
The Effect of Botanicals with Nematicidal Activity on the Structural and Functional Characteristics of the Soil Nematode Community

 ,
,  ,
,  , and
, and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Nematode Inoculum
2.2. Experimental Set Up and Sampling
2.3. Phospholipid Fatty Acid Analysis
2.4. Nematode Extraction and Analysis
2.5. Data Analysis
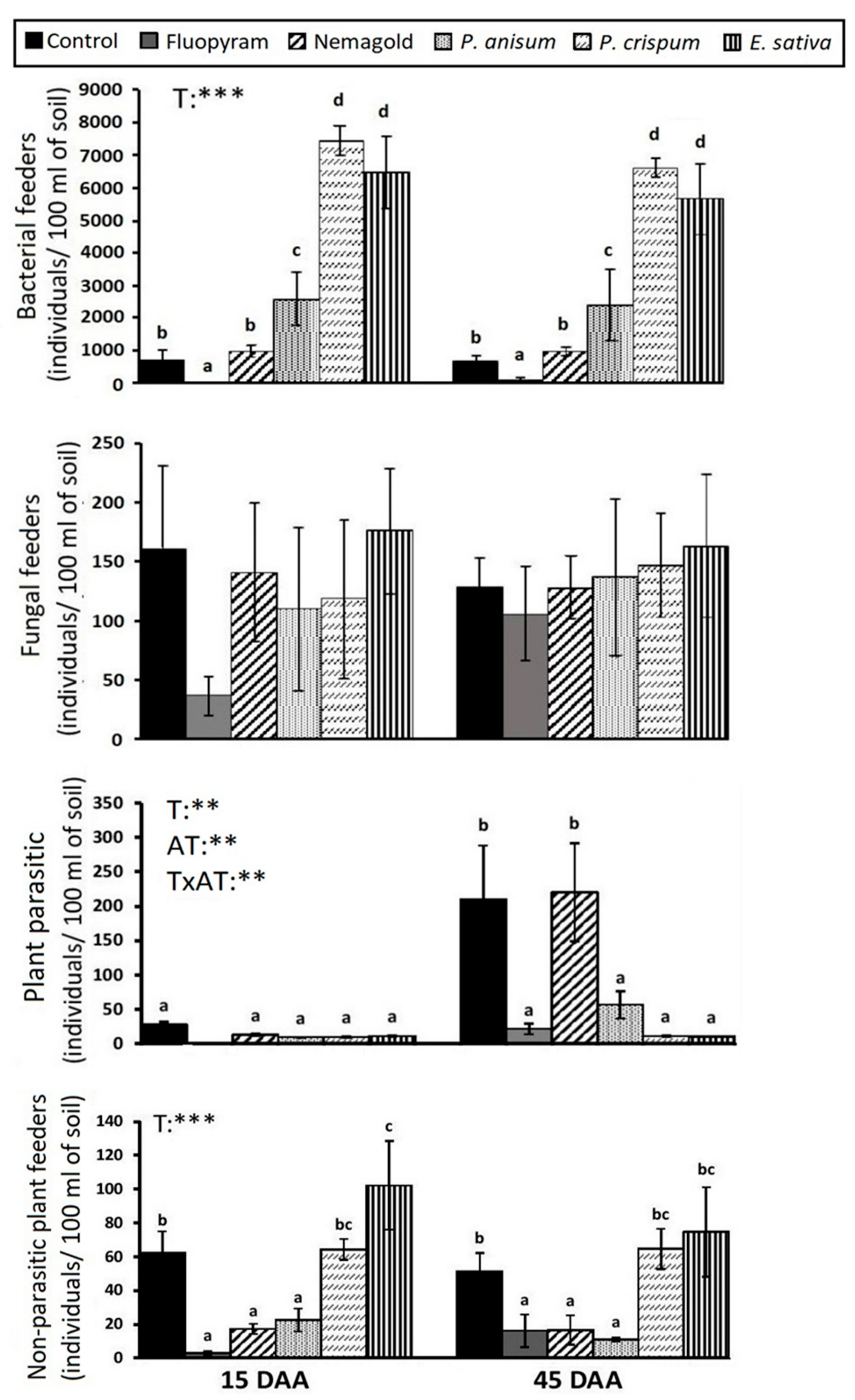
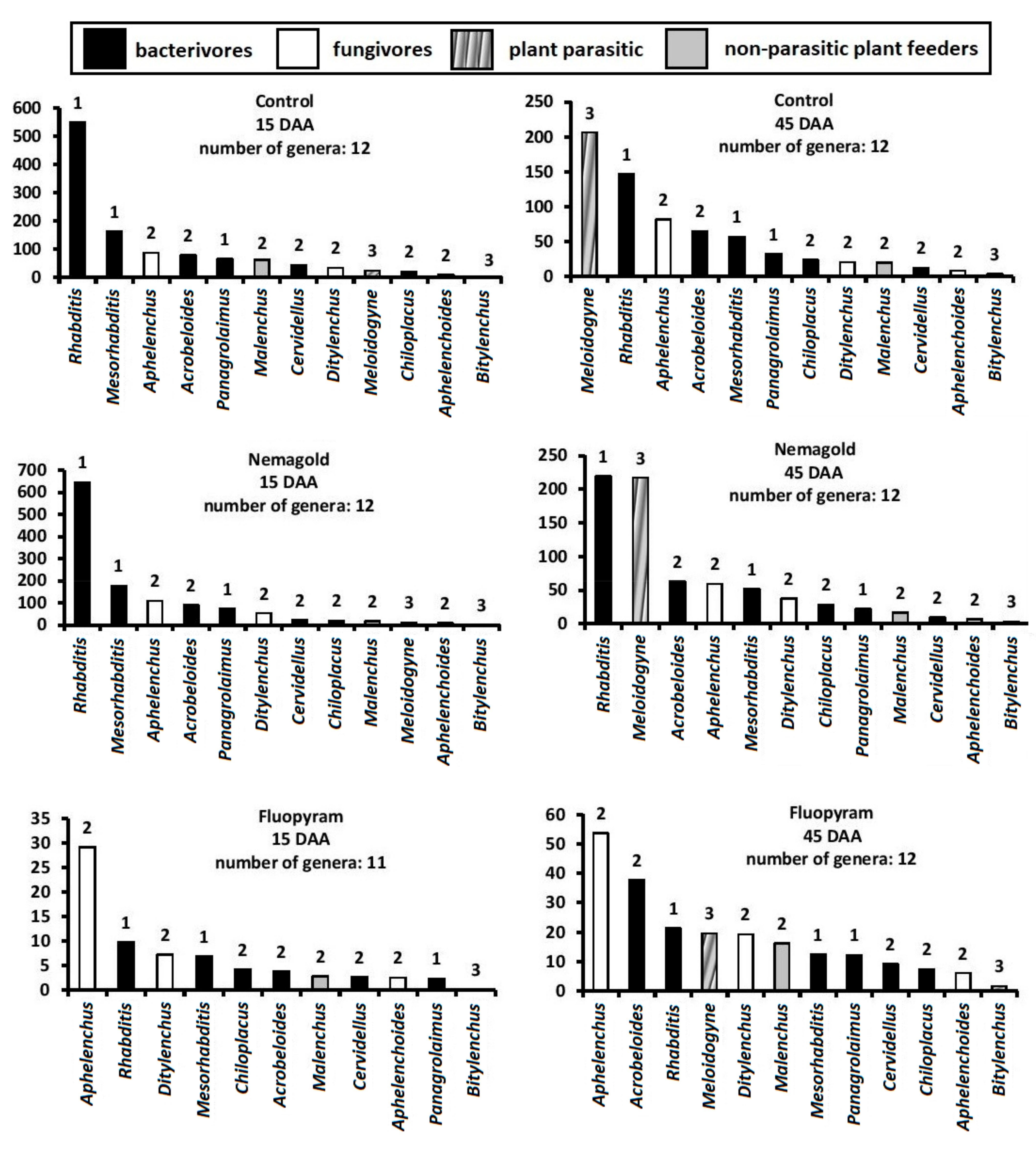
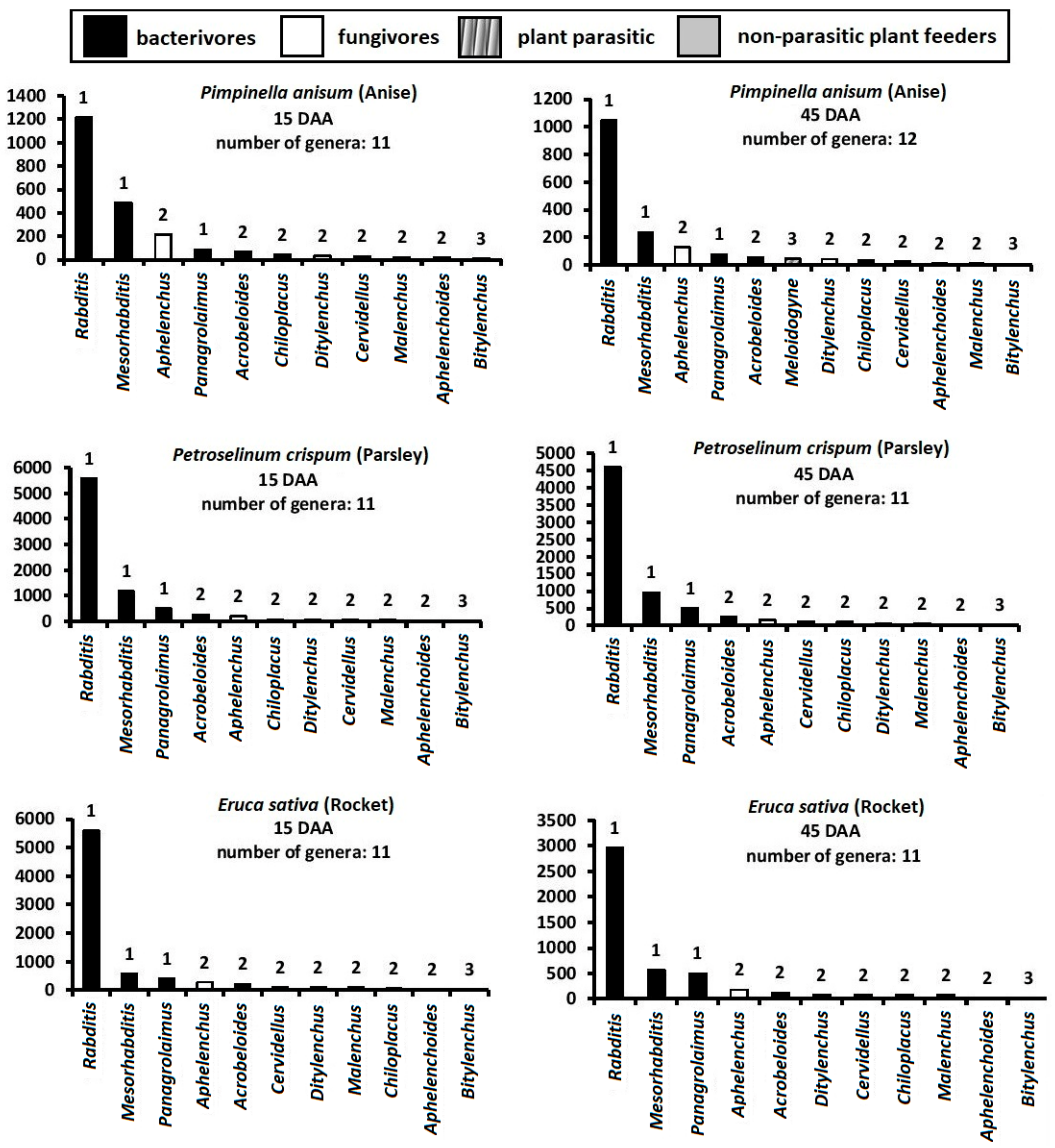
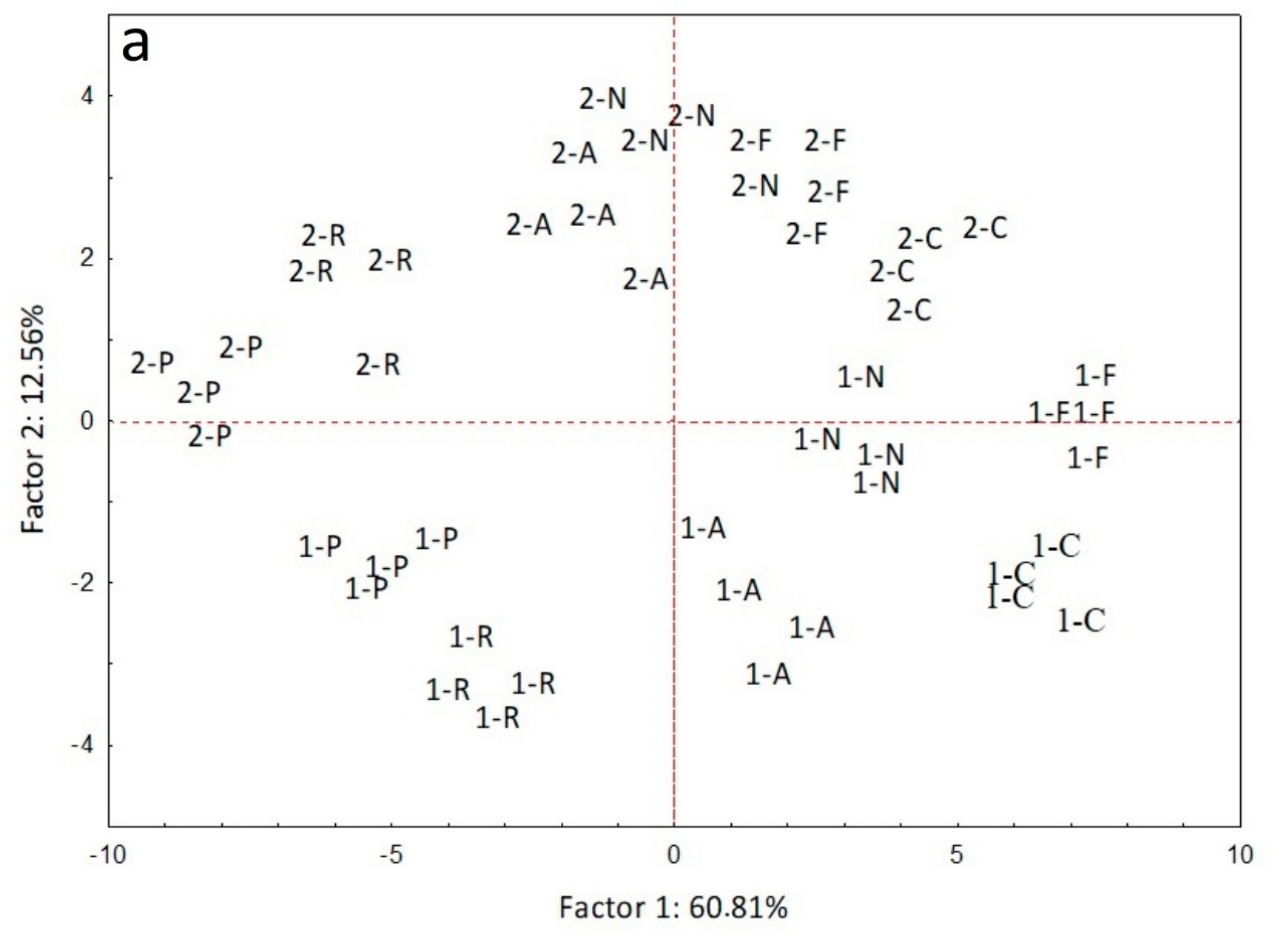
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Van Berkum, J.A.; Hoestra, H. Practical Aspects of Chemical Control of Nematodes in Soil. In Soil Disinfestation; Mulder, D., Ed.; Elsevier: Amsterdam, The Netherlands, 1979; pp. 53–120. [Google Scholar]
- Giannakou, I.O.; Karpouzas, D.G. Evaluation of chemical and integrated strategies as alternatives to methyl bromide for the control of root-knot nematodes in Greece. Pest Manag. Sci. 2003, 59, 883–892. [Google Scholar] [CrossRef]
- Ntalli, N.; Zioga, D.; Argyropoulou, D.M.; Papatheodorou, M.E.; Menkissoglu-Spiroudi, U.; Monokrousos, N. Anise, parsley and rocket as nematicidal soil amendments and their impact on non-target soil organisms. Appl. Soil Ecol. 2019, 143, 17–25. [Google Scholar] [CrossRef]
- Ntalli, N.; Monokrousos, N.; Rumbos, C.; Kontea, D.; Zioga, D.; Argyropoulou, M.D.; Menkissoglu-Spiroudi, U.; Tsiropoulos, N.G. Greenhouse biofumigation with Melia azedarach controls Meloidogyne spp. and enhances soil biological activity. J. Pest Sci. 2018, 91, 29–40. [Google Scholar] [CrossRef]
- Ramirez, R.A.; Henderson, D.R.; Riga, E.; Lacey, L.A.; Snyder, W.E. Harmful effects of mustard bio-fumigants on entomopathogenic nematodes. Biol. Control 2009, 48, 147–154. [Google Scholar] [CrossRef]
- Ferris, H.; Venette, R.C.; Scow, K.M. Soil management to enhance bacterivore and fungivore nematode populations and their nitrogen mineralisation function. Appl. Soil Ecol. 2004, 25, 19–35. [Google Scholar] [CrossRef]
- Nannipieri, P.; Ascher, J.; Ceccherini, M.T.; Landi, L.; Pietramellara, G.; Renella, G. Microbial diversity and soil functions. Eur. J. Soil Sci. 2003, 54, 655–670. [Google Scholar] [CrossRef]
- Van Der Heijden, M.G.A.; Bardgett, R.D.; Van Straalen, N.M. The unseen majority: Soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol. Lett. 2008, 11, 296–310. [Google Scholar] [CrossRef]
- Monokrousos, N.; Charalampidis, G.; Boutsis, G.; Sousanidou, V.; Papatheodorou, E.M.; Argyropoulou, M.D. Plant-induced differentiation of soil variables and nematode community structure in a Mediterranean serpentine ecosystem. Soil Res. 2014, 52, 593–603. [Google Scholar] [CrossRef]
- Ntalli, N.; Tsiafouli, M.A.; Tzani, K.; Mavridi, O.; Oplos, C.; Menkissoglu-Spiroudi, U.; Monokrousos, N. Whey: The soil bio-community enhancer that selectively controls root-knot nematodes. Plants 2019, 8, 445. [Google Scholar] [CrossRef] [PubMed]
- Ferris, H.; Matute, M.M. Structural and functional succession in the nematode fauna of a soil food web. Appl. Soil Ecol. 2003, 23, 93–110. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T. Nematode indicators of organic enrichment. J. Nematol. 2006, 38, 3–12. [Google Scholar] [PubMed]
- Hussey, R.S.; Barker, K.R. Comparison of methods of collecting inocula of Meloidogyne spp. including a new technique. Plant Dis. Rep. 1973, 57, 1025–1028. [Google Scholar]
- Ntalli, N.G.; Ferrari, F.; Menkissoglu-spiroudi, U. Synergistic and antagonistic interactions of terpenes against Meloidogyne incognita and the nematicidal activity of essential oils from seven plants indigenous to Greece. Pest Manag. Sci. 2011, 67, 341–351. [Google Scholar] [CrossRef]
- Aissani, N.; Urgeghe, P.P.; Oplos, C.; Saba, M.; Tocco, G.; Petretto, G.L.; Eloh, K.; Menkissoglu-spiroudi, U.; Ntalli, N.; Caboni, P. Nematicidal activity of the volatilome of Eruca sativa on Meloidogyne incognita. J. Agric. Food. Chem. 2015, 63, 6120–6125. [Google Scholar] [CrossRef]
- Caboni, P.; Saba, M.; Oplos, C.; Aissani, N.; Maxia, A.; Menkissoglu-spiroudi, U.; Ntalli, N. Nematicidal activity of furanocoumarins from parsley against Meloidogyne spp. Pest Manag. Sci. 2015, 71, 1099–1105. [Google Scholar] [CrossRef] [PubMed]
- Findlay, R.H. Determination of microbial community structure using phospholipid fatty acid profiles. In Molecular Microbial Ecology Manual, 2nd ed.; Kowalchuk, G.A., Bruijn, F.J.D., Head, I.M., Akkermans, A.D.L., Elsas, J.D.V., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2004; pp. 983–1004. [Google Scholar]
- Angelina, E.; Papatheodorou, E.M.; Demirtzoglou, T.; Monokrousos, N. Effects of Bacillus subtilis and Pseudomonas fluorescens inoculation on attributes of the lettuce (Lactuca sativa L.) soil rhizosphere microbial community: The role of the management system. Agronomy 2020, 10, 1428. [Google Scholar] [CrossRef]
- S’Jacob, J.J.; van Bezooijen, J. A Manual for Practical Work in Nematology; Department of Nematology, Wageningen Agricultural University: Wageningen, The Netherlands, 1984. [Google Scholar]
- Bongers, T. De Nematoden van Nederland; Pirola: Schoorl, The Netherlands, 1994. [Google Scholar]
- Yeates, G.W.; Bongers, T.; de Goede, R.G.M.; Freckman, D.W.; Georgieva, S.S. Feeding habits in nematode families and genera—An outline for soil ecologists. J. Nematol. 1993, 25, 315–331. [Google Scholar] [PubMed]
- Bongers, T. The maturity index: An ecological measure of environmental disturbance based on nematode species composition. Oecologia 1990, 83, 14–19. [Google Scholar] [CrossRef]
- Bongers, T.; Bongers, M. Functional diversity of nematodes. Appl. Soil Ecol. 1998, 10, 239–251. [Google Scholar] [CrossRef]
- Ferris, H.; Bongers, T.; de Goede, R.G.M. A framework for soil food web diagnostics: Extension of the nematode faunal analysis concept. Appl. Soil Ecol. 2001, 18, 13–29. [Google Scholar] [CrossRef]
- Eisenback, J.D.; Triantaphyllou, H.H. Root-knot Nematodes: Meloidogyne species and races. In Manual of Agricultural Nematology; Nickle, W.R., Ed.; Marcel Dekker: New York, NY, USA, 1991. [Google Scholar]
- Gebremikael, M.T.; Steel, H.; Buchan, D.; Bert, W.; De Neve, S. Nematodes enhance plant growth and nutrient uptake under C and N-rich conditions. Sci. Rep. 2016, 6, 32862. [Google Scholar] [CrossRef] [PubMed]
- Waldo, B.D.; Grabau, Z.J.; Mengistu, T.M.; Crow, W.T. Nematicide effects on non-target nematodes in bermudagrass. J. Nematol. 2019, 51, e2019-09. [Google Scholar] [CrossRef] [PubMed]
- Matadha, N.Y.; Mohapatra, S.; Siddamallaiah, L.; Udupi, V.R.; Gadigeppa, S.; Raja, D.P.; Donagar, S.P.; Hebbar, S.S. Persistence and dissipation of fluopyram and tebuconazole on bell pepper and soil under different environmental conditions. Int. J. Environ. Anal. Chem. 2020, 26, 6077–6086. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 15 DAA | MI | EI | CI | PPI |
|---|---|---|---|---|
| Control | 1.18 ± 0.03 a | 92.31 ± 1.29 b | 3.94 ±0.66 b | 2.32 ± 0.07 b |
| Nemagold | 1.23 ± 0.05 a | 92.60 ± 1.92 b | 4.53 ± 0.28 b | 2.42 ± 0.05 b |
| fluopyram | 1.63 ± 0.03 b | 70.68 ± 2.32 a | 32.78 ± 4.54 c | 2.11 ± 0.11 a |
| P. anisum | 1.17 ± 0.01 a | 94.83 ± 1.22 b | 3.62 ± 0,31 b | 2.32 ± 0.08 b |
| P. crispum | 1.17 ± 0.01 a | 97.65 ± 2.08 b | 1.02 ± 0.07 a | 2.13 ± 0.02 a |
| E. sativa | 1.09 ± 0.01 a | 97.04 ± 1.38 b | 1.57 ± 0.36 a | 2.10 ± 0.01 a |
| 45 DAA | MI | EI | CI | PPI |
| Control | 1.43 ± 0.04 b | 82.82 ± 1.79 b | 10.72 ±1.30 c | 2.92 ± 0.01 c |
| Nemagold | 1.39 ± 0.05 b | 85.08 ± 1.92 b | 9.04 ± 1.43 c | 2.94 ± 0.02 c |
| fluopyram | 1.63 ± 0.07 c | 65.22 ± 3.21 a | 32.16 ± 3.28 d | 2.62 ± 0.06 b |
| P. anisum | 1.18 ± 0.01 a | 94.89 ± 1.32 c | 3.27 ± 0.36 b | 2.81 ± 0.05 c |
| P. crispum | 1.10 ± 0.01 a | 97.02 ± 1.08 c | 1.07 ± 0.06 a | 2.15 ± 0.04 a |
| E. sativa | 1.11 ± 0.02 a | 96.63 ± 1.78 c | 1.73 ± 0.14 a | 2.14 ± 0.04 a |
| Assessment Time | *** | *** | ns | *** |
| Treatment | *** | *** | *** | *** |
| Assessment Time × Treatment | ** | ** | ns | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Monokrousos, N.; Argyropoulou, M.D.; Tzani, K.; Menkissoglou-Spiroudi, U.; Boutsis, G.; D’Addabbo, T.; Ntalli, N. The Effect of Botanicals with Nematicidal Activity on the Structural and Functional Characteristics of the Soil Nematode Community. Agriculture 2021, 11, 326. https://doi.org/10.3390/agriculture11040326
Monokrousos N, Argyropoulou MD, Tzani K, Menkissoglou-Spiroudi U, Boutsis G, D’Addabbo T, Ntalli N. The Effect of Botanicals with Nematicidal Activity on the Structural and Functional Characteristics of the Soil Nematode Community. Agriculture. 2021; 11(4):326. https://doi.org/10.3390/agriculture11040326
Chicago/Turabian StyleMonokrousos, Nikolaos, Maria D. Argyropoulou, Kalliopi Tzani, Urania Menkissoglou-Spiroudi, George Boutsis, Trifone D’Addabbo, and Nikoletta Ntalli. 2021. "The Effect of Botanicals with Nematicidal Activity on the Structural and Functional Characteristics of the Soil Nematode Community" Agriculture 11, no. 4: 326. https://doi.org/10.3390/agriculture11040326
APA StyleMonokrousos, N., Argyropoulou, M. D., Tzani, K., Menkissoglou-Spiroudi, U., Boutsis, G., D’Addabbo, T., & Ntalli, N. (2021). The Effect of Botanicals with Nematicidal Activity on the Structural and Functional Characteristics of the Soil Nematode Community. Agriculture, 11(4), 326. https://doi.org/10.3390/agriculture11040326