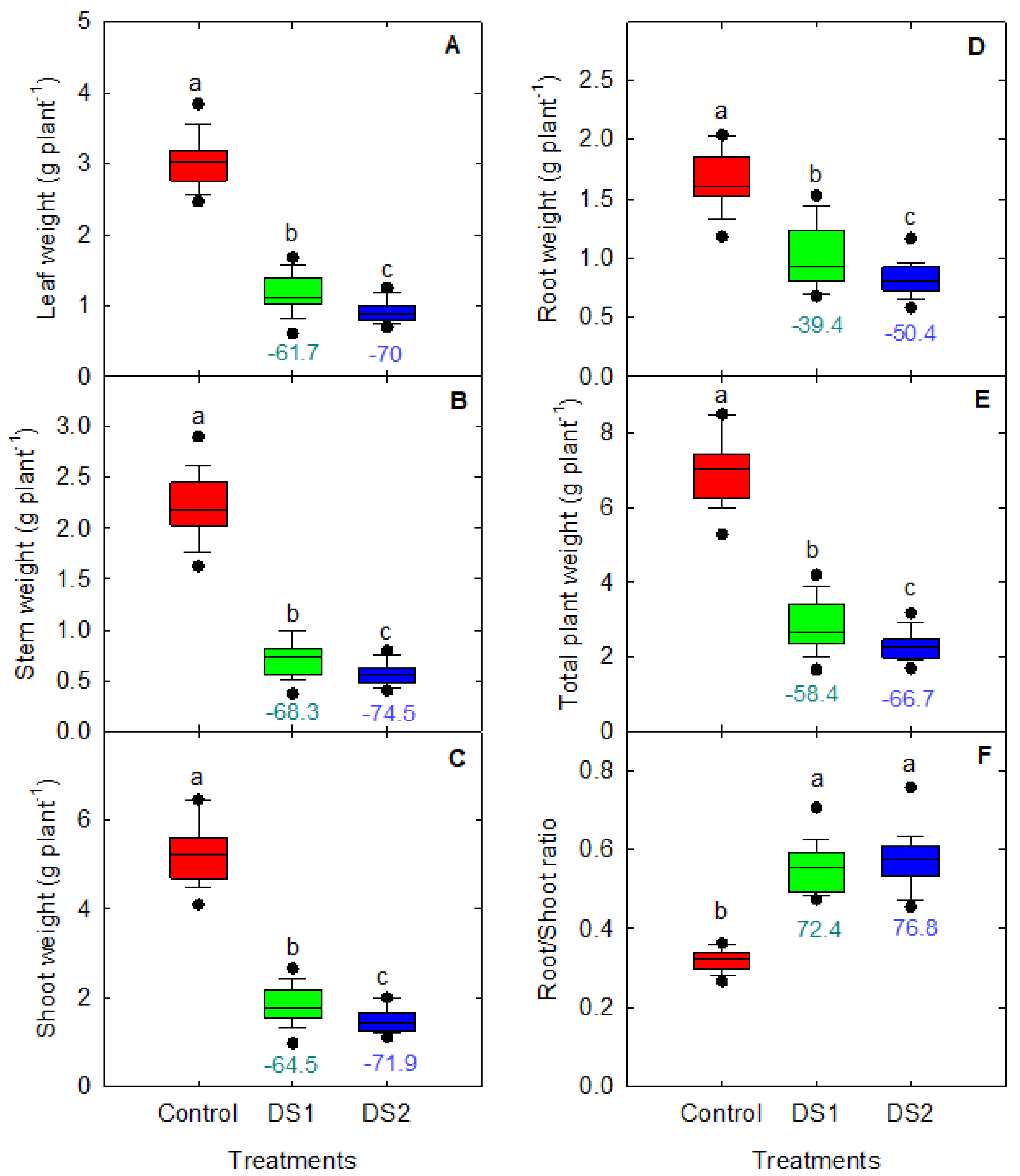
3.2. Shoot Growth and Biomass in Response to Drought Stress
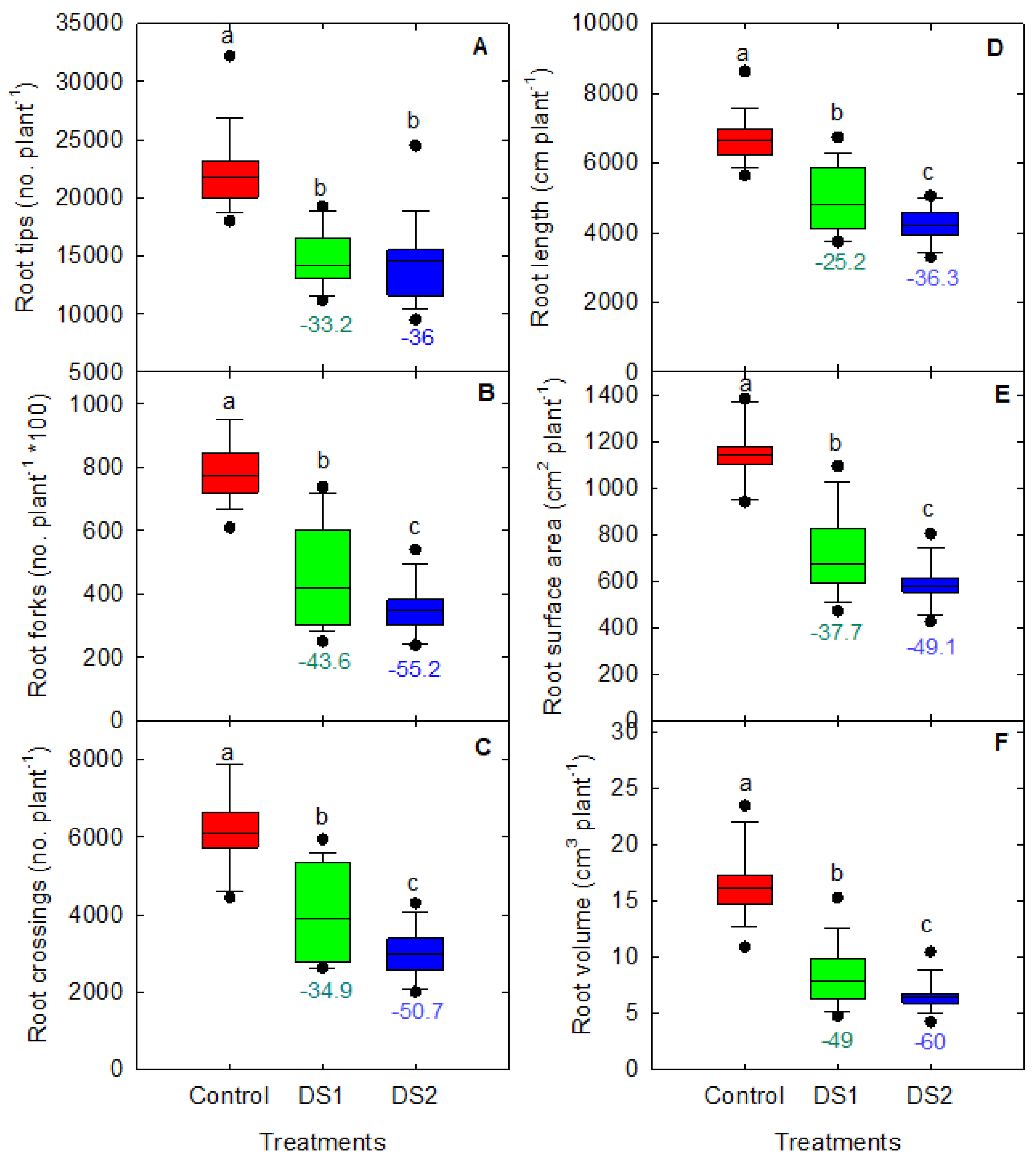
3.3. Root Traits in Response to Drought Stress
3.4. Hybrid Performance under Drought Stress
3.5. Cumulative Drought Stress Response Index
3.6. Classification of Corn Hybrids
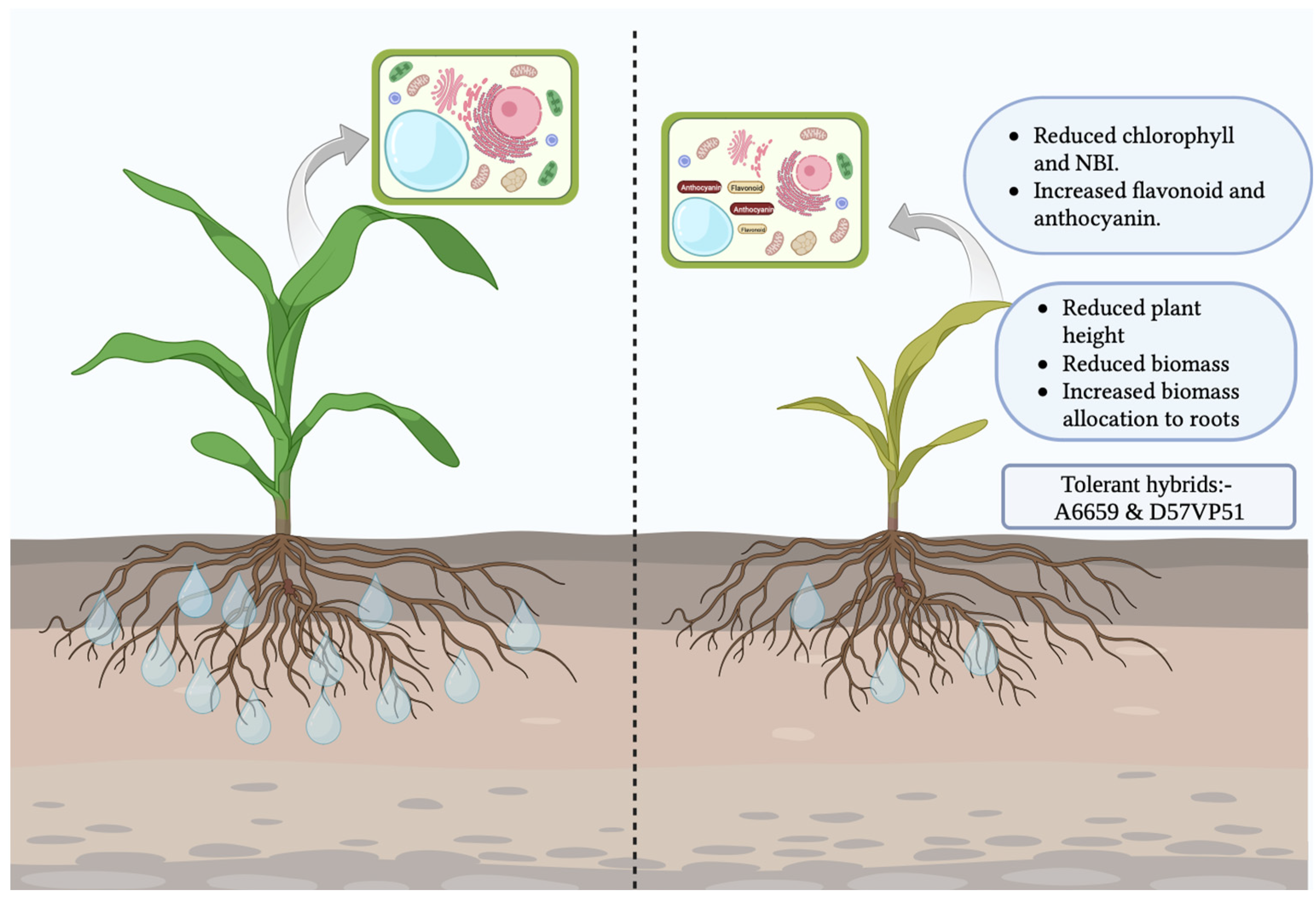
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- NASA. Global Climate Change: The Evidence for Rapid Climate Change is Compelling. 2023. Available online: https://climate.nasa.gov/evidence/ (accessed on 20 September 2023).
- NOAA. Greenhouse Gases Continued to Increase Rapidly in 2022. 2022. Available online: https://www.noaa.gov/news-release/greenhouse-gases-continued-to-increase-rapidly-in-2022 (accessed on 20 September 2023).
- NOAA-NIDIS. NOAA-National Integrated Drought Information System. Historical Drought. 2023. Available online: https://www.drought.gov/what-is-drought/historical-drought#overview (accessed on 27 November 2023).
- NOAA-NIDIS. NOAA-National Integrated Drought Information System. National Current Conditions, 20–26 September 2023. Available online: https://www.drought.gov/current-conditions (accessed on 24 November 2023).
- NDMC. National Drought Mitigation Center, U.S. Drought Monitor. 2023. Available online: https://droughtmonitor.unl.edu (accessed on 24 November 2023).
- USDA-NASS. USDA-National Agricultural Statistics Service. Corn Production. 2023. Available online: https://www.nass.usda.gov/Charts_and_Maps/Field_Crops/cornprod.php (accessed on 5 January 2024).
- USDA-FAS. USDA-Foreign Agricultural Service. U.S. Corn Exports in 2022. 2022. Available online: https://www.fas.usda.gov/data/commodities/corn (accessed on 5 January 2024).
- USDA. Agriculture in Drought. 2023. Available online: https://www.usda.gov/sites/default/files/documents/AgInDrought.pdf (accessed on 5 January 2024).
- Kukal, M.S.; Irmak, S. Irrigation-limited yield gaps: Trends and variability in the United States post-1950. Environ. Res. Commun. 2019, 1, 061005. [Google Scholar] [CrossRef]
- Vennam, R.R.; Poudel, S.; Ramamoorthy, P.; Samiappan, S.; Reddy, K.R.; Bheemanahalli, R. Impact of soil moisture stress during the silk emergence and grain-filling in maize. Physiol. Plant. 2023, 175, e14029. [Google Scholar] [CrossRef]
- Vennam, R.R.; Ramamoorthy, P.; Poudel, S.; Reddy, K.R.; Henry, W.B.; Bheemanahalli, R. Developing functional relationships between soil moisture content and corn early-season physiology, growth, and development. Plants 2023, 12, 2471. [Google Scholar] [CrossRef]
- Anderson, B. Early Spring Drought Review for US Grain and Oilseed Crops. 2022. Available online: https://www.dtnpf.com/agriculture/web/ag/news/article/2022/03/23/early-spring-drought-review (accessed on 25 November 2023).
- Harb, A.; Krishnan, A.; Ambavaram, M.M.; Pereira, A. Molecular and physiological analysis of drought stress in Arabidopsis reveals early responses leading to acclimation in plant growth. Plant Physiol. 2010, 154, 1254–1271. [Google Scholar] [CrossRef]
- Vukasovic, S.; Alahmad, S.; Christopher, J.; Snowdon, R.J.; Stahl, A.; Hickey, L.T. Dissecting the genetics of early vigor to design drought-adapted wheat. Front. Plant Sci. 2022, 12, 754439. [Google Scholar] [CrossRef]
- Condon, A.G.; Richards, R.A.; Rebetzke, G.J.; Farquhar, G.D. Breeding for high water-use efficiency. J. Exp. Bot. 2004, 55, 2447–2460. [Google Scholar] [CrossRef] [PubMed]
- Reddy, K.R.; Hodges, H.F.; Mckinion, J.M. Crop modelling and applications: A cotton example. Adv. Agron. 1997, 59, 225–290. [Google Scholar] [CrossRef]
- Lawlor, D.W.; Tezara, W. Causes of decreased photosynthetic rate and metabolic capacity in water-deficient leaf cells: A critical evaluation of mechanisms and integration of processes. Ann. Bot. 2009, 103, 561–579. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Diaz, M.F.; Kramer, P.J. Behavior of corn and Sorghum under water stress and during recovery. Plant Physiol. 1971, 48, 613–616. [Google Scholar] [CrossRef] [PubMed]
- Wijewardana, C.; Henry, W.B.; Reddy, K.R. Evaluation of drought tolerant maize germplasm to induced drought stress. J. Miss. Acad. Sci. 2017, 62, 316–329. [Google Scholar]
- Mc Fadden, J.; Smith, D.; Wechsler, S.; Wallander, S. Development, Adoption, and Management of Drought-Tolerant Corn in the United States. A Report Summary form the Economic Research Service, USDA. 2019. Available online: https://www.ers.usda.gov/webdocs/publications/91103/eib-204.pdf?v=417 (accessed on 15 January 2024).
- Lone, A.A.; Jumaa, S.H.; Wijewardana, C.; Taduri, S.; Redona, E.D.; Reddy, K.R. Drought stress tolerance screening of elite American breeding rice genotypes using low-cost pre-fabricated mini-hoop modules. Agronomy 2019, 9, 199. [Google Scholar] [CrossRef]
- Hewitt, E.J. Sand and water culture methods used in the study of plant nutrition. Soil Sci. Soc. Am. J. 1953, 17, 301. [Google Scholar] [CrossRef]
- Popat, R.; Banakara, K. Package ‘doebioresearch’: Analysis of Design of Experiments for Biological Research. CRAN Repository, R Studio. 2020. Available online: https://cran.r-project.org/web/packages/doebioresearch/index.html (accessed on 27 November 2023).
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis; Springer: New York, NY, USA, 2016; ISBN 978-3-319-24277-4. Available online: https://ggplot2.tidyverse.org (accessed on 25 December 2023).
- Seleiman, M.F.; Al-Suhaibani, N.; Ali, N.; Akmal, M.; Alotaibi, M.; Refay, Y.; Dindaroglu, T.; Abdul-Wajid, H.H.; Battaglia, M.L. Drought stress impacts on plants and different approaches to alleviate its adverse effects. Plants 2021, 10, 259. [Google Scholar] [CrossRef] [PubMed]
- Wolny, E.; Betekhtin, A.; Rojek, M.; Braszewska-Zalewska, A.; Lusinska, J.; Hasterok, R. Germination and the early stages of seedling development in Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 2916. [Google Scholar] [CrossRef] [PubMed]
- Verslues, P.E.; Agarwal, M.; Katiyar-Agarwal, S.; Zhu, J.; Zhu, J.K. Methods and concepts in quantifying resistance to drought, salt and freezing, abiotic stresses that affect plant water status. Plant J. 2006, 45, 523–539. [Google Scholar] [CrossRef] [PubMed]
- Mafakheri, A.; Siosemardeh, A.; Bahramnejad, B.; Struik, P.C.; Sohrabi, Y. Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust. J. Crop Sci. 2010, 4, 580–585. Available online: https://edepot.wur.nl/159961 (accessed on 3 January 2024).
- Chen, D.; Wang, S.; Xiong, B.; Cao, B.; Deng, X. Carbon/nitrogen imbalance associated with drought-induced leaf senescence in Sorghum bicolor. PLoS ONE 2015, 10, 0137026. [Google Scholar] [CrossRef] [PubMed]
- Zaefyzadeh, M.; Quliyev, R.A.; Babayeva, S.M.; Abbasov, M.A. The effect of the interaction between genotypes and drought stress on the superoxide dismutase and chlorophyll content in durum wheat landraces. Turk. J. Biol. 2009, 33, 1–7. [Google Scholar] [CrossRef]
- Yang, X.; Lu, M.; Wang, Y.; Wang, Y.; Liu, Z.; Chen, S. Response mechanism of plants to drought stress. Horticulturae 2021, 7, 50. [Google Scholar] [CrossRef]
- Paknejad, F.; Nasri, M.; Moghadam, H.R.T.; Zahedi, H.; Alahmadi, M.J. Effects of drought stress on chlorophyll fluorescence parameters, chlorophyll content and grain yield of wheat cultivars. J. Biol. Sci. 2007, 7, 841–847. [Google Scholar] [CrossRef]
- Osakabe, Y.; Osakabe, K.; Shinozaki, K.; Tran, L.S. Response of plants to water stress. Front. Plant Sci. 2014, 5, 86. [Google Scholar] [CrossRef] [PubMed]
- Li, B.; Fan, R.; Sun, G.; Sun, T.; Fan, Y.; Bai, S.; Guo, S.; Huang, S.; Liu, J.; Zhang, H.; et al. Flavonoids improve drought tolerance of maize seedlings by regulating the homeostasis of reactive oxygen species. Plant Soil 2021, 461, 389–405. [Google Scholar] [CrossRef]
- Mishra, N.; Jiang, C.; Chen, L.; Paul, A.; Chatterjee, A.; Shen, G. Achieving abiotic stress tolerance in plants through antioxidative defense mechanisms. Front. Plant Sci. 2023, 14, 1110622. [Google Scholar] [CrossRef] [PubMed]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Dash, S.S.S.; Lenka, D.; Sahoo, J.P.; Tripathy, S.K.; Samal, K.C.; Lenka, D.; Panda, R.K. Biochemical characterization of maize (Zea mays L.) hybrids under excessive soil moisture stress. Cereal Res. Commun. 2022, 50, 875–884. [Google Scholar] [CrossRef]
- Hossain, M.A.; Hoque, M.A.; Burritt, D.J.; Fujita, M. Proline protects plants against abiotic oxidative stress: Biochemical and molecular mechanisms. In Oxidative Damage to Plants; Academic Press: Cambridge, MA, USA, 2014; pp. 477–522. [Google Scholar]
- Yousaf, M.I.; Riaz, M.W.; Shehzad, A.; Jamil, S.; Shahzad, R.; Kanwal, S.; Ghani, A.; Ali, F.; Abdullah, M.; Ashfaq, M.; et al. Responses of maize hybrids to water stress conditions at different developmental stages: Accumulation of reactive oxygen species, activity of enzymatic antioxidants and degradation in kernel quality traits. PeerJ 2023, 11, e14983. [Google Scholar] [CrossRef]
- Chen, S.; Wu, F.; Li, Y.; Qian, Y.; Pan, X.; Li, F.; Wang, Y.; Wu, Z.; Fu, C.; Lin, H.; et al. NtMYB4 and NtCHS1 are critical factors in the regulation of flavonoid biosynthesis and are involved in salinity responsiveness. Front. Plant Sci. 2019, 10, 178. [Google Scholar] [CrossRef]
- Kovinich, N.; Kayanja, G.; Chanoca, A.; Otegui, M.S.; Grotewold, E. Abiotic stresses induce different localizations of anthocyanins in Arabidopsis. Plant Signal. Behav. 2015, 10, e1027850. [Google Scholar] [CrossRef]
- Maritim, T.K.; Korir, R.K.; Nyabundi, K.W.; Wachira, F.N.; Kamunya, S.M.; Muoki, R.C. Molecular regulation of anthocyanin discoloration under water stress and high solar irradiance in pluckable shoots of purple tea cultivar. Planta 2021, 254, 85. [Google Scholar] [CrossRef]
- Chen, X.; Zhu, Y.; Ding, Y.; Pan, R.; Shen, W.; Yu, X.; Xiong, F. The relationship between characteristics of root morphology and grain filling in wheat under drought stress. PeerJ 2021, 9, e12015. [Google Scholar] [CrossRef]
- Pham, A.T.; Maurer, A.; Pillen, K.; Brien, C.; Dowling, K.; Berger, B.; Eglinton, J.K.; March, T.J. Genome-wide association of barley plant growth under drought stress using a nested association mapping population. BMC Plant Biol. 2019, 19, 134. [Google Scholar] [CrossRef] [PubMed]
- Waheed, R.; Deeba, F.; Zulfiqar, F.; Moosa, A.; Nafees, M.; Altaf, M.A.; Arif, M.; Siddique, K.H.M. Physiology and growth of newly bred Basmati rice lines in response to vegetative-stage drought stress. Front. Plant Sci. 2023, 14, 1172255. [Google Scholar] [CrossRef] [PubMed]
- Patmi, Y.S.; Pitoyo, A.; Solichatun; Sutarno. Effect of drought stress on morphological, anatomical, and physiological characteristics of Cempo Ireng Cultivar Mutant Rice (Oryza sativa L.) strain 51 irradiated by gamma-ray. J. Phys. Conf. Ser. 2020, 1436, 012015. [Google Scholar] [CrossRef]
- Werner, C.; Correia, O.; Beyschlag, W. Two different strategies of Mediterranean macchia plants to avoid photo-inhibitory damage by excessive radiation levels during summer drought. Acta Oecologica 1999, 20, 15–23. [Google Scholar] [CrossRef]
- Xu, K.; Yang, D.; Yang, H.; Li, Z.; Qin, Y.; Shen, Y. Spatio-temporal variation of drought in China during 1961–2012: A climatic perspective. J. Hydrol. 2015, 526, 253–264. [Google Scholar] [CrossRef]
- Sehgal, A.; Sita, K.; Siddique, K.H.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; Rao, B.H.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and heat-stress effects on seed filling in food crops: Impacts on functional biochemistry, seed yields, and nutritional quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Hybrid | Company and Location |
|---|---|
| A6499 | AgriGold, AgReliant Genetics, St. Francisville, IL, USA |
| A6659 | AgriGold, AgReliant Genetics, St. Francisville, IL, USA |
| A6711 | AgriGold, AgReliant Genetics, St. Francisville, IL, USA |
| 1717 | Armor Seed, Jonesboro, AR, USA |
| A7766 | Augusta Seed Corporation, Verona, VA, USA |
| 6640 | Croplan, Saint Paul, MN, USA |
| DKC67-44 | Dekalb Genetics Corporation, DeKalb, IL, USA |
| DKC67-72 | Dekalb Genetics Corporation, DeKalb, IL, USA |
| DKC68-26 | Dekalb Genetics Corporation, DeKalb, IL, USA |
| DG2888 | Delta Grow Seed Co. Inc., England, AR, USA |
| D57VP51 | Dyna-Gro Seed, Inc., Geneseo, IL, USA |
| D57VP75 | Dyna-Gro Seed, Inc., Geneseo, IL, USA |
| MC4319 | MorCorn Hybrids, MFA Incorporated, Columbia, MO, USA |
| CNX157137 | Mycogen Hybrids, MFA Incorporated, Columbia, MO, USA |
| NK1405 | NK Seeds, Syngenta, St. Gabriel, LA, USA |
| P1316 | Pioneer Corporation, Springboro, OH, USA |
| PGY7111 | Progeny AG Products, Wynne, AR, USA |
| PGY7215 | Progeny AG Products, Wynne, AR, USA |
| REV25BHR26 | Terral Seed Inc., Lake Providence, LA, USA |
| Parameters | Drought (D) | Hybrids (H) | D × H |
|---|---|---|---|
| Soil moisture content ! | *** | - | - |
| Plant height | *** | *** | * |
| Leaf number | * | NS | NS |
| Leaf area | *** | ** | NS |
| Leaf dry weight | *** | *** | NS |
| Stem dry weight | *** | *** | NS |
| Shoot dry weight | *** | *** | NS |
| Root dry weight | *** | *** | NS |
| Total dry weight | *** | *** | NS |
| Root/shoot ratio | *** | *** | ** |
| Root length | *** | * | NS |
| Root Surface area | *** | *** | NS |
| Root avg. diameter | *** | *** | NS |
| Length per volume | *** | * | NS |
| Root volume | *** | *** | * |
| Root tips | *** | NS | NS |
| Root forks | *** | *** | NS |
| Root crossings | *** | ** | NS |
| Chlorophyll | *** | *** | * |
| Flavonoid index | *** | *** | ** |
| Anthocyanin index | *** | *** | NS |
| NBI | *** | *** | ** |
| Fv/Fm | ** | * | * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Walne, C.H.; Thenveettil, N.; Ramamoorthy, P.; Bheemanahalli, R.; Reddy, K.N.; Reddy, K.R. Unveiling Drought-Tolerant Corn Hybrids for Early-Season Drought Resilience Using Morpho-Physiological Traits. Agriculture 2024, 14, 425. https://doi.org/10.3390/agriculture14030425
Walne CH, Thenveettil N, Ramamoorthy P, Bheemanahalli R, Reddy KN, Reddy KR. Unveiling Drought-Tolerant Corn Hybrids for Early-Season Drought Resilience Using Morpho-Physiological Traits. Agriculture. 2024; 14(3):425. https://doi.org/10.3390/agriculture14030425
Chicago/Turabian StyleWalne, Charles Hunt, Naflath Thenveettil, Purushothaman Ramamoorthy, Raju Bheemanahalli, Krishna N. Reddy, and Kambham Raja Reddy. 2024. "Unveiling Drought-Tolerant Corn Hybrids for Early-Season Drought Resilience Using Morpho-Physiological Traits" Agriculture 14, no. 3: 425. https://doi.org/10.3390/agriculture14030425
APA StyleWalne, C. H., Thenveettil, N., Ramamoorthy, P., Bheemanahalli, R., Reddy, K. N., & Reddy, K. R. (2024). Unveiling Drought-Tolerant Corn Hybrids for Early-Season Drought Resilience Using Morpho-Physiological Traits. Agriculture, 14(3), 425. https://doi.org/10.3390/agriculture14030425