
Morphological and Physiological Mechanism of Activating Insoluble Inorganic Phosphorus of Different Peanut (Arachis hypogaea L.) Varieties under Low Phosphorus

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Location of Experiment
2.2. Material
2.3. Experimental Design and Implementation
2.4. Determination Items and Methods
2.4.1. Peanut Root Morphological Index
2.4.2. Peanut Phosphorus Activation Rate
2.4.3. Root–Shoot Ratio
2.4.4. Root Activity
2.4.5. Root Secretion Capacity of H+
2.4.6. Collection and Determination of Organic Acids Secreted by Roots
2.5. Statistical Analysis
3. Results
3.1. Phosphorus Activation Rate of Different Peanut Varieties
3.2. Root Morphology of Peanut Varieties with Different P Activation Efficiencies
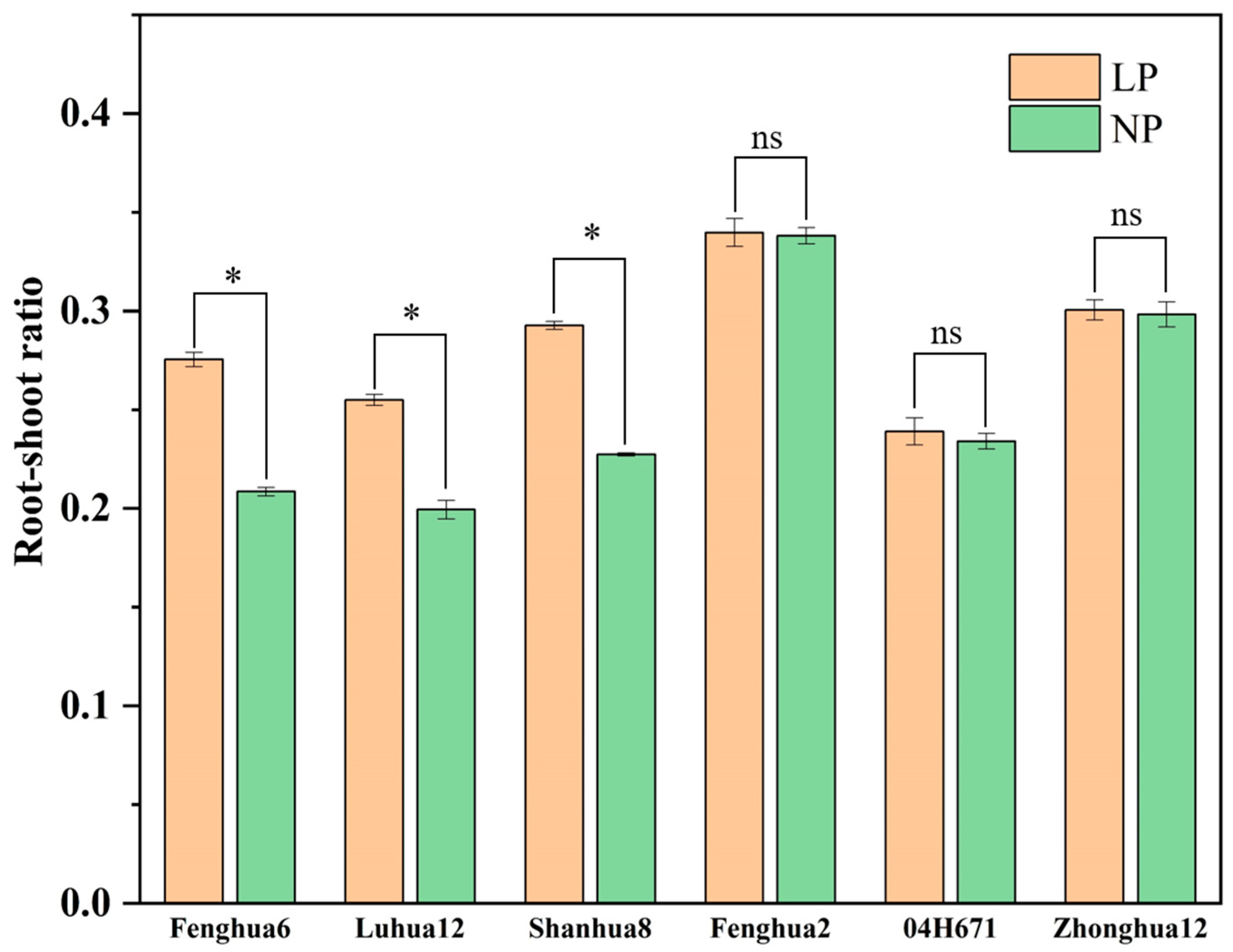
3.3. Root–Shoot Ratio of Peanut Varieties with Different P Activation Efficiencies
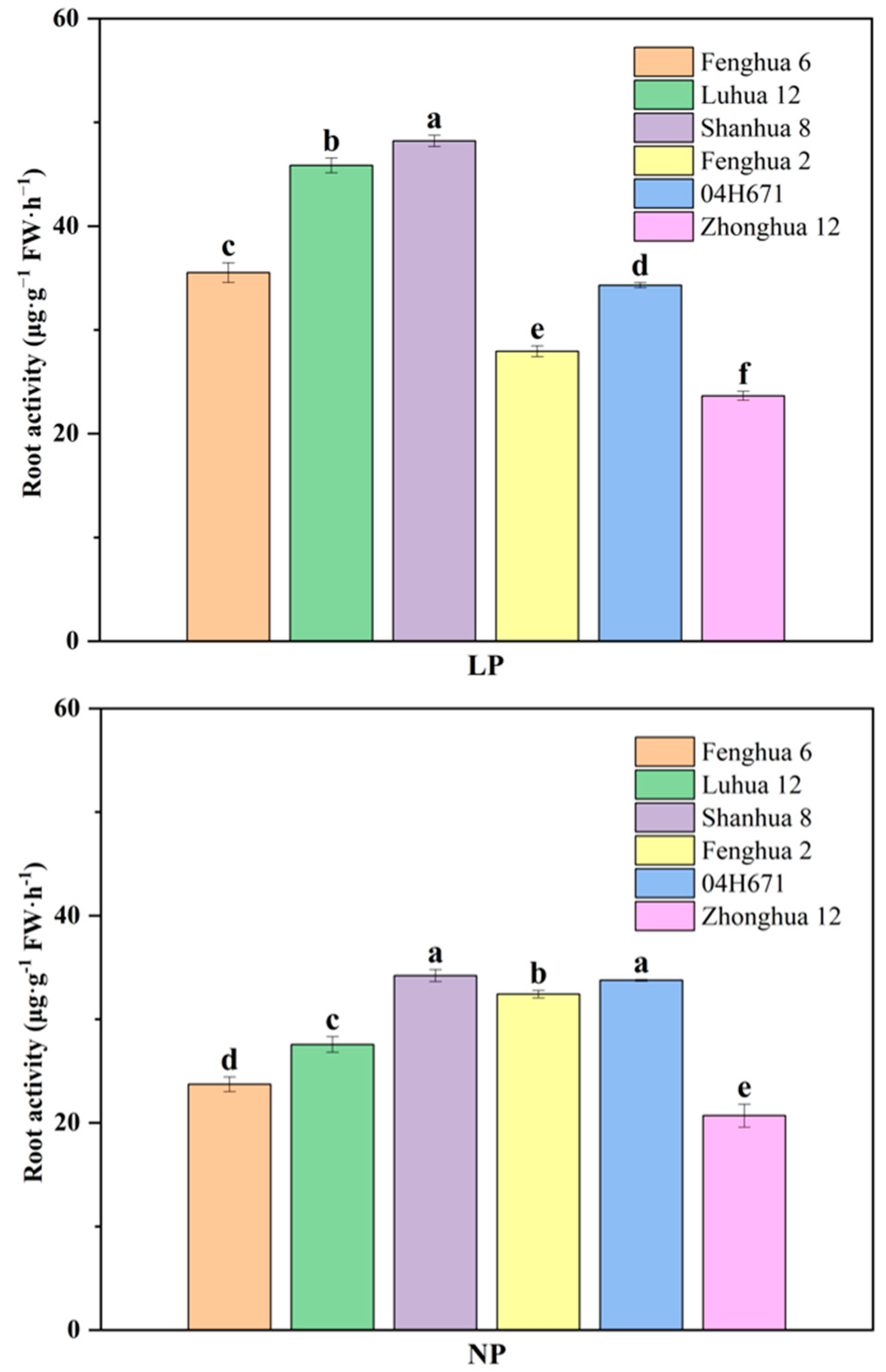
3.4. Root Activity of Peanut Varieties with Different P Activation Efficiencies
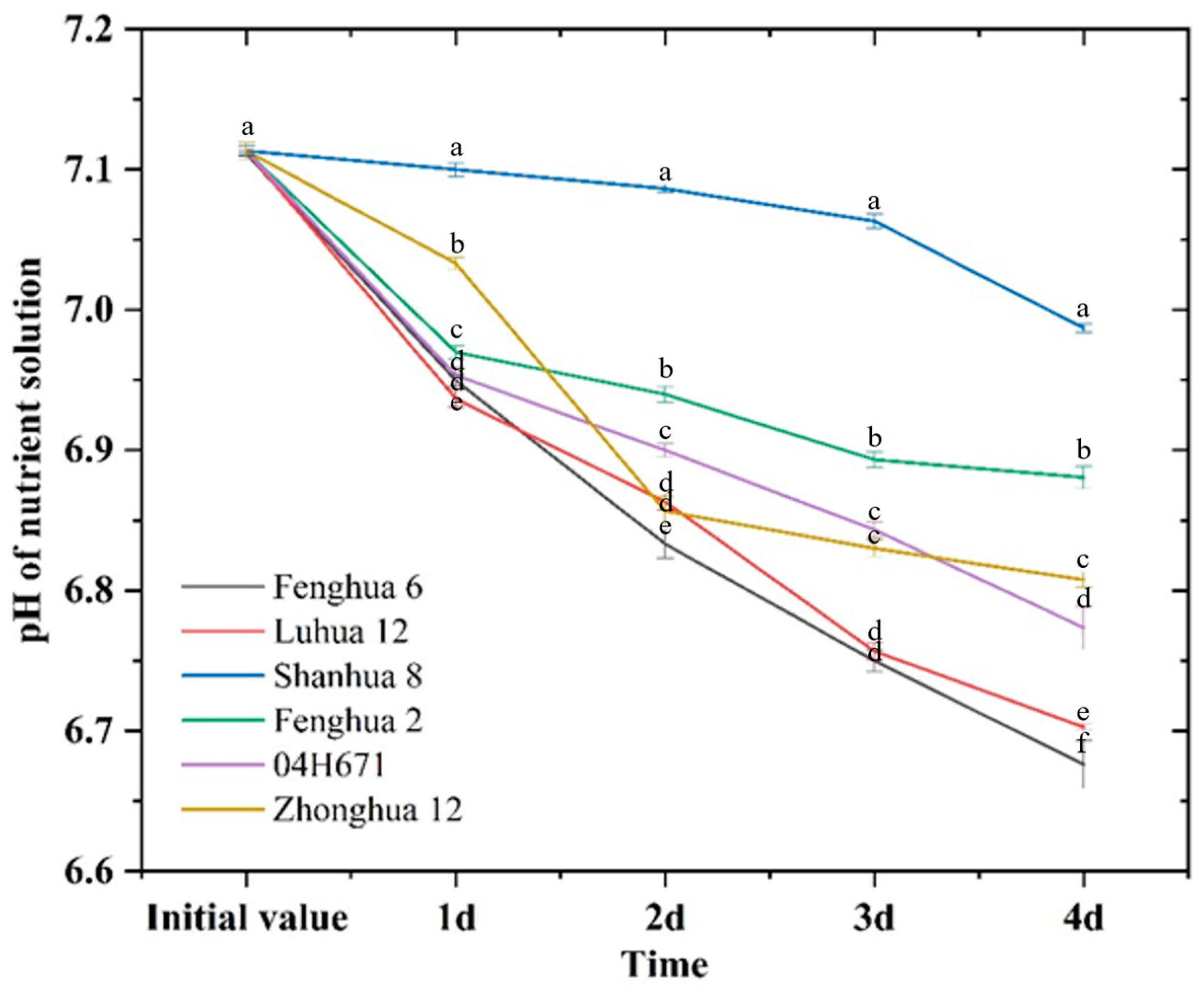
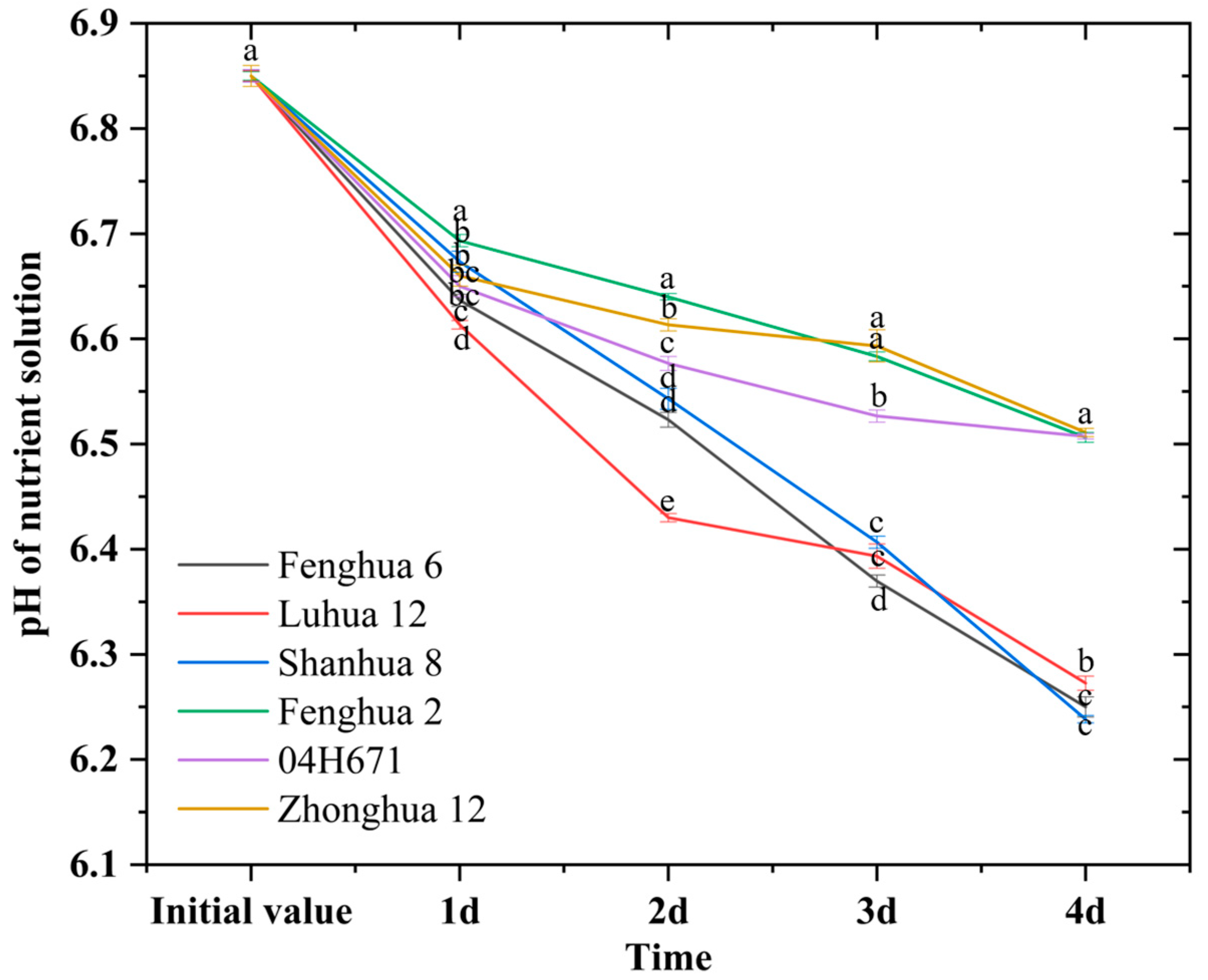
3.5. H+ Secretion Ability Peanut Varieties with Different P Activation Efficiency
3.6. Organic Acids Secreted by Roots of Peanut Varieties with Different P Activation Efficiencies
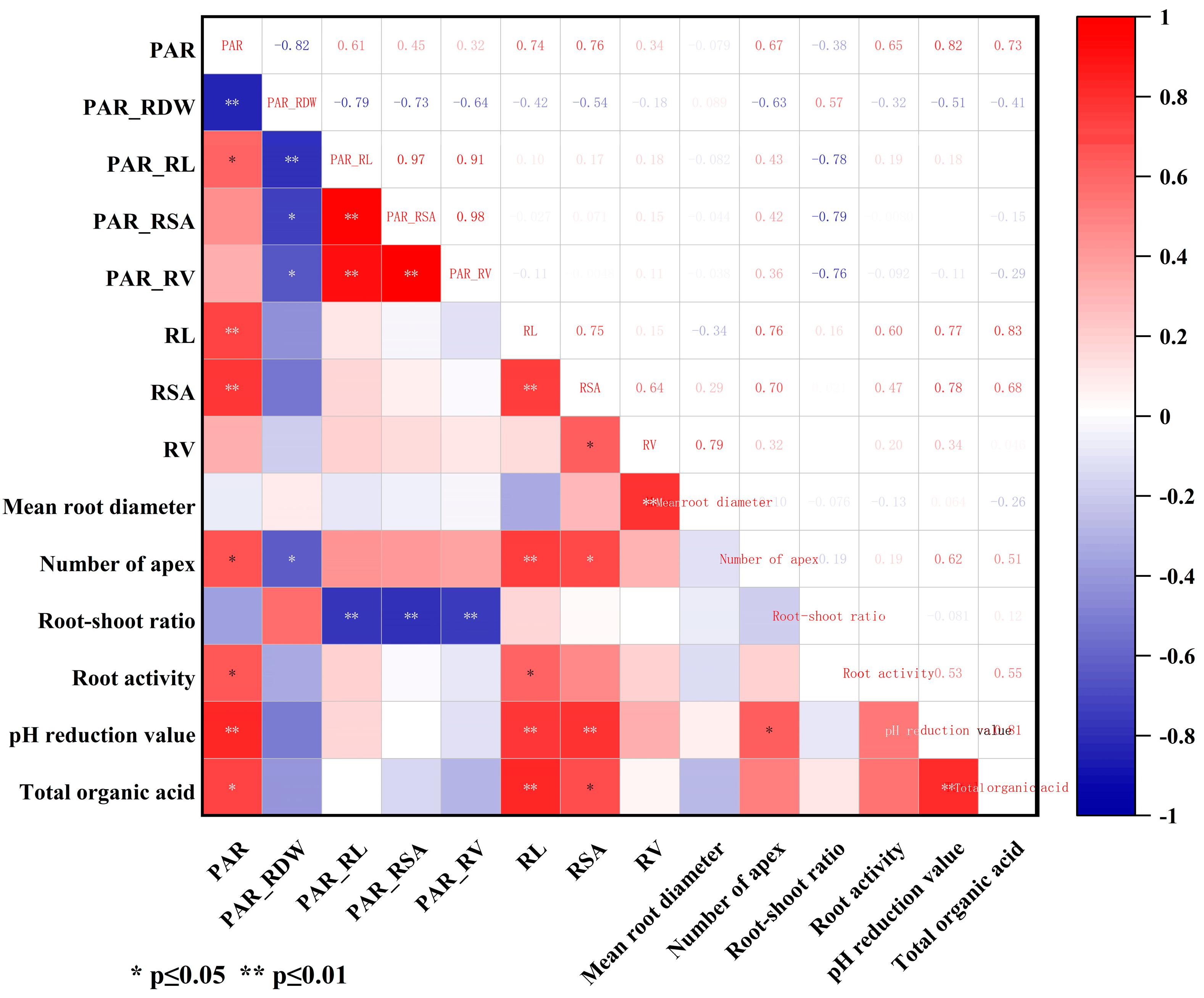
3.7. Correlation Analysis of Root Morphology and Root Exudates in Peanut Varieties with Different P Activation Efficiencies
4. Discussion
4.1. Determination of the Amount of Activated Phosphorus
4.2. Root Morphology of Peanut Varieties under Different P Concentrations
4.3. Organic Acid Secretion in Roots of Peanut Varieties under Different P Conditions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Dissanayaka, D.M.S.B.; Ghahremani, M.; Siebers, M.; Wasaki, J.; Plaxton, W.C. Recent insights into the metabolic adaptations of phosphorus-deprived plants. J. Exp. Bot. 2021, 72, 199–223. [Google Scholar] [CrossRef] [PubMed]
- Simpson, R.J.; Oberson, A.; Culvenor, R.A.; Ryan, M.H.; Veneklaas, E.J.; Lambers, H.; Lynch, J.P.; Ryan, P.R.; Delhaize, E.; Smith, F.A.; et al. Strategies and agronomic interventions to improve the phosphorus-use efficiency of farming systems. Plant Soil 2011, 349, 89–120. [Google Scholar] [CrossRef]
- Naab, J.B.; Boote, K.J.; Jones, J.W.; Porter, C.H. Adapting and evaluating the CROPGRO-peanut model for response to phosphorus on a sandy-loam soil under semi-arid tropical conditions. Field Crops Res. 2015, 176, 71–86. [Google Scholar] [CrossRef]
- Kadirimangalam, S.R.; Jadhav, Y.; Nagamadhuri, K.V. Genetic approaches for assessment of phosphorus use efficiency in groundnut (Arachis hypogaea L.). Sci. Rep. 2022, 12, 21552. [Google Scholar] [CrossRef]
- George, T.S.; Hinsinger, P.; Turner, B.L. Phosphorus in soils and plants-facing phosphorus scarcity. Plant Soil 2016, 401, 1–6. [Google Scholar] [CrossRef]
- van de Wiel, C.C.M.; van der Linden, C.G.; Scholten, O.E. Improving phosphorus use efficiency in agriculture: Opportunities for breeding. Euphytica 2015, 207, 1–22. [Google Scholar] [CrossRef]
- Li, B.; Li, P.; Zeng, X.C.; Huang, Y.F.; Wang, G.Q.; Young, B.R. Assessing the sustainability of Phosphorus use in China: Flow patterns from 1980 to 2015. Sci. Total Environ. 2019, 704, 135305. [Google Scholar] [CrossRef]
- Ghodszad, L.; Reyhanitabar, A.; Reza, M.; Lajayer, B.A. Biochar affects the fate of phosphorus in soil and water: A critical review. Chemosphere 2021, 283, 131176. [Google Scholar] [CrossRef]
- Kellogg, L.E.; Bridgham, S.D. Phosphorus retention and movement across an ombrotrophic-minerotrophic peatland gradient. Biogeochemistry 2003, 63, 299–315. [Google Scholar] [CrossRef]
- Job, M.T.P.; Souza, J.L.; Oliveira, J.R.; Filho, J.F.L.; Oliveira, T.S. Changes in inorganic phosphorus fractions in weathered soils under long-term intensive cultivation and irrigation. Arch. Agron. Soil Sci. 2023, 69, 1177–1192. [Google Scholar] [CrossRef]
- Zhu, J.; Li, M.; Whelan, M. Phosphorus activators contribute to legacy phosphorus availability in agricultural soils: A review. Sci. Total Environ. 2018, 612, 522–537. [Google Scholar] [CrossRef]
- Devau, N.; Hinsinger, P.; Cadre, E.L.; Colomb, B.; Gérard, F. Fertilization and pH effects on processes and mechanisms controlling dissolved inorganic phosphorus in soils. Geochim. Et Cosmochim. Acta 2011, 75, 2980–2996. [Google Scholar] [CrossRef]
- Wu, Y.; Zhao, C.; Zhao, X.; Yang, L.; Liu, C.; Jiang, L.; Liu, G.; Liu, P.; Luo, L. Multi-omics-based identification of purple acid phosphatases and metabolites involved in phosphorus recycling in stylo root exudates. Int. J. Biol. Macromol. 2023, 241, 124569. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, C.A.G.; de Sousa Tinoco, S.M.; de Souza, V.F.; Negri, B.F.; Gault, C.M.; Pastina, M.M.; Magalhaes, J.V.; Guimarães, L.J.M.; de Barros, E.G.; Buckler, E.S. Genome-Wide Association Study for Root Morphology and Phosphorus Acquisition Efficiency in Diverse Maize Panels. Int. J. Mol. Sci. 2023, 24, 6233. [Google Scholar] [CrossRef] [PubMed]
- Zhu, S.; Luo, L.; Zhang, X.; Zhao, M.; Wang, X.; Zhang, J.; Wan, Q.; Li, X.; Wan, Y.; Zhang, K.; et al. Study on the Relationship of Root Morphology and Phosphorus Absorption Efficiency with Phosphorus Uptake Capacity in 235 Peanut (Arachis hypogaea L.) Germplasms. Front. Environ. Sci. 2022, 10, 855815. [Google Scholar] [CrossRef]
- Wu, Y. Identification and Evaluation of Phosphorus Efficiency of New Varieties from National Consortium for Peanut Seed Research. Master’s Thesis, Shandong Agricultural University, Tai’an, China, 2022. [Google Scholar] [CrossRef]
- Wang, S.; Wu, M.; Zhong, S.; Sun, J.; Mao, X.; Qiu, N.; Zhou, F. A Rapid and Quantitative Method for Determining Seed Viability Using 2,3,5-Triphenyl Tetrazolium Chloride (TTC): With the Example of Wheat Seed. Molecules 2023, 28, 6828. [Google Scholar] [CrossRef]
- Wu, A.; Fang, Y.; Liu, S.; Wang, H.; Xu, B.; Zhang, S.; Deng, X. Root morphology and rhizosheath acid phosphatase activity in legume and graminoid species respond differently to low phosphorus supply. Rhizosphere 2021, 19, 100391. [Google Scholar] [CrossRef]
- Fan, J.W.; Du, Y.L.; Turner, N.C.; Wang, B.R.; Fang, Y.; Xi, Y.; Guo, X.R.; Li, F.M. Changes in root morphology and physiology to limited phosphorus and moisture in a locally-selected cultivar and an introduced cultivar of Medicago sativa L. growing in alkaline soil. Plant Soil 2015, 392, 215–226. [Google Scholar] [CrossRef]
- Meng, X.; Chen, W.W.; Wang, Y.Y.; Huang, Z.R.; Ye, X.; Chen, L.S.; Yang, L.T. Effects of phosphorus deficiency on the absorption of mineral nutrients, photosynthetic system performance and antioxidant metabolism in Citrus grandis. PLoS ONE 2021, 16, e0246944. [Google Scholar] [CrossRef]
- Ao, J.; Fu, J.; Tian, J. Genetic variability for root morph-architecture traits and root growth dynamics as related to phosphorus efficiency in soybean. Funct. Plant Biol. 2010, 37, 304–312. [Google Scholar] [CrossRef]
- Nguyen, V.L.; Stangoulis, J. Variation in root system architecture and morphology of two wheat genotypes is a predictor of their tolerance to phosphorus deficiency. Acta Physiol. Plant. 2019, 41, 109. [Google Scholar] [CrossRef]
- Neumann, G.; Römheld, V. Root excretion of carboxylic acids and protons in phosphorus-deficient plants. Plant Soil 1999, 211, 121–130. [Google Scholar] [CrossRef]
- Ryan, P.; Delhaize, E.; Jones, D. Function and mechanism of organic anion exudation from plant roots. Annu. Rev. Plant Physiol. Plant Mol. Biol. 2001, 52, 527–560. [Google Scholar] [CrossRef]
- Wang, Y.; Lambers, H. Root-released organic anions in response to low phosphorus availability: Recent progress, challenges and future perspectives. Plant Soil 2019, 447, 135–156. [Google Scholar] [CrossRef]
- Justi, M.; Silva, C.A.; Rosa, S.D. Organic acids as complexing agents for iron and their effects on the nutrition and growth of maize and soybean. Arch. Agron. Soil Sci. 2022, 68, 1369–1384. [Google Scholar] [CrossRef]
- Raghothama, K.G.; Karthikeyan, A.S. Phosphate Acquisition. Plant Soil 2005, 274, 37–49. [Google Scholar] [CrossRef]
- Krishnappa, R.; Aftab Hussain, I.S. Phosphorus acquisition from deficient soil: Involvement of organic acids and acid phosphatase in pigeon pea (Cajanus cajan L. mills sp.). Indian J. Plant Physiol. 2014, 19, 197–204. [Google Scholar] [CrossRef]
- Chen, Y.; Dunbabin, V.M.; Diggle, A.J.; Siddique, K.H.M.; Rengel, Z. Phosphorus starvation boosts carboxylate secretion in P-deficient genotypes of Lupinus angustifolius with contrasting root structure. Crop Pasture Sci. 2013, 64, 588–599. [Google Scholar] [CrossRef]
- Dinkelaker, B.; Römheld, V.; Marschner, H. Citric acid excretion and precipitation of calcium citrate in the rhizosphere of white lupin (Lupinus albus L.). Plant Cell Environ. 1989, 12, 285–292. [Google Scholar] [CrossRef]
- Mander, C.; Wakelin, S.; Young, S.; Condron, L.; O’Callaghan, M. Incidence and diversity of phosphate-solubilising bacteria are linked to phosphorus status in grassland soils. Soil Biol. Biochem. 2012, 44, 93–101. [Google Scholar] [CrossRef]
- Jalali, M.; Jalali, M. Effect of Low-Molecular-Weight Organic Acids on the Release of Phosphorus from Amended Calcareous Soils: Experimental and Modeling. J. Soil Sci. Plant Nutr. 2022, 22, 4179–4193. [Google Scholar] [CrossRef]
- Zhao, K.; Wu, Y. Rhizosphere calcareous soil P-extraction at the expense of organic carbon from root-exuded organic acids induced by phosphorus deficiency in several plant species. Soil Sci. Plant Nutr. 2014, 60, 640–650. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Varieties | PAR | PAR_RDW | PAR_RL | PAR_RSA | PAR_RV |
|---|---|---|---|---|---|
| mg d−1 Plant−1 | mg d−1 g−1 | μg d−1 cm−1 | μg d−1 cm−2 | mg d−1 cm−3 | |
| Fenghua 6 | 8.18 ± 0.75 a | 12.88 ± 0.39 d | 14.70 ± 0.27 b | 99.43 ± 1.11 b | 5.56 ± 0.23 a |
| Luhua 12 | 8.15 ± 0.01 a | 13.07 ± 0.51 d | 15.71 ± 0.26 a | 101.64 ± 1.00 a | 5.75 ± 0.13 a |
| Shanhua 8 | 6.08 ± 0.39 b | 24.58 ± 0.99 c | 15.44 ± 0.27 a | 97.68 ± 0.46 c | 5.67 ± 0.12 a |
| Fenghua 2 | 4.72 ± 0.34 c | 24.59 ± 0.39 c | 5.58 ± 0.43 d | 38.23 ± 0.99 e | 2.30 ± 0.01 b |
| 04H671 | 3.12 ± 0.16 d | 39.34 ± 0.83 a | 4.50 ± 0.49 e | 24.61 ± 0.63 f | 0.97 ± 0.08 d |
| Zhonghua 12 | 4.41 ± 0.26 c | 27.83 ± 0.57 b | 7.36 ± 0.26 c | 44.07 ± 0.46 d | 1.73 ± 0.01 c |
| Varieties | PAR | PAR_RDW | PAR_RL | PAR_RSA | PAR_RV |
|---|---|---|---|---|---|
| mg d−1 Plant−1 | mg d−1 g−1 | μg d−1 cm−1 | μg d−1 cm−2 | mg d−1 cm−3 | |
| Fenghua 6 | 9.89 ± 0.25 b | 14.26 ± 0.56 e | 10.03 ± 0.33 c | 57.59 ± 0.53 a | 2.88 ± 0.10 a |
| Luhua 12 | 10.72 ± 0.35 a | 14.75 ± 0.74 e | 10.97 ± 0.30 b | 57.75 ± 0.36 a | 2.41 ± 0.05 b |
| Shanhua 8 | 10.92 ± 0.40 a | 16.75 ± 0.77 d | 11.97 ± 0.59 a | 55.54 ± 0.68 b | 2.23 ± 0.02 c |
| Fenghua 2 | 3.55 ± 0.12 e | 46.67 ± 0.54 a | 3.28 ± 0.27 f | 19.08 ± 0.49 e | 0.93 ± 0.10 e |
| 04H671 | 4.71 ± 0.31 d | 34.64 ± 0.52 b | 4.81 ± 0.30 e | 23.64 ± 0.04 d | 1.01 ± 0.05 e |
| Zhonghua 12 | 6.26 ± 0.26 c | 31.33 ± 0.20 c | 5.51 ± 0.39 d | 29.20 ± 0.49 c | 1.18 ± 0.07 d |
| Varieties | Root Length (cm) | Root Surface Area (cm2) | Root Volume (cm3) | Mean Diameter (mm) | Root Tips | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| LP | NP | LP | NP | LP | NP | LP | NP | LP | NP | |
| Fenghua 6 | 1444.25 a | 1333.86 a | 297.25 a | 248.67 b | 7.53 c | 6.03 c | 0.75 c | 0.68 f | 860.67 a | 919.67 a |
| Luhua 12 | 1397.57 a | 1090.54 cd | 299.21 a | 264.99 a | 9.77 a | 9.49 a | 0.81 a | 0.86 a | 857.67 a | 738.83 c |
| Shanhua 8 | 1432.66 a | 1135.7 c | 263.48 b | 223.39 c | 6.83 e | 6.7 b | 0.67 e | 0.71 e | 766 b | 690.5 d |
| Fenghua 2 | 1232.38 b | 1240.31 b | 246.61 c | 251.79 b | 8.65 b | 6.74 b | 0.8 b | 0.73 d | 739 c | 739.2 c |
| 04H671 | 1131.12 c | 1031.01 e | 224.21 d | 213.77 c | 5.15 f | 5.79 d | 0.74 cd | 0.74 c | 615.75 e | 763.75 b |
| Zhonghua 12 | 1249.38 b | 1073.64 de | 261.06 b | 224.67 c | 7.14 d | 6.66 b | 0.74 d | 0.76 b | 715.5 d | 672.17 e |
| V | *** | *** | *** | *** | *** | |||||
| T | *** | *** | ns | ns | ns | |||||
| V × T | * | * | * | * | *** | |||||
| Varieties | Oxalic | Tartaric | Formic | Acetic | Total Amount | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| NP | LP | NP | LP | NP | LP | NP | LP | NP | LP | |
| Fenghua 6 | 0.3281 a | 0.6642 a | 0.0225 a | 0.0374 a | 0.0012 b | 0.0030 a | 0.0010 b | 0.0070 a | 0.3528 a | 0.7115 a |
| Luhua 12 | 0.0720 e | 0.6041 a | 0.0084 d | 0.0157 cd | 0.0017 a | 0.0024 b | 0.0013 a | 0.0023 b | 0.0833 f | 0.6246 b |
| Shanhua 8 | 0.1321 d | 0.6101 a | 0.0147 c | 0.0168 c | 0.0009 c | 0.0017 d | 0.0013 a | 0.0014 c | 0.1489 e | 0.6299 b |
| Fenghua 2 | 0.1797 c | 0.1428 d | 0.0161 bc | 0.0252 b | 0.0010 c | 0.0019 c | 0.0010 b | 0.0012 cd | 0.1976 d | 0.1711 e |
| 04H671 | 0.2123 b | 0.2648 c | 0.0145 c | 0.0161 cd | 0.0012 b | 0.0020 c | 0.0009 b | 0.0014 c | 0.2288 c | 0.2842 d |
| Zhonghua 12 | 0.3146 a | 0.4111 b | 0.0173 b | 0.0151 d | 0.0010 c | 0.0017 d | 0.0007 c | 0.0009 d | 0.3334 b | 0.4288 c |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, Z.; Liu, F.; Wan, Y.; Zhu, S.; Zhang, J.; Zhang, K.; Luo, L. Morphological and Physiological Mechanism of Activating Insoluble Inorganic Phosphorus of Different Peanut (Arachis hypogaea L.) Varieties under Low Phosphorus. Agriculture 2023, 13, 2270. https://doi.org/10.3390/agriculture13122270
Tan Z, Liu F, Wan Y, Zhu S, Zhang J, Zhang K, Luo L. Morphological and Physiological Mechanism of Activating Insoluble Inorganic Phosphorus of Different Peanut (Arachis hypogaea L.) Varieties under Low Phosphorus. Agriculture. 2023; 13(12):2270. https://doi.org/10.3390/agriculture13122270
Chicago/Turabian StyleTan, Zhen, Fengzhen Liu, Yongshan Wan, Suqing Zhu, Jing Zhang, Kun Zhang, and Lu Luo. 2023. "Morphological and Physiological Mechanism of Activating Insoluble Inorganic Phosphorus of Different Peanut (Arachis hypogaea L.) Varieties under Low Phosphorus" Agriculture 13, no. 12: 2270. https://doi.org/10.3390/agriculture13122270
APA StyleTan, Z., Liu, F., Wan, Y., Zhu, S., Zhang, J., Zhang, K., & Luo, L. (2023). Morphological and Physiological Mechanism of Activating Insoluble Inorganic Phosphorus of Different Peanut (Arachis hypogaea L.) Varieties under Low Phosphorus. Agriculture, 13(12), 2270. https://doi.org/10.3390/agriculture13122270