
Patterns of Copper Bioaccumulation and Translocation in Grapevine Grafts Depending on Rootstocks


Abstract
:1. Introduction
2. Materials and Methods
2.1. Experimental Setting
2.2. Growth Inhibition and Cu Accumulation Assessment
2.3. Statistical Analysis
3. Results
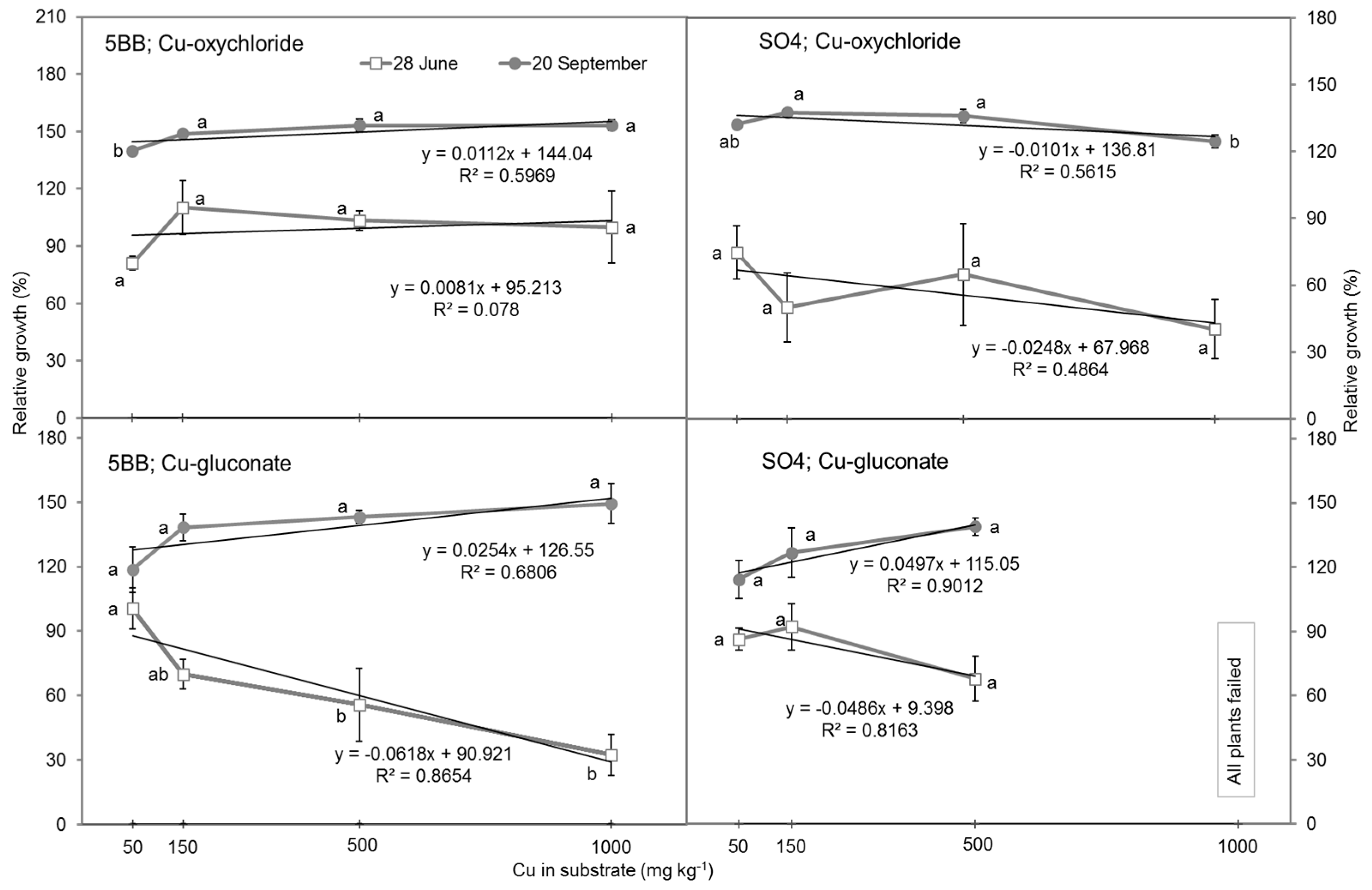
3.1. Shoot Growth Inhibition
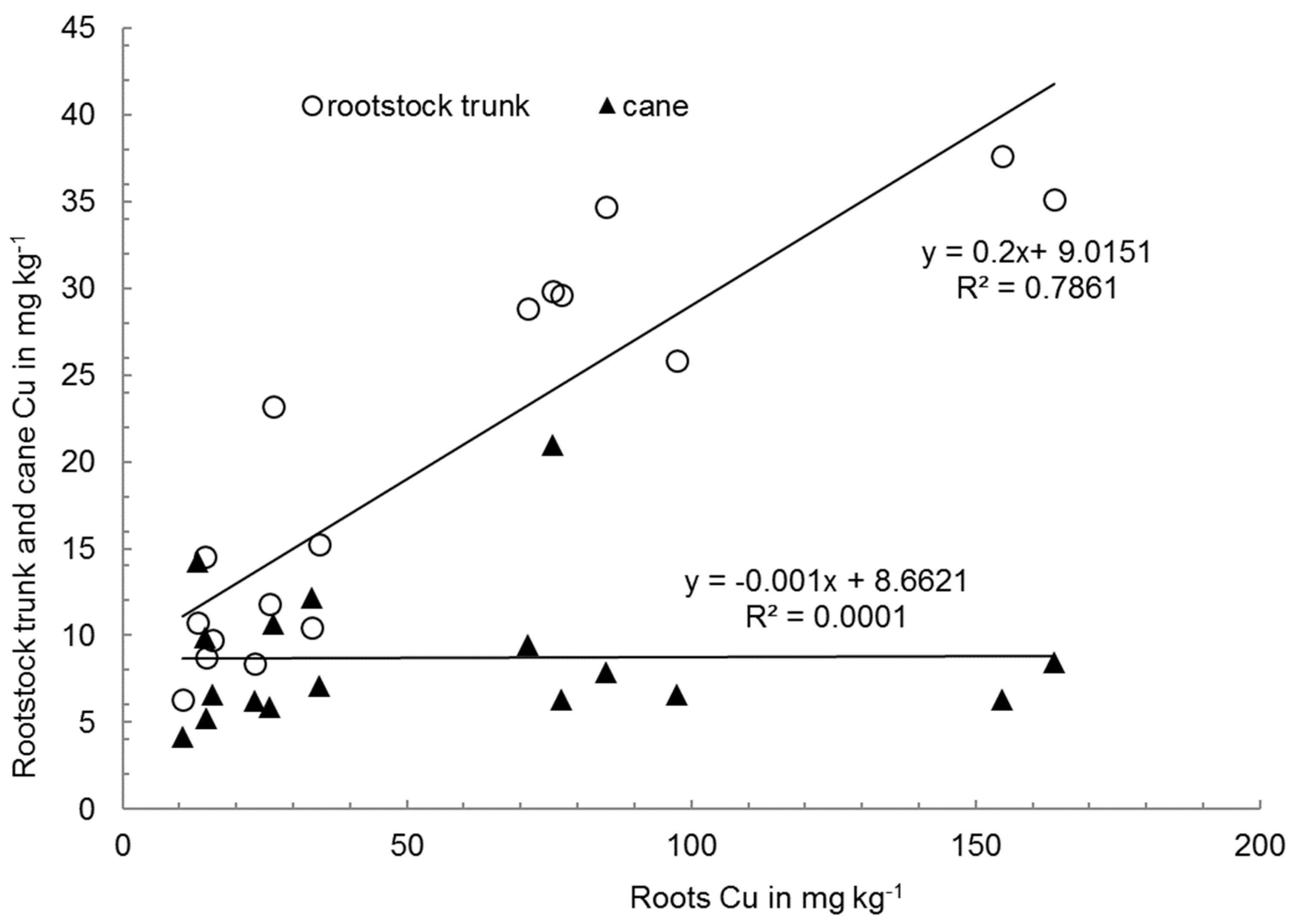
3.2. Cu in Graft Organs
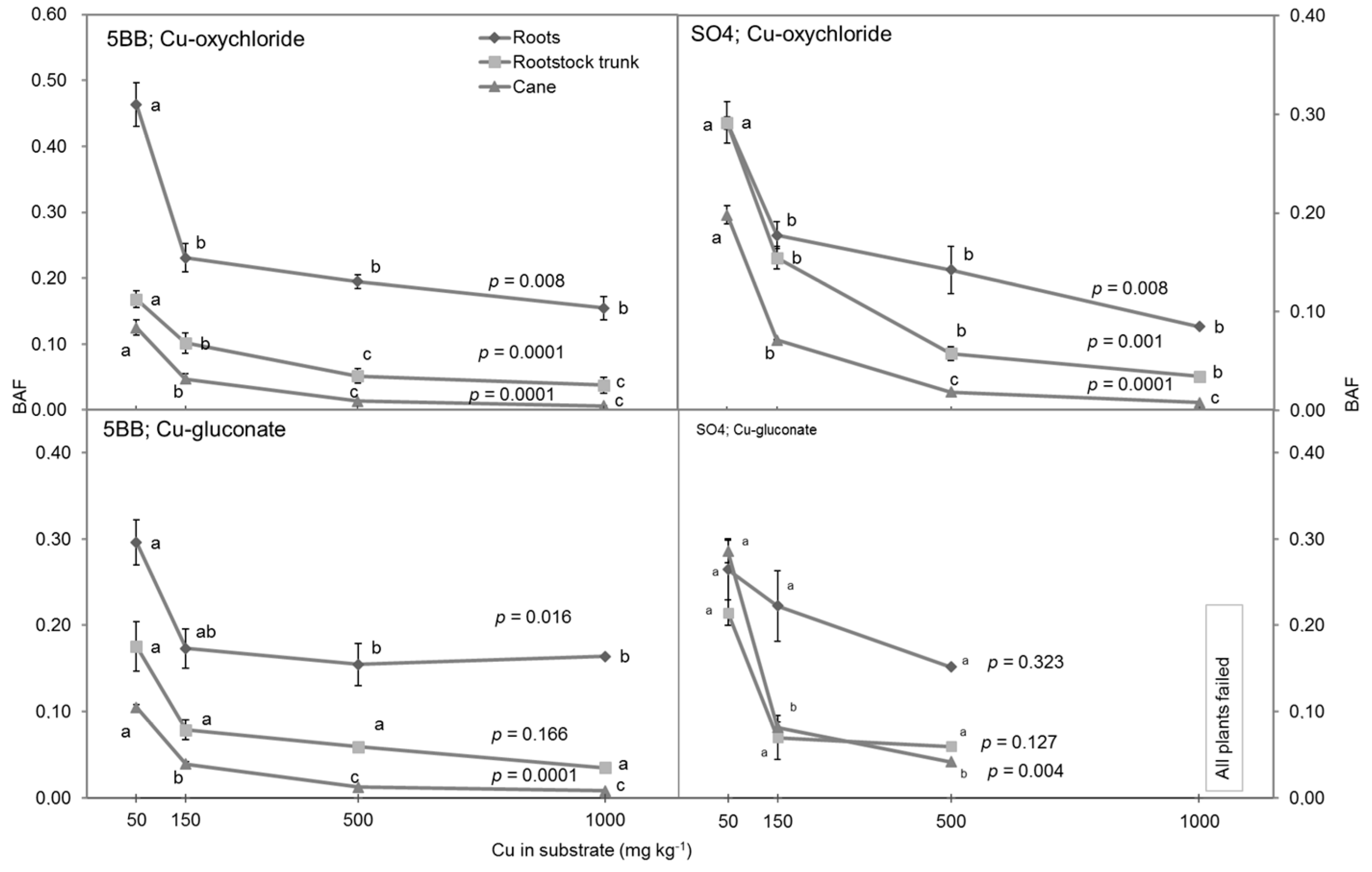
3.3. Bioaccumulation and Translocation of Cu
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brun, L.A.; Maillet, J.; Hinsinger, P.; Pepin, M. Evaluation of Copper Availability to Plants in Copper-Contaminated Vineyard Soils. Environ. Pollut. 2001, 111, 293–302. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Esparza, M.A.; Capri, E.; Pirzadeh, P.; Trevisan, M. Copper Content of Grape and Wine from Italian Farms. Food Addit. Contam. 2006, 23, 274–280. [Google Scholar] [CrossRef] [PubMed]
- Casali, C.A.; Moterle, D.F.; Rheinheimer, D.D.; Brunetto, G.; Corcini, A.L.M.; Kaminski, J.; de Melo, G.W.B. Copper Forms and Desorption in Soils under Grape Vine in the Serra Gaucha of Rio Grande Do Sul. Rev. Bras. Cienc. Solo 2008, 32, 1479–1487. [Google Scholar] [CrossRef]
- Kovačič, R.G.; Lešnik, M.; Vršič, S. An Overview of the Copper Situation and usage in Viticulture. Bulg. J. Agric. Sci. 2013, 19, 50–59. [Google Scholar]
- Chaignon, V.; Sanchez-Neira, I.; Jaillard, B.; Hinsinger, P. Copper Bioavailability and Extractability as Related to Chemical Properties of Contaminated Soils from a Vine-Growing Area. Environ. Pollut. 2003, 123, 229–238. [Google Scholar] [CrossRef]
- Mirlean, N.; Roisenberg, A.; Chies, J.O. Metal Contamination of Vineyard Soils in Wet Subtropics (Southern Brazil). Environ. Pollut. 2007, 149, 10–17. [Google Scholar] [CrossRef]
- Borkow, G.; Gabbay, J. Copper as a Biocidal Tool; Cupron Inc., Publications: Gibton, Israel, 2007; 42p. [Google Scholar] [CrossRef]
- Ware, G.W.; Whitacre, D.M. The Pesticide Book, 6th ed.; MeisterPro Information Resources: Willoughby, OH, USA, 2004; 487p. [Google Scholar]
- Helling, B.; Reineeke, S.; Reineeke, A.J. Effects of the Fungicide Copper Oxychloride on the Growth and Reproduction of Eisena (Oligochaeta). Ecotoxicol. Environ. Saf. 2000, 46, 108–116. [Google Scholar] [CrossRef]
- Schoffer, J.T.; Sauvé, S.; Neaman, A.; Ginocchio, R. Role of Leaf Litter on the Incorporation of Copper-Containing Pesticides into Soils Under Fruit Production: A Review. J. Soil Sci. Plant Nutr. 2020, 20, 990–1000. [Google Scholar] [CrossRef]
- Toselli, M.; Baldi, E.; Marcolini, G.; Malaguti, D.; Quartieri, M.; Sorrenti, G.; Marangoni, B. Response of Potted Grapevines to Increasing Soil Copper Concentration. Aust. J. Grape Wine Res. 2009, 15, 85–92. [Google Scholar] [CrossRef]
- Vršič, S.; Ivanćić, A.; Pulko, B.; Valdhuber, J. Effect of Soil Management Systems on Erosion and Nutrition Loss in Vineyards on Steep Slopes. J. Environ. Biol. 2011, 32, 289–294. [Google Scholar]
- Mengel, K.E.; Kirkbiy, A.; Kosegarten, H.; Appel, T. Principles of Plant Nutrition; Kluwer Academic Publishers: Dordecht, The Netherlands; Boston, MA, USA; London, UK, 2001; pp. 537–549. [Google Scholar]
- Kopittke, P.M.; Menzies, N.W. Effect of Cu Toxicity on Growth of Cowpea (Vigna unguiculata). Plant Soil 2006, 27, 287–296. [Google Scholar] [CrossRef]
- Mourato, M.P.; Martins, L.L.; Campos-Andrada, M.P. Physiological Responses of Lupinus luteus to Different Copper Concentrations. Biol. Plant. 2009, 53, 105–111. [Google Scholar] [CrossRef]
- Liu, W.; Yang, Y.S.; Li, P.; Zhou, Q.; Sun, T. Root Growth Inhibition and Induction of DNA Damage in Soybean (Glycine max) by Chlorobenzenes in Contaminated Soil. Chemosphere 2009, 57, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Juang, K.W.; Lee, Y.I.; Lai, H.Y.; Wang, C.H.; Chen, B.C. Copper Accumulation, Translocation, and Toxic Effects in Grapevine Cuttings. Environ. Sci. Pollut. Res. 2012, 19, 1315–1322. [Google Scholar] [CrossRef] [PubMed]
- McLaughlin, M.J.; Smolders, E.; Merckx, R. Soil Root Interface: Physicochemical Processes. In Soil Chemistry and Ecosystem Health; Huang, P.M., Ed.; Soil Science Society of America, Inc.: Madison, WI, USA, 1998; Volume 52, pp. 233–277. [Google Scholar]
- Romić, M.; Romić, D.; Ondrašek, G. Heavy Metals Accumulation in Topsoil from the Wine Growing Regions Part 2, Relationships between Soil Properties and Extractable Copper Contents. Agric. Conspec. Scentificus 2004, 69, 111–118. [Google Scholar]
- Beni, C.; Rossi, G. Conventional and Organic Farming: Estimation of some Effects on Soil, Copper Accumulation and Wine in a Central Italy Vineyard. Agrochimica 2009, 53, 145–159. [Google Scholar]
- Peralta, J.R.; Gardea-Torresdey, J.L.; Tiemann, K.J.; Gomez, E.; Arteaga, S.; Rascon, E.; Parsons, J.G. Uptake and Effects of five Heavy Metals on Seed Germination and Plant Growth in Alfalfa (Medicago sativa L.). Bull. Environ. Contam. Toxicol. 2001, 66, 727–734. [Google Scholar] [CrossRef]
- Yang, M.J.; Yang, X.E.; Römheld, V. Growth and Nutrient Composition of Elsholtzia splendens Nakai under Copper Toxicity. J. Plant Nutr. 2002, 25, 1359–1375. [Google Scholar] [CrossRef]
- Ali, M.B.; Vajpayee, P.; Tripathi, R.D.; Rai, U.N.; Singh, S.N.; Singh, S.P. Phytoremediation of Lead, Nickel and Copper by Salix acmophylla Boiss: Role of Antioxidant Enzymes and Antioxidant Substances. Bull. Environ. Contam. Toxicol. 2003, 70, 462–469. [Google Scholar] [CrossRef]
- Mackie, K.A.; Marhan, S.; Ditterich, F.; Schmidt, H.P.; Kandeler, E. The Effects of Biochar and Compost Amendments on Copper Immobilization and Soil Microorganisms in a Temperate Vineyard. Agric. Ecosyst. Environ. 2015, 201, 58–69. [Google Scholar] [CrossRef]
- Khellaf, N.; Zerdaoui, M. Growth Response of the Duckweed Lemna gibba L. to Copper and Nickel Phytoaccumulation. Ecotoxicology 2010, 19, 1363–1368. [Google Scholar] [CrossRef]
- Chopin, E.I.B.; Marin, B.; Mkoungafoko, R.; Rigaux, A.; Hopgood, M.J.; Delannoy, E.; Cances, B.; Laurain, M. Factors Affecting Distribution and Mobility of Trace Elements (Cu, Pb, Zn) in a Perennial Grapevine (Vitis vinifera L.) in the Champagne Region of France. Environ. Pollut. 2008, 156, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Luo, C.; Li, X.; Shen, Z. Copper Accumulation and Tolerance in Chrysanthemum coronarium L., and Sorghum sudanense L. Arch. Environ. Contam. Toxicol. 2008, 55, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Ma, Y.; Hua, L.; Mclaughlin, M.J. Identification of Hydroxyl Copper Toxicity to Barley (Hordeum vulgare) Root Elongation in Solution Culture. Environ. Toxicol. Chem. 2009, 28, 662–666. [Google Scholar] [CrossRef]
- Juang, K.W.; Lai, H.Y.; Chen, B.C. Coupling Bioaccumulation and Phytotoxicity to Predict Copper Removal by Switchgrass Grown Hydroponically. Ecotoxicology 2011, 20, 827–835. [Google Scholar] [CrossRef]
- Michaud, A.M.; Chappellaz, C.; Hinsinger, P. Copper Phytotoxicity Affects Root Elongation and Iron Nutrition in Durum Wheat (Triticum turgidum durum L.). Plant Soil 2008, 310, 151–165. [Google Scholar] [CrossRef]
- Brun, L.A.; Maillet, J.; Richarte, J.; Herrmann, P.; Remy, J.C. Relationships between Extractable Copper, Soil Properties and Copper Uptake by Wild Plants in Vineyard Soils. Environ. Pollut. 1998, 102, 151–161. [Google Scholar] [CrossRef]
- Devez, A.; Gomez, E.; Gilbin, R.; Elbaz-Poulichet, F.; Persin, F.; Andrieux, P.; Casellas, C. Assessment of Copper Bioavailability and Toxicity in Vineyard Runoff Waters by DPASV and Algal Bioassay. Sci. Total Environ. 2005, 348, 82–92. [Google Scholar] [CrossRef]
- Lock, K.; De Schamphelaere, K.A.C.; Becaus, S.; Criel, P.; Van Eeckhout, H.; Janssen, C.R. Development and Validation of a Terrestrial Biotic Ligand Model Predicting the Effect of Cobalt on Root Growth of Barley (Hordeum vulgare). Environ. Pollut. 2007, 147, 626–633. [Google Scholar] [CrossRef]
- Angelova, V.R.; Ivanov, A.S.; Braikov, D.M. Heavy Metals (Pb, Cu, Zn and Cd) in the System Soil-Grapevine-Grape. J. Sci. Food Agric. 1999, 79, 713–721. [Google Scholar] [CrossRef]
- Ko, B.G.; Vogeler, I.; Bolan, N.S.; Clothier, B.; Green, S.; Kennedy, J. Mobility of Copper, Chromium and Arsenic from Treated Timber into Grapevines. Sci. Total Environ. 2007, 388, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Tandy, S.; Schulin, R.; Nowack, B. The Influence of EDDS on the Uptake of Heavy Metals in Hydroponically Grown Sunflowers. Chemosphere 2006, 62, 1454–1463. [Google Scholar] [CrossRef]
- Chen, P.Y.; Lee, Y.I.; Chenc, B.C.; Juang, K.W. Effects of Calcium on Rhizotoxicity and the Accumulation and Translocation of Copper by Grapevines. Plant Physiol. Biochem. 2013, 73, 375–382. [Google Scholar] [CrossRef]
- Vršič, S. Soil Erosion and Earthworm Population Responses to Soil Management Systems in Steep-Slope Vineyards. Plant Soil Environ. 2011, 57, 258–263. [Google Scholar] [CrossRef]
- Kurnik, V.; Gaberšek, V.; Unuk, T.; Tojnko, S.; Vogrin, A.; Vajs, S.; Lešnik, M. Influence of Alternative Copper Fungicide Formulations on Copper Content in Apple Fruits. Erwerbs-Obstbau 2012, 54, 161–170. [Google Scholar] [CrossRef]
- Lai, H.Y.; Juang, K.W.; Chen, B.C. Copper Concentrations in Grapevines and Vineyard Soils in Central Taiwan. Soil Sci. Plant Nutr. 2010, 56, 601–606. [Google Scholar] [CrossRef]
- Busuioc, G.; Elekes, C.C.; Stihi, C.; Lordache, S.; Ciulei, S.C. The Bioaccumulation and Translocation of Fe, Zn, and Cu in Species of Mushrooms from Russula Genus. Environ. Sci. Pollut. Res. 2011, 18, 890–896. [Google Scholar] [CrossRef]
- Steffens, B.; Rasmussen, A. The physiology of adventitious roots. Plant Physiol. 2016, 170, 603–617. [Google Scholar] [CrossRef]
- Iwasaki, K.; Sakurai, K.; Takahashi, E. Copper Binding by the Root Cell Walls on Italian Reygrass and Red Clover. Soil Sci. Plant Nutr. 1990, 36, 431–440. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Root- Stock | Cu in Substrate (mg kg−1) | Root DW (g ± SE) | Rootstock Trunk DW (g ± SE) | Canes DW (g ± SE) | Biomass * (g ± SE) |
|---|---|---|---|---|---|
| 5BB | 0 | 10.30 bc ± 1.193 | 14.37 a ± 0.982 | 6.93 b ± 0.524 | 22.36 b ± 0.4817 |
| Oxy-50 | 8.80 c ± 1.021 | 16.17 a ± 0.913 | 6.77 b ± 0.318 | 21.17 b ± 1.9373 | |
| Oxy-150 | 7.85 c ± 0.556 | 15.85 a ± 0.822 | 8.65 ab ± 0.233 | 21.59 b ± 0.9361 | |
| Oxy-500 | 12.95 ab ± 0.388 | 16.15 a ± 0.654 | 10.08 a ± 0.466 | 28.03 ac ± 0.8164 | |
| Oxy-1000 | 14.98 a ± 1.342 | 15.15 a ± 0.210 | 10.78 a ± 1.247 | 30.17 a ± 2.0509 | |
| 0 | 10.30 b ± 1.193 | 14.37 b ± 0.982 | 6.93 b ± 0.524 | 22.36 b ± 0.4817 | |
| Glu-50 | 10.45 b ± 0.964 | 17.35 a ± 0.821 | 6.28 b ± 0.636 | 24.09 b ± 1.9974 | |
| Glu-150 | 13.48 ab ± 0.944 | 14.97 ab ± 0.522 | 7.58 b ± 0.520 | 26.58 b ± 1.4479 | |
| Glu-500 | 12.75 ab ± 1.229 | 15.90 ab ± 0.687 | 7.18 b ± 0.206 | 25.65 b ± 0.7395 | |
| Glu-1000 | 15.75 a ± 0.550 | 16.30 ab ± 0.200 | 11.35 a ± 1.550 | 32.58 a ± 2.6255 | |
| SO4 | 0 | 12.50 a ± 1.197 | 15.23 b ± 0.320 | 7.48 b ± 0.930 | 24.91 b ± 2.0395 |
| Oxy-50 | 11.77 a ± 0.296 | 16.00 b ± 0.153 | 7.33 b ± 0.384 | 24.54 b ± 0.1876 | |
| Oxy-150 | 11.83 a ± 0.828 | 15.30 ab ± 0.392 | 8.75 ab ± 0.501 | 25.52 b ± 0.7958 | |
| Oxy-500 | 14.23 a ± 1.050 | 17.17 a ± 0.554 | 10.50 a ± 0.521 | 30.90 a ± 1.5498 | |
| Oxy-1000 | 12.25 a ± 1.050 | 16.80 a ± 0.100 | 9.90 a ± 0.700 | 27.97 ab ± 0.8512 | |
| 0 | 12.50 b ± 1.197 | 15.23 a ± 0.320 | 7.48 ab ± 0.930 | 24.91 b ± 2.0395 | |
| Glu-50 | 15.70 ab ± 0.636 | 16.45 a ± 0.592 | 6.35 b ± 0.185 | 27.59 ab ± 0.4149 | |
| Glu-150 | 14.87 ab ± 1.147 | 16.50 a ± 0.306 | 8.30 ab ± 0.833 | 28.61 ab ± 1.6395 | |
| Glu-500 | 19.80 a ± 3.051 | 15.73 a ± 0.145 | 9.60 a ± 0.611 | 34.09 a ± 3.9063 | |
| Glu-1000 |
| Rootstock | Cu in Substrate (mg kg−1) | TFs | |||||
|---|---|---|---|---|---|---|---|
| Rootstock Trunk/Roots | Cane/Roots | Cane/Rootstock Trunk | |||||
| 5BB | 0 | 0.769 ± 0.381 a(AB) | 0.544 ± 0.077 (A) | 0.472 ± 0.069 a(A) | 0.248 ± 0.029 (B) (p = 0.043) | 0.762 ± 0.162 a(AB) | 0.764 ± 0.113 (A) |
| Oxy-50 | 0.396 ± 0.064 ab(AB) | 0.332 ± 0.076 b(ABC) | 0.756 ± 0.104 a(AB) | ||||
| Oxy-150 | 0.453 ± 0.087 ab(AB) | 0.256 ± 0.024 b(CD) | 0.503 ± 0.133 ab(ABC) | ||||
| Oxy-500 | 0.261 ± 0.041 b(B) | 0.105 ± 0.009 c(E) | 0.290 ± 0.062 b(BC) | ||||
| Oxy-1000 | 0.238 ± 0.062 b(B) | 0.072 ± 0.010 c(E) | 0.218 ± 0.056 b(C) | ||||
| 0 | 0.769 ± 0.381 a(AB) | 0.472 ± 0.069 a(A) | 0.762 ± 0.162 a(AB) | ||||
| Glu-50 | 0.914 ± 0.247 a(AB) | 0.449 ± 0.050 a(AB) | 0.992 ± 0.292 a(A) | ||||
| Glu-150 | 0.974 ± 0.370 a(AB) | 0.315 ± 0.074 ab(BC) | 0.646 ± 0.170 ab(ABC) | ||||
| Glu-500 | 0.399 ± 0.032 a(A) | 0.110 ± 0.020 c(DE) | 0.366 ± 0.076 ab(BC) | ||||
| Glu-1000 | 0.477 ± 0.169 a(A) | 0.151 ± 0.043 bc(DE) | 0.229 ± 0.028 b(C) | ||||
| SO4 | 0 | 0.624 ± 0.069 a(AB) | 0.499 ± 0.044 (A) | 0.790 ± 0.205 a(A) | 0.379 ± 0.055 (A) | 0.654 ± 0.071 a(AB) | 0.533 ± 0.064 (A) |
| Oxy-50 | 0.627 ± 0.161 a(AB) | 0.511 ± 0.072 ab(AB) | 0.635 ± 0.133 a(AB) | ||||
| Oxy-150 | 0.477 ± 0.071 ab(ABC) | 0.360 ± 0.065 bc(AB) | 0.497 ± 0.093 ab(B) | ||||
| Oxy-500 | 0.392 ± 0.073 ab(BC) | 0.140 ± 0.015 bc(B) | 0.232 ± 0.041 b(B) | ||||
| Oxy-1000 | 0.215 ± 0.003 b(C) | 0.064 ± 0.018 c(B) | 0.244 ± 0.017 b(B) | ||||
| 0 | 0.624 ± 0.069 aa(AB) | 0.790 ± 0.205 a(A) | 0.654 ± 0.071 a(AB) | ||||
| Glu-50 | 0.768 ± 0.145 a(A) | 0.521 ± 0.090 a(AB) | 1.640 ± 0.473 a(A) | ||||
| Glu-150 | 0.313 ± 0.017 b(AB) | 0.277 ± 0.069 ab(AB) | 1.197 ± 0.133 a(AB) | ||||
| Glu-500 | 0.390 ± 0.009 b(AB) | 0.164 ± 0.049 b(B) | 0.854 ± 0.186 a(AB) | ||||
| Glu-1000 | All plants failed | ||||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vršič, S.; Gumzej, M.; Lešnik, M.; Perko, A.; Pulko, B. Patterns of Copper Bioaccumulation and Translocation in Grapevine Grafts Depending on Rootstocks. Agriculture 2023, 13, 1768. https://doi.org/10.3390/agriculture13091768
Vršič S, Gumzej M, Lešnik M, Perko A, Pulko B. Patterns of Copper Bioaccumulation and Translocation in Grapevine Grafts Depending on Rootstocks. Agriculture. 2023; 13(9):1768. https://doi.org/10.3390/agriculture13091768
Chicago/Turabian StyleVršič, Stanko, Mojca Gumzej, Mario Lešnik, Andrej Perko, and Borut Pulko. 2023. "Patterns of Copper Bioaccumulation and Translocation in Grapevine Grafts Depending on Rootstocks" Agriculture 13, no. 9: 1768. https://doi.org/10.3390/agriculture13091768
APA StyleVršič, S., Gumzej, M., Lešnik, M., Perko, A., & Pulko, B. (2023). Patterns of Copper Bioaccumulation and Translocation in Grapevine Grafts Depending on Rootstocks. Agriculture, 13(9), 1768. https://doi.org/10.3390/agriculture13091768