
Effects of Tebuconazole on the Earthworm Dendrobaena veneta: Full Life Cycle Approach


Abstract
1. Introduction
2. Materials and Methods
2.1. Test Organisms
2.2. Test Materials and Soil Spiking
2.3. Experimental Design
2.3.1. Cocoon Hatching Success and Juvenile Growth Dynamics (Cocoon Exposure)
2.3.2. Reproduction Test
2.4. Data Presentation and Statistical Analysis
3. Results
3.1. Cocoon Hatching Dynamics, Juvenile Mass Hatching, Juvenile Growth Dynamics
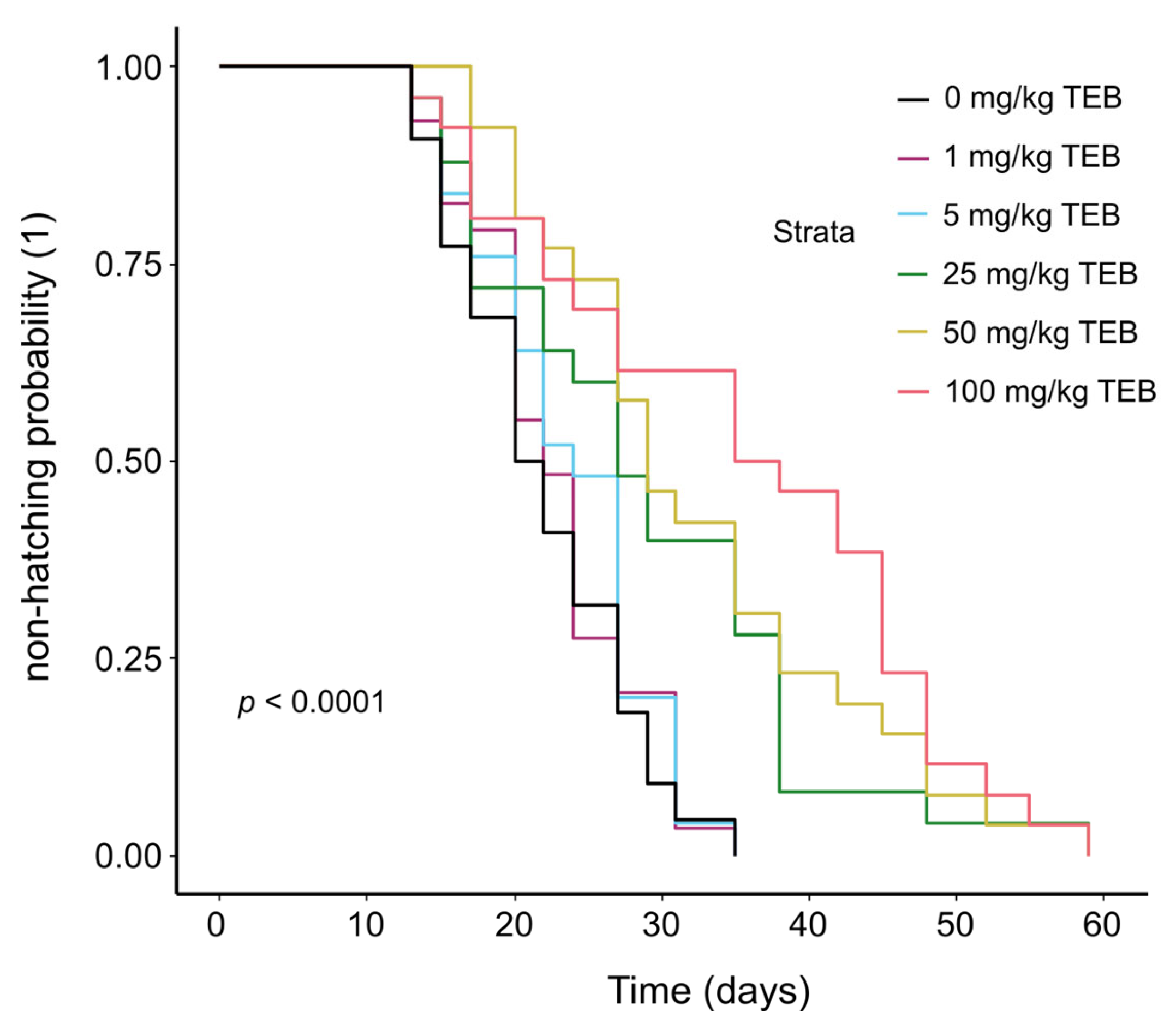
3.1.1. Cocoon Hatching Dynamics
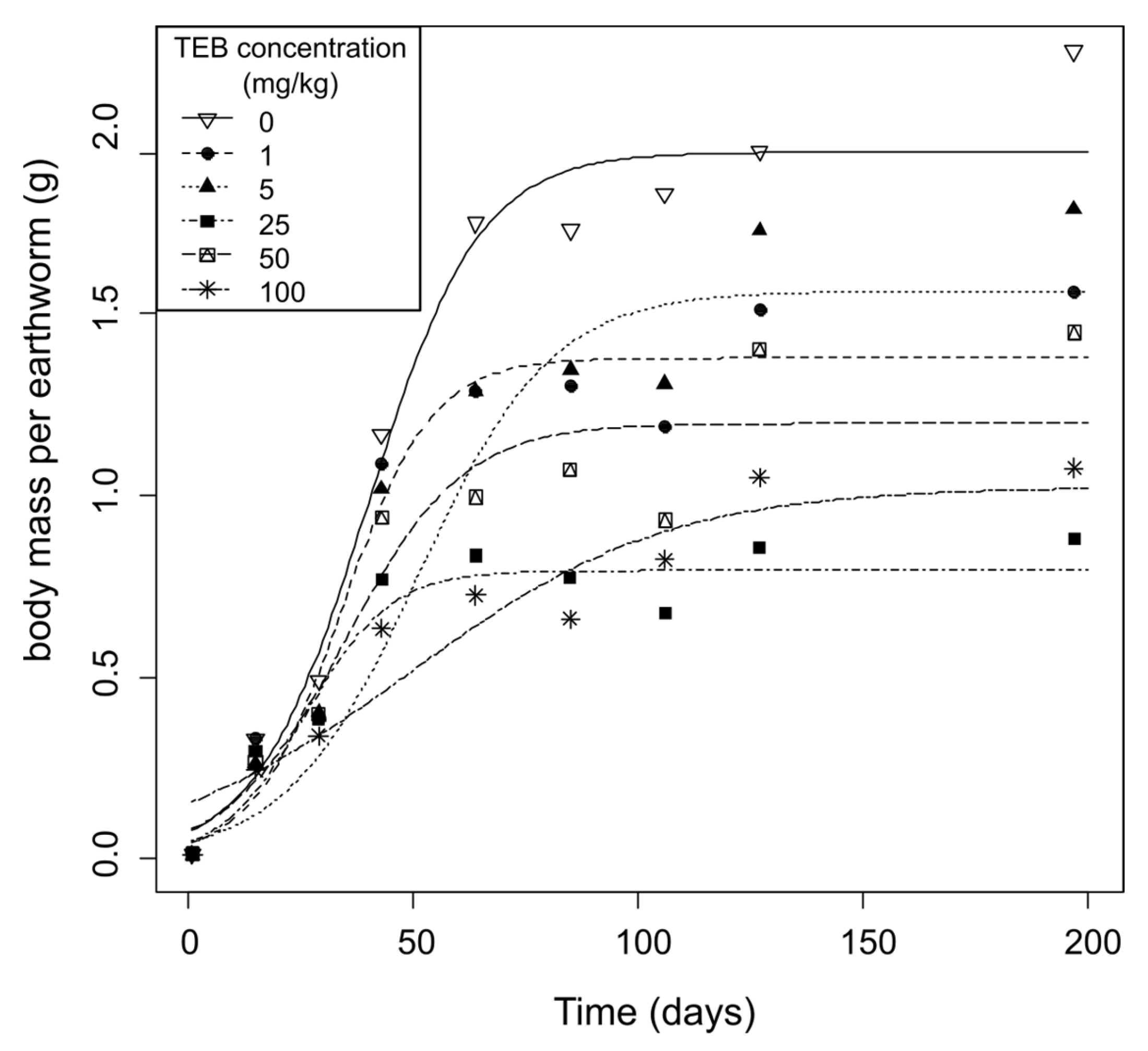
3.1.2. Juvenile Mass at Hatching and Growth Dynamics
3.2. Reproduction Test
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- World Food and Agriculture–Statistical Yearbook 2021; FAO: Rome, Italy, 2021.
- Facts and Figures about Toxic Chemicals in Agriculture 2022, 2nd ed.; Heinrich-Böll-Stiftung: Berlin, Germany, 2022; ISBN 9789464007473.
- Van Dijk, M.; Morley, T.; Rau, M.L.; Saghai, Y. A Meta-Analysis of Projected Global Food Demand and Population at Risk of Hunger for the Period 2010–2050. Nat. Food 2021, 2, 494–501. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Wang, Z.; Zhang, B.; Zhang, Q. Degradation and Adsorption of Tebuconazole and Tribenuron-Methyl in Wheat Soil, Alone and in Combination. Chil. J. Agric. Res. 2017, 77, 281–286. [Google Scholar] [CrossRef][Green Version]
- Varelis, P. Pesticide Residues: Conazoles. Ref. Modul. Food Sci. 2024, 476–479. [Google Scholar] [CrossRef]
- Silva, V.; Mol, H.G.J.; Zomer, P.; Tienstra, M.; Ritsema, C.J.; Geissen, V. Pesticide Residues in European Agricultural Soils—A Hidden Reality Unfolded. Sci. Total Environ. 2019, 653, 1532–1545. [Google Scholar] [CrossRef] [PubMed]
- Hvězdová, M.; Kosubová, P.; Košíková, M.; Scherr, K.E.; Šimek, Z.; Brodský, L.; Šudoma, M.; Škulcová, L.; Sáňka, M.; Svobodová, M.; et al. Currently and Recently Used Pesticides in Central European Arable Soils. Sci. Total Environ. 2018, 613–614, 361–370. [Google Scholar] [CrossRef] [PubMed]
- Bošković, N.; Brandstätter-Scherr, K.; Sedláček, P.; Bílková, Z.; Bielská, L.; Hofman, J. Adsorption of Epoxiconazole and Tebuconazole in Twenty Different Agricultural Soils in Relation to Their Properties. Chemosphere 2020, 261, 127637. [Google Scholar] [CrossRef] [PubMed]
- Lewis, K.A.; Tzilivakis, J.; Warner, D.J.; Green, A. An International Database for Pesticide Risk Assessments and Management. Hum. Ecol. Risk Assess. 2016, 22, 1050–1064. [Google Scholar] [CrossRef]
- Xu, C.; Sun, X.; Jin, M.; Yang, X.; Zhang, L.; Yao, Y.; Niu, L.; Shentu, J.; Liu, J.; Liu, W. Maternal Exposure to Chiral Triazole Fungicide Tebuconazole Induces Enantioselective Thyroid Disruption in Zebrafish Offspring. Ecotoxicol. Environ. Saf. 2023, 251, 114540. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, C.; Yang, G.; Wang, X.; Wang, Q.; Weng, H.; Zhang, Z.; Qian, Y. Combined Lethal Toxicity, Biochemical Responses, and Gene Expression Variations Induced by Tebuconazole, Bifenthrin and Their Mixture in Zebrafish (Danio rerio). Ecotoxicol. Environ. Saf. 2022, 230, 113116. [Google Scholar] [CrossRef]
- Sancho, E.; Villarroel, M.J.; Ferrando, M.D. Assessment of Chronic Effects of Tebuconazole on Survival, Reproduction and Growth of Daphnia magna after Different Exposure Times. Ecotoxicol. Environ. Saf. 2016, 124, 10–17. [Google Scholar] [CrossRef]
- Sancho, E.; Villarroel, M.J.; Andreu, E.; Ferrando, M.D. Disturbances in Energy Metabolism of Daphnia magna after Exposure to Tebuconazole. Chemosphere 2009, 74, 1171–1178. [Google Scholar] [CrossRef] [PubMed]
- Rico, A.; Sabater, C.; Castillo, M.Á. Lethal and Sub-Lethal Effects of Five Pesticides Used in Rice Farming on the Earthworm Eisenia fetida. Ecotoxicol. Environ. Saf. 2016, 127, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Li, G.; Li, D.; Rao, H.; Liu, X. Potential Neurotoxicity, Immunotoxicity, and Carcinogenicity Induced by Metribuzin and Tebuconazole Exposure in Earthworms (Eisenia fetida) Revealed by Transcriptome Analysis. Sci. Total Environ. 2022, 807, 150760. [Google Scholar] [CrossRef] [PubMed]
- Yu, D.; Li, J.; Zhang, Y.; Wang, H.; Guo, B.; Zheng, L. Enantioseletive Bioaccumulation of Tebuconazole in Earthworm Eisenia fetida. J. Environ. Sci. 2012, 24, 2198–2204. [Google Scholar] [CrossRef]
- Zhang, R.; Zhou, Z.; Zhu, W. Evaluating the Effects of the Tebuconazole on the Earthworm, Eisenia fetida by H-1 NMR-Based Untargeted Metabolomics and MRNA Assay. Ecotoxicol. Environ. Saf. 2020, 194, 110370. [Google Scholar] [CrossRef] [PubMed]
- Gomes, S.I.L.; Ammendola, A.; Casini, S.; Amorim, M.J.B. Toxicity of Fungicides to Terrestrial Non-Target Fauna–Formulated Products versus Active Ingredients (Azoxystrobin, Cyproconazole, Prothioconazole, Tebuconazole)—A Case Study with Enchytraeus crypticus (Oligochaeta). Sci. Total Environ. 2021, 754, 142098. [Google Scholar] [CrossRef] [PubMed]
- Römbke, J.; Jänsch, S.; Didden, W. The Use of Earthworms in Ecological Soil Classification and Assessment Concepts. Ecotoxicol. Environ. Saf. 2005, 62, 249–265. [Google Scholar] [CrossRef]
- Pelosi, C.; Barot, S.; Capowiez, Y.; Hedde, M.; Vandenbulcke, F. Pesticides and Earthworms. A Review. Agron. Sustain. Dev. 2014, 34, 199–228. [Google Scholar] [CrossRef]
- Reinecke, S.A.; Reinecke, A.J. The Impact of Organophosphate Pesticides in Orchards on Earthworms in the Western Cape, South Africa. Ecotoxicol. Environ. Saf. 2007, 66, 244–251. [Google Scholar] [CrossRef]
- Bicho, R.C.; Santos, F.C.F.; Scott-Fordsmand, J.J.; Amorim, M.J.B. Effects of Copper Oxide Nanomaterials (CuONMs) Are Life Stage Dependent–Full Life Cycle in Enchytraeus crypticus. Environ. Pollut. 2017, 224, 117–124. [Google Scholar] [CrossRef]
- Bicho, R.C.; Santos, F.C.F.; Gonçalves, M.F.M.; Soares, A.M.V.M.; Amorim, M.J.B. Enchytraeid Reproduction TestPLUS: Hatching, Growth and Full Life Cycle Test—An Optional Multi-Endpoint Test with Enchytraeus crypticus. Ecotoxicology 2015, 24, 1053–1063. [Google Scholar] [CrossRef] [PubMed]
- Podolak, A.; Kostecka, J.; Mazur-Paczka, A.; Garczyńska, M.; Paczka, G.; Szura, R. Life Cycle of the Eisenia fetida and Dendrobaena veneta Earthworms (Oligohaeta, Lumbricidae). J. Ecol. Eng. 2020, 21, 40–45. [Google Scholar] [CrossRef] [PubMed]
- Hackenberger, D.K.; Stjepanović, N.; Lončarić, Ž.; Hackenberger, B.K. Acute and Subchronic Effects of Three Herbicides on Biomarkers and Reproduction in Earthworm Dendrobaena veneta. Chemosphere 2018, 208, 722–730. [Google Scholar] [CrossRef]
- OECD. Guidelines for the Testing of Chemicals (222/2016) Earthworms Reproduction Test (Eisenia fetida/Eisenia andrei); Organization for Economic Cooperation and Development: Paris, France, 2016; pp. 1–21. [Google Scholar]
- Viljoen, S.A.; Reinecke, A.J.; Hartman, L. Life-Cycle of the European Compost Worm Dendrobaena veneta (Oligochaeta). Afr. Zool. 1991, 26, 43–48. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Core Team: Vienna, Austria, 2022; Available online: https://www.R-project.org/ (accessed on 5 August 2023).
- RStudio Team. RStudio: Integrated Development for R; T.M. Survival: Survival Analysis; RStudio, PBC: Boston, MA, USA, 2020; Available online: http://www.rstudio.com/Therneau (accessed on 5 August 2023).
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.5-5; R Core Team: Vienna, Austria, 2023; Available online: https://CRAN.R-project.org/package=survival (accessed on 5 August 2023).
- Kassambara, A.; Kosinski, M.; Biecek, P. Survminer: Drawing Survival Curves Using Ggplot2; R Core Team: Vienna, Austria, 2021. [Google Scholar]
- Sprouffske, K. Growthcurver: Simple Metrics to Summarize Growth Curves; R Core Team: Vienna, Austria, 2020. [Google Scholar]
- Castro-Ferreira, M.P.; Roelofs, D.; van Gestel, C.A.M.; Verweij, R.A.; Soares, A.M.V.M.; Amorim, M.J.B. Enchytraeus crypticus as Model Species in Soil Ecotoxicology. Chemosphere 2012, 87, 1222–1227. [Google Scholar] [CrossRef] [PubMed]
- Kovačević, M.; Hackenberger, D.K.; Hackenberger, B.K. Effects of Strobilurin Fungicides (Azoxystrobin, Pyraclostrobin, and Trifloxystrobin) on Survival, Reproduction and Hatching Success of Enchytraeus crypticus. Sci. Total Environ. 2021, 790, 148143. [Google Scholar] [CrossRef] [PubMed]
- Bart, S.; Barraud, A.; Amossé, J.; Péry, A.R.R.; Mougin, C.; Pelosi, C. Effects of Two Common Fungicides on the Reproduction of Aporrectodea caliginosa in Natural Soil. Ecotoxicol. Environ. Saf. 2019, 181, 518–524. [Google Scholar] [CrossRef]
- Chen, J.; Saleem, M.; Wang, C.; Liang, W.; Zhang, Q. Individual and Combined Effects of Herbicide Tribenuron-Methyl and Fungicide Tebuconazole on Soil Earthworm Eisenia fetida. Sci. Rep. 2018, 8, 2967. [Google Scholar] [CrossRef]
- Johnston, A.S.A.; Hodson, M.E.; Thorbek, P.; Alvarez, T.; Sibly, R.M. An Energy Budget Agent-Based Model of Earthworm Populations and Its Application to Study the Effects of Pesticides. Ecol. Model. 2014, 280, 5–17. [Google Scholar] [CrossRef]
- Velki, M.; Hackenberger, B.K. Different Sensitivities of Biomarker Responses in Two Epigeic Earthworm Species after Exposure to Pyrethroid and Organophosphate Insecticides. Arch. Environ. Contam. Toxicol. 2013, 65, 498–509. [Google Scholar] [CrossRef]
- Xiao, N.; Jing, B.; Ge, F.; Liu, X. The Fate of Herbicide Acetochlor and Its Toxicity to Eisenia fetida under Laboratory Conditions. Chemosphere 2006, 62, 1366–1373. [Google Scholar] [CrossRef]
- Mosleh, Y.Y.; Ismail, S.M.M.; Ahmed, M.T.; Ahmed, Y.M. Comparative Toxicity and Biochemical Responses of Certain Pesticides to the Mature Earthworm Aporrectodea caliginosa under Laboratory Conditions. Environ. Toxicol. 2003, 18, 338–346. [Google Scholar] [CrossRef] [PubMed]
- Mosleh, Y.Y.; Paris-Palacios, S.; Couderchet, M.; Vernet, G. Acute and Sublethal Effects of Two Insecticides on Earthworms (Lumbricus terrestris L.) under Laboratory Conditions. Environ. Toxicol. 2003, 18, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Zhou, S.; Duan, C.; Wang, X.; Michelle, W.H.G.; Yu, Z.; Fu, H. Assessing Cypermethrin-Contaminated Soil with Three Different Earthworm Test Methods. J. Environ. Sci. 2008, 20, 1381–1385. [Google Scholar] [CrossRef] [PubMed]
- Booth, L.H.; O’Halloran, K. A Comparison of Biomarker Responses in the Earthworm Aporrectodea caliginosa to the Organophosphorus Insecticides Diazinon and Chlorpyrifos. Environ. Toxicol. Chem. 2001, 20, 2494–2502. [Google Scholar] [CrossRef] [PubMed]
- Briones, M.J.I.; Álvarez-Otero, R. Body Wall Thickness as a Potential Functional Trait for Assigning Earthworm Species to Ecological Categories. Pedobiologia 2018, 67, 26–34. [Google Scholar] [CrossRef]
- Suthar, S. Toxicity of Methyl Parathion on Growth and Reproduction of Three Ecologically Different Tropical Earthworms. Int. J. Environ. Sci. Technol. 2014, 11, 191–198. [Google Scholar] [CrossRef]
- Santadino, M.; Coviella, C.; Momo, F. Glyphosate Sublethal Effects on the Population Dynamics of the Earthworm Eisenia fetida (Savigny, 1826). Water Air Soil Pollut. 2014, 225, 1–8. [Google Scholar] [CrossRef]
- Costa, P.A.; Guilhermino, L.; Laskowski, R. Effects of Pesticide Formulations Containing Cypermethrin or Tebuconazole, Individually and in Mixture, on the Earthworm Eisenia fetida. Appl. Soil Ecol. 2024, 193, 105139. [Google Scholar] [CrossRef]
- Beaumelle, L.; Tison, L.; Eisenhauer, N.; Hines, J.; Malladi, S.; Pelosi, C.; Thouvenot, L.; Phillips, H.R.P. Pesticide Effects on Soil Fauna Communities—A Meta-Analysis. J. Appl. Ecol. 2023, 60, 1239–1253. [Google Scholar] [CrossRef]
- Kaka, H.; Opute, P.A.; Maboeta, M.S. Potential Impacts of Climate Change on the Toxicity of Pesticides towards Earthworms. J. Toxicol. 2021, 2021, 8527991. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Treatment (mg/kg TEB) | Median Hatching Time | Average Hatching Success (%) | Average Number of Juveniles per Cocoon | Juvenile Survival Probability (%) |
|---|---|---|---|---|
| 0 | 21 [20, 27] | 73.3 ± 27.3 | 1.03 ± 0.08 | 0.95 ± 0.04 |
| 1 | 22 [20, 24] | 86.6 ± 16.32 | 1.03 ± 0.08 | 0.70 ± 0.16 |
| 5 | 24 [20, 27] | 83.3 ± 23.4 | 1.13 ± 0.33 | 0.77 ± 0.06 |
| 25 | 27 [22, 38] * | 76.7 ± 26.6 | 1.07 ± 0.16 | 0.63 ± 0.15 |
| 50 | 29 [27, 38] * | 70.0 ± 20.9 | 1.03 ± 0.08 | 0.40 ± 0.10 • |
| 100 | 36.5 [27, 45] * | 83.3 ± 23.4 | 1.10 ± 0.11 | 0.33 ± 0.10 • |
| Model Parameters | 0 mg/kg TEB | 1 mg/kg TEB | 5 mg/kg TEB | 25 mg/kg TEB | 50 mg/kg TEB | 100 mg/kg TEB |
|---|---|---|---|---|---|---|
| r | 0.081 | 0.101 | 0.068 | 0.106 | 0.077 | 0.035 * |
| K | 1.943 | 1.377 | 1.561 | 0.793 * | 1.197 * | 1.024 * |
| σ | 0.154 | 0.154 | 0.183 | 0.092 | 0.195 | 0.126 |
| Treatment (mg/kg TEB) | Adult Survival | Number of Juveniles | Mass per Juvenile (mg) | Number of Juveniles per Earthworm | Number of Unhatched Cocoons |
|---|---|---|---|---|---|
| 0 | 0.94 ± 0.09 | 22.1 ± 3.3 | 30.9 ± 2.8 | 5.2 ± 1.6 | 4.3 ± 0.95 |
| 1 | 0.97 ± 0.08 | 20.2 ± 2.9 | 25.1 ± 9.0 | 4.2 ± 0.6 | 2.1 ± 1.3 * |
| 5 | 0.91 ± 0.11 | 20.3 ± 5.0 | 27.5 ± 5.1 | 4.4 ± 0.8 | 2.7 ± 1.1 |
| 25 | 0.83 ± 0.14 | 15.7 ± 4.9 * | 27.0 ± 2.0 | 4.9 ± 0.9 | 4.9 ± 1.2 |
| 50 | 0.91 ± 0.11 | 15.4 ± 4.2 * | 21.8 ± 5.8 * | 3.7 ± 0.6 * | 4.6 ± 1.1 |
| 100 | 0.83 ± 0.18 | 11.6 ± 1.3 * | 15.1 ± 2.2 * | 2.9 ± 0.77 * | 4.1 ± 1.2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kovačević, M.; Stjepanović, N.; Zelić, L.; Lončarić, Ž. Effects of Tebuconazole on the Earthworm Dendrobaena veneta: Full Life Cycle Approach. Agriculture 2023, 13, 2119. https://doi.org/10.3390/agriculture13112119
Kovačević M, Stjepanović N, Zelić L, Lončarić Ž. Effects of Tebuconazole on the Earthworm Dendrobaena veneta: Full Life Cycle Approach. Agriculture. 2023; 13(11):2119. https://doi.org/10.3390/agriculture13112119
Chicago/Turabian StyleKovačević, Marija, Nikolina Stjepanović, Luca Zelić, and Željka Lončarić. 2023. "Effects of Tebuconazole on the Earthworm Dendrobaena veneta: Full Life Cycle Approach" Agriculture 13, no. 11: 2119. https://doi.org/10.3390/agriculture13112119
APA StyleKovačević, M., Stjepanović, N., Zelić, L., & Lončarić, Ž. (2023). Effects of Tebuconazole on the Earthworm Dendrobaena veneta: Full Life Cycle Approach. Agriculture, 13(11), 2119. https://doi.org/10.3390/agriculture13112119