Effects of Drip Irrigation with Plastic on Photosynthetic Characteristics and Biomass Distribution of Muskmelon



Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Measured Indicators
2.2.1. Leaf Photosynthetic Characteristics
2.2.2. Biomass, Yield, and Total Nitrogen Contents
2.2.3. Soil Urease Activity
2.3. Data Processing
3. Results
3.1. Net CO2 Assimilation Rate (Pn)
3.2. Stomatal Conductance (Gs)
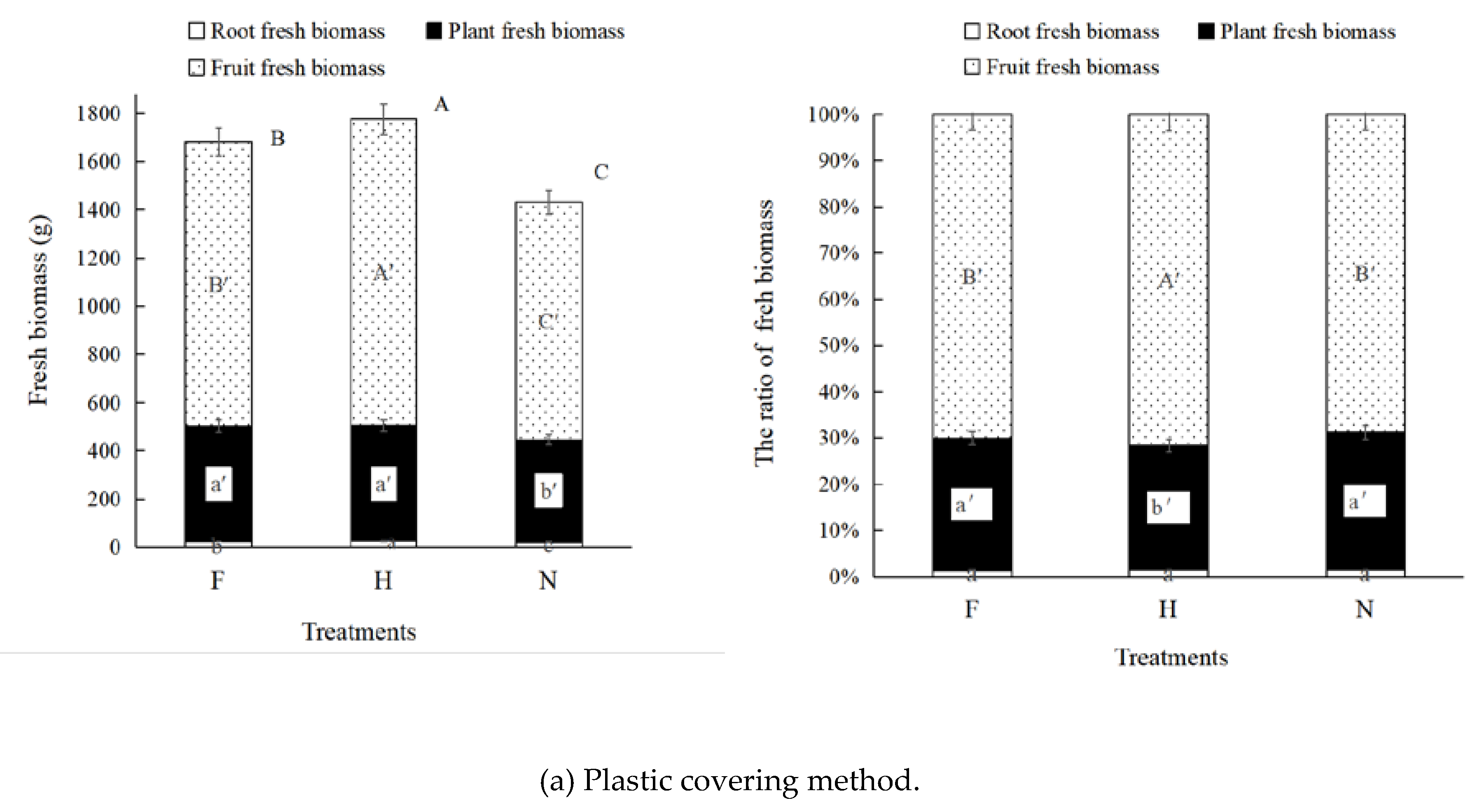
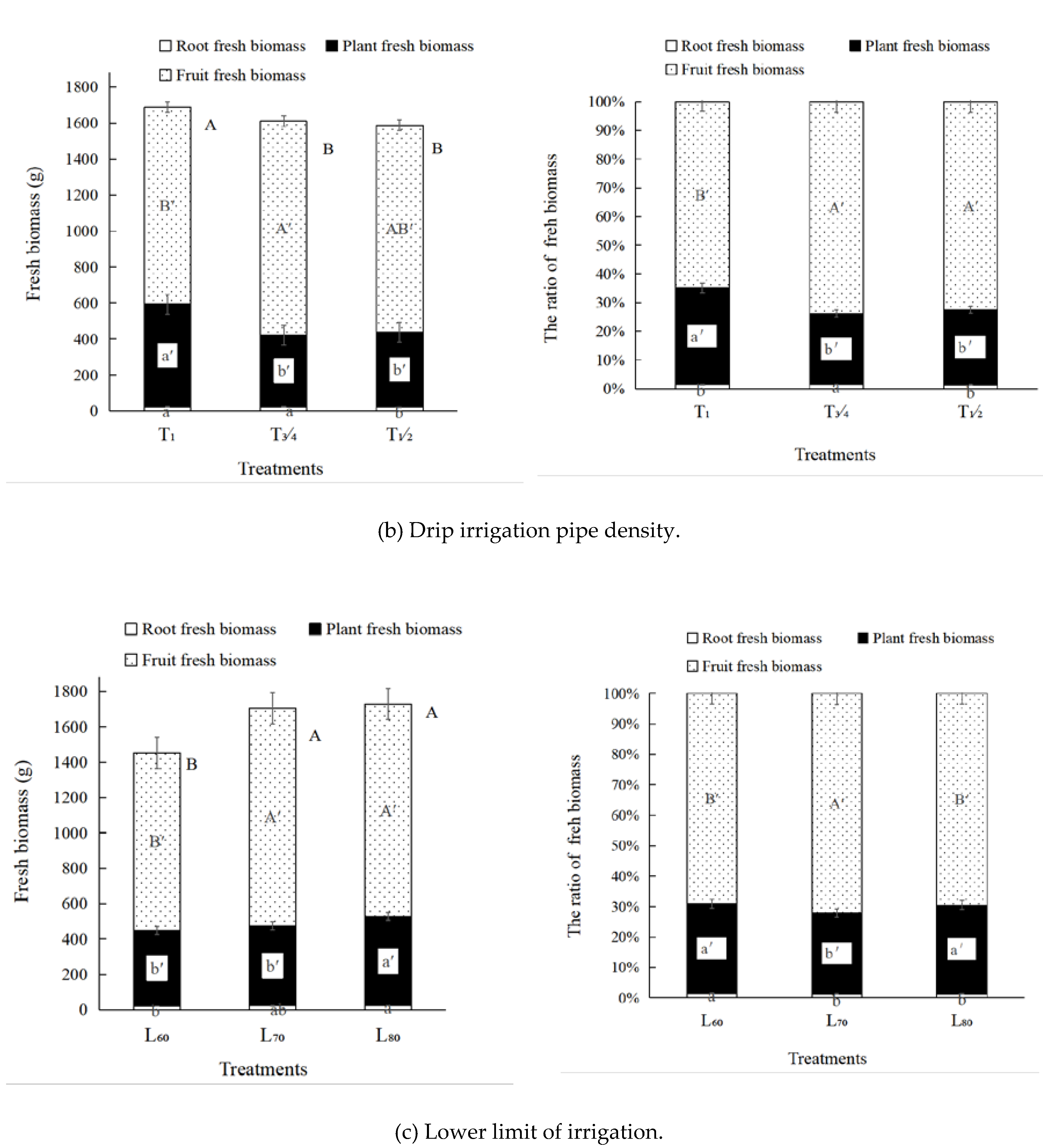
3.3. Muskmelon Biomass
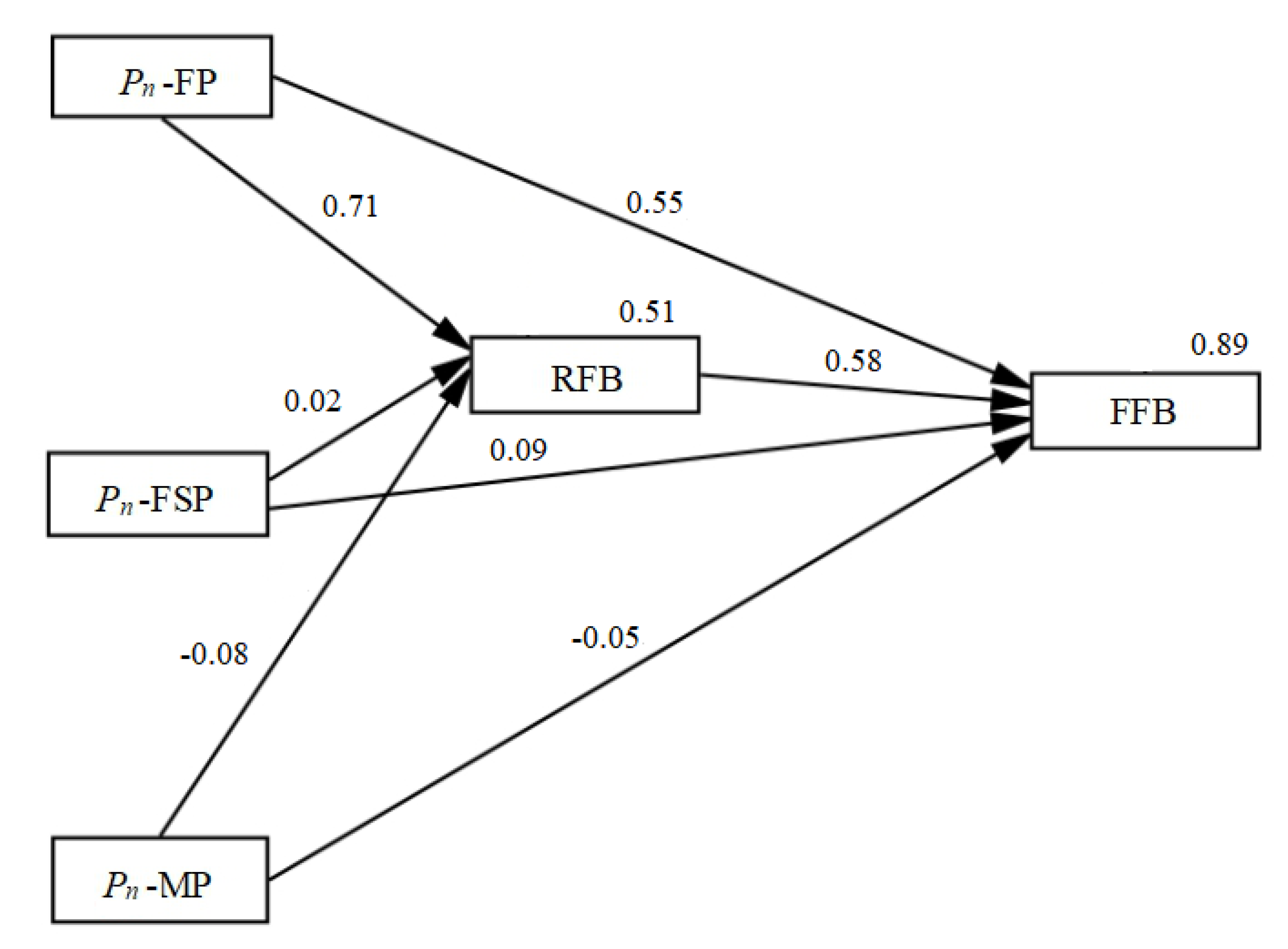
3.4. Simple Correlation Analysis and Structural Equation Model
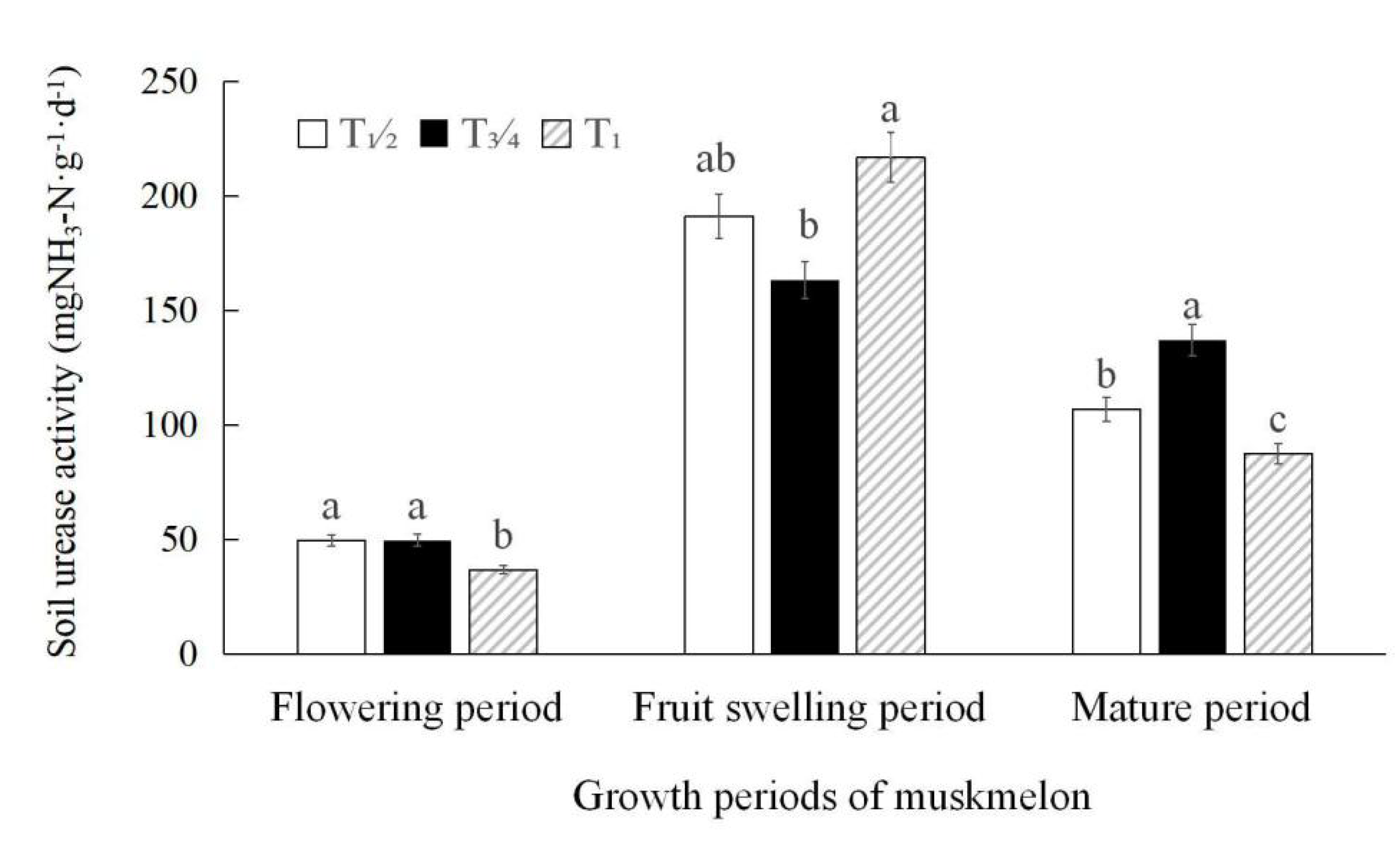
3.5. Soil Urease Activity
4. Discussion
4.1. Half Plastic Covering was Beneficial to Pn and Fresh Biomass
4.2. The Smaller Drip Irrigation Pipe Density was Beneficial to Pn
4.3. Lower Limit of Irrigation Significantly Affected Pn during Fruit Swelling Period
4.4. Determination of the Combination of Different Mulching Drip Irrigation Measures
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Urban, O.; Hlaváčová, M.; Klem, K.; Novotná, K.; Rapantová, B.; Smutná, P.; Horáková, V.; Hlavinka, P.; Karpa, P.; Trnka, M. Combined effects of drought and high temperature on photosynthetic characteristics in four winter wheat genotypes. Fild. Crop. Res. 2018, 223, 137–149. [Google Scholar] [CrossRef]
- Fang, X.M.; Li, Y.S.; Nie, J.; Wang, C.; Huang, K.H.; Zhang, Y.K.; Zhang, Y.L.; She, H.Z.; Liu, X.B.; Ruan, R.W.; et al. Effects of nitrogen fertilizer and planting density on the leaf photosynthetic characteristics, agronomic traits and grain yield in common buckwheat (Fagopyrum esculentum M.). Fild. Crop. Res. 2018, 219, 160–168. [Google Scholar] [CrossRef]
- Ali, S.; Xu, Y.Y.; Jia, Q.M.; Ahmad, I.; Wei, T.; Ren, X.; Zhang, P.; Din, R.; Cai, T.; Jia, Z. Cultivation techniques combined with deficit irrigation improves winter wheat photosynthetic characteristics, dry matter translocation and water use efficiency under simulated rainfall conditions. Agric. Water. Manag. 2018, 201, 207–218. [Google Scholar] [CrossRef]
- Reynolds, P.E.; Simpson, J.A.; Thevathasan, N.V.; Gordon, A.M. Effects of tree competition on corn and soybean photosynthesis, growth, and yield in a temperate tree-based agroforestry intercropping system in southern Ontario, Canada. Ecol. Eng. 2007, 29, 362–371. [Google Scholar] [CrossRef]
- Wasaya, A.; Tahir, M.; Ali, H.; Hussain, M.; Yasir, T.A.; Sher, A.; Ijaz, M. Influence of varying tillage systems and nitrogen application on crop allometry, chlorophyll contents, biomass production and net returns of maize (Zea mays L.). Soil. Till. Res. 2017, 170, 18–26. [Google Scholar] [CrossRef]
- Liu, H.; Gan, W.; Rengel, Z.; Zhao, P. Effects of zinc fertilizer rate and application method on photosynthetic characteristics and grain yield of summer maize. J. Soil. Sci. Plant. Nut. 2016, 16, 550–562. [Google Scholar] [CrossRef]
- Khaliq, A.; Ali, S.; Hameed, A.; Farooq, M.A.; Farid, M.; Shakoor, M.B.; Mahmood, K.; Rizwan, M. Silicon alleviates nickel toxicity in cotton seedlings through enhancing growth, photosynthesis, and suppressing Ni uptake and oxidative stress. Arch. Agron. Soil. Sci. 2016, 62, 633–647. [Google Scholar] [CrossRef]
- Khoshbakht, D.; Asghari, M.R.; Haghighi, M. Influence of foliar application of polyamines on growth, gas-exchange characteristics, and chlorophyll fluorescence in Bakraii citrus under saline conditions. Photosynthetica 2018, 56, 731–742. [Google Scholar] [CrossRef]
- Wu, K.; Li, J.X.; Luo, J.P.; Liu, Y.K.; Song, Y.C.; Liu, N.L.; Rafiq, M.T.; Li, T.Q. Effects of elevated CO2, and endophytic bacterium on photosynthetic characteristics and cadmium accumulation in, Sedum alfredii. Sci. Total. Environ. 2018, 643, 357–366. [Google Scholar] [CrossRef] [PubMed]
- Zornoza, R.; Guerrero, C.; Mataix-Solera, J.; Arcenegui, V.; García-Orenes, F.; Mataix-Beneyto, J. Assessing air-drying and rewetting pre-treatment effect on some soil enzyme activities under Mediterranean conditions. Soil. Biol. Biochem. 2006, 38, 2125–2134. [Google Scholar] [CrossRef]
- Hu, M.J.; Jiang, L.C.; Li, S.Z.; Zhou, L.F.; Tong, C. Effects of different mulching and drip irrigation patterns on photosynthetic characteristics of maize leaves in the hetao irrigation district, Inner Mongolia, China. J. Appl. Ecol. 2017, 12, 144–153. [Google Scholar]
- Yuan, X.K.; Yang, Z.Q.; Li, Y.X.; Liu, Q.; Han, W. Effects of different levels of water stress on leaf photosynthetic characteristics and antioxidant enzyme activities of greenhouse tomato. Photosynthetica 2016, 54, 28–39. [Google Scholar] [CrossRef]
- Liu, E.K.; Mei, X.R.; Yan, C.R.; Gong, D.Z.; Zhang, Y.Q. Effects of water stress on photosynthetic characteristics, dry matter translocation and WUE in two winter wheat genotypes. Agric. Water. Manag. 2016, 167, 75–85. [Google Scholar] [CrossRef]
- Wang, C.Y.; Deng, Y.; Zhao, L.; Zhang, L.G.; Wang, J.; Guo, H.X.; Wang, L.J. Effects of different film-mulching modes on photosynthetic characters and yield of maize. Chin. Agric. Sci. Bull. 2015, 31, 62–67. [Google Scholar]
- Ndubuisi, M.C. Physical properties of an ultisol under plastic film and no-mulches and their effect on the yield of maize. Am. J. Sci. 2009, 5, 25–30. [Google Scholar]
- Wang, J.; Niu, W.; Xu, J.; Li, Y. Effects of drip irrigation under plastic film on muskmelon soil environment and yield in greenhouse. Trans. Chin. Soc. Agric. Eng. 2016, 32, 232–241. [Google Scholar]
- Wang, J.W.; Niu, W.Q.; Dyck, M.; Zhang, M.Z.; Li, Y. Drip irrigation with film covering improves soil enzymes and muskmelon growth in the greenhouse. Soil Res. 2018, 56, 59–70. [Google Scholar] [CrossRef]
- Ðukic, D.; Mandic, L.; Veskovic, S.; Kapor, I. Correlation between fertilizer type and rate, urease activity and total nitrogen and ammonium nitrogen contents in a Chernozem soil under wheat. Agric. For. 2015, 61, 43–51. [Google Scholar]
- Su, W.; Qu, Y.; Feng, B.L.; Chai, Y. Photosynthesis characteristics and yield of broomcorn millet under film mulching on ridge-furrow for harvesting rainwater model in semi-arid region of northern shaanxi. Trans. Chin. Soc. Agric. Eng. 2014, 30, 137–145. [Google Scholar]
- Wang, Z.D.; Zhao, J.; Mao, S.H.; Wu, J.X. Study on the regional advantage layout of chinese muskmelon industry. Chin. J. Agric. Resour. Reg. Plan. 2014, 35, 128–133. [Google Scholar]
- Xue, L.; Ma, Z.M.; Du, S.P.; Feng, S.J.; Ran, S.B. Effects of application of nitrogen on melon yield, nitrogen balance and soil nitrogen accumulation under plastic mulching with drip Irrigation. Sci. Agric. Sin. 2019, 52, 690–700. [Google Scholar]
- Zhao, M.; Shi, Y.; Chen, X.; Ma, J. Soil nitrogen accumulation in different ages of vegetable greenhouses. Procedia Environ. Sci. 2011, 8, 21–25. [Google Scholar] [CrossRef]
- Mu, L.; Fang, L. Effects of different mulching practices on the photosynthetic characteristics of hot pepper (Capsicum annuum L.) in a greenhouse in Northwest China. Acta Agric. Scand. 2015, 65, 735–746. [Google Scholar]
- Utkhede, A.D.; Gadge, S.B.; Gorantiwar, S.D. Response of Muskmelon to Spectral Modification of Shading Nets under Different Drip Irrigation Regimes. Int. J. Curr. Microbiol. App. Sci. 2019, 8, 419–428. [Google Scholar] [CrossRef]
- Sharma, S.P.; Leskovar, D.I.; Volder, A.; Crosby, K.M.; Ibrahim, A.M.H. Root distribution patterns of reticulatus and inodorus melon (Cucumis melo L.) under subsurface deficit irrigation. Irrig. Sci. 2018, 36, 301–317. [Google Scholar] [CrossRef]
- Farquhar, G.D.; Sharkey, T.D. Stomatal conductance and photosynthesis. Annu. Rev. Plant Physiol. 1982, 33, 317–345. [Google Scholar] [CrossRef]
- Chen, G.Y.; Chen, J.; Xu, D.Q. Thinking about the relationship between net photosynthetic rate and intercellular CO2 concentration. Plant Physiol. Commun. 2010, 46, 64–66. [Google Scholar]
- Li, S.X.; Liu, W.G.; Gao, Y.; Liu, T.; Zhou, T.; Du, Y.L.; Yang, H.; Zhang, H.; Liu, J.D.; Yang, W.J. Effects of silicon on plant growth and photosynthetic characteristics of soybean seedlings under artificial shade stress. Sci. Agric. Sin. 2018, 51, 45–54. [Google Scholar]
- Wang, J.W.; Li, Y.; Niu, W.Q. Deficit Alternate Drip Irrigation Increased Root-Soil-Plant Interaction, Tomato Yield, and Quality. Int. J. Environ. Res. Public Health. 2020, 17, 781. [Google Scholar] [CrossRef]
- Wang, J.W.; Niu, W.Q.; Guo, L.L.; Li, Y. Drip irrigation with film mulch improves soil alkaline phosphatase and phosphorus uptake. Agric. Water. Manag. 2018, 201, 258–267. [Google Scholar] [CrossRef]
- Zhang, Z.J.; Tang, L.; Li, Y.J.; Liu, X.J. Effect of drip-irrigation pipe arrangement on distribution of water and salt in soil as well as the yield of asparagus lettuce under raised-bed cultivation. J. Irrig. Drain. 2018, 37, 23–27. [Google Scholar]
- Shang, J.; Liu, Y.G.; Jiang, W.; Zhang, H.S.; Lin, Q.; Zhao, C.X.; Zhang, Y.M. Effects of different distance to drip tape on dry matter accumulation, distribution and translocation in winter wheat. Chin. Agric. Sci. Bull. 2014, 30, 182–188. [Google Scholar]
- Zhang, X.; Niu, W.Q.; Jia, Z.X. Influences of rhizosphere aeration supplies on soil enzyme activities for potted tomato after irrigation. J. Nat. Resour. 2012, 27, 1296–1303. [Google Scholar]
- Maleva, M.; Borisova, G.; Chukina, N.; Nekrasova, G.; Prasad, M.N.V. Influence of exogenous urea on photosynthetic pigments, 14CO2 uptake, and urease activity in Elodea densa-environmental implications. Environ. Sci. Pollut. R. 2013, 20, 6172–6177. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, A.; Mhatre, A.; Pandit, R.; Lali, A.M. Synergistic biorefinery of Scenedesmus obliquus and Ulva lactuca in poultry manure towards sustainable bioproduct generation. Bioresour. Technol. 2020, 297, 122462. [Google Scholar] [CrossRef]
- Peña-Rojas, K.; Aranda, X.; Fleck, I. Stomatal limitation to CO2 assimilation and down-regulation of photosynthesis in Quercus ilex resprouts in response to slowly imposed drought. Tree Physiol. 2004, 24, 813–822. [Google Scholar] [CrossRef]
- Time, A.; Garrido, M.; Acevedo, E. Water relations and growth response to drought stress of Prosopis tamarugo Phil. A review. J. Soil. Sci. Plant. Nut. 2018, 18, 329–343. [Google Scholar] [CrossRef]
- Mao, W.G.; Wu, Z.; Huang, J.; Guo, S.R. Effects of moisture and light intensity on ecophysiological characteristics of muskmelon seedlings. Chin. J. Appl. Ecol. 2007, 18, 2475–2479. [Google Scholar]
- Zeng, C.Z.; Bie, Z.L.; Yuan, B.Z. Determination of optimum irrigation water amount for drip-irrigated muskmelon (Cucumis melo L.) in plastic greenhouse. Agric. Water. Manag. 2009, 96, 595–602. [Google Scholar] [CrossRef]
- Kanai, S.; Adu-Gymfi, J.; Lei, K.; Ito, J.; Ohkura, K.; Moghaieb, R.E.A.; EI-Shemy, H.; Mohapatra, P.K.; Saneoka, H.; Fujita, K. N-deficiency damps out circadian rhythmic changes of stem diameter dynamics in tomato plant. Plant. Sci. 2008, 174, 183–191. [Google Scholar] [CrossRef]
- Sengupta, D.; Guha, A.; Reddy, A.R. Interdependence of plant water status with photosynthetic performance and root defense responses in Vigna radiata (L.) Wilczek under progressive drought stress and recovery. J. Photochem. Photobiol. B Biol. 2013, 127, 170–181. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Factors | Levels | Experimental Setting | Materials |
|---|---|---|---|
| Plastic covering method | Full plastic covering (F) | Both rows and inter-row were covered. | White high-pressure low-density polyethylene plastic, with 0.014 mm thick. |
| Half plastic covering (H) | Rows were covered. | ||
| No plastic covering (N) | No rows and inter-row were covered. | ||
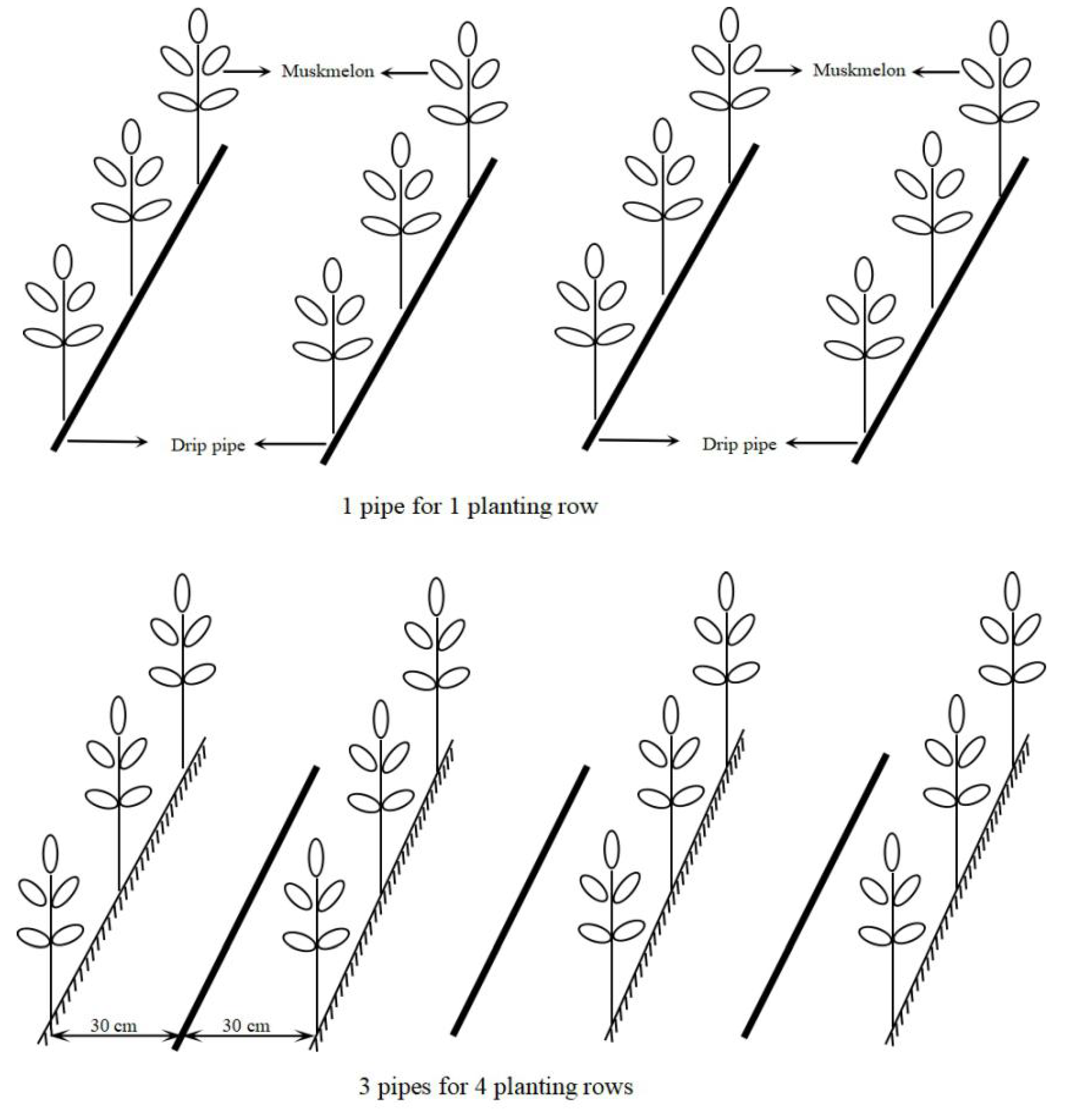
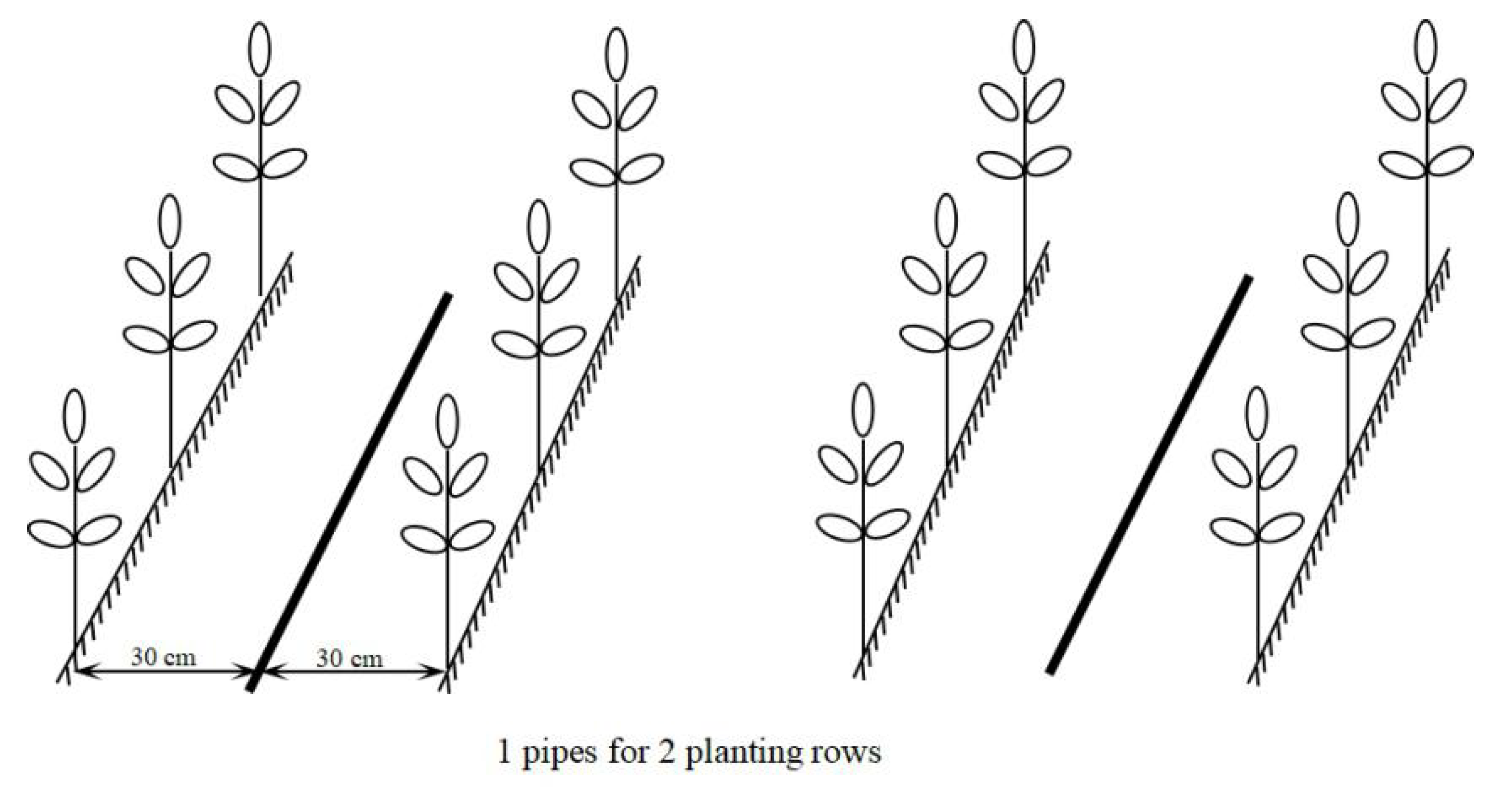
| Drip irrigation pipe density (Figure 1) | 1 pipe per 1row (T1) | One pipe was laid for each row. | The embedded inner inlay flat drip irrigation pipes, with 16 mm diameter, 0.3 mm wall thickness, 30 cm emitter distance, 0.1 Mpa working pressure, and 1.2 L·h-1 flow. |
| 3 pipes for 4 rows (T3/4) | Three pipes were laid in three spaces between four rows of a group. | ||
| 1 pipe for 2 rows (T1/2) | One pipe was laid between two prows. | ||
| Lower limit of irrigation | 60% field water capacity (F) (L60) | The upper limit was 70% F. | Irrigation water was derived from local agricultural water. |
| 70% field water capacity (L70) | The upper limit was 80% F. | ||
| 80% field water capacity (L80) | The upper limit was 90% F. |
| Growth Periods | Single Experimental Factors | |||
|---|---|---|---|---|
| Flowering period (FP) | P * | F: 13.37 b | H: 17.32 a | N: 13.82 b |
| T * | T1: 14.03 b | T3/4: 15.80 a | T1/2: 14.68 ab | |
| Lns | L60: 14.50 a | L70: 14.79 a | L80: 15.23 a | |
| Fruit swelling period (FSP) | P * | F: 19.53 b | H: 21.39 a | N: 19.40 b |
| T * | T1: 19.12 b | T3/4: 18.02 c | T1/2: 22.99 a | |
| L * | L60: 16.00 c | L70: 20.78 b | L80: 23.14 a | |
| Mature period (MP) | P * | F: 21.35 b | H: 23.87 a | N: 23.42 a |
| T * | T1: 24.41 a | T3/4: 21.09 b | T1/2: 23.59 a | |
| Lns | L60: 22.77 a | L70: 23.01 a | L80: 22.98 a | |
| Growth Periods | Single Experimental Factors | |||
|---|---|---|---|---|
| Flowering period (FP) | Pns | F: 0.21 a | H: 0.23 a | N: 0.22 a |
| T * | T1: 0.17 c | T3/4: 0.22 b | T1/2: 0.26 a | |
| L * | L60: 0.23 a | L70: 0.19 b | L80: 0.23 a | |
| Fruit swelling period (FSP) | P * | F: 0.27 c | H: 0.31 a | N: 0.29 b |
| T * | T1: 0.30 a | T3/4: 0.26 c | T1/2: 0.29 b | |
| L * | L60: 0.26 c | L70: 0.37 a | L80: 0.28 b | |
| Mature period (MP) | Pns | F: 0.40 a | H: 0.38 a | N: 0.41 a |
| T * | T1: 0.37 c | T3/4: 0.43 a | T1/2: 0.40 b | |
| L * | L60: 0.39 b | L70: 0.38 b | L80: 0.42 a | |
| Fruit Fresh Biomass | Plant Fresh Biomass | Root Fresh Biomass | Total Fresh Biomass | Pn in FP | Pn in FSP | Pn in MP | Ci in MP | |
|---|---|---|---|---|---|---|---|---|
| Fruit fresh biomass | 1.00 | 0.20 | 0.78 * | 0.87 ** | 0.75 * | 0.55 | −0.03 | 0.00 |
| Plant fresh biomass | 1.00 | 0.43 | 0.63 | −0.18 | 0.25 | 0.33 | −0.10 | |
| Root fresh biomass | 1.00 | 0.87 ** | 0.73 * | 0.25 | 0.03 | −0.17 | ||
| Total fresh biomass | 1.00 | 0.57 | 0.55 | 0.25 | −0.13 | |||
| Pn in FP | 1.00 | 0.32 | 0.02 | −0.17 | ||||
| Pn in FSP | 1.00 | 0.32 | 0.40 | |||||
| Pn in MP | 1.00 | −0.55 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Niu, W.; Li, Y. Effects of Drip Irrigation with Plastic on Photosynthetic Characteristics and Biomass Distribution of Muskmelon. Agriculture 2020, 10, 84. https://doi.org/10.3390/agriculture10030084
Wang J, Niu W, Li Y. Effects of Drip Irrigation with Plastic on Photosynthetic Characteristics and Biomass Distribution of Muskmelon. Agriculture. 2020; 10(3):84. https://doi.org/10.3390/agriculture10030084
Chicago/Turabian StyleWang, Jingwei, Wenquan Niu, and Yuan Li. 2020. "Effects of Drip Irrigation with Plastic on Photosynthetic Characteristics and Biomass Distribution of Muskmelon" Agriculture 10, no. 3: 84. https://doi.org/10.3390/agriculture10030084
APA StyleWang, J., Niu, W., & Li, Y. (2020). Effects of Drip Irrigation with Plastic on Photosynthetic Characteristics and Biomass Distribution of Muskmelon. Agriculture, 10(3), 84. https://doi.org/10.3390/agriculture10030084