

What Do Cross-Range Germination, Growth, and Interaction Studies Reveal about the Behaviour of an Expansive Plant Species?

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Plants
2.2. Seeds and Propagules Procurement
2.3. Growing Media Types
2.4. Cross-Range Germination Test
2.5. Growth Experiment
2.6. Interaction Experiments
2.7. Data Analysis
2.7.1. Germination Test
2.7.2. Growth Experiment
2.7.3. Interaction Experiments
3. Results
3.1. Germination Test
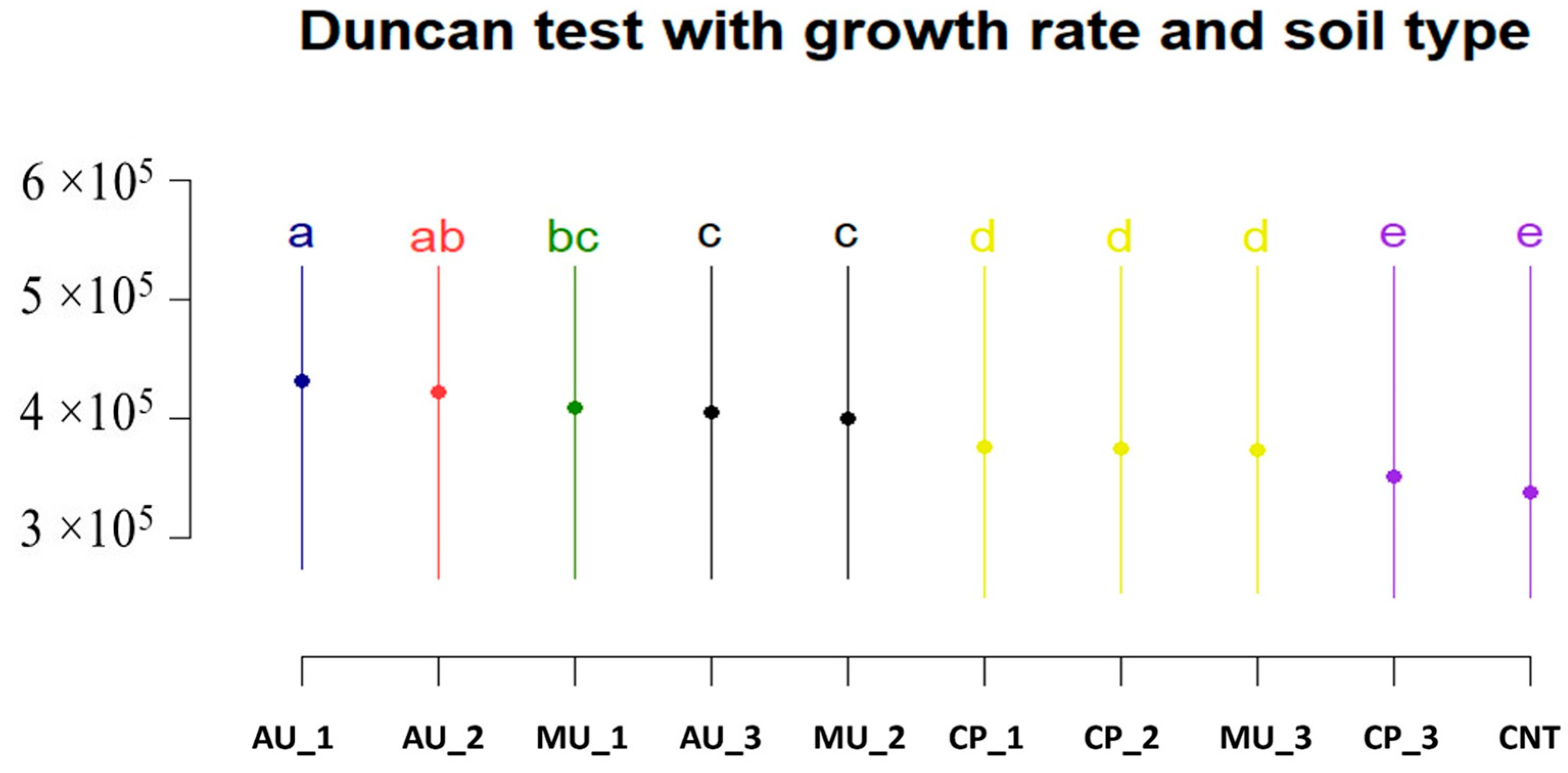
3.2. Growth Experiment
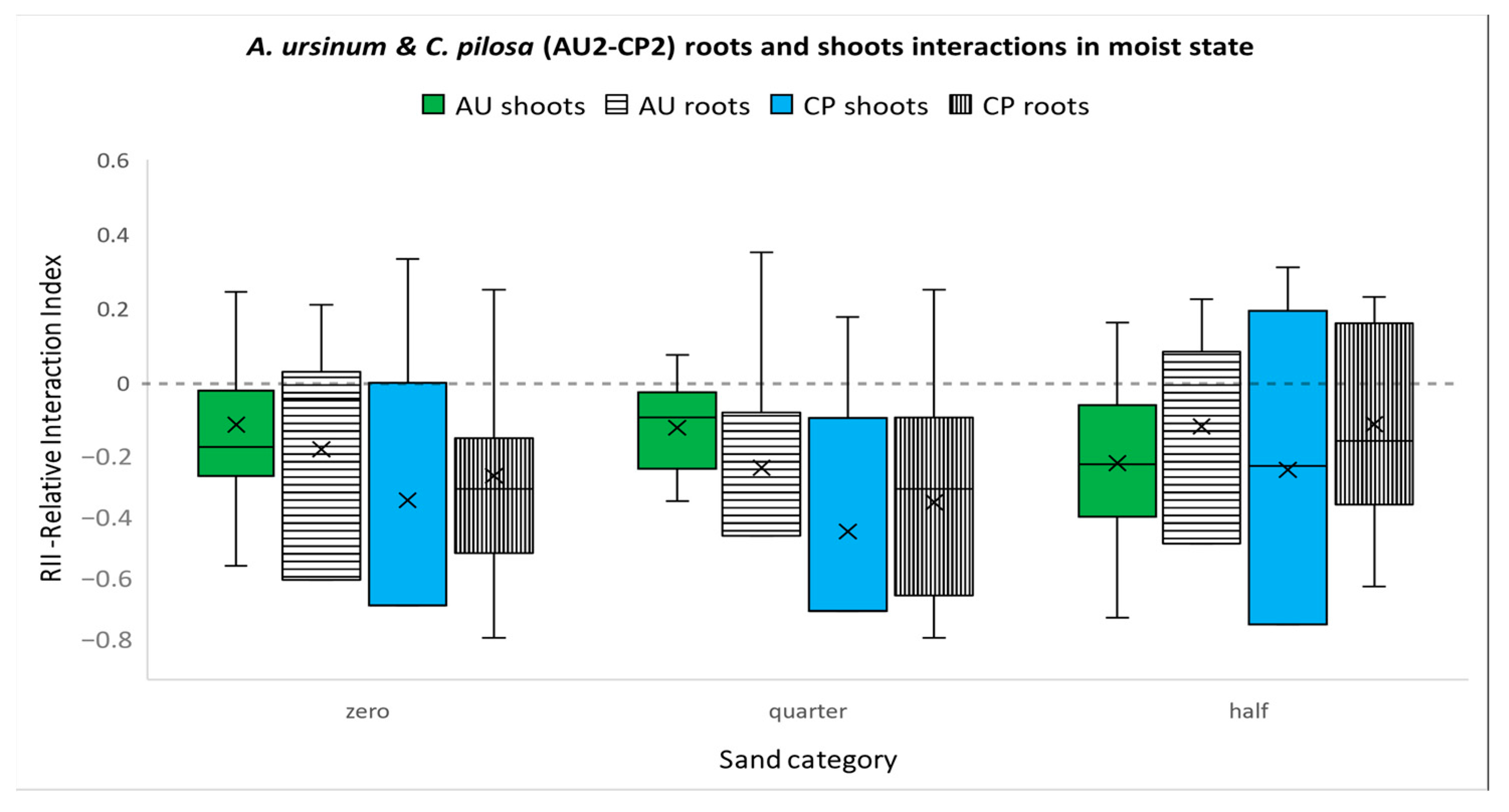
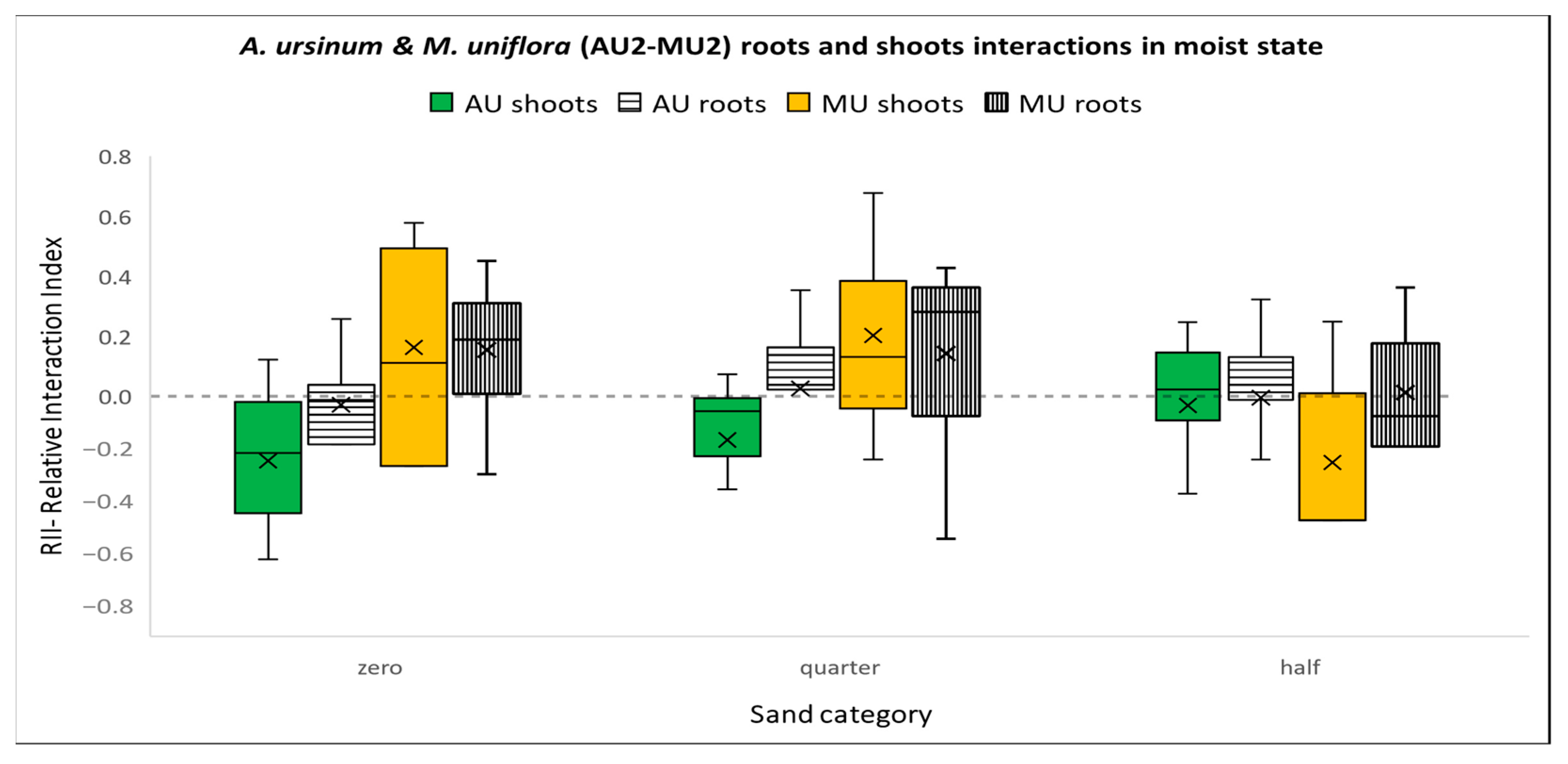
3.3. Interaction Experiments
3.3.1. Competition between A. ursinum and C. majalis under Three Moisture Categories (Figure 4)

3.3.2. Competition between A. ursinum and C. pilosa in a Wet Environment (Figure 5)

3.3.3. Competition between A. ursinum and M. uniflora in a Wet Environment (Figure 6)

3.3.4. Competition between A. ursinum and C. pilosa in Moist Environments (Figure 7)

3.3.5. Competition between A. ursinum and M. uniflora in a Moist Environment (Figure 8)

4. Discussion
4.1. Cross-Range Germination Test
4.2. Growth Experiment
4.3. Interaction Experiments
5. Conclusive Remarks
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harper, J.L. Population Biology of Plants; Academic Press Inc.: London, UK, 1977; ISBN 19782321379. [Google Scholar]
- Valladares, F.; Bastias, C.C.; Godoy, O.; Granda, E.; Escudero, A. Species Coexistence in a Changing World. Front. Plant Sci. 2015, 6, 866. [Google Scholar] [CrossRef]
- Švamberková, E.; Vítová, A.; Lepš, J. The Role of Biotic Interactions in Plant Community Assembly: What Is the Community Species Pool? Acta Oecologica 2017, 85, 150–156. [Google Scholar] [CrossRef]
- Saura-Mas, S.; Bonas, A.; Lloret, F. Plant Community Response to Drought-Induced Canopy Defoliation in a Mediterranean Quercus Ilex Forest. Eur. J. For. Res. 2015, 134, 261–272. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Anderegg, L.D.L.; Sherman, C.; Karp, D.S. Effects of Widespread Drought-induced Aspen Mortality on Understory Plants. Conserv. Biol. 2012, 26, 1082–1090. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.T. Succession Analysis of Plant Communities in Abandoned Croplands in the Eastern Loess Plateau of China. J. Arid Environ. 2005, 63, 458–474. [Google Scholar] [CrossRef]
- Gilliam, F.S. Response of the Herbaceous Layer of Forest Ecosystems to Excess Nitrogen Deposition. J. Ecol. 2006, 94, 1176–1191. [Google Scholar] [CrossRef]
- Gilliam, F.S.; Welch, N.T.; Phillips, A.H.; Billmyer, J.H.; Peterjohn, W.T.; Fowler, Z.K.; Walter, C.A.; Burnham, M.B.; May, J.D.; Adams, M.B. Twenty-Five-Year Response of the Herbaceous Layer of a Temperate Hardwood Forest to Elevated Nitrogen Deposition. Ecosphere 2016, 7, e01250. [Google Scholar] [CrossRef]
- Bricca, A.; Conti, L.; Tardella, M.F.; Catorci, A.; Iocchi, M.; Theurillat, J.-P.; Cutini, M. Community Assembly Processes along a Sub-Mediterranean Elevation Gradient: Analyzing the Interdependence of Trait Community Weighted Mean and Functional Diversity. Plant Ecol. 2019, 220, 1139–1151. [Google Scholar] [CrossRef]
- Bricca, A.; Tardella, F.M.; Tolu, F.; Goia, I.; Ferrara, A.; Catorci, A. Disentangling the Effects of Disturbance from Those of Dominant Tall Grass Features in Driving the Functional Variation of Restored Grassland in a Sub-Mediterranean Context. Diversity 2020, 12, 11. [Google Scholar] [CrossRef]
- Callaway, R.M.; Aschehoug, E.T. Invasive Plants versus Their New and Old Neighbors: A Mechanism for Exotic Invasion. Science 2000, 290, 521–523. [Google Scholar] [CrossRef]
- Götzenberger, L.; de Bello, F.; Bråthen, K.A.; Davison, J.; Dubuis, A.; Guisan, A.; Lepš, J.; Lindborg, R.; Moora, M.; Pärtel, M.; et al. Ecological Assembly Rules in Plant Communities-Approaches, Patterns and Prospects. Biol. Rev. 2012, 87, 111–127. [Google Scholar] [CrossRef]
- Halassy, M.; Botta-Dukát, Z.; Csecserits, A.; Szitár, K.; Török, K. Trait-based Approach Confirms the Importance of Propagule Limitation and Assembly Rules in Old-field Restoration. Restor. Ecol. 2019, 27, 840–849. [Google Scholar] [CrossRef]
- Kaushik, K.; Bricca, A.; Mugnai, M.; Viciani, D.; Rudolf, K.; Somfalvi-tóth, K.; Morschhauser, T. Effects of a Dominant Species on the Functional Diversity of Coexisting Species in Temperate Deciduous Understorey. Plants 2021, 10, 2252. [Google Scholar] [CrossRef] [PubMed]
- Lepš, J. Scale- and Time-Dependent Effects of Fertilization, Mowing and Dominant Removal on a Grassland Community during a 15-Year Experiment. J. Appl. Ecol. 2014, 51, 978–987. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating Resources in Plant Communities: A General Theory of Invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Hierro, J.L.; Maron, J.L.; Callaway, R.M. A Biogeographical Approach to Plant Invasions: The Importance of Studying Exotics in Their Introduced and Native Range. J. Ecol. 2005, 93, 5–15. [Google Scholar] [CrossRef]
- Callaway, R.M.; Maron, J.L. What Have Exotic Plant Invasions Taught Us over the Past 20 Years? Trends Ecol. Evol. 2006, 21, 369–374. [Google Scholar] [CrossRef] [PubMed]
- Catford, J.A.; Vesk, P.A.; White, M.D.; Wintle, B.A. Hotspots of Plant Invasion Predicted by Propagule Pressure and Ecosystem Characteristics. Divers. Distrib. 2011, 17, 1099–1110. [Google Scholar] [CrossRef]
- Catford, J.A.; Jansson, R.; Nilsson, C. Reducing Redundancy in Invasion Ecology by Integrating Hypotheses into a Single Theoretical Framework. Divers. Distrib. 2009, 15, 22–40. [Google Scholar] [CrossRef]
- Hulme, P.E.; Bacher, S.; Kenis, M.; Klotz, S.; Kühn, I.; Minchin, D.; Nentwig, W.; Olenin, S.; Panov, V.; Pergl, J.; et al. Grasping at the Routes of Biological Invasions: A Framework for Integrating Pathways into Policy. J. Appl. Ecol. 2008, 45, 403–414. [Google Scholar] [CrossRef]
- Hulme, P.E. Trade, Transport and Trouble: Managing Invasive Species Pathways in an Era of Globalization. J. Appl. Ecol. 2009, 46, 10–18. [Google Scholar] [CrossRef]
- Pyšek, P.; Richardson, D.M.; Rejmánek, M.; Webster, G.L.; Williamson, M.; Kirschner, J. Alien Plants in Checklists and Floras: Towards Better Communication between Taxonomists and Ecologists. Taxon 2004, 53, 131–143. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmánek, M.; Barbour, M.G.; Dane Panetta, F.; West, C.J. Naturalization and Invasion of Alien Plants: Concepts and Definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Wang, Y.J.; Chen, D.; Yan, R.; Yu, F.H.; van Kleunen, M. Invasive Alien Clonal Plants Are Competitively Superior over Co-Occurring Native Clonal Plants. Perspect. Plant Ecol. Evol. Syst. 2019, 40, 125484. [Google Scholar] [CrossRef]
- Linder, H.P.; Lehmann, C.E.R.; Archibald, S.; Osborne, C.P.; Richardson, D.M. Global Grass (Poaceae) Success Underpinned by Traits Facilitating Colonization, Persistence and Habitat Transformation. Biol. Rev. 2018, 93, 1125–1144. [Google Scholar] [CrossRef] [PubMed]
- Edmund, T.; Linders, W.; Abebe, A.; Kevin, S.; Lisanework, C.; Shiferaw, H.; Allan, E. Direct and Indirect Effects of Invasive Species : Biodiversity Loss Is a Major Mechanism by Which an Invasive Tree Affects Ecosystem Functioning. J. Ecol. 2019, 107, 2660–2672. [Google Scholar] [CrossRef]
- Early, R.; Bradley, B.A.; Dukes, J.S.; Lawler, J.J.; Olden, J.D.; Blumenthal, D.M. Global Threats from Invasive Alien Species in the Twenty-First Century and National Response Capacities. Nat. Commun. 2016, 7, 12485. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Pergl, J.; Sádlo, J.; Chytrý, M.; Jarošík, V. Invasion Success of Alien Plants: Do Habitat Affinities in the Native Distribution Range Matter? Glob. Ecol. Biogeogr. 2009, 18, 372–382. [Google Scholar] [CrossRef]
- Hejda, M.; Pyšek, P.; Jarošík, V. Impact of Invasive Plants on the Species Richness, Diversity and Composition of Invaded Communities. J. Ecol. 2009, 97, 393–403. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A Global Assessment of Invasive Plant Impacts on Resident Species, Communities and Ecosystems: The Interaction of Impact Measures, Invading Species’ Traits and Environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Hejda, M.; Chytrý, M.; Pergl, J.; Pyšek, P. Native-Range Habitats of Invasive Plants: Are They Similar to Invaded-Range Habitats and Do They Differ According to the Geographical Direction of Invasion? Divers. Distrib. 2015, 21, 312–321. [Google Scholar] [CrossRef]
- Fried, G.; Laitung, B.; Pierre, C.; Chagué, N.; Panetta, F.D. Impact of Invasive Plants in Mediterranean Habitats: Disentangling the Effects of Characteristics of Invaders and Recipient Communities. Biol. Invasions 2014, 16, 1639–1658. [Google Scholar] [CrossRef]
- Fried, G.; Carboni, M.; Mahaut, L.; Violle, C. Functional Traits Modulate Plant Community Responses to Alien Plant Invasion. Perspect. Plant Ecol. Evol. Syst. 2019, 37, 53–63. [Google Scholar] [CrossRef]
- Pysek, P. Clonality and plant invasions: Can a trait make a difference? In The Ecology and Evolution of Clonal Plants; Backhuys Publishers: Leiden, The Netherlands, 1997; pp. 405–427. [Google Scholar]
- Liu, J.; Dong, M.; Miao, S.L.; Li, Z.Y.; Song, M.H.; Wang, R.Q. Invasive Alien Plants in China: Role of Clonality and Geographical Origin. Biol. Invasions 2006, 8, 1461–1470. [Google Scholar] [CrossRef]
- Price, E.A.C.; Marshall, C. Clonal Plants and Environmental Heterogeneity—An Introduction to the Proceedings. Plant Ecol. 1999, 141, 3–7. [Google Scholar] [CrossRef]
- You, W.-H.; Han, C.-M.; Fang, L.-X.; Du, D.-L. Propagule Pressure, Habitat Conditions and Clonal Integration Influence the Establishment and Growth of an Invasive Clonal Plant, Alternanthera Philoxeroides. Front. Plant Sci. 2016, 7, 568. [Google Scholar] [CrossRef]
- Crall, A.W.; Renz, M.; Panke, B.J.; Newman, G.J.; Chapin, C.; Graham, J.; Bargeron, C. Developing Cost-Effective Early Detection Networks for Regional Invasions. Biol. Invasions 2012, 14, 2461–2469. [Google Scholar] [CrossRef]
- Reaser, J.K.; Burgiel, S.W.; Kirkey, J.; Brantley, K.A.; Veatch, S.D.; Burgos-Rodríguez, J. The Early Detection of and Rapid Response (EDRR) to Invasive Species: A Conceptual Framework and Federal Capacities Assessment. Biol. Invasions 2020, 22, 1–19. [Google Scholar] [CrossRef]
- Bruno, J.F.; Rand, T.A.; Emery, N.C.; Bertness, M.D. Facilitative and Competitive Interaction Components among New England Salt Marsh Plants. PeerJ 2017, 2017, e4049. [Google Scholar] [CrossRef][Green Version]
- Bertness, M.D.; Leonard, G.H.; Levine, J.M.; Schmidt, P.R.; Ingraham, A.O. Testing the Relative Contribution of Positive and Negative Interactions in Rocky Intertidal Communities. Ecology 1999, 80, 2711–2726. [Google Scholar] [CrossRef]
- Armas, C.; Ordiales, R.; Pugnaire, F.I. Measuring Plant Interactions: A New Comparative Index. Ecology 2004, 85, 2682–2686. [Google Scholar] [CrossRef]
- Pal, R.W.; Maron, J.L.; Nagy, D.U.; Waller, L.P.; Tosto, A.; Liao, H.; Callaway, R.M. What Happens in Europe Stays in Europe: Apparent Evolution by an Invader Does Not Help at Home. Ecology 2020, 101, e03072. [Google Scholar] [CrossRef]
- Dierschke, H. Long-Term Dynamics in the Herb Layer of a Calcareous Beech Forest: Investigations of Permanent Plots, 1981–2001. Pol. Bot. Stud. 2006, 22, 165–172. [Google Scholar]
- Dierschke, H. Constancy and Dynamics in a Species-Rich Calcareous Beech Forest. Changes within a Large Transect 1981–2011 (German: Konstanz Und Dynamik in Einem Artenreichen Kalkbuchenwald. Veränderungen in Einem Großtransekt 1981–2011). Tuexenia 2013, 33, 49–92. [Google Scholar]
- Morschhauser, T.; Rudolf, K.; Botta-Dukát, Z.; Oborny, B. Density-Dependence in the Establishment of Juvenile Allium ursinum Individuals in a Monodominant Stand of Conspecific Adults. Acta Oecol. 2009, 35, 621–629. [Google Scholar] [CrossRef]
- Oborny, B.; Botta-Dukát, Z.; Rudolf, K.; Morschhauser, T. Population Ecology of Allium ursinum, a Space-Monopolizing Clonal Plant. Acta Bot. Hung. 2011, 53, 371–388. [Google Scholar] [CrossRef]
- Ernst, W.H.O. Population Biology of Allium ursinum in Northern Germany. J. Ecol. 1979, 67, 347. [Google Scholar] [CrossRef]
- Borhidi, A.; Kevey, B.; Lendvai, G.; Seregélyes, T. Plant Communities of Hungary; Akademiai Kiado: Budapest, Hungary, 2012; ISBN 9789630592789. [Google Scholar]
- Rychnovská, M.; Bednář, V. Floodplain Forest: Herb Layer as Indicator of Its Ecological Status. Acta Univ. Palacki Olomuc. Fac. Rer. Nat. Biol. 1998, 36, 7–15. [Google Scholar]
- Sobolewska, D.; Podolak, I.; Makowska-Wąs, J. Allium ursinum: Botanical, Phytochemical and Pharmacological Overview. Phytochem. Rev. 2015, 14, 81–97. [Google Scholar] [CrossRef] [PubMed]
- Voća, S.; Žlabur, J.Š.; Uher, S.F.; Peša, M.; Opačić, N.; Radman, S. Neglected Potential of Wild Garlic (Allium ursinum L.)—Specialized Metabolites Content and Antioxidant Capacity of Wild Populations in Relation to Location and Plant Phenophase. Horticulturae 2022, 8, 24. [Google Scholar] [CrossRef]
- Lata, M. An Ethnobotanical Survey of Medicinal Plants Used by Tribal Migratory Shepherds in Hills of Tungasigarh of Thunag Subdivision of District Mandi Himachal Pradesh. Int. J. Chem. Stud. 2020, 8, 1071–1078. [Google Scholar] [CrossRef]
- Upadhyay, R.K. Garlic: A Potential Source of Pharmaceuticals and Pesticides: A Review. Int. J. Green Pharm. 2016, 10, S1–S28. [Google Scholar]
- Schmitt, B.; Schulz, H.; Storsberg, J.; Keusgen, M. Chemical Characterization of Allium ursinum L. Depending on Harvesting Time. J. Agric. Food Chem. 2005, 53, 7288–7294. [Google Scholar] [CrossRef] [PubMed]
- Błazewicz-Woźniak, M.; Kesik, T.; Ewa Michowska, A. Flowering of Bear Garlic (Allium ursinum L.) Cultivated in the Field at Varied Nitrogen Nutrition and Mulching. Acta Sci. Pol. Hortorum Cultus 2011, 10, 133–144. [Google Scholar]
- Kesik, T.; BáaĪewicz-WoĨniak, M.; Michowska, A.E. Influence of Mulching and Nitrogen Nutrition on Bear Garlic (Allium ursinum L.) Growth. Acta Sci. Pol. Hortorum Cultus 2011, 10, 133–144. [Google Scholar]
- Znamirowska, A.; Szajnar, K.; Rożek, P.; Kalicka, D.; Kuźniar, P.; Hanus, P.; Kotula, K.; Obirek, M.; Kluz, M. Effect of Addition of Wild Garlic (Allium ursinum) on the Quality of Kefirs from Sheep’s Milk. Acta Sci. Pol. Technol. Aliment. 2017, 16, 209–215. [Google Scholar] [CrossRef]
- Błażewicz-Woźniak, M.; Michowska, A. The Growth, Flowering and Chemical Composition of Leaves of Three Ecotypes of Allium ursinum L. Acta Agrobot. 2012, 64, 171–180. [Google Scholar] [CrossRef]
- Ballesteros-Canovas, J.A.; Mohan, D.; Stoffel, M.; Khan, M.A.; Kuriger, J.; Sinha, S. Science Brief: Ecosystem-Based Adaptation: An Integrated Response to Climate Change in the Indian Himalayan Region; IHCAP: Delhi, India, 2017. [Google Scholar]
- Salgotra, R.K.; Sharma, M.; Pandotra, P. Biotechnological Interventions for Sustainable Conservation of Plant Genetic Resources in the Scenario of Climate Change. Nat. Resour. Conserv. Res. 2019, 2, 754. [Google Scholar] [CrossRef]
- Amagova, Z.; Matsadze, V.; Kavarnakaeva, Z.; Golubkina, N.; Antoshkina, M.; Sękara, A.; Tallarita, A.; Caruso, G. Joint Cultivation of Allium ursinum and Armoracia rusticana under Foliar Sodium Selenate Supply. Plants 2022, 11, 2778. [Google Scholar] [CrossRef]
- Farkas, Á.; Molnár, R.; Morschhauser, T.; Hahn, I. Variation in Nectar Volume and Sugar Concentration of Allium ursinum L. ssp. ucrainicum in Three Habitats. Sci. World J. 2012, 2012, 138579. [Google Scholar] [CrossRef]
- Bodó, A.; Farkas, Á.; Nagy, D.U.; Rudolf, K.; Hoffmann, R.; Kocsis, M.; Morschhauser, T. Soil Humus, Iron, Sulphate and Magnesium Content Affect Nectar Traits of Wild Garlic (Allium ursinum L.). Plants 2021, 10, 597. [Google Scholar] [CrossRef]
- Kuebbing, S.E.; Nuñez, M.A.; Simberloff, D. Current Mismatch between Research and Conservation Efforts: The Need to Study Co-Occurring Invasive Plant Species. Biol. Conserv. 2013, 160, 121–129. [Google Scholar] [CrossRef]
- FNAEC Convallaria. Flora of North America; FNA Editorial Committee, Ed.; Oxford University Press: New York, NY, USA, 2003; p. 205. Available online: http://floranorthamerica.org/Main_Page (accessed on 19 September 2023).
- POWO Plants of the World Online. Facilitated by the Royal Botanic Gardens, Kew. Published on the Internet. Available online: http://www.Plantsoftheworldonline.Org/ (accessed on 19 June 2023).
- Tutin, T.G. Biological Flora of the British Isles: Allium ursinum L. J. Ecol. 1957, 45, 1003–1010. [Google Scholar] [CrossRef]
- Eggert, A. Dry Matter Economy and Reproduction of a Temperate Forest Spring Geophyte—Allium ursinum. Ecography 1992, 15, 45–55. [Google Scholar] [CrossRef]
- Stranczinger, S.; Rudolf, K.; Morschhauser, T.; Stranczinger, S.; Rudolf, K.; Farkas, Á.; Morschhauser, T.; Stranczinger, S.; Rudolf, K.; Farkas, Á. Characteristics of Reproductive Strategies in Wild Garlic (Allium ursinum L.). Int. J. Plant Reprod. Biol. 2014, 6, 21–29. [Google Scholar]
- Grime, J.P.; Hodgson, J.G.; Hunt, R. Comparative Plant Ecology. A Functional Approach to Common British Species, 1st ed.; Springer: London, UK, 1988; ISBN 9401710945. [Google Scholar]
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea. Vol. 1. Lycopodiaceae to Platanaceae.; Cambridge University Press: London, UK, 1964; Available online: https://www.cambridge.org/us/universitypress/subjects/life-sciences/botanical-reference/flora-europaea-volume-1-2nd-edition (accessed on 19 September 2023).
- Kovacs, J.A. Data to Vegetation Biology and Coenological Relations of Allium ursinum L. Stands in Eastern Transylvania. Kanitzia 2007, 15, 63–76. [Google Scholar]
- Sonkoly, J.; Tóth, E.; Balogh, N.; Balogh, L.; Bartha, D.; Csendesné Bata, K.; Bátori, Z.; Békefi, N.; Botta-Dukát, Z.; Bölöni, J.; et al. PADAPT 1.0—the Pannonian Dataset of Plant Traits. Sci. Data 2023, 10, 742. [Google Scholar] [CrossRef] [PubMed]
- Borhidi, A. Social Behaviour Types of the Hungarian Flora, Its Naturalness and Relative Ecological Indicator Values. Janus Pannon. Tudom. Kiadv. Pecs. Hung. 1993, 39, 97–181. [Google Scholar]
- Soó, R. Synopsis of the Systematic-Geobotanical Flora and Vegetation of Hungary VI (Latin: Synonpsis Systematico-Geobotanica Florae Vegetatonisque Hungarae); Akademiai Kiado: Budapest, Hungary, 1980; ISBN 9630520915. [Google Scholar]
- Kosiński, I. The Influence of Shoot Harvesting on the Age Structure of Convallaria majalis L. Populations. Acta Soc. Bot. Pol. 2003, 72, 53–59. [Google Scholar] [CrossRef]
- Eriksson, O. Seed Size Variation and Its Effect on Germination and Seedling Performance in the Clonal Herb Convallaria majalis. Acta Oecol. 1999, 20, 61–66. [Google Scholar] [CrossRef]
- Oinonen, E. The Time Table of Vegetative Spreading of the Lily-of-the-Valley (Convallaria majalis L.) and the Wood Small-Reed (Calamagrostis epigeios (L.) Roth) in Southern Finland. Acta For. Fenn. 1969, 97, 7607. [Google Scholar] [CrossRef][Green Version]
- Braune, S.; Lieske, K.; Frey, W.; Pfeiffer, T. Vegetative Multiplication and Spatial Genetic Structure in Patches of Convallaria majalis L. (Ruscaceae). Plant Divers. Evol. 2011, 129, 7–26. [Google Scholar] [CrossRef]
- Zlatník, A. Forest Phytocenology (In Czech: Lesnická Fytocenologie); Státní Zemědělské Nakladatelství: Praha, Czech Republic, 1976. [Google Scholar]
- Dostál, J. New Flora of the C.S.S.R.; Academia ČSAV: Praha, Czech Republic, 1989. (In Czech) [Google Scholar]
- Rejzková, E.; Fér, T.; Vojta, J.; Marhold, K. Phylogeography of the Forest Herb Carex pilosa (Cyperaceae). Bot. J. Linn. Soc. 2008, 158, 115–130. [Google Scholar] [CrossRef]
- Iliško, E.; Soms, J. Geographic Distribution of Protected Sedge Species Carex Pilosa Scop. In Latvia with Reference to Forest Ecosystems. In ENVIRONMENT. TECHNOLOGIES. RESOURCES. Proceedings of the International Scientific and Practical Conference; 2015; Volume 2, p. 325. Available online: http://journals.ru.lv/index.php/ETR/article/view/975 (accessed on 19 September 2023). [CrossRef]
- Ellenberg, H.; Weber, H.E.; Dull, R.; Wirth, V.; Werner, W.; Paulißen, D. Indicator Values of Plants in Central Europe (In German: Zeigerwerte von Pflanzen in Mitteleuropa 2nd Ed). Scr. Geobot. 1992, 18, 1–260. [Google Scholar]
- Oberdorfer, E. Southern German Plant Societies, Part IV, Forests and Shrubs (German: Süddeutsche Pflanzengesellschaften, Teil IV, Wälder und Gebüsche), 2nd ed.; Gustav Fischér Verlag: New York, NY, USA, 1992. [Google Scholar]
- Chytry, M.; Vicherek, J. Forest Vegetation of Podyjí/Thayatal National Park (Czech/German: Lesní Vegetace Národního Parku Podyjí/Thayatal. Die Waldvegetation Des Nationalparks Podyjí/Thayatal); Academia: Praha, Czech Republic, 1995. [Google Scholar]
- Bohn, U.; Gollub, G.; Hettwer, C.; Neuhäuslová, Z.; Raus, T.; Schlüter, H.; Weber, H. Map of the Natural Vegetation of Europe. Scale 1:2,500,000 (German: Karte Der Natürlichen Vegetation Europas, Maßstab 1:2,500,000); Bonn-Bad Godesberg: Bundesamt für Naturschutz, Germany, 2003; Available online: https://library.wur.nl/WebQuery/titel/1631571 (accessed on 19 September 2023).
- Zubek, S.; Rola, K.; Rożek, K.; Błaszkowski, J.; Stanek, M.; Chmolowska, D.; Chowaniec, K.; Zalewska-Gałosz, J.; Stefanowicz, A.M. Experimental Assessment of Forest Floor Geophyte and Hemicryptophyte Impact on Arbuscular Mycorrhizal Fungi Communities. Plant Soil 2022, 480, 651–673. [Google Scholar] [CrossRef]
- R Core Team. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Djurdjevic, L.; Dinic, A.; Pavlovic, P.; Mitrovic, M.; Karadzic, B.; Tesevic, V. Allelopathic Potential of Allium ursinum L. Biochem. Syst. Ecol. 2004, 32, 533–544. [Google Scholar] [CrossRef]
- Janík, R.; Schieber, B. Effect of Stocking Densities on the Biomass Production of Carex pilosa L. in Sub-Mountain Beech Stands. Beskydy 2010, 3, 151–158. [Google Scholar]
- Campillo, C.; Fortes, R.; Henar Prieto, M. del Solar radiation effect on crop production. In Solar Radiation; InTech: Rijeka, Croatia, 2012. [Google Scholar]
- Muchow, R.C.; Sinclair, T.R.; Bennett, J.M. Temperature and Solar Radiation Effects on Potential Maize Yield across Locations. Agron. J. 1990, 82, 338–343. [Google Scholar] [CrossRef]
- Střelec, J. Influence of Cutting Operation in a Beech Stand on Changes in Illumination (Slovak: Vplyv Ťažbového Zásahu v Bu-Kovom Poraste Na Zmeny Osvetlenia). Lesn. Časopis–Forestry J. 1992, 38, 551–558. [Google Scholar]
- Mayr, R.; Godoy, R. Seasonal Patterns in Vesicular-Arbuscular Mycorrhiza in Melic-Beech Forest. Agric. Ecosyst. Environ. 1990, 29, 281–288. [Google Scholar] [CrossRef]
- Stefanowicz, A.M.; Kapusta, P.; Stanek, M.; Rola, K.; Zubek, S. Herbaceous Plant Species Support Soil Microbial Performance in Deciduous Temperate Forests. Sci. Total Environ. 2022, 810, 151313. [Google Scholar] [CrossRef]
- Neuenkamp, L.; Zobel, M.; Koorem, K.; Jairus, T.; Davison, J.; Öpik, M.; Vasar, M.; Moora, M. Light Availability and Light Demand of Plants Shape the Arbuscular Mycorrhizal Fungal Communities in Their Roots. Ecol. Lett. 2021, 24, 426–437. [Google Scholar] [CrossRef]
- Olsson, M.O.; Falkengren-Grerup, U. Partitioning of Nitrate Uptake between Trees and Understory in Oak Forests. For. Ecol. Manag. 2003, 179, 311–320. [Google Scholar] [CrossRef]
- Muller, R. Nutrient relations of the herbaceous layer in deciduous forest ecosystems; In Gilliam & Roberts. In The Herbaceous Layer in Forests of Eastern North America; Life Sciences; Oxford University Press: New York, NY, USA, 2003; pp. 15–37. ISBN 9780195140880. Available online: https://global.oup.com/academic/product/the-herbaceous-layer-in-forests-of-eastern-north-america-9780199837656?q=The%20Herbaceous%20Layer%20in%20Forests%20of%20Eastern%20North%20America&lang=en&cc=us (accessed on 19 September 2023).
- Gilliam, F.S. The Ecological Significance of the Herbaceous Layer in Temperate Forest Ecosystems. Bioscience 2007, 57, 845–858. [Google Scholar] [CrossRef]
- Rothstein, D.E. Spring Ephemeral Herbs and Nitrogen Cycling in a Northern Hardwood Forest: An Experimental Test of the Vernal Dam Hypothesis. Oecologia 2000, 124, 446–453. [Google Scholar] [CrossRef]
- Grime, J.P. Plant Strategies, Vegetation Processes, and Ecosystem Properties; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2006; ISBN 0471496014. Available online: https://www.wiley.com/en-ie/Plant+Strategies%2C+Vegetation+Processes%2C+and+Ecosystem+Properties%2C+2nd+Edition-p-9780470850404 (accessed on 19 September 2023).
- Tutin, T.G.; Heywood, V.H.; Burges, N.A.; Valentine, D.H.; Walters, S.M.; Webb, D.A. Flora Europaea: Plantaginaceae to Compositae (and Rubiaceae); Cambridge University Press: Cambridge, UK, 1976; Volume 4, ISBN 0521087171. [Google Scholar]
- Kevey, B. Phytogeographic Characterization of Allium ursinum with Particular Interest in Its Occurrence in Hungary. Ph.D. Thesis, Lajos Kossuth University of Debrecen, Debrecen, Hungary, 1977. Available online: https://doktori.hu/index.php?menuid=120&lang=EN (accessed on 19 September 2023). (In Hungarian).
- Grime, J.P.; Crick, J.C.; Rincon, J.E. The Ecological Significance of Plasticity. Symp. Soc. Exp. Biol. 1986, 40, 5–29. [Google Scholar]
- Kuklová, M.; Kukla, J.; Schieber, B. Individual and Population Parameters of Carex Pilosa Scop. (Cyperaceae) in Four Forest Sites in Western Carpathians (Slovakia). Pol. J. Ecol. 2005, 53, 427–434. [Google Scholar]
- Gudžinskas, Z.; Krampis, I.; Laiviņš, M. Spread of Carex Pilosa Scop. in Latvia and Lithuania. Latv. Veģetācija 2014, 21, 127–132. [Google Scholar]
- Gribova, S.; Isachenko, T.; Lavrenko, E.M. The Vegetation of the European Part of the USSR (In Russian: Rastitelnost Evropeiskoi Chasti SSSR); Nauka Publisher: Leningrad, Russia, 1980; Available online: https://www.naukapublishers.ru/ (accessed on 19 September 2023).
- Luo, W.; Zhao, W. Burial Depth and Diameter of the Rhizome Fragments Affect the Regenerative Capacity of a Clonal Shrub. Ecol. Complex. 2015, 23, 34–40. [Google Scholar] [CrossRef]
- Janík, R. Light Conditions in Submountain Beech Stands in Dependence on Cutting Intensity. Folia Oecol. 2009, 36, 67–71. [Google Scholar]
- Lubbe, F.C.; Henry, H.A.L. Plant Rhizome Positioning in the Soil and under Litter: Trade-offs of Frost Avoidance versus Growth. Perspect. Plant Ecol. Evol. Syst. 2019, 41, 125500. [Google Scholar] [CrossRef]
- Bazzaz, F.A.; Ackerly, D.D.; Reekie, E.G. Reproductive allocation in plants. In Seeds: The Ecology of Regeneration in Plant Communities; CABI Publishing: Wallingford, UK, 2000; pp. 1–29. [Google Scholar]
- Zhang, C.; Yang, C.; Dong, M. The Significance of Rhizome Connection of Semi-Shrub Hedysarum Laeve in an Inner Mongolian Dune, China. Acta Oecol. 2002, 23, 109–114. [Google Scholar] [CrossRef]
- Conti, L.; Block, S.; Parepa, M.; Münkemüller, T.; Thuiller, W.; Acosta, A.T.R.; van Kleunen, M.; Dullinger, S.; Essl, F.; Dullinger, I.; et al. Functional Trait Differences and Trait Plasticity Mediate Biotic Resistance to Potential Plant Invaders. J. Ecol. 2018, 106, 1607–1620. [Google Scholar] [CrossRef]
- Westbrooks, R.G.; Eplee, R.E. Early Detection and Rapid Response. In Encyclopedia of Biological Invasions; Simberloff, D., Rejmánek, M., Eds.; University of California Press: Berkeley, CA, USA, 2019; pp. 169–177. Available online: https://pubs.usgs.gov/publication/70156706 (accessed on 19 September 2023).
- Baskin, C.C.; Baskin, J.M. Germination Ecology of Seeds with Physical Dormancy. In Seeds; Academic Press: San Diego, CA, USA, 2014. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Soil Type | p-Value |
|---|---|
| HUN vs. USA | 0.2362 |
| AU vs. USA | <0.01 |
| AU vs. CP + MU | <0.0001 |
| USA vs. CP + MU | <0.001 |
| Df | Sum Sq. | Mean Sq. | F Value | Pr (>F) | |
|---|---|---|---|---|---|
| Solar | 1 | 2.415 × 1012 | 2.415 × 1012 | 119.365 | <0.001 |
| Soil Type | 9 | 7.684 × 1012 | 8.538 × 1011 | 42.206 | <0.001 |
| Solar: Soil Type | 9 | 3.246 × 1012 | 3.607 × 1011 | 17.831 | <0.001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, K.; Pal, R.W.; Somfalvi-Tóth, K.; Riyazuddin, R.; Rudolf, K.; Morschhauser, T. What Do Cross-Range Germination, Growth, and Interaction Studies Reveal about the Behaviour of an Expansive Plant Species? Agriculture 2023, 13, 2171. https://doi.org/10.3390/agriculture13112171
Kaushik K, Pal RW, Somfalvi-Tóth K, Riyazuddin R, Rudolf K, Morschhauser T. What Do Cross-Range Germination, Growth, and Interaction Studies Reveal about the Behaviour of an Expansive Plant Species? Agriculture. 2023; 13(11):2171. https://doi.org/10.3390/agriculture13112171
Chicago/Turabian StyleKaushik, Krishan, Robert W. Pal, Katalin Somfalvi-Tóth, Riyazuddin Riyazuddin, Kinga Rudolf, and Tamás Morschhauser. 2023. "What Do Cross-Range Germination, Growth, and Interaction Studies Reveal about the Behaviour of an Expansive Plant Species?" Agriculture 13, no. 11: 2171. https://doi.org/10.3390/agriculture13112171
APA StyleKaushik, K., Pal, R. W., Somfalvi-Tóth, K., Riyazuddin, R., Rudolf, K., & Morschhauser, T. (2023). What Do Cross-Range Germination, Growth, and Interaction Studies Reveal about the Behaviour of an Expansive Plant Species? Agriculture, 13(11), 2171. https://doi.org/10.3390/agriculture13112171