
The Expression Level of SOX Family Transcription Factors’ mRNA as a Diagnostic Marker for Osteoarthritis

 , , , , , , , and
, , , , , , , and
Abstract
1. Introduction
2. Materials and Methods
2.1. Clinical Characteristics of OA Patients
2.2. Biological Material
2.3. RNA Isolation and Qualitative/Quantitative RNA Evaluation
2.4. Relative Expression Level (RQ) of Studied Genes
2.5. Statistical Analysis
2.6. Microarray Data Processing and Analysis
2.7. Construction of Protein–Protein Interaction (PPI) Network
3. Results
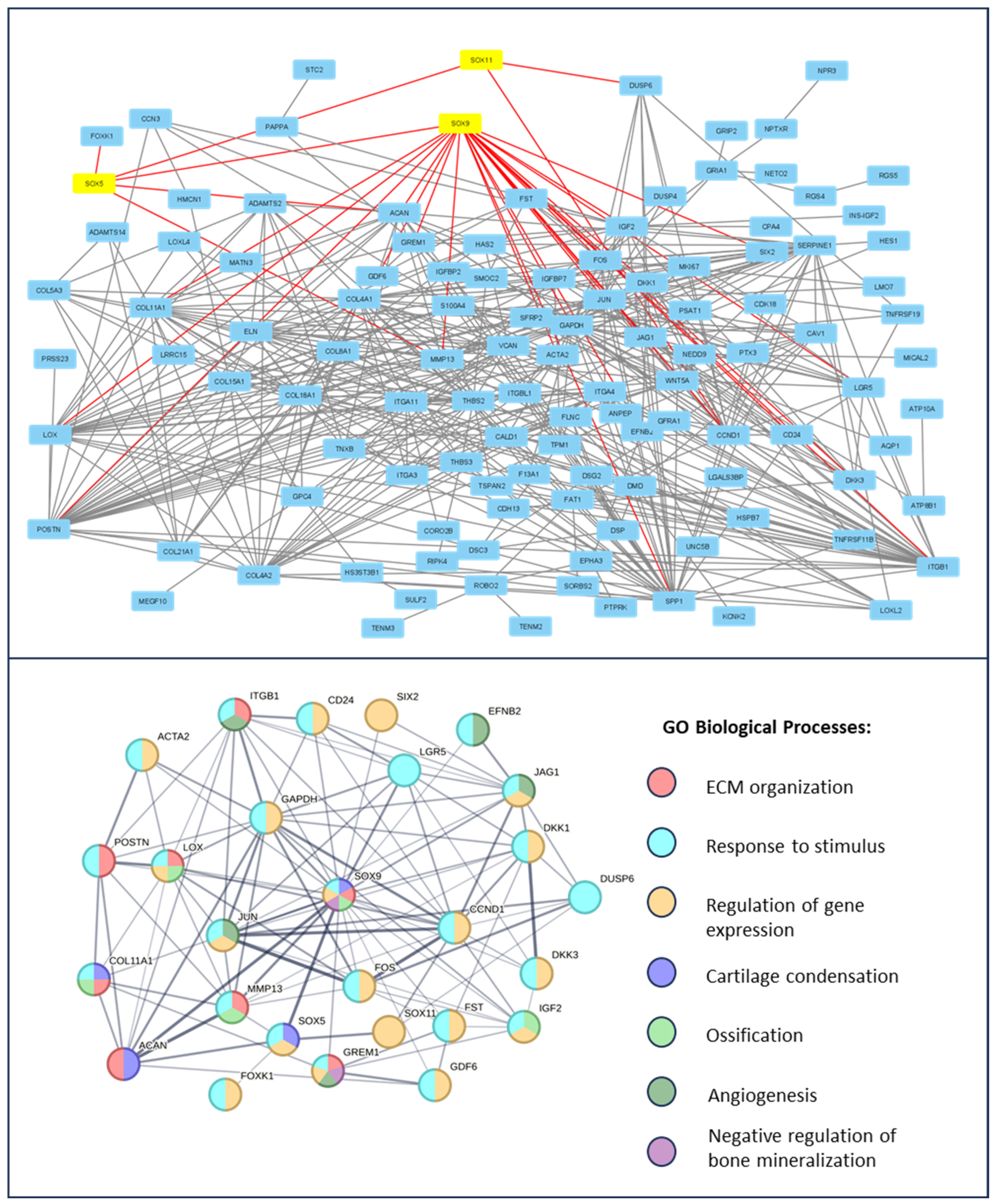
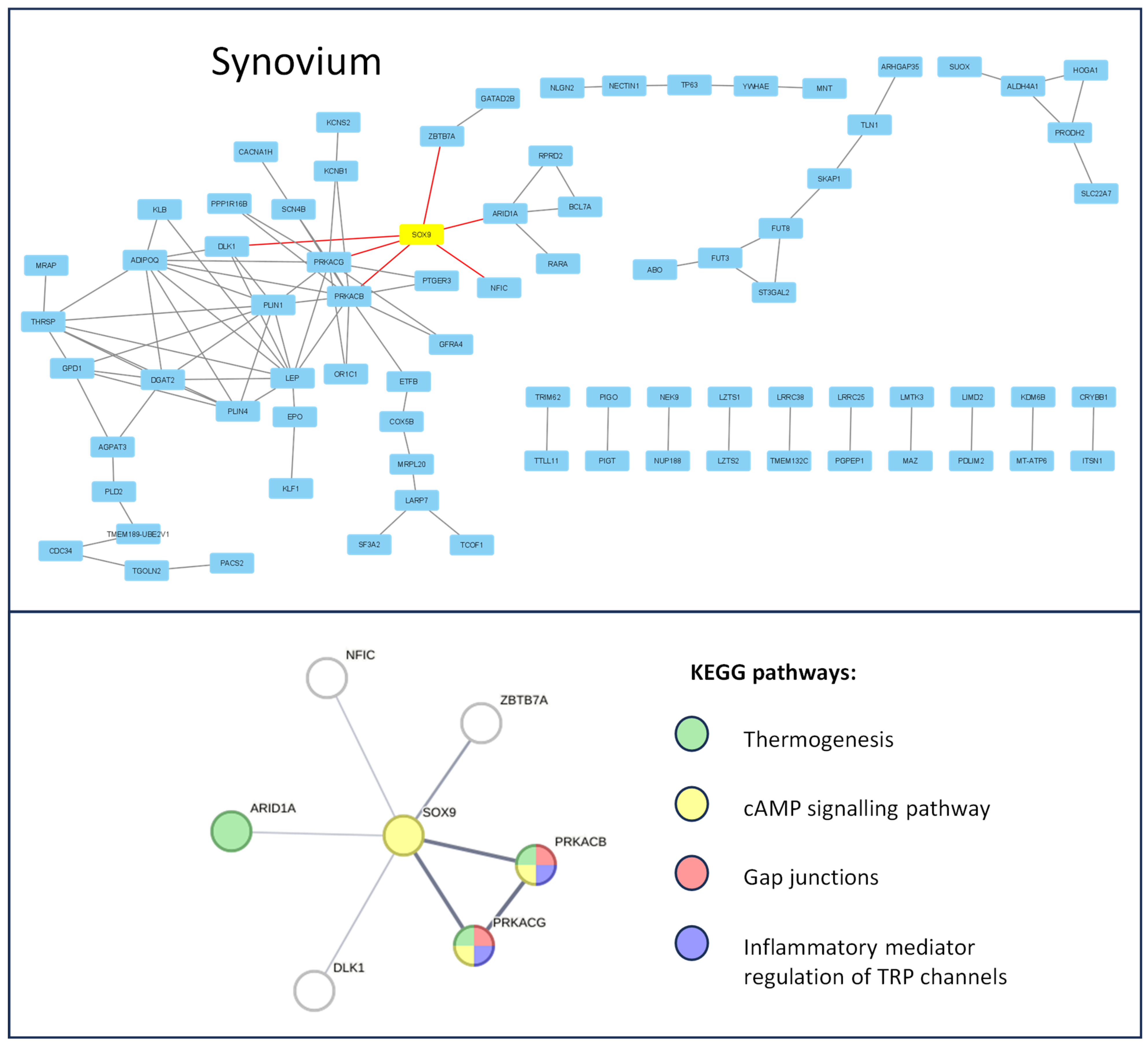
3.1. Protein–Protein Interaction in Chndrocytes and Synovium
3.2. Relative Expression Level (RQ) of SOX5, SOX9, and SOX11 mRNA in Articular Cartilage with Subchondral Bone in OA Patients vs. Control Tissue
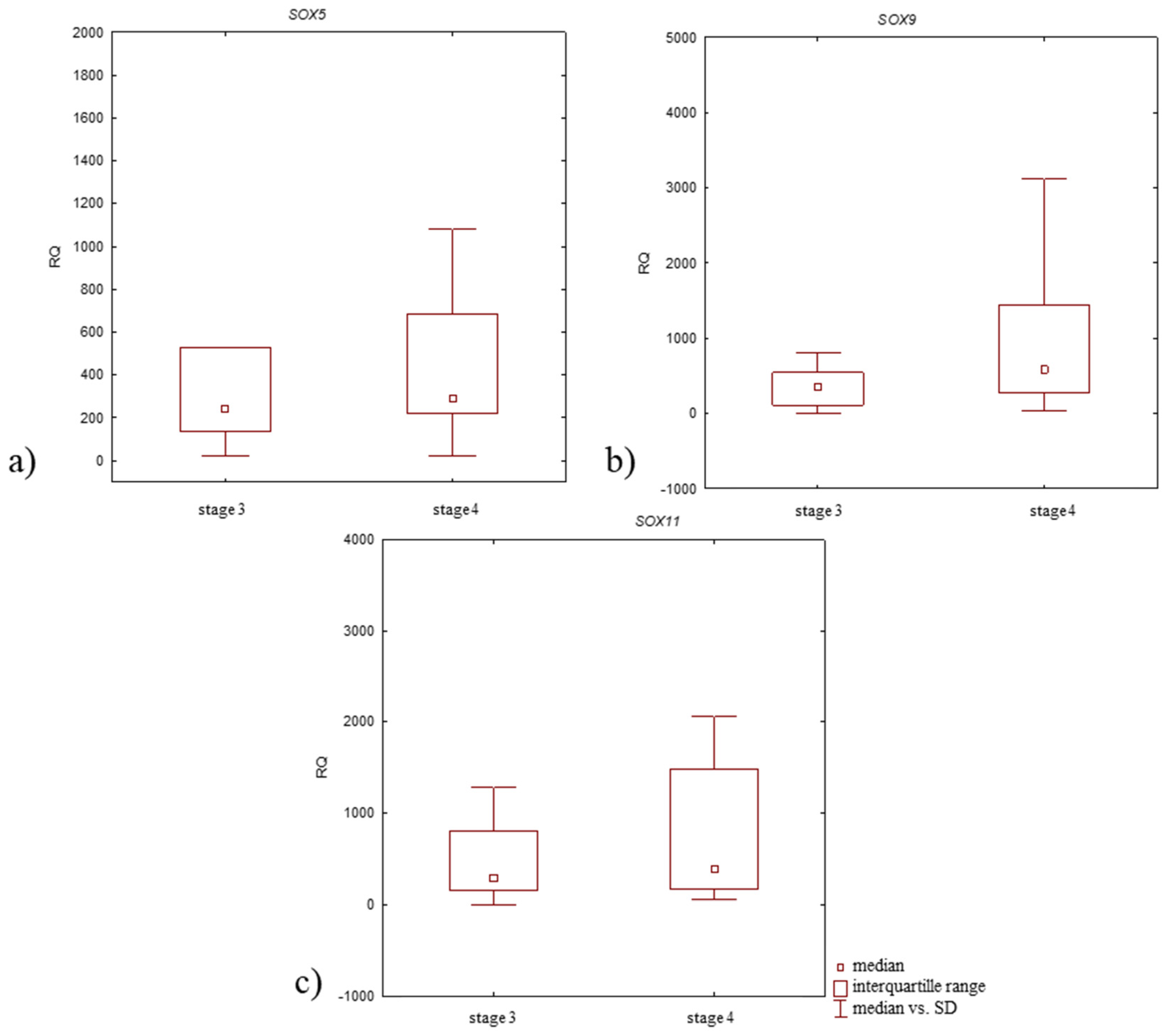
3.3. Relative Expression Level (RQ) of Studied Genes in Articular Cartilage with Subchondral Bone of OA Patients According to Clinical Characteristics of Patients
3.4. Relative Expression Level (RQ) of the SOX5, SOX9, and SOX11 mRNA in Synovium of OA Patients vs. Control Tissue
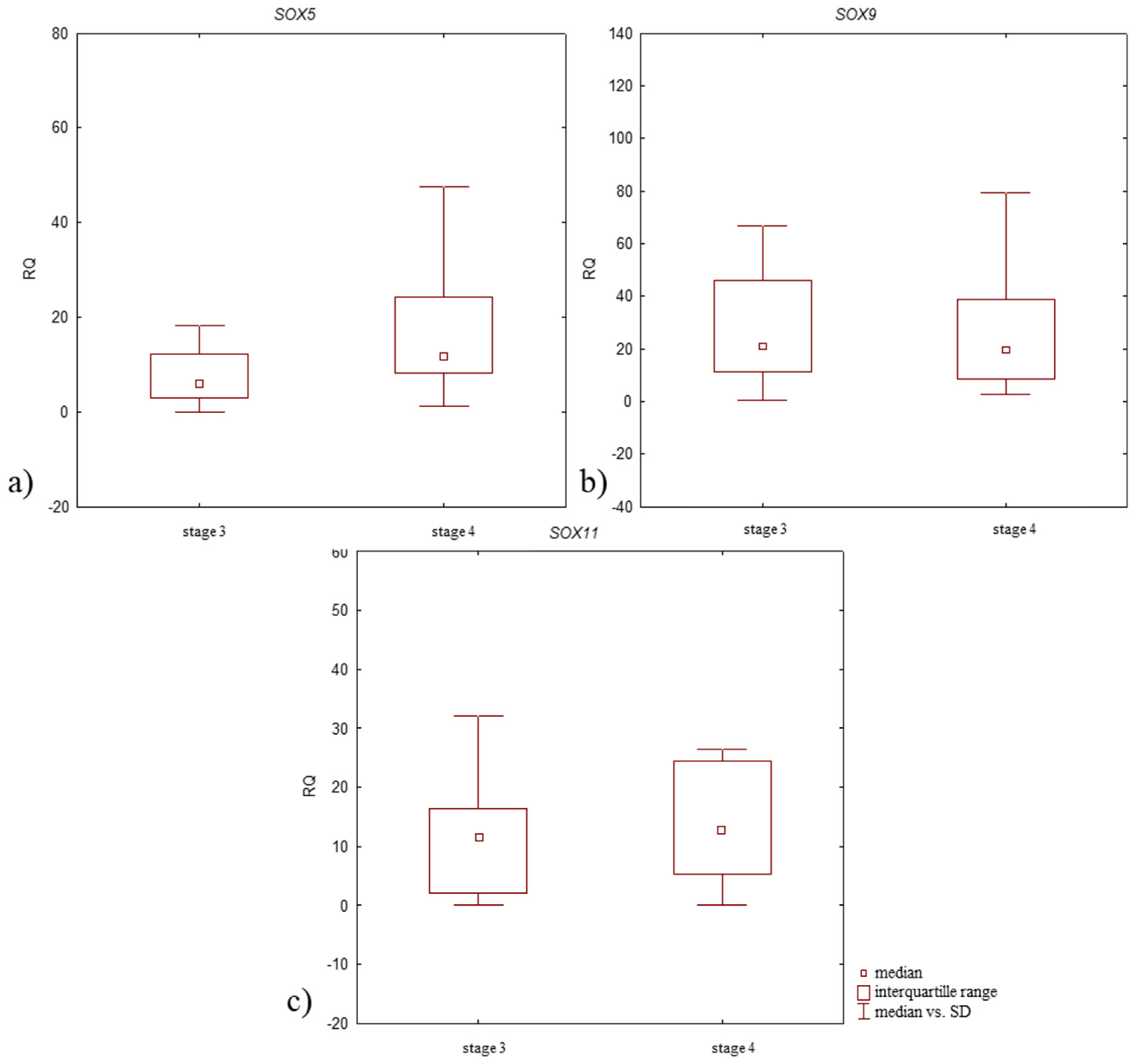
3.5. Relative Expression Level (RQ) of Studied Genes in Synovium of OA Patients According to the Clinical Characteristics of the Patients and Pathological Tissue Examination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Long, H.; Liu, Q.; Yin, H.; Wang, K.; Diao, N.; Zhang, Y.; Lin, J.; Guo, A. Prevalence Trends of Site-Specific Osteoarthritis From 1990 to 2019: Findings From the Global Burden of Disease Study 2019. Arthritis Rheumatol. 2022, 74, 1172–1183. [Google Scholar] [CrossRef] [PubMed]
- GBD 2015 Disease and Injury Incidence and Prevalence Collaborators. Global, regional, and national incidence, prevalence, and years lived with disability for 310 diseases and injuries, 1990-2015: A systematic analysis for the Global Burden of Disease Study 2015. Lancet 2016, 388, 1545–1602. [Google Scholar] [CrossRef]
- Palazzo, C.; Nguyen, C.; Lefevre-Colau, M.-M.; Rannou, F.; Poiraudeau, S. Risk factors and burden of osteoarthritis. Ann. Phys. Rehabil. Med. 2016, 59, 134–138. [Google Scholar] [CrossRef]
- Katz, J.N.; Arant, K.R.; Loeser, R.F. Diagnosis and Treatment of Hip and Knee Osteoarthritis: A Review. JAMA 2021, 325, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Abramoff, B.; Caldera, F.E. Osteoarthritis: Pathology, Diagnosis, and Treatment Options. Med. Clin. N. Am. 2020, 104, 293–311. [Google Scholar] [CrossRef]
- Steinhaus, M.E.; Christ, A.B.; Cross, M.B. Total Knee Arthroplasty for Knee Osteoarthritis: Support for a Foregone Conclusion? HSS J. 2017, 13, 207–210. [Google Scholar] [CrossRef] [PubMed]
- Bruyère, O.; Cooper, C.; Arden, N.; Branco, J.; Brandi, M.L.; Herrero-Beaumont, G.; Berenbaum, F.; Dennison, E.; Devogelaer, J.-P.; Hochberg, M.; et al. Can we identify patients with high risk of osteoarthritis progression who will respond to treatment? A focus on epidemiology and phenotype of osteoarthritis. Drugs Aging 2015, 32, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Yao, Q.; Wu, X.; Tao, C.; Gong, W.; Chen, M.; Qu, M.; Zhong, Y.; He, T.; Chen, S.; Xiao, G. Osteoarthritis: Pathogenic signaling pathways and therapeutic targets. Signal Transduct. Target. Ther. 2023, 8, 1–31. [Google Scholar] [CrossRef] [PubMed]
- Grässel, S.; Zaucke, F.; Madry, H. Osteoarthritis: Novel Molecular Mechanisms Increase Our Understanding of the Disease Pathology. J. Clin. Med. 2021, 10, 1938. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H. Control of chondrogenesis by the transcription factor Sox9. Mod. Rheumatol. 2008, 18, 213–219. [Google Scholar] [CrossRef] [PubMed]
- Akiyama, H.; Chaboissier, M.-C.; Martin, J.F.; Schedl, A.; de Crombrugghe, B. The transcription factor Sox9 has essential roles in successive steps of the chondrocyte differentiation pathway and is required for expression of Sox5 and Sox6. Genes. Dev. 2002, 16, 2813–2828. [Google Scholar] [CrossRef] [PubMed]
- Neefjes, M.; van Caam, A.P.M.; van der Kraan, P.M. Transcription Factors in Cartilage Homeostasis and Osteoarthritis. Biology 2020, 9, 290. [Google Scholar] [CrossRef] [PubMed]
- Bullejos, M.; de la Guardia, R.D.; Barragán, M.J.L.; Sánchez, A. HMG-box sequences from microbats homologous to the human SOX30 HMG-box*. Genetica 2000, 110, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, Y.; Kondoh, H. Sox proteins: Regulators of cell fate specification and differentiation. Development 2013, 140, 4129–4144. [Google Scholar] [CrossRef]
- Bowles, J.; Schepers, G.; Koopman, P. Phylogeny of the SOX family of developmental transcription factors based on sequence and structural indicators. Dev. Biol. 2000, 227, 239–255. [Google Scholar] [CrossRef]
- Harley, V.R.; Lovell-Badge, R.; Goodfellow, P.N. Definition of a consensus DNA binding site for SRY. Nucleic Acids Res. 1994, 22, 1500–1501. [Google Scholar] [CrossRef] [PubMed]
- Kamachi, Y.; Uchikawa, M.; Kondoh, H. Pairing SOX off: With partners in the regulation of embryonic development. Trends Genet. 2000, 16, 182–187. [Google Scholar] [CrossRef]
- Stevanovic, M.; Drakulic, D.; Lazic, A.; Ninkovic, D.S.; Schwirtlich, M.; Mojsin, M. SOX Transcription Factors as Important Regulators of Neuronal and Glial Differentiation During Nervous System Development and Adult Neurogenesis. Front. Mol. Neurosci. 2021, 14, 654031. [Google Scholar] [CrossRef] [PubMed]
- Ming, Z.; Vining, B.; Bagheri-Fam, S.; Harley, V. SOX9 in organogenesis: Shared and unique transcriptional functions. Cell Mol. Life Sci. 2022, 79, 522. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.; Huang, W.; Whitworth, D.J.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; de Crombrugghe, B. Haploinsufficiency of Sox9 results in defective cartilage primordia and premature skeletal mineralization. Proc. Natl. Acad. Sci. USA 2001, 98, 6698–6703. [Google Scholar] [CrossRef] [PubMed]
- Lefebvre, V.; Huang, W.; Harley, V.R.; Goodfellow, P.N.; de Crombrugghe, B. SOX9 is a potent activator of the chondrocyte-specific enhancer of the pro alpha1(II) collagen gene. Mol. Cell Biol. 1997, 17, 2336–2346. [Google Scholar] [CrossRef] [PubMed]
- Oh, C.; Lu, Y.; Liang, S.; Mori-Akiyama, Y.; Chen, D.; de Crombrugghe, B.; Yasuda, H. SOX9 Regulates Multiple Genes in Chondrocytes, Including Genes Encoding ECM Proteins, ECM Modification Enzymes, Receptors, and Transporters. PLoS ONE 2014, 9, e107577. [Google Scholar] [CrossRef]
- Zhou, G.; Zheng, Q.; Engin, F.; Munivez, E.; Chen, Y.; Sebald, E.; Krakow, D.; Lee, B. Dominance of SOX9 function over RUNX2 during skeletogenesis. Proc. Natl. Acad. Sci. USA 2006, 103, 19004–19009. [Google Scholar] [CrossRef]
- Hattori, T.; Müller, C.; Gebhard, S.; Bauer, E.; Pausch, F.; Schlund, B.; Bösl, M.R.; Hess, A.; Surmann-Schmitt, C.; von der Mark, H.; et al. SOX9 is a major negative regulator of cartilage vascularization, bone marrow formation and endochondral ossification. Development 2010, 137, 901–911. [Google Scholar] [CrossRef]
- Zwickl, H.; Niculescu-Morzsa, E.; Halbwirth, F.; Bauer, C.; Jeyakumar, V.; Reutterer, A.; Berger, M.; Nehrer, S. Correlation Analysis of SOX9, -5, and -6 as well as COL2A1 and Aggrecan Gene Expression of Collagen I Implant-Derived and Osteoarthritic Chondrocytes. Cartilage 2016, 7, 185–192. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Yu, J.; Wang, Z.; Ni, C.; Xia, L.; Tang, T. SOX11 promotes osteoarthritis through induction of TNF-α. Pathol. Res. Pract. 2019, 215, 152442. [Google Scholar] [CrossRef] [PubMed]
- Pucci, B.; Adams, C.S.; Fertala, J.; Snyder, B.C.; Mansfield, K.D.; Tafani, M.; Freeman, T.; Shapiro, I.M. Development of the terminally differentiated state sensitizes epiphyseal chondrocytes to apoptosis through caspase-3 activation. J. Cell. Physiol. 2007, 210, 609–615. [Google Scholar] [CrossRef]
- Kohn, M.D.; Sassoon, A.A.; Fernando, N.D. Classifications in Brief: Kellgren-Lawrence Classification of Osteoarthritis. Clin. Orthop. Relat. Res. 2016, 474, 1886–1893. [Google Scholar] [CrossRef]
- Lin, X.; Li, H. Obesity: Epidemiology, Pathophysiology, and Therapeutics. Front. Endocrinol. 2021, 12, 706978. [Google Scholar] [CrossRef]
- Kryczka, J.; Boncela, J. Characteristics of ABCC4 and ABCG2 High Expression Subpopulations in CRC—A New Opportunity to Predict Therapy Response. Cancers 2023, 15, 5623. [Google Scholar] [CrossRef] [PubMed]
- Kryczka, J.; Boncela, J. Integrated Bioinformatics Analysis of the Hub Genes Involved in Irinotecan Resistance in Colorectal Cancer. Biomedicines 2022, 10, 1720. [Google Scholar] [CrossRef] [PubMed]
- Felson, D.T.; Lawrence, R.C.; Dieppe, P.A.; Hirsch, R.; Helmick, C.G.; Jordan, J.M.; Kington, R.S.; Lane, N.E.; Nevitt, M.C.; Zhang, Y.; et al. Osteoarthritis: New insights. Part 1: The disease and its risk factors. Ann. Intern. Med. 2000, 133, 635–646. [Google Scholar] [CrossRef] [PubMed]
- Cui, A.; Li, H.; Wang, D.; Zhong, J.; Chen, Y.; Lu, H. Global, regional prevalence, incidence and risk factors of knee osteoarthritis in population-based studies. eClinicalMedicine 2020, 29–30, 100587. [Google Scholar] [CrossRef]
- Patel, J.; Chen, S.; Katzmeyer, T.; Pei, Y.A.; Pei, M. Sex-dependent variation in cartilage adaptation: From degeneration to regeneration. Biol. Sex. Differ. 2023, 14, 17. [Google Scholar] [CrossRef] [PubMed]
- Jarecki, J.; Małecka-Masalska, T.; Kosior-Jarecka, E.; Widuchowski, W.; Krasowski, P.; Gutbier, M.; Dobrzyński, M.; Blicharski, T. Concentration of Selected Metalloproteinases and Osteocalcin in the Serum and Synovial Fluid of Obese Women with Advanced Knee Osteoarthritis. Int. J. Environ. Res. Public Health 2022, 19, 3530. [Google Scholar] [CrossRef]
- Tummala, S.; Schiphof, D.; Byrjalsen, I.; Dam, E.B. Gender Differences in Knee Joint Congruity Quantified from MRI: A Validation Study with Data from Center for Clinical and Basic Research and Osteoarthritis Initiative. Cartilage 2018, 9, 38–45. [Google Scholar] [CrossRef]
- King, L.K.; March, L.; Anandacoomarasamy, A. Obesity & osteoarthritis. Indian. J. Med. Res. 2013, 138, 185. [Google Scholar] [PubMed]
- Coggon, D.; Reading, I.; Croft, P.; McLaren, M.; Barrett, D.; Cooper, C. Knee osteoarthritis and obesity. Int. J. Obes. Relat. Metab. Disord. 2001, 25, 622–627. [Google Scholar] [CrossRef] [PubMed]
- Yucesoy, B.; Charles, L.E.; Baker, B.; Burchfiel, C.M. Occupational and genetic risk factors for osteoarthritis: A review. Work 2015, 50, 261–273. [Google Scholar] [CrossRef] [PubMed]
- Driban, J.B.; Hootman, J.M.; Sitler, M.R.; Harris, K.P.; Cattano, N.M. Is Participation in Certain Sports Associated With Knee Osteoarthritis? A Systematic Review. J. Athl. Train. 2017, 52, 497–506. [Google Scholar] [CrossRef]
- Haag, J.; Gebhard, P.M.; Aigner, T. SOX gene expression in human osteoarthritic cartilage. Pathobiology 2008, 75, 195–199. [Google Scholar] [CrossRef]
- Aigner, T.; Gebhard, P.M.; Schmid, E.; Bau, B.; Harley, V.; Pöschl, E. SOX9 expression does not correlate with type II collagen expression in adult articular chondrocytes. Matrix Biol. 2003, 22, 363–372. [Google Scholar] [CrossRef] [PubMed]
- Mobasheri, A.; Thudium, C.S.; Bay-Jensen, A.-C.; Maleitzke, T.; Geissler, S.; Duda, G.N.; Winkler, T. Biomarkers for osteoarthritis: Current status and future prospects. Best. Pract. Res. Clin. Rheumatol. 2023, 37, 101852. [Google Scholar] [CrossRef] [PubMed]
- Braaten, J.A.; Banovetz, M.T.; DePhillipo, N.N.; Familiari, F.; Russo, R.; Kennedy, N.I.; LaPrade, R.F. Biomarkers for Osteoarthritis Diseases. Life 2022, 12, 1799. [Google Scholar] [CrossRef]
- Ahmed, E.A.; Alzahrani, A.M. SOXC Transcription Factors as Diagnostic Biomarkers and Therapeutic Targets for Arthritis. Int. J. Mol. Sci. 2023, 24, 4215. [Google Scholar] [CrossRef]
- Zhang, L.; Hu, J.; Athanasiou, K.A. The Role of Tissue Engineering in Articular Cartilage Repair and Regeneration. Crit. Rev. Biomed. Eng. 2009, 37, 1–57. [Google Scholar] [CrossRef] [PubMed]
- Peng, Z.; Sun, H.; Bunpetch, V.; Koh, Y.; Wen, Y.; Wu, D.; Ouyang, H. The regulation of cartilage extracellular matrix homeostasis in joint cartilage degeneration and regeneration. Biomaterials 2021, 268, 120555. [Google Scholar] [CrossRef]
- Fisch, K.M.; Gamini, R.; Alvarez-Garcia, O.; Akagi, R.; Saito, M.; Muramatsu, Y.; Sasho, T.; Koziol, J.A.; Su, A.I.; Lotz, M.K. Identification of transcription factors responsible for dysregulated networks in human osteoarthritis cartilage by global gene expression analysis. Osteoarthr. Cartil. 2018, 26, 1531–1538. [Google Scholar] [CrossRef] [PubMed]
- Mitsis, T.; Efthimiadou, A.; Bacopoulou, F.; Vlachakis, D.; Chrousos, G.P.; Eliopoulos, E. Transcription factors and evolution: An integral part of gene expression (Review). World Acad. Sci. J. 2020, 2, 3–8. [Google Scholar] [CrossRef]
- Pan, Y.; Tsai, C.-J.; Ma, B.; Nussinov, R. Mechanisms of transcription factor selectivity. Trends Genet. 2010, 26, 75–83. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Garcia, O.; Fisch, K.M.; Wineinger, N.E.; Akagi, R.; Saito, M.; Sasho, T.; Su, A.I.; Lotz, M.K. Increased DNA Methylation and Reduced Expression of Transcription Factors in Human Osteoarthritis Cartilage. Arthritis Rheumatol. 2016, 68, 1876–1886. [Google Scholar] [CrossRef] [PubMed]
- Rim, Y.A.; Nam, Y.; Ju, J.H. The Role of Chondrocyte Hypertrophy and Senescence in Osteoarthritis Initiation and Progression. Int. J. Mol. Sci. 2020, 21, 2358. [Google Scholar] [CrossRef] [PubMed]
- Nagy, A.; Kénesi, E.; Rentsendorj, O.; Molnár, A.; Szénási, T.; Sinkó, I.; Zvara, A.; Oommen, S.T.; Barta, E.; Puskás, L.G.; et al. Evolutionarily conserved, growth plate zone-specific regulation of the matrilin-1 promoter: L-Sox5/Sox6 and Nfi factors bound near TATA finely tune activation by Sox9. Mol. Cell Biol. 2011, 31, 686–699. [Google Scholar] [CrossRef]
- Song, H.; Park, K.-H. Regulation and function of SOX9 during cartilage development and regeneration. Semin. Cancer Biol. 2020, 67, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Herlofsen, S.R.; Høiby, T.; Cacchiarelli, D.; Zhang, X.; Mikkelsen, T.S.; Brinchmann, J.E. Brief Report: Importance of SOX8 for In Vitro Chondrogenic Differentiation of Human Mesenchymal Stromal Cells. Stem Cells 2014, 32, 1629–1635. [Google Scholar] [CrossRef] [PubMed]
- Smits, P.; Li, P.; Mandel, J.; Zhang, Z.; Deng, J.M.; Behringer, R.R.; de Crombrugghe, B. The Transcription Factors L-Sox5 and Sox6 Are Essential for Cartilage Formation. Developmental Cell 2001, 1, 277–290. [Google Scholar] [CrossRef]
- Park, S.B.; Seo, K.W.; So, A.Y.; Seo, M.S.; Yu, K.R.; Kang, S.K.; Kang, K.S. SOX2 has a crucial role in the lineage determination and proliferation of mesenchymal stem cells through Dickkopf-1 and c-MYC. Cell Death Differ. 2012, 19, 534–545. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Dong, S. The Signaling Pathways Involved in Chondrocyte Differentiation and Hypertrophic Differentiation. Stem Cells Int. 2016, 2016, e2470351. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.; Deng, J.M.; Zhang, Z.; Behringer, R.R.; De Crombrugghe, B. Sox9 is required for cartilage formation. Nat. Genet. 1999, 22, 85–89. [Google Scholar] [CrossRef]
- Ikeda, T.; Kawaguchi, H.; Kamekura, S.; Ogata, N.; Mori, Y.; Nakamura, K.; Ikegawa, S.; Chung, U. Distinct roles of Sox5, Sox6, and Sox9 in different stages of chondrogenic differentiation. J. Bone Min. Metab. 2005, 23, 337–340. [Google Scholar] [CrossRef] [PubMed]
- Csukasi, F.; Duran, I.; Zhang, W.; Martin, J.H.; Barad, M.; Bamshad, M.; Weis, M.A.; Eyre, D.; Krakow, D.; Cohn, D.H. Dominant-negative SOX9 mutations in campomelic dysplasia. Hum. Mutat. 2019, 40, 2344–2352. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.-F.; Lefebvre, V. The transcription factors SOX9 and SOX5/SOX6 cooperate genome-wide through super-enhancers to drive chondrogenesis. Nucleic Acids Res. 2015, 43, 8183–8203. [Google Scholar] [CrossRef] [PubMed]
- Frith, J.; Genever, P. Transcriptional Control of Mesenchymal Stem Cell Differentiation. Transfus. Med. Hemother 2008, 35, 216–227. [Google Scholar] [CrossRef] [PubMed]
- Dvir-Ginzberg, M.; Gagarina, V.; Lee, E.-J.; Hall, D.J. Regulation of Cartilage-specific Gene Expression in Human Chondrocytes by SirT1 and Nicotinamide Phosphoribosyltransferase. J. Biol. Chem. 2008, 283, 36300–36310. [Google Scholar] [CrossRef] [PubMed]
- Kypriotou, M.; Fossard-Demoor, M.; Chadjichristos, C.; Ghayor, C.; De Crombrugghe, B.; Pujol, J.-P.; Galéra, P. SOX9 Exerts a Bifunctional Effect on Type II Collagen Gene (COL2A1) Expression in Chondrocytes Depending on the Differentiation State. DNA Cell Biol. 2003, 22, 119–129. [Google Scholar] [CrossRef]
- Lefebvre, V.; Li, P.; de Crombrugghe, B. A new long form of Sox5 (L-Sox5), Sox6 and Sox9 are coexpressed in chondrogenesis and cooperatively activate the type II collagen gene. EMBO J. 1998, 17, 5718–5733. [Google Scholar] [CrossRef] [PubMed]
- Bridgewater, L.C.; Lefebvre, V.; de Crombrugghe, B. Chondrocyte-specific enhancer elements in the Col11a2 gene resemble the Col2a1 tissue-specific enhancer. J. Biol. Chem. 1998, 273, 14998–15006. [Google Scholar] [CrossRef] [PubMed]
- de Crombrugghe, B.; Lefebvre, V.; Behringer, R.R.; Bi, W.; Murakami, S.; Huang, W. Transcriptional mechanisms of chondrocyte differentiation. Matrix Biol. 2000, 19, 389–394. [Google Scholar] [CrossRef]
- Han, Y.; Lefebvre, V. L-Sox5 and Sox6 drive expression of the aggrecan gene in cartilage by securing binding of Sox9 to a far-upstream enhancer. Mol. Cell Biol. 2008, 28, 4999–5013. [Google Scholar] [CrossRef]
- Hu, G.; Codina, M.; Fisher, S. Multiple enhancers associated with ACAN suggest highly redundant transcriptional regulation in cartilage. Matrix Biol. 2012, 31, 328–337. [Google Scholar] [CrossRef]
- Zhang, Q.; Ji, Q.; Wang, X.; Kang, L.; Fu, Y.; Yin, Y.; Li, Z.; Liu, Y.; Xu, X.; Wang, Y. SOX9 is a regulator of ADAMTSs-induced cartilage degeneration at the early stage of human osteoarthritis. Osteoarthr. Cartil. 2015, 23, 2259–2268. [Google Scholar] [CrossRef]
- Dy, P.; Wang, W.; Bhattaram, P.; Wang, Q.; Wang, L.; Ballock, R.T.; Lefebvre, V. Sox9 directs hypertrophic maturation and blocks osteoblast differentiation of growth plate chondrocytes. Dev. Cell 2012, 22, 597–609. [Google Scholar] [CrossRef] [PubMed]
- Gu, J.; Lu, Y.; Li, F.; Qiao, L.; Wang, Q.; Li, N.; Borgia, J.A.; Deng, Y.; Lei, G.; Zheng, Q. Identification and characterization of the novel Col10a1 regulatory mechanism during chondrocyte hypertrophic differentiation. Cell Death Dis. 2014, 5, e1469. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Tsang, K.Y.; Tang, H.C.; Chan, D.; Cheah, K.S.E. Hypertrophic chondrocytes can become osteoblasts and osteocytes in endochondral bone formation. Proc. Natl. Acad. Sci. USA 2014, 111, 12097–12102. [Google Scholar] [CrossRef] [PubMed]
- Peacock, J.D.; Huk, D.J.; Ediriweera, H.N.; Lincoln, J. Sox9 Transcriptionally Represses Spp1 to Prevent Matrix Mineralization in Maturing Heart Valves and Chondrocytes. PLoS ONE 2011, 6, e26769. [Google Scholar] [CrossRef] [PubMed]
- Bai, R.-J.; Li, Y.-S.; Zhang, F.-J. Osteopontin, a bridge links osteoarthritis and osteoporosis. Front. Endocrinol. 2022, 13, 1012508. [Google Scholar] [CrossRef]
- Cheng, C.; Zhang, F.-J.; Tian, J.; Tu, M.; Xiong, Y.-L.; Luo, W.; Li, Y.-S.; Song, B.-B.; Gao, S.-G.; Lei, G.-H. Osteopontin inhibits HIF-2α mRNA expression in osteoarthritic chondrocytes. Exp. Ther. Med. 2015, 9, 2415–2419. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.-G.; Zeng, C.; Song, Y.; Tian, J.; Cheng, C.; Yang, T.; Li, H.; Zhang, F.-J.; Lei, G.-H. Effect of osteopontin on the mRNA expression of ADAMTS4 and ADAMTS5 in chondrocytes from patients with knee osteoarthritis. Exp. Ther. Med. 2015, 9, 1979–1983. [Google Scholar] [CrossRef] [PubMed]
- Attur, M.G.; Dave, M.N.; Stuchin, S.; Kowalski, A.J.; Steiner, G.; Abramson, S.B.; Denhardt, D.T.; Amin, A.R. Osteopontin: An intrinsic inhibitor of inflammation in cartilage. Arthritis Rheum. 2001, 44, 578–584. [Google Scholar] [CrossRef]
- Luo, W.; Lin, Z.; Yuan, Y.; Wu, Z.; Zhong, W.; Liu, Q. Osteopontin (OPN) alleviates the progression of osteoarthritis by promoting the anabolism of chondrocytes. Genes. Dis. 2022, 10, 1714–1725. [Google Scholar] [CrossRef]
- Liu, Q.; Zeng, H.; Yuan, Y.; Wang, Z.; Wu, Z.; Luo, W. Osteopontin inhibits osteoarthritis progression via the OPN/CD44/PI3K signal axis. Genes. Dis. 2022, 9, 128–139. [Google Scholar] [CrossRef]
- Rosenthal, A.K.; Gohr, C.M.; Uzuki, M.; Masuda, I. Osteopontin promotes pathologic mineralization in articular cartilage. Matrix Biol. 2007, 26, 96–105. [Google Scholar] [CrossRef]
- Li, Y.; Jiang, W.; Wang, H.; Deng, Z.; Zeng, C.; Tu, M.; Li, L.; Xiao, W.; Gao, S.; Luo, W.; et al. Osteopontin Promotes Expression of Matrix Metalloproteinase 13 through NF-?B Signaling in Osteoarthritis. BioMed Res. Int. 2016, 2016, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Honsawek, S.; Tanavalee, A.; Sakdinakiattikoon, M.; Chayanupatkul, M.; Yuktanandana, P. Correlation of plasma and synovial fluid osteopontin with disease severity in knee osteoarthritis. Clin. Biochem. 2009, 42, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.; Gao, S.; Lei, G. Association of osteopontin with osteoarthritis. Rheumatol. Int. 2014, 34, 1627–1631. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.G.; Li, K.H.; Zeng, K.B.; Tu, M.; Xu, M.; Lei, G.H. Elevated osteopontin level of synovial fluid and articular cartilage is associated with disease severity in knee osteoarthritis patients. Osteoarthr. Cartil. 2010, 18, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Ito, T.; Yadav, N.; Lee, J.; Furumatsu, T.; Yamashita, S.; Yoshida, K.; Taniguchi, N.; Hashimoto, M.; Tsuchiya, M.; Ozaki, T.; et al. Arginine methyltransferase CARM1/PRMT4 regulates endochondral ossification. BMC Dev. Biol. 2009, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Lin, W.; Xu, L.; Li, G. Molecular Insights Into Lysyl Oxidases in Cartilage Regeneration and Rejuvenation. Front. Bioeng. Biotechnol. 2020, 8, 359. [Google Scholar] [CrossRef]
- Alshenibr, W.; Tashkandi, M.M.; Alsaqer, S.F.; Alkheriji, Y.; Wise, A.; Fulzele, S.; Mehra, P.; Goldring, M.B.; Gerstenfeld, L.C.; Bais, M.V. Anabolic role of lysyl oxidase like-2 in cartilage of knee and temporomandibular joints with osteoarthritis. Arthritis Res. Ther. 2017, 19, 179. [Google Scholar] [CrossRef] [PubMed]
- Waldrep, K.M.; Rodgers, J.I.; Garrett, S.M.; Wolf, B.J.; Feghali-Bostwick, C.A. The Role of SOX9 in IGF-II-Mediated Pulmonary Fibrosis. Int. J. Mol. Sci. 2023, 24, 11234. [Google Scholar] [CrossRef] [PubMed]
- Uchimura, T.; Foote, A.T.; Smith, E.L.; Matzkin, E.G.; Zeng, L. Insulin-Like Growth Factor II (IGF-II) Inhibits IL-1β-Induced Cartilage Matrix Loss and Promotes Cartilage Integrity in Experimental Osteoarthritis. J. Cell Biochem. 2015, 116, 2858–2869. [Google Scholar] [CrossRef]
- Luo, S.; Shi, Q.; Li, W.; Wu, W.; Zha, Z. ITGB1 promotes the chondrogenic differentiation of human adipose-derived mesenchymal stem cells by activating the ERK signaling. J. Mol. Histol. 2020, 51, 729–739. [Google Scholar] [CrossRef]
- Xie, L.; Li, Z.; Chen, Z.; Li, M.; Tao, J. ITGB1 alleviates osteoarthritis by inhibiting cartilage inflammation and apoptosis via activating cAMP pathway. J. Orthop. Surg. Res. 2023, 18, 849. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-G.; Yu, S.-S.; Poo, H.; Chun, J.-S. c-Jun/activator protein-1 mediates interleukin-1beta-induced dedifferentiation but not cyclooxygenase-2 expression in articular chondrocytes. J. Biol. Chem. 2005, 280, 29780–29787. [Google Scholar] [CrossRef]
- He, X.; Ohba, S.; Hojo, H.; McMahon, A.P. AP-1 family members act with Sox9 to promote chondrocyte hypertrophy. Development 2016, 143, 3012–3023. [Google Scholar] [CrossRef]
- Ouyang, Y.; Wang, W.; Tu, B.; Zhu, Y.; Fan, C.; Li, Y. Overexpression of SOX9 alleviates the progression of human osteoarthritis in vitro and in vivo. Drug Des. Devel Ther. 2019, 13, 2833–2842. [Google Scholar] [CrossRef]
- Newman, S.; Ahmed, H.; Rehmatullah, N. Radiographic vs. MRI vs. arthroscopic assessment and grading of knee osteoarthritis—Are we using appropriate imaging? J. Exp. Orthop. 2022, 9, 2. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Liu, X.; Yang, Y.; Sun, Z.; Deng, S.; Jiang, Z.; Li, W.; Wu, F. NEAT1/miR-193a-3p/SOX5 axis regulates cartilage matrix degradation in human osteoarthritis. Cell Biol. Int. 2020, 44, 947–957. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Zhang, H.; Sun, J.; Gao, X.; Liu, C. CircSEC24A promotes IL-1β-induced apoptosis and inflammation in chondrocytes by regulating miR-142-5p/SOX5 axis. Biotechnol. Appl. Biochem. 2022, 69, 701–713. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, N.; Wu, X. Circular RNA_0003800 exacerbates IL-1β-induced chondrocyte injury via miR-197-3p/SOX5 axis. Int. Immunopharmacol. 2023, 115, 109643. [Google Scholar] [CrossRef]
- Fu, M.; Huang, G.; Zhang, Z.; Liu, J.; Zhang, Z.; Huang, Z.; Yu, B.; Meng, F. Expression profile of long noncoding RNAs in cartilage from knee osteoarthritis patients. Osteoarthr. Cartil. 2015, 23, 423–432. [Google Scholar] [CrossRef] [PubMed]
- Raud, B.; Gay, C.; Guiguet-Auclair, C.; Bonnin, A.; Gerbaud, L.; Pereira, B.; Duclos, M.; Boirie, Y.; Coudeyre, E. Level of obesity is directly associated with the clinical and functional consequences of knee osteoarthritis. Sci. Rep. 2020, 10, 3601. [Google Scholar] [CrossRef] [PubMed]
- Nedunchezhiyan, U.; Wu, X.; Prasadam, I. Obesity, Inflammation, and Immune System in Osteoarthritis. Front. Immunol. 2022, 13, 907750. [Google Scholar] [CrossRef] [PubMed]
- Losina, E.; Weinstein, A.M.; Reichmann, W.M.; Burbine, S.A.; Solomon, D.H.; Daigle, M.E.; Rome, B.N.; Chen, S.P.; Hunter, D.J.; Suter, L.G.; et al. Lifetime Risk and Age at Diagnosis of Symptomatic Knee Osteoarthritis in the US. Arthritis Care Res. 2013, 65, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Ilia, I.; Nitusca, D.; Marian, C. Adiponectin in Osteoarthritis: Pathophysiology, Relationship with Obesity and Presumptive Diagnostic Biomarker Potential. Diagnostics 2022, 12, 455. [Google Scholar] [CrossRef] [PubMed]
- Laurberg, T.B.; Frystyk, J.; Ellingsen, T.; Hansen, I.T.; Jørgensen, A.; Tarp, U.; Hetland, M.L.; Hørslev-Petersen, K.; Hornung, N.; Poulsen, J.H.; et al. Plasma adiponectin in patients with active, early, and chronic rheumatoid arthritis who are steroid- and disease-modifying antirheumatic drug-naive compared with patients with osteoarthritis and controls. J. Rheumatol. 2009, 36, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- de Boer, T.N.; van Spil, W.E.; Huisman, A.M.; Polak, A.A.; Bijlsma, J.W.J.; Lafeber, F.P.J.G.; Mastbergen, S.C. Serum adipokines in osteoarthritis; comparison with controls and relationship with local parameters of synovial inflammation and cartilage damage. Osteoarthr. Cartil. 2012, 20, 846–853. [Google Scholar] [CrossRef] [PubMed]
- Cuzdan Coskun, N.; Ay, S.; Evcik, F.D.; Oztuna, D. Adiponectin: Is it a biomarker for assessing the disease severity in knee osteoarthritis patients? Int. J. Rheum. Dis. 2017, 20, 1942–1949. [Google Scholar] [CrossRef]
- Francin, P.-J.; Abot, A.; Guillaume, C.; Moulin, D.; Bianchi, A.; Gegout-Pottie, P.; Jouzeau, J.-Y.; Mainard, D.; Presle, N. Association between adiponectin and cartilage degradation in human osteoarthritis. Osteoarthr. Cartil. 2014, 22, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Pérez, R.F.; Tejedor, J.R.; Santamarina-Ojeda, P.; Martínez, V.L.; Urdinguio, R.G.; Villamañán, L.; Candiota, A.P.; Sarró, N.V.; Barradas, M.; Fernandez-Marcos, P.J.; et al. Conservation of Aging and Cancer Epigenetic Signatures across Human and Mouse. Mol. Biol. Evol. 2021, 38, 3415–3435. [Google Scholar] [CrossRef]
- Senapati, P.; Miyano, M.; Sayaman, R.W.; Basam, M.; Trac, C.; Leung, A.; LaBarge, M.A.; Schones, D.E. Aging leads to DNA methylation alterations associated with loss of lineage fidelity and breast cancer in mammary luminal epithelial cells. bioRxiv 2022. [Google Scholar] [CrossRef]
- Stevanovic, M.; Lazic, A.; Schwirtlich, M.; Stanisavljevic Ninkovic, D. The Role of SOX Transcription Factors in Ageing and Age-Related Diseases. Int. J. Mol. Sci. 2023, 24, 851. [Google Scholar] [CrossRef] [PubMed]
- Carrasco-Garcia, E.; Moreno-Cugnon, L.; Garcia, I.; Borras, C.; Revuelta, M.; Izeta, A.; Lopez-Lluch, G.; de Pancorbo, M.M.; Vergara, I.; Vina, J.; et al. SOX2 expression diminishes with ageing in several tissues in mice and humans. Mech. Ageing Dev. 2019, 177, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Larson, B.L.; Ylostalo, J.; Lee, R.H.; Gregory, C.; Prockop, D.J. Sox11 is expressed in early progenitor human multipotent stromal cells and decreases with extensive expansion of the cells. Tissue Eng. Part. A 2010, 16, 3385–3394. [Google Scholar] [CrossRef]
- Dy, P.; Smits, P.; Silvester, A.; Penzo-Méndez, A.; Dumitriu, B.; Han, Y.; de la Motte, C.A.; Kingsley, D.M.; Lefebvre, V. Synovial joint morphogenesis requires the chondrogenic action of Sox5 and Sox6 in growth plate and articular cartilage. Dev. Biol. 2010, 341, 346–359. [Google Scholar] [CrossRef]
- Zhang, X.; Wu, S.; Zhu, Y.; Chu, C.-Q. Exploiting Joint-Resident Stem Cells by Exogenous SOX9 for Cartilage Regeneration for Therapy of Osteoarthritis. Front. Med. 2021, 8, 622609. [Google Scholar] [CrossRef] [PubMed]
- Park, J.S.; Yang, H.N.; Woo, D.G.; Jeon, S.Y.; Do, H.-J.; Lim, H.-Y.; Kim, J.-H.; Park, K.-H. Chondrogenesis of human mesenchymal stem cells mediated by the combination of SOX trio SOX5, 6, and 9 genes complexed with PEI-modified PLGA nanoparticles. Biomaterials 2011, 32, 3679–3688. [Google Scholar] [CrossRef]
- Gadi, J.; Jung, S.-H.; Lee, M.-J.; Jami, A.; Ruthala, K.; Kim, K.-M.; Cho, N.-H.; Jung, H.-S.; Kim, C.-H.; Lim, S.-K. The Transcription Factor Protein Sox11 Enhances Early Osteoblast Differentiation by Facilitating Proliferation and the Survival of Mesenchymal and Osteoblast Progenitors. J. Biol. Chem. 2013, 288, 25400. [Google Scholar] [CrossRef]
- Yang, Y.-H.K. Aging of mesenchymal stem cells: Implication in regenerative medicine. Regen. Ther. 2018, 9, 120–122. [Google Scholar] [CrossRef]
- Stolzing, A.; Jones, E.; McGonagle, D.; Scutt, A. Age-related changes in human bone marrow-derived mesenchymal stem cells: Consequences for cell therapies. Mech. Ageing Dev. 2008, 129, 163–173. [Google Scholar] [CrossRef]
- Choudhery, M.S.; Badowski, M.; Muise, A.; Pierce, J.; Harris, D.T. Donor age negatively impacts adipose tissue-derived mesenchymal stem cell expansion and differentiation. J. Transl. Med. 2014, 12, 8. [Google Scholar] [CrossRef] [PubMed]
- Bhattaram, P.; Muschler, G.; Wixler, V.; Lefebvre, V. Inflammatory Cytokines Stabilize SOXC Transcription Factors to Mediate the Transformation of Fibroblast-Like Synoviocytes in Arthritic Disease. Arthritis Rheumatol. 2018, 70, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Feng, X.; Shi, Y.; Xu, L.; Peng, Q.; Wang, F.; Wang, X.; Sun, W.; Lu, Y.; Tsao, B.P.; Zhang, M.; et al. Modulation of IL-6 induced RANKL expression in arthritic synovium by a transcription factor SOX5. Sci. Rep. 2016, 6, 32001. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y.; Wu, Q.; Xuan, W.; Feng, X.; Wang, F.; Tsao, B.P.; Zhang, M.; Tan, W. Transcription Factor SOX5 Promotes the Migration and Invasion of Fibroblast-Like Synoviocytes in Part by Regulating MMP-9 Expression in Collagen-Induced Arthritis. Front. Immunol. 2018, 9, 749. [Google Scholar] [CrossRef] [PubMed]
- Wijesinghe, S.N.; Badoume, A.; Nanus, D.E.; Sharma-Oates, A.; Farah, H.; Certo, M.; Alnajjar, F.; Davis, E.T.; Mauro, C.; Lindsay, M.A.; et al. Obesity defined molecular endotypes in the synovium of patients with osteoarthritis provides a rationale for therapeutic targeting of fibroblast subsets. Clin. Transl. Med. 2023, 13, e1232. [Google Scholar] [CrossRef] [PubMed]
- Nanus, D.E.; Wijesinghe, S.N.; Pearson, M.J.; Hadjicharalambous, M.R.; Rosser, A.; Davis, E.T.; Lindsay, M.A.; Jones, S.W. Regulation of the Inflammatory Synovial Fibroblast Phenotype by Metastasis-Associated Lung Adenocarcinoma Transcript 1 Long Noncoding RNA in Obese Patients with Osteoarthritis. Arthritis Rheumatol. 2020, 72, 609–619. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inclusion Criteria |
|---|
| Diagnosis of primary knee OA with predominantly medial involvement |
| Advanced degenerative changes found in the radiological examination The third or fourth degree of disease, according to Kellgren–Lawrence classification |
| Giving voluntary, informed, written consent to participate in the study |
| Exclusion criteria |
| Rheumatoid arthritis |
| Hemophilia |
| Psoriatic arthritis |
| Neurological disorders |
| Patient Characteristics | Number of Patients (n) | Total Percentage of Patients |
|---|---|---|
| Sex | ||
| Female | 21 | 68% |
| Male | 10 | 32% |
| Age | ||
| ≤65 years | 14 | 45% |
| >65 years | 17 | 55% |
| Weight | ||
| ≤88 kg | 16 | 52% |
| >88 kg | 15 | 48% |
| BMI a | ||
| 25.0–29.9 (overweight) | 4 | 13% |
| 30.0–34.9 (class I obesity) | 20 | 65% |
| 35.0–39.0 (class II obesity) | 6 | 19% |
| ≥40 (class III obesity) | 1 | 3% |
| Stage of knee OA according to Kellgren–Lawrence classification | ||
| Stage 3 | 14 | 45% |
| Stage 4 | 17 | 55% |
| SOX5 | SOX9 | SOX11 | ||||
|---|---|---|---|---|---|---|
| (Median RQ) | (Median RQ) | (Median RQ) | ||||
| OA-affected articular cartilage with subchondral bone control tissue | p = 0.00015 | p = 0.0024 | p > 0.05 | |||
| 237.935 | 522.929 | 354.66 | ||||
| 69.395 | 178.271 | 317.992 | ||||
| female | 220.128 | p > 0.05 | 556.385 | p > 0.05 | 354.66 | p > 0.05 |
| male | 339.09 | 400.887 | 332.913 | |||
| ≤65 years | 339.09 | p = 0.04 | 606.861 | p = 0.017 | 372.414 | p > 0.05 |
| >65 years | 189.304 | 272.911 | 285.509 | |||
| Overweight | 210.926 | p > 0.05 | 478.408 | p > 0.05 | 556.954 | p > 0.05 |
| Class I obesity | 285.76 | 459.286 | 280.584 | |||
| Class II + III obesity | 310.183 | 800.773 | 390.167 | |||
| Stage 3 | 240.662 | p > 0.05 | 348.422 | p > 0.05 | 280.584 | p > 0.05 |
| Stage 4 | 287.935 | 585.338 | 390.167 | |||
| SOX5 | SOX9 | SOX11 | ||||
|---|---|---|---|---|---|---|
| (Median RQ) | (Median RQ) | (Median RQ) | ||||
| OA-affected synovium | 10.193 | p = 0.0003 | 19.722 | p > 0.05 | 12.436 | p = 0.0007 |
| Control tissue | 23.297 | 22.874 | 44.471 | |||
| female | 11.056 | p > 0.05 | 24.511 | p > 0.05 | 10.717 | p > 0.05 |
| male | 7.354 | 14.373 | 15.003 | |||
| ≤65 years | 12.583 | p > 0.05 | 20.699 | p > 0.05 | 15.003 | p > 0.05 |
| >65 years | 8.271 | 19.699 | 5.8 | |||
| Overweight | 4.265 | p > 0.05 | 14.333 | p > 0.05 | 6.308 | p > 0.05 |
| Class I obesity | 9.281 | 21.511 | 12.875 | |||
| Class II + III obesity | 14.658 | 21.849 | 6.95 | |||
| Stage 3 | 5.884 | p > 0.05 | 20.786 | p > 0.05 | 11.577 | p > 0.05 |
| Stage 4 | 11.674 | 19.549 | 12.746 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baran, K.; Brzeziańska-Lasota, E.; Kryczka, J.; Boncela, J.; Czechowska, A.; Kopacz, K.; Padula, G.; Nowak, K.; Domżalski, M. The Expression Level of SOX Family Transcription Factors’ mRNA as a Diagnostic Marker for Osteoarthritis. J. Clin. Med. 2025, 14, 1176. https://doi.org/10.3390/jcm14041176
Baran K, Brzeziańska-Lasota E, Kryczka J, Boncela J, Czechowska A, Kopacz K, Padula G, Nowak K, Domżalski M. The Expression Level of SOX Family Transcription Factors’ mRNA as a Diagnostic Marker for Osteoarthritis. Journal of Clinical Medicine. 2025; 14(4):1176. https://doi.org/10.3390/jcm14041176
Chicago/Turabian StyleBaran, Kamila, Ewa Brzeziańska-Lasota, Jakub Kryczka, Joanna Boncela, Aleksandra Czechowska, Karolina Kopacz, Gianluca Padula, Krzysztof Nowak, and Marcin Domżalski. 2025. "The Expression Level of SOX Family Transcription Factors’ mRNA as a Diagnostic Marker for Osteoarthritis" Journal of Clinical Medicine 14, no. 4: 1176. https://doi.org/10.3390/jcm14041176
APA StyleBaran, K., Brzeziańska-Lasota, E., Kryczka, J., Boncela, J., Czechowska, A., Kopacz, K., Padula, G., Nowak, K., & Domżalski, M. (2025). The Expression Level of SOX Family Transcription Factors’ mRNA as a Diagnostic Marker for Osteoarthritis. Journal of Clinical Medicine, 14(4), 1176. https://doi.org/10.3390/jcm14041176