The Potential of Termite Mound Spreading for Soil Fertility Management under Low Input Subsistence Agriculture


 and
and
Abstract
:1. Introduction
2. Materials and Methods
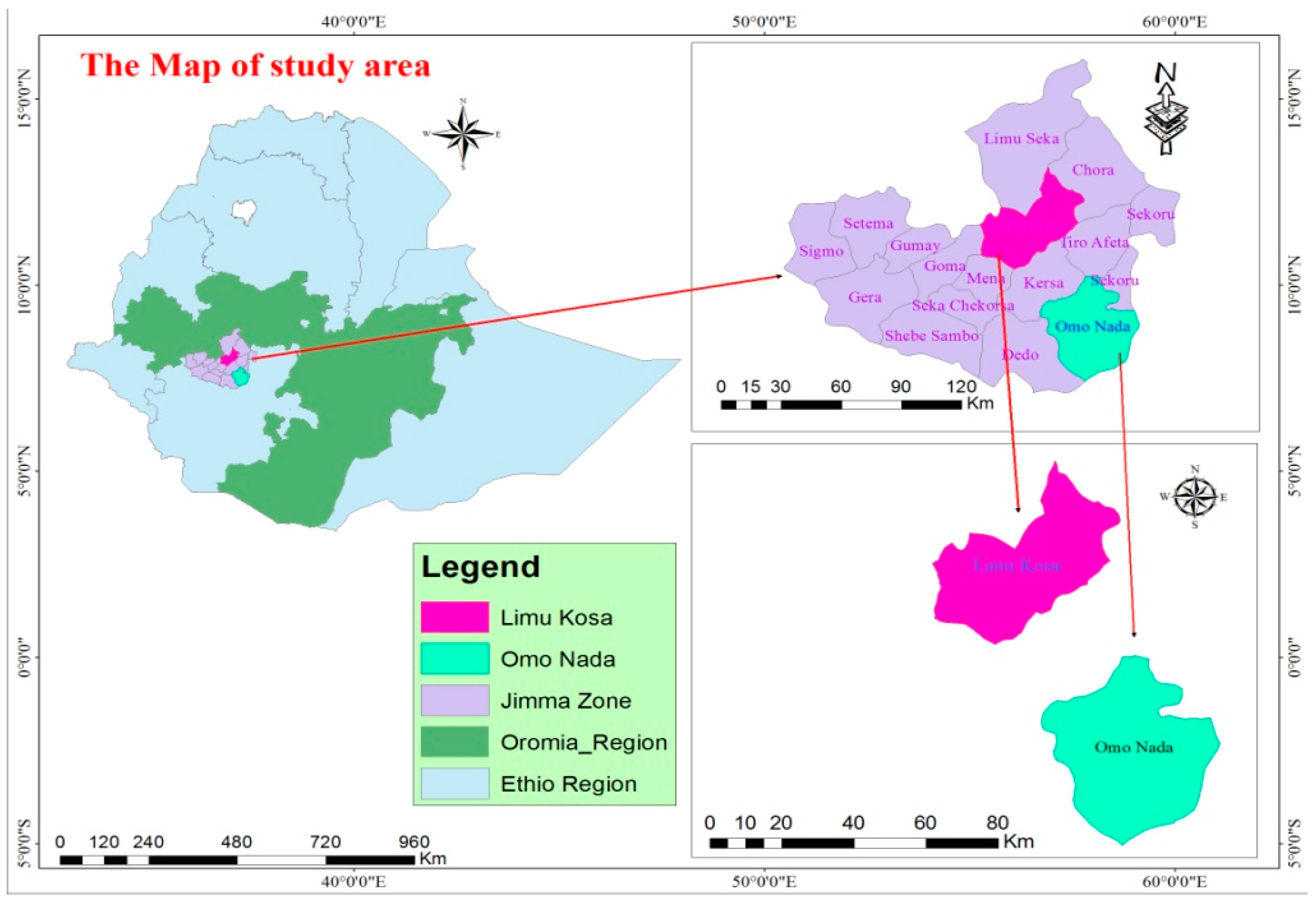
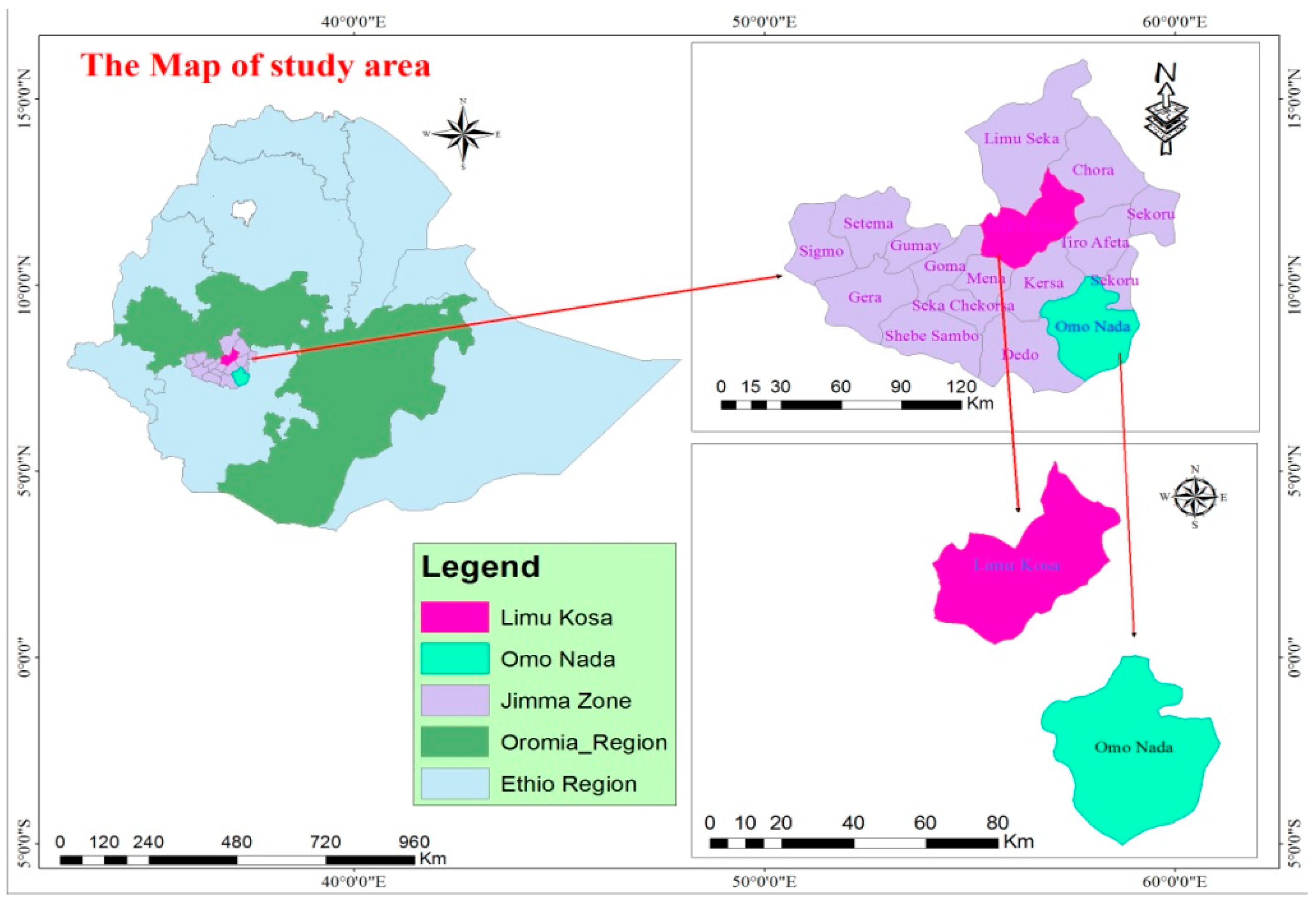
2.1. Site Description
2.2. Soil Sampling
2.3. Soil Chemical Properties
2.4. Soil Physical Properties
2.5. Estimation of Mounds Abundance and Potential of Mound Materials
2.6. Data Analysis
3. Results
3.1. Termite Mound Density and Characteristics
3.2. Physical Soil Properties
3.3. Soil Organic Carbon and Total Nitrogen and Plant-Available Nutrients
3.4. Exchangeable Base Cations and Cation Exchange Capacity
4. Discussion
4.1. Selective Enrichment of Mineral Matter in Mounds
4.2. Aggregate Stability
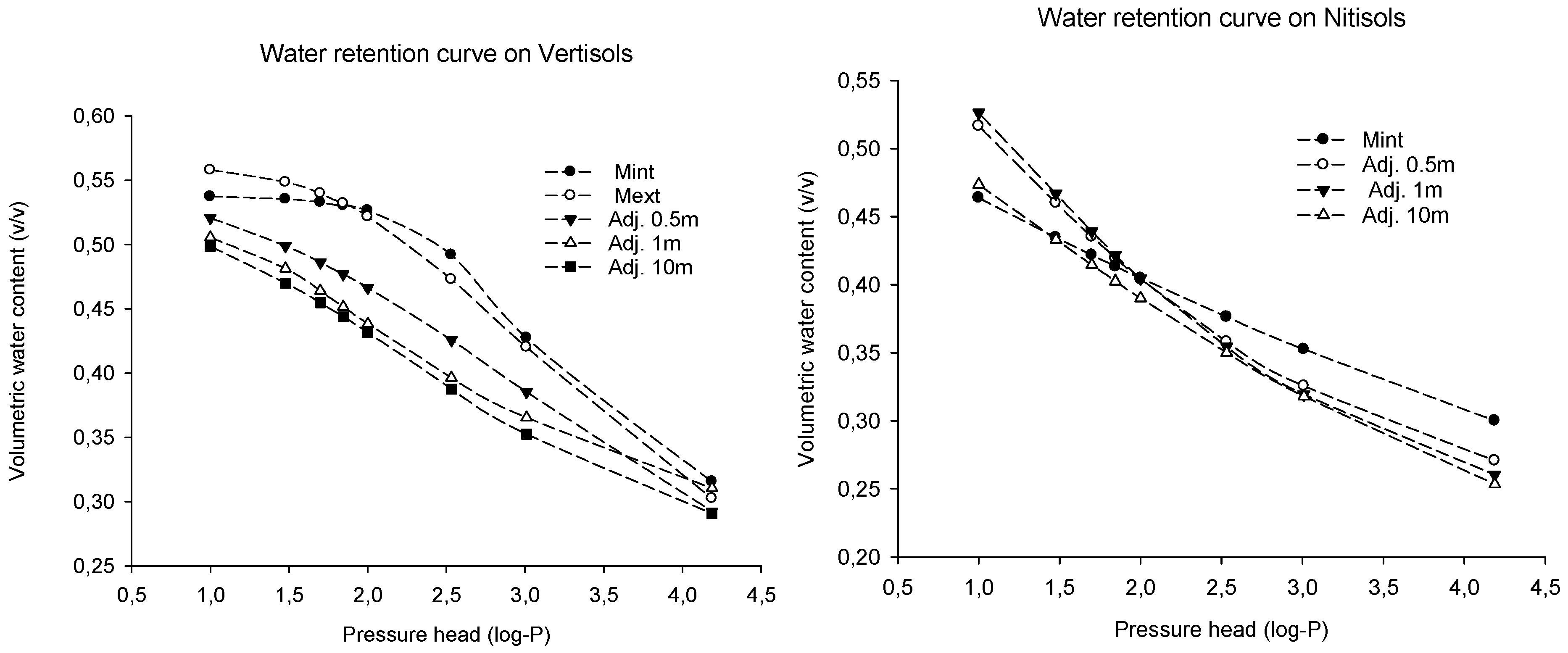
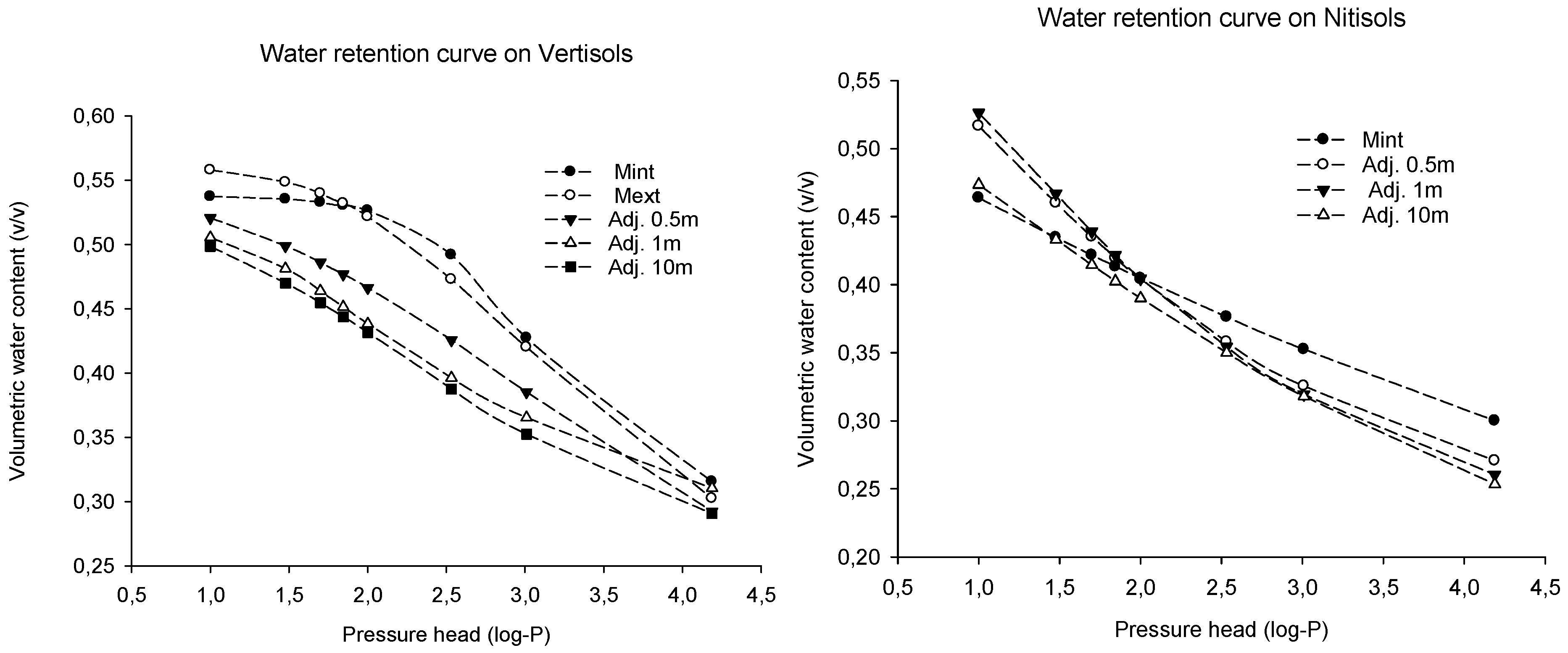
4.3. Effect on Water Retention
4.4. Organic Matter and Total Nitrogen
4.5. Effect on Available Phosphorous
4.6. Exchangeable Base Cations and CEC
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wakweya, T.; Nigussie, A.; Worku, G.; Biresaw, A.; Aticho, A.; Hirko, O.; Ambaw, G.; Mamuye, M.; Dume, B.; Ahmed, M. Long-term effects of bone char and lignocellulosic biochar-based soil amendments on phosphorus adsorption-desorption and crop yield in low-input acidic soils. Soil Use Manag. 2021. [Google Scholar] [CrossRef]
- Schriever, V.A.; Mori, E.; Petters, W.; Boerner, C.; Smitka, M.; Hummel, T. The “Sniffin’ Kids” test—A 14-Item odor identification test for children. PLoS ONE 2014, 9, e101086. [Google Scholar] [CrossRef]
- ATA. Agricultural Transformation Agency; Ethiopian Ministry of Agriculture: Addis Ababa, Ethiopia, 2012.
- Yanggen, D.; Kelly, V.A.; Reardon, T.; Naseem, A. Incentives for Fertilizer Use in Sub-Saharan Africa: A Review of Empirical Evidence on Fertilizer Response and Profitability; Michigan State University, Michigan Department of Agricultural, Food, and Resource Economics: East Lansing, MI, USA, 1998.
- Jiang, J.; Shang, X.; Wang, H.; Xu, Y.-B.; Gao, Y.; Zhou, Q. Diagnostic value of contrast-enhanced ultrasound in thyroid nodules with calcification. Kaohsiung J. Med. Sci. 2015, 31, 138–144. [Google Scholar] [CrossRef] [Green Version]
- Adamou, I.; Pierre, N.J.; Pogenet, P.; Tchimbi, B.; Gonlaina, G. Soil degradation in the Sudano-Guinea savannas of Mbe, Cameroon: Farmers’ perception, indicators and soil fertility management strategies. Res. J. Agric. Biol. 2007, 3, 907–916. [Google Scholar]
- Chianu, J.N.; Nkonya, E.M.; Mairura, F.S.; Chianu, J.N.; Akinnifesi, F.K. Biological nitrogen fixation and socioeconomic factors for legume production in sub-Saharan Africa: A review. Agron. Sustain. Dev. 2011, 31, 139–154. [Google Scholar] [CrossRef] [Green Version]
- Otsuka, K.; Yamano, T. The possibility of a green revolution in sub-Saharan Africa: Evidence from Kenya. J. Agric. Dev. 2005, 2, 8–19. [Google Scholar]
- Toenniessen, G.; Adesina, A.; Devries, J. Building an Alliance for a Green Revolution in Africa. Ann. N. Y. Acad. Sci. 2008, 1136, 233–242. [Google Scholar] [CrossRef]
- MacDonald, G.K.; Bennet, E.M.; Potter, P.A.; Ramankutty, N. Agronomic phosphorus imbalances across the world’s croplands. Proc. Natl. Acad. Sci. USA 2011, 108, 3086–3091. [Google Scholar] [CrossRef] [Green Version]
- Narum, S.M.; Le, T.; Le, D.P.; Lee, J.C.; Donahue, N.D.; Yang, W.; Wilhelm, S. Chapter 4—Passive targeting in nanomedicine: Fundamental concepts, body interactions, and clinical potential. In Nanoparticles for Biomedical Applications; Chung, E.J., Leon, L., Rinaldi, C., Eds.; Elsevier: Amsterdam, The Netherlands, 2020; pp. 19, 37–53. [Google Scholar] [CrossRef]
- Nebiyu, A.; Vandorpe, A.; Diels, J.; Boeckx, P. Nitrogen and phosphorus benefits from faba bean (Vicia faba L.) residues to subsequent wheat crop in the humid highlands of Ethiopia. Nutr. Cycl. Agroecosyst. 2014, 98, 253–266. [Google Scholar] [CrossRef]
- Jeffery, S.; Verheijen, F.; van der Velde, M.; Bastos, A.C. A quantitative review of the effects of biochar application to soils on crop productivity using meta-analysis. Agric. Ecosyst. Environ. 2011, 144, 175–187. [Google Scholar] [CrossRef]
- Nigussie, A.; Kissi, E.; Misganaw, M.; Ambaw, G. Effect of biochar application on soil properties and nutrient uptake of lettuces (Lactuca sativa) grown in chromium polluted soils. Am. Eurasian J. Agric. Environ. Sci. 2012, 12, 369–376. [Google Scholar]
- Zhang, A.; Bian, R.; Pan, G.; Cui, L.; Hussain, Q.; Li, L.; Zheng, J.; Zheng, J.; Zhang, X.; Han, X.; et al. Effects of biochar amendment on soil quality, crop yield and greenhouse gas emission in a Chinese rice paddy: A field study of 2 consecutive rice growing cycles. Field Crops Res. 2012, 127, 153–160. [Google Scholar] [CrossRef]
- Araya, T.; Cornelis, W.M.; Nyssen, J.; Govaerts, B.; Getnet, F.; Bauer, H.; Amare, K.; Raes, D.; Haile, M.; Deckers, J. Medium-term effects of conservation agriculture based cropping systems for sustainable soil and water management and crop productivity in the Ethiopian highlands. Field Crops Res. 2012, 132, 53–62. [Google Scholar] [CrossRef]
- Mugwe, J.; Mugendi, D.; Mucheru-Muna, M.; Merckx, R.; Chianu, J.; Vanlauwe, B. Determinants of the decision to adopt integrated soil fertility management practices by smallholder farmers in the central highlands of kenya. Exp. Agric. 2009, 45, 61–75. [Google Scholar] [CrossRef] [Green Version]
- Tilahun, A.; Kebede, F.; Yamoah, C.; Erens, H.; Mujinya, B.; Verdoodt, A.; Van Ranst, E. Quantifying the masses of Macrotermessubhyalinus mounds and evaluating their use as a soil amendment. Agric. Ecosyst. Environ. 2012, 157, 54–59. [Google Scholar] [CrossRef]
- Adhikary, N.; Erens, H.; Weemaels, L.; Deweer, E.; Mees, F.; Mujinya, B.; Baert, G.; Boeckx, P.; Van Ranst, E. Effects of Spreading Out Termite Mound Material on Ferralsol Fertility, Katanga, D.R. Congo. Commun. Soil Sci. Plant Anal. 2016, 47, 1089–1100. [Google Scholar] [CrossRef]
- Lavelle, P.; Blanchart, E.; Martin, A.; Spain, A.V.; Martin, S. Impact of Soil Fauna on the Properties of Soils in the Humid Tropics. Myth. Sci. Soils Trop. 1992, 29, 157–185. [Google Scholar] [CrossRef]
- Donovan, S.E.; Eggleton, P.; Dubbin, W.E.; Batchelder, M.; Dibog, L. The effect of a soil-feeding termite, Cubitermes fungifaber (Isoptera: Termitidae) on soil properties: Termites may be an important source of soil microhabitat heterogeneity in tropical forests. Pedobiologia 2001, 45, 1–11. [Google Scholar] [CrossRef]
- Jouquet, P.; Tessier, D.; Lepage, M. The soil structural stability of termite nests: Role of clays in Macrotermesbellicosus (Isoptera, Macrotermitinae) mound soils. Eur. J. Soil Biol. 2004, 40, 23–29. [Google Scholar] [CrossRef]
- Mujinya, B.; Van Ranst, E.; Verdoodt, A.; Baert, G.; Ngongo, L. Termite bioturbation effects on electro-chemical properties of Ferralsols in the Upper Katanga (D.R. Congo). Geoderma 2010, 158, 233–241. [Google Scholar] [CrossRef]
- Lavelle, P.; Decaëns, T.; Aubert, M.; Barot, S.; Blouin, M.; Bureau, F.; Margerie, P.; Mora, P.; Rossi, J.-P. Soil invertebrates and ecosystem services. Eur. J. Soil Biol. 2006, 42, S3–S15. [Google Scholar] [CrossRef]
- Kaschuk, G.; Santos, J.C.P.; DE Almeida, J.A.; Sinhorati, D.C.; Berton-Junior, J.F. Termite activity in relation to natural grassland soil attributes. Sci. Agric. Sci. 2006, 63, 583–588. [Google Scholar] [CrossRef]
- Ackerman, I.L.; Teixeira, W.G.; Riha, S.J.; Lehmann, J.; Fernandes, E.C. The impact of mound-building termites on surface soil properties in a secondary forest of Central Amazonia. Appl. Soil Ecol. 2007, 37, 267–276. [Google Scholar] [CrossRef]
- Sarcinelli, T.S.; Schaefer, C.E.G.; Lynch, L.D.S.; Arato, H.D.; Viana, J.H.M.; Filho, M.R.D.A.; Gonçalves, T.T. Chemical, physical and micromorphological properties of termite mounds and adjacent soils along a toposequence in Zona da Mata, Minas Gerais State, Brazil. Catena 2009, 76, 107–113. [Google Scholar] [CrossRef]
- Jouquet, P.; Guilleux, N.; Caner, L.; Chintakunta, S.; Ameline, M.; Shanbhag, R. Influence of soil pedological properties on termite mound stability. Geoderma 2016, 262, 45–51. [Google Scholar] [CrossRef]
- Black, H.; Okwakol, M. Agricultural intensification, soil biodiversity and agroecosystem function in the tropics: The role of termites. Appl. Soil Ecol. 1997, 6, 37–53. [Google Scholar] [CrossRef]
- Debelo, D.G. Faunal survey of the termites of the genus Macrotermes (Isoptera: Termitidae) of Ethiopia. J. Entomol. Nematol. 2018, 10, 50–64. [Google Scholar]
- Jiménez, J.; Decaëns, T. Chemical variations in the biostructures produced by soil ecosystem engineers. Examples from the neotropical savannas. Eur. J. Soil Biol. 2006, 42, S92–S102. [Google Scholar] [CrossRef] [Green Version]
- Mahaney, W.C.; Zippin, J.; Milner, M.W.; Sanmugadas, K.; Hancock, R.G.V.; Aufreiter, S.; Campbell, S.; Huffman, M.A.; Wink, M.; Malloch, D.; et al. Chemistry, mineralogy and microbiology of termite mound soil eaten by the chimpanzees of the Mahale Mountains, Western Tanzania. J. Trop. Ecol. 1999, 15, 565–588. [Google Scholar] [CrossRef]
- Grohmann, C. Termite Mediated Heterogeneity of Soil and Vegetation Patterns in a Semi-Arid Savanna Ecosystem in Namibia. Ph.D. Thesis, The Jullius Maximilians University of Würzburg, Würzburg, Germany, 2010. [Google Scholar]
- Walkley, A.; Black, I.A. An examination of the degtjareff method for determining soil organic matter, and a proposed modification of the chromic acid titration method. Soil Sci. 1934, 37, 29–38. [Google Scholar] [CrossRef]
- Jackson, M.L. Soil Chemical Analysis: Advanced Course; UW-Madison Libraries Parallel Press: Madison, WI, USA, 2005. [Google Scholar]
- Bowman, R.A. A Rapid Method to Determine Total Phosphorus in Soils. Soil Sci. Soc. Am. J. 1988, 52, 1301–1304. [Google Scholar] [CrossRef]
- Cornelis, W.M.; Khlosi, M.; Hartmann, R.; Van Meirvenne, M.; De Vos, B. Comparison of Unimodal Analytical Expressions for the Soil-Water Retention Curve. Soil Sci. Soc. Am. J. 2005, 69, 1902–1911. [Google Scholar] [CrossRef] [Green Version]
- Kemper, W.D.; Rosenau, R.C. Aggregate Stability and Size Distribution. 2018, 425–442. Available online: https://doi.org/10.2136/sssabookser5.1.2ed.c17 (accessed on 12 October 2021).
- Zhang, H.; Schroder, J.L.; Fuhrman, J.K.; Basta, N.T.; Storm, D.E.; Payton, M.E. Path and multiple regression analyses of phosphorous sorption capacity. Soil Sci. Soc. Am. J. 2005, 69, 96–106. [Google Scholar] [CrossRef]
- McComie, L.; Dhanarajan, G. The physical and chemical composition of mounds of Macrotermes carbonarius (Hagen) (Termitidae, Macrotermitinae), in Penang, Malaysia. J. Soil Sci. 1993, 44, 427–433. [Google Scholar] [CrossRef]
- Muwawa, E.M.; Makonde, H.M.; Budambula, N.; Osiemo, Z.L. Chemical properties associated with guts, soil and nest materials of Odontotermes and Macrotermes species from Kenya. J. Biol. Environ. Sci. 2014, 4, 253–263. [Google Scholar]
- Gholami, A.; Riazi, F. Impact of Termite Activity on Physical and Chemical Properties. J. Basic Appl. Sci. Res. 2012, 2, 5581–5582. [Google Scholar]
- Brady, N.C.; Weil, R.R. The Nature and Properties of Soils; Prentice-Hall Inc.: Upper Saddle River, NJ, USA, 1996. [Google Scholar]
- WRB—World Reference Base for Soil Resources. International Soil Classification System for Naming Soils and Creating Legends for Soil Maps; Food and agriculture organization (FAO): Rome, Italy, 2015. [Google Scholar]
- Jouquet, P.; Lepage, M.; Velde, B. Termite soil preferences and particle selections: Strategies related to ecological requirements. Insectes Sociaux 2002, 49, 1–7. [Google Scholar] [CrossRef]
- Six, J.; Bossuyt, H.; Degryze, S.; Denef, K. A history of research on the link between (micro) aggregates, soil biota, and soil organic matter dynamics. Soil Tillage Res. 2004, 79, 7–31. [Google Scholar] [CrossRef]
- Oades, J. The role of biology in the formation, stabilization and degradation of soil structure. Geoderma 1993, 56, 377–400. [Google Scholar] [CrossRef]
- Jastrow, J.; Miller, R. Methods for assessing the effects of biota on soil structure. Agric. Ecosyst. Environ. 1991, 34, 279–303. [Google Scholar] [CrossRef]
- Jungerius, P.; Ancker, J.V.D.; Mücher, H. The contribution of termites to the microgranular structure of soils on the Uasin Gishu Plateau, Kenya. CATENA 1999, 34, 349–363. [Google Scholar] [CrossRef]
- Bignell, D.E.; Holt, J.A. Termites. In Encyclopedia of Soil Science; Lal, R., Ed.; Marcel Dekker Inc.: New York, NY, USA, 2002; pp. 1305–1307. [Google Scholar]
- Denef, K.; Six, J. Clay mineralogy determines the importance of biological versus abiotic processes for macroaggregate formation and stabilization. Eur. J. Soil Sci. 2004, 56, 469–479. [Google Scholar] [CrossRef]
- Rawls, W.; Pachepsky, Y.; Ritchie, J.; Sobecki, T.; Bloodworth, H. Effect of soil organic carbon on soil water retention. Geoderma 2003, 116, 61–76. [Google Scholar] [CrossRef]
- Bauer, A.; Black, A.L. Soil Carbon, Nitrogen, and Bulk Density Comparisons in Two Cropland Tillage Systems after 25 Years and in Virgin Grassland. Soil Sci. Soc. Am. J. 1981, 45, 1166–1170. [Google Scholar] [CrossRef]
- Suzuki, S.; Noble, A.D.; Ruaysoongnern, S.; Chinabut, N. Improvement in Water-Holding Capacity and Structural Stability of a Sandy Soil in Northeast Thailand. Arid. Land Res. Manag. 2007, 21, 37–49. [Google Scholar] [CrossRef]
- Garba, M.; Cornelis, W.M.; Steppe, K. Effect of termite mound material on the physical properties of sandy soil and on the growth characteristics of tomato (Solanum lycopersicum L.) in semi-arid Niger. Plant Soil 2010, 338, 451–466. [Google Scholar] [CrossRef]
- Fall, S.; Brauman, A.; Chotte, J.-L. Comparative distribution of organic matter in particle and aggregate size fractions in the mounds of termites with different feeding habits in Senegal: Cubitermesniokoloensis and Macrotermesbellicosus. Appl. Soil Ecol. 2001, 17, 131–140. [Google Scholar] [CrossRef]
- Adekayode, F.; Ogunkoya, M. Comparative study of clay and organic matter content of termite mounds and the surrounding soils. In African Crop Science Conference Proceedings; African Crop Science Society: Kampala, Uganda, 2009; Volume 9, pp. 379–384. [Google Scholar]
- Wood, T.; Johnson, R.; Anderson, J. Modification of soils in Nigerian savanna by soil-feeding Cubitermes (Isoptera, Termitidae). Soil Biol. Biochem. 1983, 15, 575–579. [Google Scholar] [CrossRef]
- Gillman, L.R.; Jefferies, M.K.; Richards, G. Non-soil constituents of termite (Coptotermes acinaciformis) mounds. Aust. J. Biol. Sci. 1972, 25, 1005–1014. [Google Scholar] [CrossRef]
- Schaefer, C.E.R. Brazilian latosols and their B horizon microstructure as long-term biotic constructs. Soil Res. 2001, 39, 909–926. [Google Scholar] [CrossRef]
- Verlinden, A.; Seely, M.; Hillyer, A. Settlement, trees and termites in Central North Namibia: A case of indigenous resource management. J. Arid Environ. 2006, 66, 307–335. [Google Scholar] [CrossRef]
- Okwakol, M.J.N.; Sekamatte, M.B. Soil macrofauna research in ecosystems in Uganda. Afr. J. Ecol. 2007, 45, 2–8. [Google Scholar] [CrossRef]
- Stevenson, F.J. Cycles of Soils: Carbon, Nitrogen, Phosphorus, Sulfur, Micronutrients; John Wiley & Sons: Hoboken, NJ, USA, 1999. [Google Scholar]
- Hue, N.V. Effects of organic acids/anions on p sorption and phytoavailability in soils with different mineralogies. Soil Sci. 1991, 152, 463–471. [Google Scholar] [CrossRef]
- Pomeroy, D.E. Some effects of mound-building termites on the soils of a semi-arid area of Kenya. J. Soil Sci. 1983, 34, 555–570. [Google Scholar] [CrossRef]
- Aloys, K. The Influence of Mound-Building Termites on the Mineralogical Composition of Ferralsols, Geology and Soil Science. Master’s Thesis, Ghent University, Ghent, Belgium, 2013. [Google Scholar]
- Wielemaker, W. Soil Formation by Termites: A Study in the Kisii Area, Kenya. Ph.D. Thesis, Department of Soil Science and Geology, Agricultural University of Wageningen, Wageningen, The Netherlands, 1984. [Google Scholar]
- Coventry, R.; Holt, J.; Sinclair, D. Nutrient cycling by mound building termites in low fertility soils of semi-arid tropical Australia. Soil Res. 1988, 26, 375–390. [Google Scholar] [CrossRef]
- Jouquet, P.; Bottinelli, N.; Lata, J.-C.; Mora, P.; Caquineau, S. Role of the fungus-growing termite Pseudacanthotermes spiniger (Isoptera, Macrotermitinae) in the dynamic of clay and soil organic matter content. An experimental analysis. Geoderma 2007, 139, 127–133. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
| Mound Characteristics | Omo Nadda, Vertisols (n = 22) | Limmu Kosa, Nitisols (n = 19) | ||||
|---|---|---|---|---|---|---|
| Minimum | Mean ± SEM | Maximum | Minimum | Mean ± SEM | Maximum | |
| Height (m) | 0.3 | 0.7 ± 0.2 | 1.1 | 0.2 | 0.6 ± 0.2 | 1 |
| Basal area (m2) | 0.8 | 3.2 ± 1.5 | 10.9 | 3.1 | 13.2 ± 4.9 | 31.9 |
| Basal area (m2 ha−1) | 3.6 | 14.0 ± 6.5 | 47.2 | 27.5 | 119.0 ± 44.8 | 286.7 |
| Volume (m3) | 0.2 | 1.3 ± 0.7 | 4.5 | 0.4 | 4.4 ± 2.6 | 14.2 |
| Mass kg per mound | 139 | 1454 ± 8 | 5033 | 327 | 4320 ± 2900 | 14,134 |
| Mass per ha (t ha−1) | 0.6 | 6.3 ± 3.3 | 21.8 | 2.9 | 38.9 ± 23.5 | 127.2 |
| Sites | Clay (%) | Silt (%) | Sand (%) | USDA Textural Class | ρb (Mgm−3) |
|---|---|---|---|---|---|
| Omo Nadda, Vertisols | |||||
| Mint | 60.7 ± 2.7 ba | 23.3 ± 2.7 a | 16.0 ± 0.0 a | Clay | 1.29 ± 0.0 a |
| Mext | 58 ± 2.3 ab | 23.3 ± 2.7 a | 18.7 ± 1.3 a | Clay | 1.21 ± 0.0 a |
| Adj. 0.5 m | 52.67 ± 0.7 ab | 28.0 ± 1.2 a | 19.3 ± 1.3 a | Clay | 1.14 ± 0.0 a |
| Adj. 1 m | 50.0 ± 1.2 a | 30.0 ± 1.2 a | 20.0 ± 0.0 a | Clay | 1.16 ± 0.1 a |
| Adj. 10 m | 51.1 ± 3.3 ab | 30.7 ± 0.7 a | 18.0 ± 3.2 a | Clay | 1.25 ± 0.0 a |
| p-value | * | NS | NS | NS | |
| Limmu kosa, Nitisols | |||||
| Mint | 80.0 ± 1.2 c | 15 ± 0.7 a | 5 ± 1.3 a | Clay | 1.10 ± 0.0 a |
| Mext | 73.3 ± 4.7 c | 17 ± 1.3 a | 9 ± 3.3 ab | Clay | 1.11 ± 0.1 a |
| Adj. 0.5 m | 41.0 ± 1.76 ab | 31 ± 1.3 c | 27 ± 0.7 bc | Clay | 1.02 ± 0.1 a |
| Adj. 1 m | 36.0 ± 1.2 a | 32 ± 1.2 c | 32 ± 2.0 cd | Clay L. | 1.08 ± 0.0 a |
| Adj. 10 m | 40.0 ± 1.2 ab | 31 ± 2.7 bc | 29 ± 3.7 c | Clay L. | 1.07 ± 0.0 a |
| p-value | * | * | * | NS | |
| Sites | WSA-FW | WSA-SW | MWD-FW | MWD-SW |
|---|---|---|---|---|
| Omo Nadda, Vertisols | ||||
| Mint | 71.5 ± 4.4 a | 89.1 ± 1.6 ab | 0.59 ± 0.1 a | 0.9 ± 0.0 a |
| Mext | 77.4 ± 1.5 ab | 83.2 ± 0.6 a | 0.75 ± 0.1 ab | 1.0 ± 0.0 a |
| Adj. 0.5 m | 83.2 ± 3.7 ab | 92.7 ± 2.1 b | 0.86 ± 0.1 b | 1. ± 0.1 a |
| Adj. 1 m | 88.1 ± 1.1 b | 94.7 ± 0.8 b | 0.98 ± 0.0 b | 1.0 ± 0.0 a |
| Adj. 10 m | 84.1 ± 5.1 ab | 92.0 ± 2.5 b | 0.91 ± 0.1 b | 0.9 ± 0.0 a |
| p-value | * | * | * | NS |
| Limmu kosa, Nitisols | ||||
| Mint | 66.8 ± 5.6 a | 98.3 ± 0.4 a | 0.73 ± 0.1 a | 1.1 ± 0.0 a |
| Mext | 66.1 ± 2.5 a | 97.8 ± 0.4 a | 0.73 ± 0.1 a | 1.1 ± 0.0 a |
| Adj. 0.5 m | 96.6 ± 1.1 b | 99.4 ± 0.3 a | 1.05 ± 0.0 b | 1.1 ± 0.0 a |
| Adj. 1 m | 95.3 ± 0.8 b | 99.5 ± 0.3 a | 0.98 ± 0.1 b | 1.1 ± 0.0 a |
| Adj. 10 m | 96.0 ± 1.4 b | 99.5 ± 0.0 a | 1.00 ± 0.1 b | 1.0 ± 0.1 a |
| p-value | * | NS | * | NS |
| Site | FC (v/v) | PWP (v/v) | PAW (v/v) | Porosity |
|---|---|---|---|---|
| Omo Nadda, Vertisols | ||||
| Mint | 0.53 ± 0.02 a | 0.32 ± 0.00 a | 0.21 ± 0.03 a | 51.21 ± 0.71 a |
| Mext | 0.52 ± 0.03 a | 0.30 ± 0.01 a | 0.22 ± 0.02 a | 54.23 ± 1.30 a |
| Adj. 0.5 m | 0.47 ± 0.03 a | 0.29 ± 0.02 a | 0.17 ± 0.01 a | 57.01 ± 1.15 a |
| Adj. 1 m | 0.44 ± 0.01 a | 0.31 ± 0.02 a | 0.13 ± 0.02 a | 56.13 ± 1.85 a |
| Adj.10 m | 0.43 ± 0.05 a | 0.29 ± 0.06 a | 0.14 ± 0.00 a | 52.71 ± 1.48 a |
| p-value | NS | NS | NS | NS |
| Limmu kosa, Nitisols | ||||
| Mint | 0.40 ± 0.02 a | 0.30 ± 0.01 a | 0.10 ± 0.01 a | 58.31 ± 0.12 a |
| Mext | 0.41 ± 0.02 a | 0.28 ± 0.01 a | 0.13 ± 0.01 a | 58.17 ± 2.66 a |
| Adj. 0.5 m | 0.40 ± 0.01 a | 0.27 ± 0.01 a | 0.13 ± 0.01 a | 61.41 ± 1.70 a |
| Adj. 1 m | 0.40 ± 0.02 a | 0.26 ± 0.01 a | 0.14 ± 0.01 a | 59.15 ± 0.47 a |
| Adj.10 m | 0.39 ± 0.01 a | 0.25 ± 0.00 a | 0.14 ± 0.01 a | 59.68 ± 0.63 a |
| p-value | NS | NS | NS | NS |
| Sites | pH (H2O) | SOC (%) | Nt (%) | Pav (mg kg−1) |
|---|---|---|---|---|
| Omo Nadda, Vertisols | ||||
| Mint | 7.1 ± 0.23 a | 1.68 ± 0.27 a | 0.12 ± 0.02 a | 6.53 ± 0.29 a |
| Mext | 7.1 ± 0.12 a | 1.53 ± 0.50 a | 0.08 ± 0.00 a | 6.76 ± 0.11 a |
| Adj.0.5 m | 6.8 ± 0.06 a | 2.88 ± 0.48 b | 0.13 ± 0.01 a | 11.33 ± 1.42 b |
| Adj. 1 m | 6.6 ± 0.09 a | 2.79 ± 0.012 b | 0.14 ± 0.01 a | 9.37 ± 0.27 ab |
| Adj. 10 m | 6.9 ± 0.10 a | 2.40 ± 0.42 ab | 0.10 ± 0.01 a | 5.34 ± 0.77 a |
| p-value | NS | * | NS | * |
| Limmu kosa, Nitisols | ||||
| Mint | 5.6 ± 0.20 ab | 1.49 ± 0.04 a | 0.12 ± 0.01 a | 0.00 ± 0.00 a |
| Mext | 5.6 ± 0.12 ab | 1.79 ± 0.57 bc | 0.10 ± 0.01 a | 0.19 ± 0.19 a |
| Adj.0.5 m | 5.9 ± 0.15 b | 3.21 ± 0.66 cd | 0.14 ± 0.01 a | 1.11 ± 0.13 a |
| Adj. 1 m | 5.8 ± 0.03 b | 3.52 ± 0.19 d | 0.14 ± 0.01 a | 1.45 ± 0.30 a |
| Adj. 10 m | 5.8 ± 0.06 b | 2.81 ± 0.29 bcd | 0.11 ± 0.02 a | 2.03 ± 1.09 a |
| p-value | * | * | NS | NS |
| Site | Alox (g kg−1) | Feox (g kg−1) | Ptot (g kg−1) | Pox (g kg−1) | Pav(mg kg−1) | PSI (%) |
|---|---|---|---|---|---|---|
| Limmu kosa, Nitisols | ||||||
| Mint | 3.63 ± 0.23 a | 7.11 ± 0.10 a | 0.47 ± 0.03 a | 0.05 ± 0.01 a | 0.00 ± 0.00 a | 0.61 ± 0.1 a |
| Mext | 3.44 ± 0.14 a | 6.84 ± 0.29 a | 0.48 ± 0.00 a | 0.06 ± 0.01 a | 0.19 ± 0.19 a | 0.77 ± 0.1 a |
| Adj.0.5 | 3.15 ± 0.21 a | 11.87 ± 0.97 bc | 0.68 ± 0.02 bc | 0.14 ± 0.01 b | 1.11 ± 0.13 a | 1.39 ± 0.1 b |
| Adj.1 m | 3.38 ± 0.29 a | 12.86 ± 0.80 c | 0.69 ± 0.04 c | 0.15 ± 0.02 b | 1.45 ± 0.30 a | 1.40 ± 0.2 b |
| Adj. 10 | 3.59 ± 0.40 a | 11.92 ± 0.82 bc | 0.62 ± 0.01 bc | 0.15 ± 0.01 b | 2.03 ± 1.09 a | 1.38 ± 0.2 b |
| p-value | NS | * | * | NS | NS | * |
| Site | Ca | Mg | K | Na | CEC |
|---|---|---|---|---|---|
| Omo Nadda, Vertisols | |||||
| Mint | 24.49 ± 1.25 b | 4.83 ± 0.37 a | 1.24 ± 0.17 a | 0.67 ± 0.16 b | 30.06 ± 0.60 b |
| Mext | 24.44 ± 0.51 b | 4.86 ± 0.51 a | 1.04 ± 0.12 a | 0.66 ± 0.10 b | 29.01 ± 1.01 ab |
| Adj.0.5 m | 22.74 ± 1.1 ab | 4.87 ± 0.34 a | 1.01 ± 0.03 a | 0.28 ± 0.0 ab | 28.56 ± 0.43 ab |
| Adj. 1 m | 20.87 ± 1.8 ab | 4.59 ± 0.13 a | 0.89 ± 0.04 a | 0.22 ± 0.03 a | 27.69 ± 1.60 ab |
| Adj. 10 m | 18.35 ± 1.04 a | 4.03 ± 0.05 a | 0.87 ± 0.06 a | 0.46 ± 0.1 ab | 24.38 ± 1.65 a |
| p-value | * | NS | NS | * | * |
| Limmu kosa, Nitisols | |||||
| Mint | 4.30 ± 0.71 a | 1.63 ± 0.06 a | 1.02 ± 0.17 a | 0.09 ± 0.02 a | 18.60 ± 0.63 a |
| Mext | 4.22 ± 0.15 a | 1.97 ± 0.11 ab | 0.82 ± 0.27 a | 0.07 ± 0.01 a | 19.41 ± 0.24 ab |
| Adj.0.5 m | 6.55 ± 0.13 b | 2.52 ± 0.12 cd | 0.76 ± 0.07 a | 0.09 ± 0.02 a | 21.46 ± 0.25 bc |
| Adj. 1 m | 6.81 ± 0.40 b | 2.52 ± 0.18 cd | 0.68 ± 0.06 a | 0.07 ± 0.01 a | 21.82 ± 0.78 c |
| Adj. 10 m | 6.33 ± 0.27 ab | 2.28 ± 0.1 bcd | 0.69 ± 0.18 a | 0.08 ± 0.01 a | 21.46 ± 0.10 bc |
| p-value | * | * | NS | NS | * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tilahun, A.; Cornelis, W.; Sleutel, S.; Nigussie, A.; Dume, B.; Van Ranst, E. The Potential of Termite Mound Spreading for Soil Fertility Management under Low Input Subsistence Agriculture. Agriculture 2021, 11, 1002. https://doi.org/10.3390/agriculture11101002
Tilahun A, Cornelis W, Sleutel S, Nigussie A, Dume B, Van Ranst E. The Potential of Termite Mound Spreading for Soil Fertility Management under Low Input Subsistence Agriculture. Agriculture. 2021; 11(10):1002. https://doi.org/10.3390/agriculture11101002
Chicago/Turabian StyleTilahun, Amsalu, Wim Cornelis, Steven Sleutel, Abebe Nigussie, Bayu Dume, and Eric Van Ranst. 2021. "The Potential of Termite Mound Spreading for Soil Fertility Management under Low Input Subsistence Agriculture" Agriculture 11, no. 10: 1002. https://doi.org/10.3390/agriculture11101002
APA StyleTilahun, A., Cornelis, W., Sleutel, S., Nigussie, A., Dume, B., & Van Ranst, E. (2021). The Potential of Termite Mound Spreading for Soil Fertility Management under Low Input Subsistence Agriculture. Agriculture, 11(10), 1002. https://doi.org/10.3390/agriculture11101002