
The Role of the Microbiota Gut–Liver Axis during HCV Chronic Infection: A Schematic Overview

 , , , , and
, , , , and 
Abstract
1. Introduction
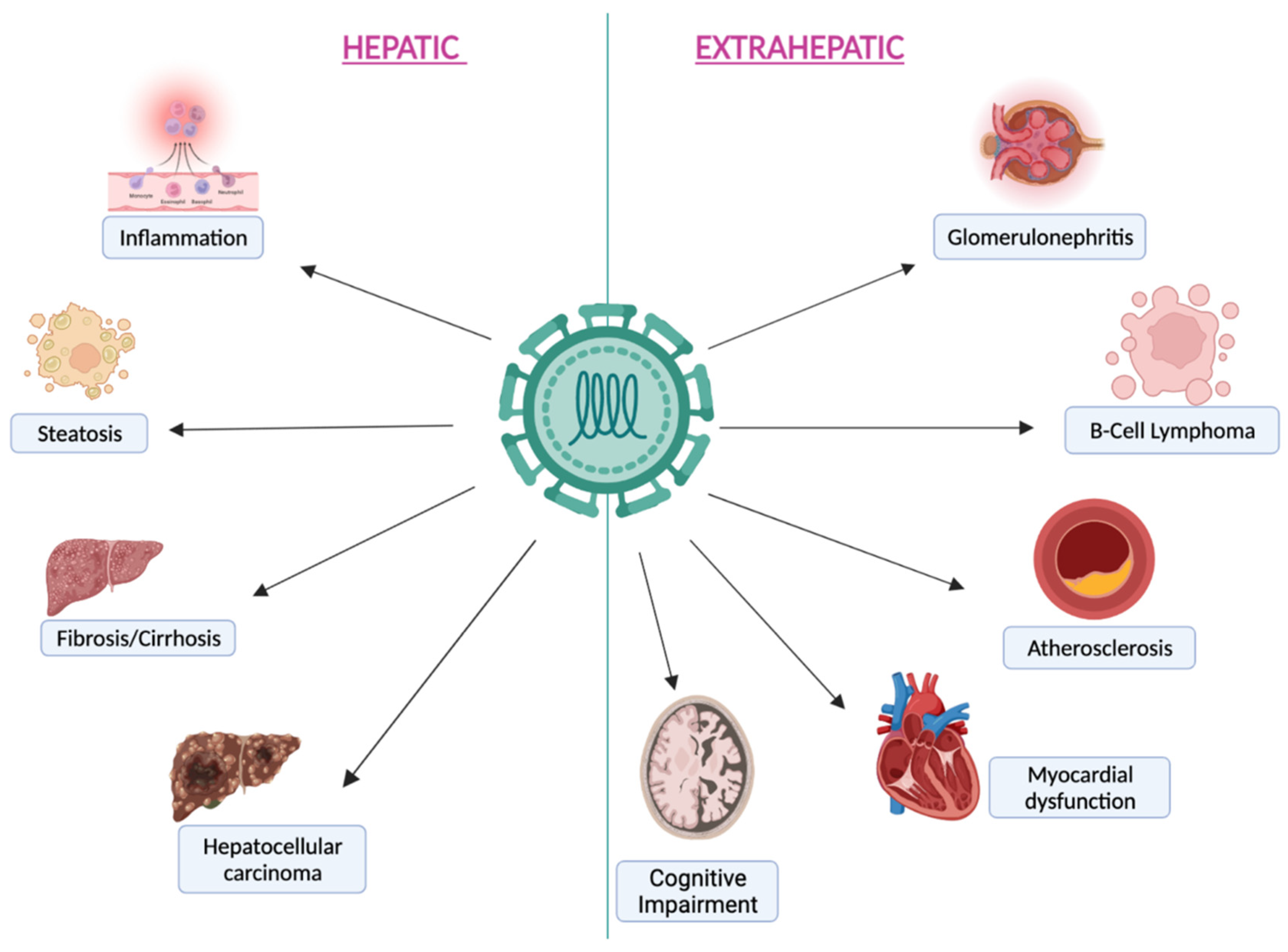
2. HCV Infection: Epidemiology and Pathogenesis
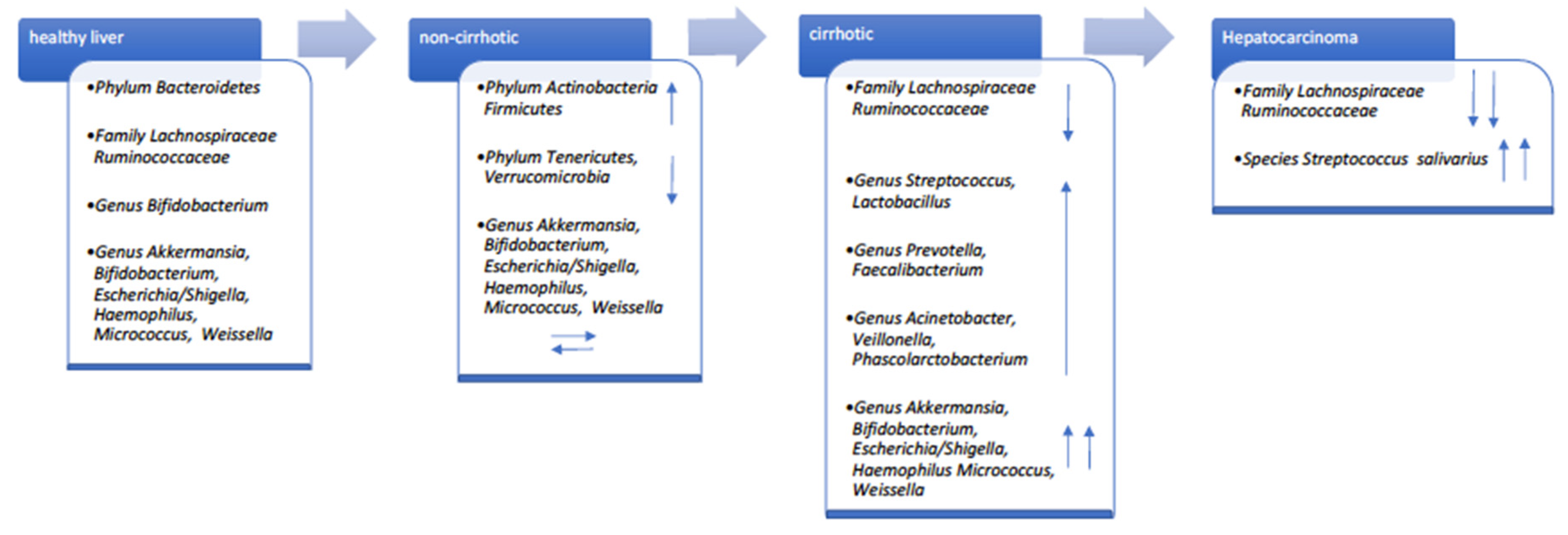
3. HCV Infection Effect on the Gut–Liver Axis
4. Gut Microbiota and HCV Therapy
5. Gut Microbiota and Therapeutic Manipulation
6. Conclusions
- Study designs should be performed taking into account dietary features and geographic location of positive patients.
- The control group of non-infected subjects should be matched for age, gender, and diet.
- Experimental studies should be designed according to HCV genotype/subtype and liver disease status of positive patients.
- Pre-clinical and clinical studies, including a large cohort of patients, are required to better understand the link between the gut and liver during chronic infection.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- World Health Organization (WHO). Available online: https://www.who.int (accessed on 5 August 2022).
- European Association for the Study of the Liver. EASL recommendations on treatment of hepatitis C: Final update of the series. J. Hepatol. 2020, 73, 1170–1218. [Google Scholar] [CrossRef]
- Blach, S.; Kondili, L.A.; Aghemo, A.; Cai, Z.; Dugan, E.; Estes, C.; Gamkrelidze, I.; Ma, S.; Pawlotsky, J.M.; Razavi-Shearer, D.; et al. Impact of COVID-19 on global HCV elimination efforts. J. Hepatol. 2021, 74, 31–36. [Google Scholar] [CrossRef]
- Van Dijk, M.; Brakenhoff, S.M.; Isfordink, C.J.; Cheng, W.H.; Blokzijl, H.; Boland, G.; Dofferhoff, A.S.M.; van Hoek, B.; van Nieuwkoop, C.; Sonneveld, M.J.; et al. The Netherlands Is on Track to Meet the World Health Organization Hepatitis C Elimination Targets by 2030. J. Clin. Med. 2021, 10, 4562. [Google Scholar] [CrossRef] [PubMed]
- Marascio, N.; Rotundo, S.; Quirino, A.; Matera, G.; Liberto, M.C.; Costa, C.; Russo, A.; Trecarichi, E.M.; Torti, C. Similarities, differences, and possible interactions between hepatitis E and hepatitis C viruses: Relevance for research and clinical practice. World J. Gastroenterol. 2022, 28, 1226–1238. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Marcell, L.; Kottilil, S. Systemic manifestations of hepatitis C Infection. Infect Agent Cancer. 2016, 11, 29. [Google Scholar] [CrossRef] [PubMed]
- Marascio, N.; Quirino, A.; Barreca, G.S.; Galati, L.; Costa, C.; Pisani, V.; Mazzitelli, M.; Matera, G.; Liberto, M.C.; Focà, A.; et al. Discussion on critical points for a tailored therapy to cure hepatitis C virus infection. Clin. Mol. Hepatol. 2019, 25, 30–36. [Google Scholar] [CrossRef]
- Wellhöner, F.; Döscher, N.; Woelfl, F.; Vital, M.; Plumeier, I.; Kahl, S.; Potthoff, A.; Manns, M.P.; Pieper, D.H.; Cornberg, M.; et al. Eradication of Chronic HCV Infection: Improvement of Dysbiosis Only in Patients without Liver Cirrhosis. Hepatology 2021, 74, 72–82. [Google Scholar] [CrossRef]
- Chen, W.; Wei, Y.; Xiong, A.; Li, Y.; Guan, H.; Wang, Q.; Miao, Q.; Bian, Z.; Xiao, X.; Lian, M.; et al. Comprehensive Analysis of Serum and Fecal Bile Acid Profiles and Interaction with Gut Microbiota in Primary Biliary Cholangitis. Clin. Rev. Allergy Immunol. 2020, 58, 25–38. [Google Scholar] [CrossRef]
- Granito, A.; Muratori, P.; Muratori, L. Editorial: Gut microbiota profile in patients with autoimmune hepatitis-aclue for adjunctive probiotic therapy? Aliment. Pharmacol. Ther. 2020, 52, 392–394. [Google Scholar] [CrossRef]
- Petersen, C.; Round, J.L. Defining dysbiosis and its influence on host immunity and disease. Cell Microbiol. 2014, 16, 1024–1033. [Google Scholar] [CrossRef]
- Sultan, S.; El-Mowafy, M.; Elgaml, A.; El-Mesery, M.; El Shabrawi, A.; Elegezy, M.; Hammami, R.; Mottawea, W. Alterations of the Treatment-Naive Gut Microbiome in Newly Diagnosed Hepatitis C Virus Infection. ACS Infect. Dis. 2021, 7, 1059–1068. [Google Scholar] [CrossRef] [PubMed]
- Ponziani, F.R.; Putignani, L.; Paroni Sterbini, F.; Petito, V.; Picca, A.; Del Chierico, F.; Reddel, S.; Calvani, R.; Marzetti, E.; Sanguinetti, M.; et al. Influence of hepatitis C virus eradication with direct-acting antivirals on the gut microbiota in patients with cirrhosis. Aliment Pharmacol. Ther. 2018, 48, 1301–1311. [Google Scholar] [CrossRef]
- Zheng, R.; Wang, G.; Pang, Z.; Ran, N.; Gu, Y.; Guan, X.; Yuan, Y.; Zuo, X.; Pan, H.; Zheng, J.; et al. Liver cirrhosis contributes to the disorder of gut microbiota in patients with hepatocellular carcinoma. Cancer Med. 2020, 9, 4232–4250. [Google Scholar] [CrossRef]
- El-Mowafy, M.; Elgaml, A.; El-Mesery, M.; Sultan, S.; Ahmed, T.A.E.; Gomaa, A.I.; Aly, M.; Mottawea, W. Changes of Gut-Microbiota-Liver Axis in Hepatitis C Virus Infection. Biology 2021, 10, 55. [Google Scholar] [CrossRef] [PubMed]
- Yamane, D.; McGivern, D.R.; Masaki, T.; Lemon, S.M. Liver injury and disease pathogenesis in chronic hepatitis C. Curr. Top. Microbiol. Immunol. 2013, 369, 263–288. [Google Scholar] [CrossRef] [PubMed]
- Gower, E.; Estes, C.; Blach, S.; Razavi-Shearer, K.; Razavi, H. Global epidemiology and genotype distribution of the hepatitis C virus infection. J. Hepatol. 2014, 61 (Suppl. 1), S45–S57. [Google Scholar] [CrossRef]
- Soza, A.; Riquelme, A.; Arrese, M. Routes of transmission of hepatitis C virus. Ann Hepatol. 2010, 9 (Suppl. 1), 33. [Google Scholar] [CrossRef]
- Nijmeijer, B.M.; Koopsen, J.; Schinkel, J.; Prins, M.; Geijtenbeek, T.B. Sexually transmitted hepatitis C virus infections: Current trends, and recent advances in understanding the spread in men who have sex with men. J. Int. AIDS Soc. 2019, 22 (Suppl. 6), e25348. [Google Scholar] [CrossRef]
- Negro, F. Natural History of Hepatic and Extrahepatic Hepatitis C Virus Diseases and Impact of Interferon-Free HCV Therapy. Cold Spring Harb. Perspect. Med. 2020, 10, a036921. [Google Scholar] [CrossRef]
- Jackowiak, P.; Figlerowicz, M.; Kurzyńska-Kokorniak, A.; Figlerowicz, M. Mechanisms involved in the development of chronic hepatitis C as potential targets of antiviral therapy. Curr Pharm. Biotechnol. 2011, 12, 1774–1780. [Google Scholar] [CrossRef]
- Schoggins, J.W.; Rice, C.M. Innate immune responses to hepatitis C virus. Curr. Top. Microbiol. Immunol. 2013, 369, 219–242. [Google Scholar] [CrossRef] [PubMed]
- Irshad, M.; Mankotia, D.S.; Irshad, K. An insight into the diagnosis and pathogenesis of hepatitis C virus infection. World J. Gastroenterol. 2013, 19, 7896–7909. [Google Scholar] [CrossRef] [PubMed]
- Vento, S.; Nobili, V.; Cainelli, F. Clinical course of infection with hepatitis C is still poorly understood. BMJ 2006, 332, 374–375. [Google Scholar] [CrossRef] [PubMed]
- Janiak, M.; Perlejewski, K.; Grabarczyk, P.; Kubicka-Russel, D.; Zagordi, O.; Berak, H.; Osuch, S.; Pawełczyk, A.; Bukowska-Ośko, I.; Płoski, R.; et al. Hepatitis C virus (HCV) genotype 1b displays higher genetic variability of hypervariable region 1 (HVR1) than genotype 3. Sci. Rep. 2019, 9, 12846. [Google Scholar] [CrossRef] [PubMed]
- Goel, A.; Bhadauria, D.S.; Aggarwal, R. Hepatitis C virus infection and chronic renal disease: A review. Indian J. Gastroenterol. 2018, 37, 492–503. [Google Scholar] [CrossRef] [PubMed]
- Khatun, M.; Ray, R.B. Mechanisms underlying hepatitis C virus-associated hepatic fibrosis. Cells 2019, 8, 1249. [Google Scholar] [CrossRef] [PubMed]
- Ioannou, G.N.; Beste, L.A.; Green, P.K.; Singal, A.G.; Tapper, E.B.; Waljee, A.K.; Sterling, R.K.; Feld, J.J.; Kaplan, D.E.; Taddei, T.H.; et al. Increased Risk for Hepatocellular Carcinoma Persists Up to 10 Years After HCV Eradication in Patients With Baseline Cirrhosis or High FIB-4 Scores. Gastroenterology 2019, 157, 1264–1278.e4. [Google Scholar] [CrossRef] [PubMed]
- Strazzulla, A.; Iemmolo, R.M.R.; Carbone, E.; Postorino, M.C.; Mazzitelli, M.; De Santis, M.; Di Benedetto, F.; Cristiani, C.M.; Costa, C.; Pisani, V.; et al. The Risk of Hepatocellular Carcinoma After Directly Acting Antivirals for Hepatitis C Virus Treatment in Liver Transplanted Patients: Is It Real? Hepat. Mon. 2016, 16, e41933. [Google Scholar] [CrossRef] [PubMed]
- Tremaroli, V.; Bäckhed, F. Functional interactions between the gut microbiota and host metabolism. Nature 2012, 489, 242–249. [Google Scholar] [CrossRef] [PubMed]
- Cremer, J.; Arnoldini, M.; Hwa, T. Effect of water flow and chemical environment on microbiota growth and composition in the human colon. Proc. Natl. Acad. Sci. USA 2017, 114, 6438–6443. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Gao, Z.; Zhang, J.; Ye, X.; Xu, A.; Ye, J.; Jia, W. Sodium butyrate stimulates expression of fibroblast growth factor 21 in liver by inhibition of histone deacetylase 3. Diabetes 2012, 61, 797–806. [Google Scholar] [CrossRef] [PubMed]
- Brandl, K.; Kumar, V.; Eckmann, L. Gut-liver axis at the frontier of host-microbial interactions. Am. J. Physiol. Gastrointest. Liver Physiol. 2017, 312, G413–G419. [Google Scholar] [CrossRef] [PubMed]
- Chiang, J.Y.L.; Ferrell, J.M. Bile Acid Metabolism in Liver Pathobiology. Gene Expr. 2018, 18, 71–87. [Google Scholar] [CrossRef] [PubMed]
- Wahlström, A.; Kovatcheva-Datchary, P.; Ståhlman, M.; Bäckhed, F.; Marschall, H.-U. Crosstalk between Bile Acids and Gut Microbiota and Its Impact on Farnesoid X Receptor Signalling. Dig. Dis. 2017, 35, 246–250. [Google Scholar] [CrossRef]
- Ridlon, J.M.; Kang, D.-J.; Hylemon, P.B. Bile salt biotransformations by human intestinal bacteria. J. Lipid Res. 2006, 47, 241–259. [Google Scholar] [CrossRef]
- Swann, J.R.; Want, E.J.; Geier, F.M.; Spagou, K.; Wilson, I.D.; Sidaway, J.E.; Nicholson, J.K.; Holmes, E. Systemic gut microbial modulation of bile acid metabolism in host tissue compartments. Proc. Natl. Acad. Sci. USA 2011, 108 (Suppl. 1), 4523–4530. [Google Scholar] [CrossRef]
- Kakiyama, G.; Pandak, W.M.; Gillevet, P.M.; Hylemon, P.B.; Heuman, D.M.; Daita, K.; Takei, H.; Muto, A.; Nittono, H.; Ridlon, J.M.; et al. Modulation of the fecal bile acid profile by gut microbiota in cirrhosis. J. Hepatol. 2013, 58, 949–955. [Google Scholar] [CrossRef]
- Suk, K.T.; Kim, D.J. Gut microbiota: Novel therapeutic target for nonalcoholic fatty liver disease. Expert Rev. Gastroenterol. Hepatol. 2019, 13, 193–204. [Google Scholar] [CrossRef]
- Duboc, H.; Rajca, S.; Rainteau, D.; Benarous, D.; Maubert, M.A.; Quervain, E.; Thomas, G.; Barbu, V.; Humbert, L.; Despras, G.; et al. Connecting dysbiosis, bile-acid dysmetabolism and gut inflammation in inflammatory bowel diseases. Gut 2013, 62, 531–539. [Google Scholar] [CrossRef]
- Paul, B.; Lewinska, M.; Andersen, J.B. Lipid alterations in chronic liver disease and liver cancer. JHEP Rep. 2022, 4, 100479. [Google Scholar] [CrossRef]
- Inoue, T.; Nakayama, J.; Moriya, K.; Kawaratani, H.; Momoda, R.; Ito, K.; Iio, E.; Nojiri, S.; Fujiwara, K.; Yonedaet, M.; et al. Gut Dysbiosis associated with hepatitis C virus infection. Clin. Infect. Dis. 2018, 67, 869–877. [Google Scholar] [CrossRef] [PubMed]
- Preveden, T.; Scarpellini, E.; Milic, N.; Luzza, F.; Abenavoli, L. Gut microbiota changes and chronic hepatitis C virus infection. Expert Rev. Gastroenterol. Hepatol. 2017, 11, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Gupta, H.; Youn, G.S.; Shin, M.J.; Suk, K.T. Role of Gut Microbiota in Hepatocarcinogenesis. Microorganisms 2019, 7, 121. [Google Scholar] [CrossRef] [PubMed]
- Aly, A.M.; Adel, A.; El-Gendy, A.O.; Essam, T.M.; Aziz, R.K. Gut microbiome alterations in patients with stage 4 hepatitis C. Gut Pathog. 2016, 8, 42. [Google Scholar] [CrossRef]
- Pérez-Matute, P.; Íñiguez, M.; Villanueva-Millán, M.J.; Recio-Fernández, E.; Vázquez, A.M.; Sánchez, S.C.; Morano, L.E.; Oteo, J.A. Short-term effects of direct-acting antiviral agents on inflammation and gut microbiota in hepatitis C-infected patients. Eur. J. Intern. Med. 2019, 67, 47–58. [Google Scholar] [CrossRef]
- Heidrich, B.; Vital, M.; Plumeier, I.; Döscher, N.; Kahl, S.; Kirschner, J.; Ziegert, S.; Solbach, P.; Lenzen, H.; Potthoff, A.; et al. Intestinal microbiota in patients with chronic hepatitis C with and without cirrhosis compared with healthy controls. Liver Int. 2018, 38, 50–58. [Google Scholar] [CrossRef]
- Cosseau, C.; Devine, D.A.; Dullaghan, E.; Gardy, J.L.; Chikatamarla, A.; Gellatly, S.; Yu, L.L.; Pistolic, J.; Falsafi, R.; Tagg, J.; et al. The commensal Streptococcus salivarius K12 downregulates the innate immune responses of human epithelial cells and promotes host-microbe homeostasis. Infect. Immun. 2008, 76, 4163–4175. [Google Scholar] [CrossRef]
- Manns, M.P.; McHutchison, J.G.; Gordon, S.C.; Rustgi, V.K.; Shiffman, M.; Reindollar, R.; Goodman, Z.D.; Koury, K.; Ling, M.; Albrecht, J.K. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: A randomised trial. Lancet 2001, 358, 958–965. [Google Scholar] [CrossRef]
- European Association for the Study of the Liver. EASL recommendations on treatment of hepatitis C 2016. J. Hepatol. 2017, 66, 153–194. [Google Scholar] [CrossRef]
- Bartenschlager, R.; Penin, F.; Lohmann, V.; André, P. Assembly of infectious hepatitis C virus particles. Trends Microbiol. 2011, 19, 95–103. [Google Scholar] [CrossRef]
- Pawlotsky, J.M. Hepatitis C virus resistance to direct-acting antiviral drugs in interferon-free regimens. Gastroenterology 2016, 151, 70–86. [Google Scholar] [CrossRef]
- Molino, S.; Martin, M.T. Hepatitis C virus resistance testing in genotype 1, the changing role in clinical utility. Ann Pharmacother. 2017, 51, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Bajaj, J.S.; Sterling, R.K.; Betrapally, N.S.; Nixon, D.E.; Fuchs, M.; Daita, K.; Heuman, D.M.; Sikaroodi, M.; Hylemon, P.B.; White, M.B.; et al. HCV eradication does not impact gut dysbiosis or systemic inflammation in cirrhotic patients. Aliment. Pharmacol Ther. 2016, 44, 638–643. [Google Scholar] [CrossRef]
- Begley, M.; Gahan, C.G.; Hill, C. The interaction between bacteria and bile. FEMS Microbiol. Rev. 2005, 29, 625–651. [Google Scholar] [CrossRef] [PubMed]
- D’Ambrosio, R.; Aghemo, A.; Rumi, M.G.; Ronchi, G.; Donato, M.F.; Paradis, V.; Colombo, M.; Bedossa, P. A morphometric and immune histochemical study to assess the benefit of a sustained virological response in hepatitis C virus patients with cirrhosis. Hepatology 2012, 56, 532–543. [Google Scholar] [CrossRef]
- Yilmaz, B.; Ruckstuhl, L.; Müllhaupt, B.; Magenta, L.; Kuster, M.H.; Clerc, O.; Torgler, R.; Semmo, N. Pilot Sub-Study of the Effect of Hepatitis C Cure by Glecaprevir/Pibrentasvir on the Gut Microbiome of Patients with Chronic Hepatitis C Genotypes 1 to 6 in the Mythen Study. Pharmaceuticals 2021, 14, 931. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A.; et al. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef]
- Wang, L.; Wan, Y.Y. The role of gut microbiota in liver disease development and treatment. Liver Res. 2019, 3, 3–18. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Kassam, Z.; Fagan, A.; Gavis, E.A.; Liu, E.; Cox, I.J.; Kheradman, R.; Heuman, D.; Wang, J.; Gurry, T.; et al. Fecal microbiota transplant from a rational stool donor improves hepatic encephalopathy: A randomized clinical trial. Hepatology 2017, 66, 1727–1738. [Google Scholar] [CrossRef]
- Bajaj, J.S.; Heuman, D.M.; Sanyal, A.J.; Hylemon, P.B.; Sterling, R.K.; Stravitz, R.T.; Fuchs, M.; Ridlon, J.M.; Daita, K.; Monteith, P.; et al. Modulation of the metabiome by rifaximin in patients with cirrhosis and minimal hepatic encephalopathy. PLoS ONE 2013, 8, e60042. [Google Scholar] [CrossRef]
- Stern, J.; Miller, G.; Li, X.; Saxena, D. Virome and bacteriome: Two sides of the same coin. Curr. Opin. Virol. 2019, 37, 37–43. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Antiviral Therapy | Liver Status | Gut Composition and Time of Evaluation |
|---|---|---|
| SOC | Cirrhosis | Improvement of dysbiosis 15 months later SVR [54] |
| DAA | Cirrhosis | Improvement of dysbiosis 12 months later SVR [13] |
| DAA | Cirrhosis | No significant bacterial diversity between baseline and 24/48 weeks later SVR [8] |
| DAA | No cirrhosis | Improvement of dysbiosis 24/48 weeks later SVR [8] |
| DAA | Cirrhosis/No cirrhosis | No significant bacterial diversity between baseline and 12 weeks later SVR [57] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Marascio, N.; De Caro, C.; Quirino, A.; Mazzitelli, M.; Russo, E.; Torti, C.; Matera, G. The Role of the Microbiota Gut–Liver Axis during HCV Chronic Infection: A Schematic Overview. J. Clin. Med. 2022, 11, 5936. https://doi.org/10.3390/jcm11195936
Marascio N, De Caro C, Quirino A, Mazzitelli M, Russo E, Torti C, Matera G. The Role of the Microbiota Gut–Liver Axis during HCV Chronic Infection: A Schematic Overview. Journal of Clinical Medicine. 2022; 11(19):5936. https://doi.org/10.3390/jcm11195936
Chicago/Turabian StyleMarascio, Nadia, Carmen De Caro, Angela Quirino, Maria Mazzitelli, Emilio Russo, Carlo Torti, and Giovanni Matera. 2022. "The Role of the Microbiota Gut–Liver Axis during HCV Chronic Infection: A Schematic Overview" Journal of Clinical Medicine 11, no. 19: 5936. https://doi.org/10.3390/jcm11195936
APA StyleMarascio, N., De Caro, C., Quirino, A., Mazzitelli, M., Russo, E., Torti, C., & Matera, G. (2022). The Role of the Microbiota Gut–Liver Axis during HCV Chronic Infection: A Schematic Overview. Journal of Clinical Medicine, 11(19), 5936. https://doi.org/10.3390/jcm11195936