Lipid Rafts in Signalling, Diseases, and Infections: What Can Be Learned from Fluorescence Techniques?


Abstract
1. Introduction
1.1. The Lipid Raft Hypothesis
1.2. Lipid Raft Functions
1.3. Lipid Rafts and Diseases
2. Methods of Studying Lipid Organization
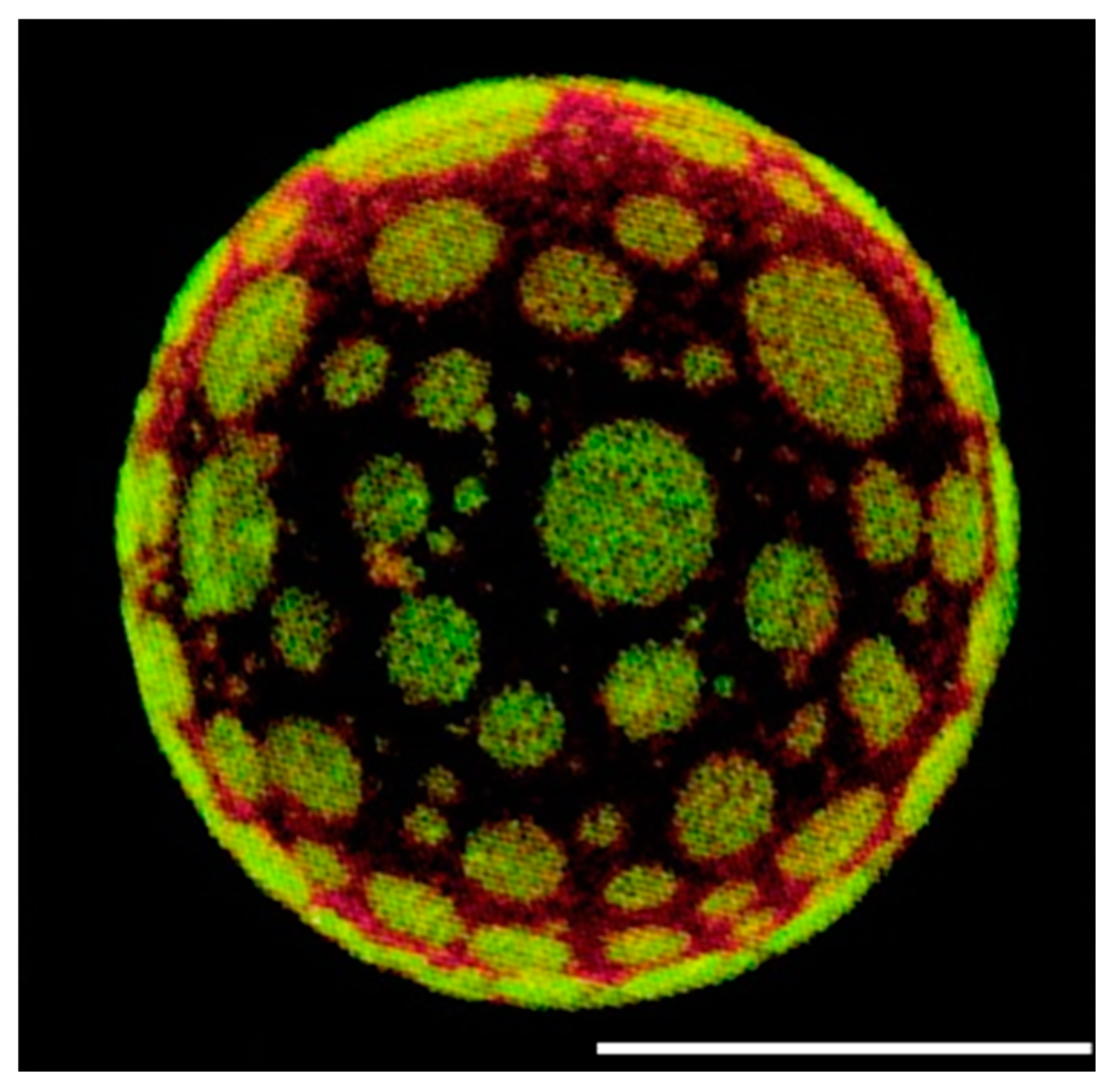
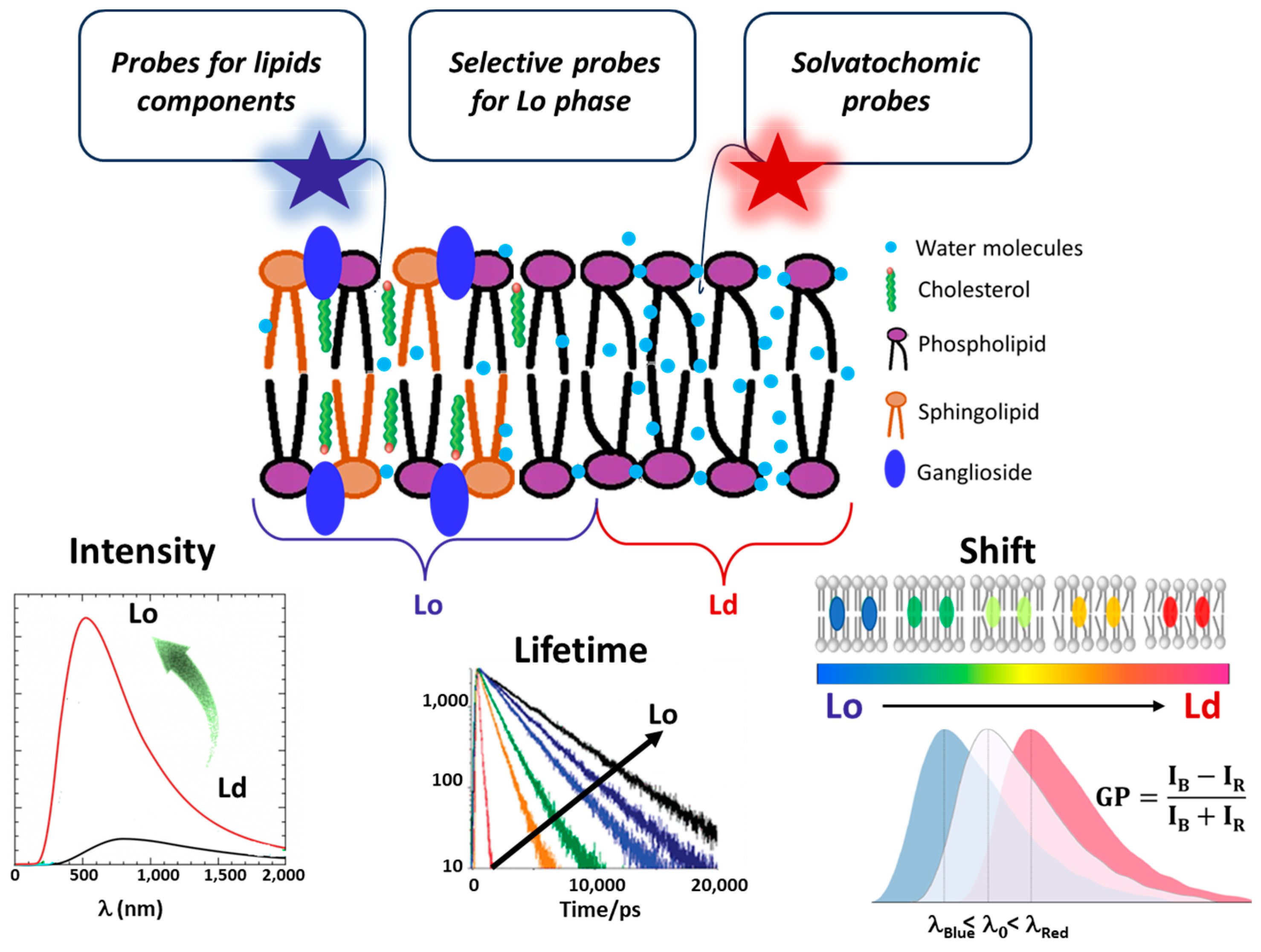
3. Fluorescent Probes for Studying Lipid Rafts
3.1. Probes Selective for Lipid Components
3.2. Probes Selective for the Lo Membrane Phase
3.3. Environment-Sensitive Dyes
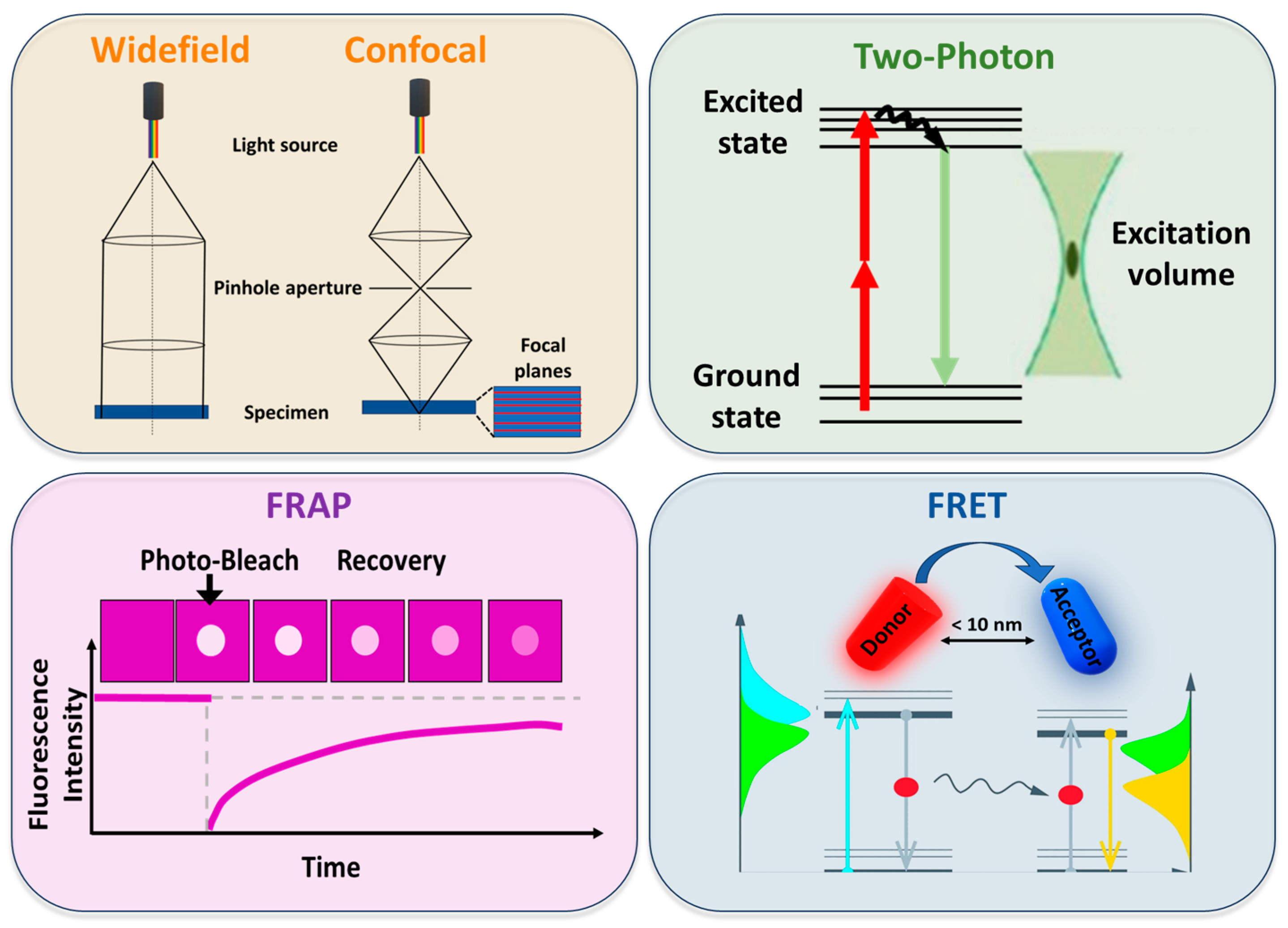
4. Fluorescence-Based Techniques for Studying Lipid Rafts
5. Advanced Fluorescence Imaging Methods
5.1. Nanoscale Imaging of Lipid Rafts


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| STED | SIM | SMLM | |
|---|---|---|---|
| Lateral resolution | ≈20–30 nm | ≈60–100 nm | ≈20–30 nm |
| Microscope system | Scanning | Widefield | Widefield |
| Acquisition time | Seconds–minutes | Seconds–minutes | Minutes–hours |
| Dyes | Suited for emission stimulation | Conventional | Photoswitchable–Photoactivable |
| Post-processing | no | yes | yes |
| Limitations | High energy lasers | High computational analysis | Uniform labelling density |
| Refs | [149,166] | [153] | [158] |
5.2. Probing Dynamics of Lipid Rafts
6. Lipid Rafts: Recent Insights into Signalling, Diseases, and Infections Using Fluorescence Techniques
6.1. Involvement of Lipid Rafts in Diseases
6.1.1. Neuronal Diseases
6.1.2. Viruses and Bacteria
6.2. Lipid Rafts as Signalling Hubs
6.3. Targeting Lipid Rafts Using FRET-Based Biosensors
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jacobson, K.; Mouritsen, O.G.; Anderson, R.G.W. Lipid rafts: At a crossroad between cell biology and physics. Nat. Cell Biol. 2007, 9, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Jacobson, K.; Sheets, E.D.; Simson, R. Revisiting the Fluid Mosaic Model of Membranes. Science 1995, 268, 1441–1442. [Google Scholar] [CrossRef] [PubMed]
- Parton, R.G.; Simons, K. Digging into Caveolae. Science 1995, 269, 1398–1399. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ikonen, E. Functional rafts in cell membranes. Nature 1997, 387, 569–572. [Google Scholar] [CrossRef] [PubMed]
- Levental, I.; Levental, K.R.; Heberle, F.A. Lipid Rafts: Controversies Resolved, Mysteries Remain. Trends Cell Biol. 2020, 30, 341–353. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; London, E. Structure and Origin of Ordered Lipid Domains in Biological Membranes. J. Membr. Biol. 1998, 164, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.M.; Gaus, K. Imaging lipid domains in cell membranes: The advent of super-resolution fluorescence microscopy. Front. Plant Sci. 2013, 4, 503. [Google Scholar] [CrossRef] [PubMed]
- Kusumi, A.; Koyama-Honda, I.; Suzuki, K. Molecular Dynamics and Interactions for Creation of Stimulation-Induced Stabilized Rafts from Small Unstable Steady-State Rafts. Traffic 2004, 5, 213–230. [Google Scholar] [CrossRef] [PubMed]
- Pike, L.J. Rafts defined: A report on the Keystone symposium on lipid rafts and cell function. J. Lipid Res. 2006, 47, 1597–1598. [Google Scholar] [CrossRef]
- Davis, J.H.; Clair, J.J.; Juhasz, J. Phase Equilibria in DOPC/DPPC-d62/Cholesterol Mixtures. Biophys. J. 2009, 96, 521–539. [Google Scholar] [CrossRef] [PubMed]
- Magenau, A.; Benzing, C.; Proschogo, N.; Don, A.S.; Hejazi, L.; Karunakaran, D.; Jessup, W.; Gaus, K. Phagocytosis of IgG-Coated Polystyrene Beads by Macrophages Induces and Requires High Membrane Order. Traffic 2011, 12, 1730–1743. [Google Scholar] [CrossRef] [PubMed]
- Kumar, M.; Ivanova, M.I.; Ramamoorthy, A. Non-micellar ganglioside GM1 induces an instantaneous conformational change in Aβ42 leading to the modulation of the peptide amyloid-fibril pathway. Biophys. Chem. 2023, 301, 107091. [Google Scholar] [CrossRef]
- Head, B.P.; Patel, H.H.; Insel, P.A. Interaction of membrane/lipid rafts with the cytoskeleton: Impact on signaling and function. Biochim. Biophys. Acta (BBA)-Biomembr. 2014, 1838, 532–545. [Google Scholar] [CrossRef] [PubMed]
- Laganowsky, A.; Reading, E.; Allison, T.M.; Ulmschneider, M.B.; Degiacomi, M.T.; Baldwin, A.J.; Robinson, C.V. Membrane proteins bind lipids selectively to modulate their structure and function. Nature 2014, 510, 172–175. [Google Scholar] [CrossRef] [PubMed]
- Parton, R.G.; Simons, K. The multiple faces of caveolae. Nat. Rev. Mol. Cell Biol. 2007, 8, 185–194. [Google Scholar] [CrossRef]
- Pereda, D.; Puertas-Avendaño, R.; Zjalic, M.; Heffer, M.; Gonzalez-Marrero, I.; González-Gómez, M.; Diaz, M.; Marin, R. A Closer Look At Neurotoxity. In Extracellular Propagation of Lipid Raft Molecular Species Involved in Parkinson’s Disease Neurodegeneration; Nova Science Publishers: New York, NY, USA, 2020; Chapter 2. [Google Scholar]
- Marques-Da-Silva, D.; Lagoa, R. Rafting on the Evidence for Lipid Raft-like Domains as Hubs Triggering Environmental Toxicants’ Cellular Effects. Molecules 2023, 28, 6598. [Google Scholar] [CrossRef]
- Pryor, S.; McCaffrey, G.; Young, L.R.; Grimes, M.L. NGF Causes TrkA to Specifically Attract Microtubules to Lipid Rafts. PLoS ONE 2012, 7, e35163. [Google Scholar] [CrossRef] [PubMed]
- Dupree, J.L.; Pomicter, A.D. Myelin, DIGs, and membrane rafts in the central nervous system. Prostaglandins Other Lipid Mediat. 2010, 91, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Gee, Y.J.; Sea, Y.L.; Lal, S.K. Viral modulation of lipid rafts and their potential as putative antiviral targets. Rev. Med. Virol. 2022, 33, e2413. [Google Scholar] [CrossRef]
- Viljetić, B.; Blažetić, S.; Labak, I.; Ivić, V.; Zjalić, M.; Heffer, M.; Balog, M. Lipid Rafts: The Maestros of Normal Brain Development. Biomolecules 2024, 14, 362. [Google Scholar] [CrossRef] [PubMed]
- Peeters, R.; Jellusova, J. Lipid metabolism in B cell biology. Mol. Oncol. 2023, 18, 1795–1813. [Google Scholar] [CrossRef]
- Mollinedo, F.; Gajate, C. Lipid rafts as signaling hubs in cancer cell survival/death and invasion: Implications in tumor progression and therapy. J. Lipid Res. 2020, 61, 611–635. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef] [PubMed]
- Murai, T. Lipid Raft-Mediated Regulation of Hyaluronan–CD44 Interactions in Inflammation and Cancer. Front. Immunol. 2015, 6, 420. [Google Scholar] [CrossRef] [PubMed]
- Norambuena, A.; Schwartz, M.A. Effects of integrin-mediated cell adhesion on plasma membrane lipid raft components and signaling. Mol. Biol. Cell 2011, 22, 3456–3464. [Google Scholar] [CrossRef]
- George, K.S.; Wu, S. Lipid raft: A floating island of death or survival. Toxicol. Appl. Pharmacol. 2012, 259, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Ribas, H.T.; Sogayar, M.C.; Dolga, A.M.; Winnischofer, S.M.; Trombetta-Lima, M. Lipid profile in breast cancer: From signaling pathways to treatment strategies. Biochimie 2023, 219, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Grassi, S.; Giussani, P.; Mauri, L.; Prioni, S.; Sonnino, S.; Prinetti, A. Lipid rafts and neurodegeneration: Structural and functional roles in physiologic aging and neurodegenerative diseases. J. Lipid Res. 2020, 61, 636–654. [Google Scholar] [CrossRef] [PubMed]
- Mirdha, L. Aggregation Behavior of Amyloid Beta Peptide Depends Upon the Membrane Lipid Composition. J. Membr. Biol. 2024, 257, 151–164. [Google Scholar] [CrossRef]
- Morgan, P.K.; Fang, L.; Lancaster, G.I.; Murphy, A.J. Hematopoiesis is regulated by cholesterol efflux pathways and lipid rafts: Connections with cardiovascular diseases. J. Lipid Res. 2020, 61, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Kulkarni, R.; Wiemer, E.A.C.; Chang, W. Role of Lipid Rafts in Pathogen-Host Interaction - A Mini Review. Front. Immunol. 2022, 12, 815020. [Google Scholar] [CrossRef] [PubMed]
- Dasgupta, A.; Gangai, S.; Narayan, R.; Kapoor, S. Mapping the Lipid Signatures in COVID-19 Infection: Diagnostic and Therapeutic Solutions. J. Med. Chem. 2023, 66, 14411–14433. [Google Scholar] [CrossRef] [PubMed]
- Sanders, D.W.; Jumper, C.C.; Ackerman, P.J.; Bracha, D.; Donlic, A.; Kim, H.; Kenney, D.; Castello-Serrano, I.; Suzuki, S.; Tamura, T.; et al. SARS-CoV-2 requires cholesterol for viral entry and pathological syncytia formation. eLife 2021, 10, e65962. [Google Scholar] [CrossRef] [PubMed]
- Heberle, F.A.; Petruzielo, R.S.; Pan, J.; Drazba, P.; Kučerka, N.; Standaert, R.F.; Feigenson, G.W.; Katsaras, J. Bilayer Thickness Mismatch Controls Domain Size in Model Membranes. J. Am. Chem. Soc. 2013, 135, 6853–6859. [Google Scholar] [CrossRef] [PubMed]
- Bunge, A.; Müller, P.; Stöckl, M.; Herrmann, A.; Huster, D. Characterization of the Ternary Mixture of Sphingomyelin, POPC, and Cholesterol: Support for an Inhomogeneous Lipid Distribution at High Temperatures. Biophys. J. 2008, 94, 2680–2690. [Google Scholar] [CrossRef] [PubMed]
- Veatch, S.L.; Soubias, O.; Keller, S.L.; Gawrisch, K. Critical fluctuations in domain-forming lipid mixtures. Proc. Natl. Acad. Sci. USA 2007, 104, 17650–17655. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, T.; Tsuchikawa, H.; Murata, M.; Matsumori, N. Deuterium NMR of Raft Model Membranes Reveals Domain-Specific Order Profiles and Compositional Distribution. Biophys. J. 2015, 108, 2502–2506. [Google Scholar] [CrossRef]
- Bhojoo, U.; Chen, M.; Zou, S. Temperature induced lipid membrane restructuring and changes in nanomechanics. Biochim. Biophys. Acta (BBA)-Biomembr. 2018, 1860, 700–709. [Google Scholar] [CrossRef]
- Sullan, R.M.A.; Li, J.K.; Hao, C.; Walker, G.C.; Zou, S. Cholesterol-Dependent Nanomechanical Stability of Phase-Segregated Multicomponent Lipid Bilayers. Biophys. J. 2010, 99, 507–516. [Google Scholar] [CrossRef]
- Drolle, E.; Negoda, A.; Hammond, K.; Pavlov, E.; Leonenko, Z. Changes in lipid membranes may trigger amyloid toxicity in Alzheimer’s disease. PLoS ONE 2017, 12, e0182194. [Google Scholar] [CrossRef]
- E Wolf, D.; Maynard, V.M.; A McKinnon, C.; Melchior, D.L. Lipid domains in the ram sperm plasma membrane demonstrated by differential scanning calorimetry. Proc. Natl. Acad. Sci. USA 1990, 87, 6893–6896. [Google Scholar] [CrossRef] [PubMed]
- de Almeida, R.F.; Fedorov, A.; Prieto, M. Sphingomyelin/Phosphatidylcholine/Cholesterol Phase Diagram: Boundaries and Composition of Lipid Rafts. Biophys. J. 2003, 85, 2406–2416. [Google Scholar] [CrossRef] [PubMed]
- de la Serna, J.B.; Perez-Gil, J.; Simonsen, A.C.; Bagatolli, L.A. Cholesterol Rules. J. Biol. Chem. 2004, 279, 40715–40722. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, J.W.; Conchello, J.-A. Fluorescence microscopy. Nat. Methods 2005, 2, 910–919. [Google Scholar] [CrossRef]
- Sezgin, E.; Schwille, P. Fluorescence Techniques to Study Lipid Dynamics. Cold Spring Harb. Perspect. Biol. 2011, 3, a009803. [Google Scholar] [CrossRef] [PubMed]
- Klymchenko, A.S.; Kreder, R. Fluorescent Probes for Lipid Rafts: From Model Membranes to Living Cells. Chem. Biol. 2014, 21, 97–113. [Google Scholar] [CrossRef]
- Klymchenko, A.S. Fluorescent Probes for Lipid Membranes: From the Cell Surface to Organelles. Acc. Chem. Res. 2022, 56, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, S.; Sancataldo, G.; Nielsen, H.M.; Foderà, V.; Vetri, V. Peptide–Membrane Interactions Monitored by Fluorescence Lifetime Imaging: A Study Case of Transportan 10. Langmuir 2021, 37, 13148–13159. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, S.; Sancataldo, G.; Baiamonte, C.; Pizzolanti, G.; Vetri, V. Transportan 10 Induces Perturbation and Pores Formation in Giant Plasma Membrane Vesicles Derived from Cancer Liver Cells. Biomolecules 2023, 13, 492. [Google Scholar] [CrossRef]
- Malacrida, L.; Gratton, E. LAURDAN fluorescence and phasor plots reveal the effects of a H2O2 bolus in NIH-3T3 fibroblast membranes dynamics and hydration. Free Radic. Biol. Med. 2018, 128, 144–156. [Google Scholar] [CrossRef] [PubMed]
- Merritt, E.A.; Sarfaty, S.; Akker, F.V.D.; L’Hoir, C.; Martial, J.A.; Hol, W.G. Crystal structure of cholera toxin B-pentamer bound to receptor GM1 pentasaccharide. Protein Sci. 1994, 3, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Mertz, J.A.; McCann, J.A.; Picking, W.D. Fluorescence Analysis of Galactose, Lactose, and Fucose Interaction with the Cholera Toxin B Subunit. Biochem. Biophys. Res. Commun. 1996, 226, 140–144. [Google Scholar] [CrossRef] [PubMed]
- Mikhalyov, I.; Samsonov, A. Lipid raft detecting in membranes of live erythrocytes. Biochim. Biophys. Acta (BBA)-Biomembr. 2011, 1808, 1930–1939. [Google Scholar] [CrossRef] [PubMed]
- Arthur, J.R.; Heinecke, K.A.; Seyfried, T.N. Filipin recognizes both GM1 and cholesterol in GM1 gangliosidosis mouse brain. J. Lipid Res. 2011, 52, 1345–1351. [Google Scholar] [CrossRef]
- Drabikowski, W.; Łagwińska, E.; Sarzała, M. Filipin as a fluorescent probe for the location of cholesterol in the membranes of fragmented sarcoplasmic reticulum. Biochim. Biophys. Acta (BBA)-Biomembr. 1973, 291, 61–70. [Google Scholar] [CrossRef]
- Harzer, K.; Kustermann-Kuhn, B. Quantified increases of cholesterol, total lipid and globotriaosylceramide in filipin-positive Niemann-Pick type C fibroblasts. Clin. Chim. Acta 2001, 305, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Robalo, J.R.; Canto, A.M.T.M.D.; Carvalho, A.J.P.; Ramalho, J.P.P.; Loura, L.M.S. Behavior of Fluorescent Cholesterol Analogues Dehydroergosterol and Cholestatrienol in Lipid Bilayers: A Molecular Dynamics Study. J. Phys. Chem. B 2013, 117, 5806–5819. [Google Scholar] [CrossRef] [PubMed]
- Sezgin, E.; Can, F.B.; Schneider, F.; Clausen, M.P.; Galiani, S.; Stanly, T.A.; Waithe, D.; Colaco, A.; Honigmann, A.; Wuestner, D.; et al. A comparative study on fluorescent cholesterol analogs as versatile cellular reporters. J. Lipid Res. 2016, 57, 299–309. [Google Scholar] [CrossRef]
- Pourmousa, M.; Róg, T.; Mikkeli, R.; Vattulainen, L.; Solanko, L.M.; Wüstner, D.; List, N.H.; Kongsted, J.; Karttunen, M. Dehydroergosterol as an Analogue for Cholesterol: Why It Mimics Cholesterol So Well—or Does It? J. Phys. Chem. B 2014, 118, 7345–7357. [Google Scholar] [CrossRef]
- Ramirez, D.M.C.; Ogilvie, W.W.; Johnston, L.J. NBD-cholesterol probes to track cholesterol distribution in model membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2010, 1798, 558–568. [Google Scholar] [CrossRef] [PubMed]
- Jurášek, M.; Valečka, J.; Novotný, I.; Kejík, Z.; Fähnrich, J.; Marešová, A.; Tauchen, J.; Bartůněk, P.; Dolenský, B.; Jakubek, M.; et al. Synthesis and biological evaluation of cationic TopFluor cholesterol analogues. Bioorg. Chem. 2021, 117, 105410. [Google Scholar] [CrossRef]
- Nusshold, C.; Uellen, A.; Bernhart, E.; Hammer, A.; Damm, S.; Wintersperger, A.; Reicher, H.; Hermetter, A.; Malle, E.; Sattler, W. Endocytosis and intracellular processing of BODIPY-sphingomyelin by murine CATH.a neurons. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2013, 1831, 1665–1678. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Ano, H.; Murata, M.; Shigetomi, K.; Ikenouchi, J.; Matsumori, N. Emphatic visualization of sphingomyelin-rich domains by inter-lipid FRET imaging using fluorescent sphingomyelins. Sci. Rep. 2017, 7, 16801. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, T.; Hunt, G.; Farkas, E.R.; Webb, W.W.; Feigenson, G.W. Fluorescence probe partitioning between Lo/Ld phases in lipid membranes. Biochim. Biophys. Acta (BBA)-Biomembr. 2007, 1768, 2182–2194. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, S.A.; Tricerri, M.A.; Gunther, G.; Gratton, E. Laurdan Generalized Polarization: From Cuvette to Microscope; ©FORMATEX: Budapest, Hungary, 2007. [Google Scholar]
- Parasassi, T.; De Stasio, G.; D’Ubaldo, A.; Gratton, E. Phase fluctuation in phospholipid membranes revealed by Laurdan fluorescence. Biophys. J. 1990, 57, 1179–1186. [Google Scholar] [CrossRef]
- Parasassi, T.; Krasnowska, E.K.; Bagatolli, L.; Gratton, E. Laurdan and Prodan as Polarity-Sensitive Fluorescent Membrane Probes. J. Fluoresc. 1998, 8, 365–373. [Google Scholar] [CrossRef]
- Tamai, N.; Matsuki, H.; Goto, M. Phase Imaging of Phosphatidylcholine Bilayer Membranes by Prodan Fluorescence. Membranes 2022, 12, 1219. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.M.; Choo, H.; Jung, S.; Ko, Y.; Park, W.; Jeon, S.; Kim, C.H.; Joo, T.; Cho, B.R. A Two-Photon Fluorescent Probe for Lipid Raft Imaging: C-Laurdan. ChemBioChem 2007, 8, 553–559. [Google Scholar] [CrossRef] [PubMed]
- Jin, L.; Millard, A.C.; Wuskell, J.P.; Dong, X.; Wu, D.; Clark, H.A.; Loew, L.M. Characterization and Application of a New Optical Probe for Membrane Lipid Domains. Biophys. J. 2006, 90, 2563–2575. [Google Scholar] [CrossRef]
- Zhanghao, K.; Liu, W.; Li, M.; Wu, Z.; Wang, X.; Chen, X.; Shan, C.; Wang, H.; Chen, X.; Dai, Q.; et al. High-dimensional super-resolution imaging reveals heterogeneity and dynamics of subcellular lipid membranes. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kucherak, O.A.; Oncul, S.; Darwich, Z.; Yushchenko, D.A.; Arntz, Y.; Didier, P.; Mély, Y.; Klymchenko, A.S. Switchable Nile Red-Based Probe for Cholesterol and Lipid Order at the Outer Leaflet of Biomembranes. J. Am. Chem. Soc. 2010, 132, 4907–4916. [Google Scholar] [CrossRef] [PubMed]
- Klymchenko, A.S.; Oncul, S.; Didier, P.; Schaub, E.; Bagatolli, L.; Duportail, G.; Mély, Y. Visualization of lipid domains in giant unilamellar vesicles using an environment-sensitive membrane probe based on 3-hydroxyflavone. Biochim. Biophys. Acta (BBA)-Biomembr. 2008, 1788, 495–499. [Google Scholar] [CrossRef]
- Kim, H.M.; Jeong, B.H.; Hyon, J.-Y.; An, M.J.; Seo, M.S.; Hong, J.H.; Lee, K.J.; Kim, C.H.; Joo, T.; Hong, S.-C.; et al. Two-Photon Fluorescent Turn-On Probe for Lipid Rafts in Live Cell and Tissue. J. Am. Chem. Soc. 2008, 130, 4246–4247. [Google Scholar] [CrossRef] [PubMed]
- Lim, C.S.; Kim, H.J.; Lee, J.H.; Tian, Y.S.; Kim, C.H.; Kim, H.M.; Joo, T.; Cho, B.R. A Two-Photon Turn-On Probe for Lipid Rafts with Minimum Internalization. ChemBioChem 2010, 12, 392–395. [Google Scholar] [CrossRef] [PubMed]
- Haidekker, M.A.; Ling, T.; Anglo, M.; Stevens, H.Y.; Frangos, J.A.; Theodorakis, E.A. New fluorescent probes for the measurement of cell membrane viscosity. Chem. Biol. 2001, 8, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Mika, J.T.; Thompson, A.J.; Dent, M.R.; Brooks, N.J.; Michiels, J.; Hofkens, J.; Kuimova, M.K. Measuring the Viscosity of the Escherichia coli Plasma Membrane Using Molecular Rotors. Biophys. J. 2016, 111, 1528–1540. [Google Scholar] [CrossRef]
- Polita, A.; Stancikaitė, M.; Žvirblis, R.; Maleckaitė, K.; Dodonova-Vaitkūnienė, J.; Tumkevičius, S.; Shivabalan, A.P.; Valinčius, G. Designing a green-emitting viscosity-sensitive 4,4-difluoro-4-bora-3a,4a-diaza-s-indacene (BODIPY) probe for plasma membrane viscosity imaging. RSC Adv. 2023, 13, 19257–19264. [Google Scholar] [CrossRef]
- Rumble, C.; Rich, K.; He, G.; Maroncelli, M. CCVJ Is Not a Simple Rotor Probe. J. Phys. Chem. A 2012, 116, 10786–10792. [Google Scholar] [CrossRef]
- Blank, N.; Schiller, M.; Krienke, S.; Wabnitz, G.; Ho, A.D.; Lorenz, H. Cholera toxin binds to lipid rafts but has a limited specificity for ganglioside GM1. Immunol. Cell Biol. 2007, 85, 378–382. [Google Scholar] [CrossRef] [PubMed]
- Cervin, J.; Boucher, A.; Youn, G.; Björklund, P.; Wallenius, V.; Mottram, L.; Sampson, N.S.; Yrlid, U. Fucose-Galactose Polymers Inhibit Cholera Toxin Binding to Fucosylated Structures and Galactose-Dependent Intoxication of Human Enteroids. ACS Infect. Dis. 2020, 6, 1192–1203. [Google Scholar] [CrossRef] [PubMed]
- Boldyrev, I.A.; Zhai, X.; Momsen, M.M.; Brockman, H.L.; Brown, R.E.; Molotkovsky, J.G. New BODIPY lipid probes for fluorescence studies of membranes. J. Lipid Res. 2007, 48, 1518–1532. [Google Scholar] [CrossRef]
- Wüstner, D. Fluorescent sterols as tools in membrane biophysics and cell biology. Chem. Phys. Lipids 2007, 146, 1–25. [Google Scholar] [CrossRef] [PubMed]
- Bruno, F.; Camuso, S.; Capuozzo, E.; Canterini, S. The Antifungal Antibiotic Filipin as a Diagnostic Tool of Cholesterol Alterations in Lysosomal Storage Diseases and Neurodegenerative Disorders. Antibiotics 2023, 12, 122. [Google Scholar] [CrossRef]
- Orlandi, P.A.; Fishman, P.H. Filipin-dependent Inhibition of Cholera Toxin: Evidence for Toxin Internalization and Activation through Caveolae-like Domains. J. Cell Biol. 1998, 141, 905–915. [Google Scholar] [CrossRef] [PubMed]
- Ariola, F.S.; Li, Z.; Cornejo, C.; Bittman, R.; Heikal, A.A. Membrane Fluidity and Lipid Order in Ternary Giant Unilamellar Vesicles Using a New Bodipy-Cholesterol Derivative. Biophys. J. 2009, 96, 2696–2708. [Google Scholar] [CrossRef] [PubMed]
- Juhasz, J.; Davis, J.H.; Sharom, F.J. Fluorescent probe partitioning in giant unilamellar vesicles of ‘lipid raft’ mixtures. Biochem. J. 2010, 430, 415–423. [Google Scholar] [CrossRef] [PubMed]
- Klymchenko, A.S. Solvatochromic and Fluorogenic Dyes as Environment-Sensitive Probes: Design and Biological Applications. Acc. Chem. Res. 2017, 50, 366–375. [Google Scholar] [CrossRef] [PubMed]
- Gunther, G.; Malacrida, L.; Jameson, D.M.; Gratton, E.; Sánchez, S.A. LAURDAN since Weber: The Quest for Visualizing Membrane Heterogeneity. Acc. Chem. Res. 2021, 54, 976–987. [Google Scholar] [CrossRef] [PubMed]
- van Maarschalkerweerd, A.; Vetri, V.; Langkilde, A.E.; Foderà, V.; Vestergaard, B. Protein/Lipid Coaggregates are Formed During α-Synuclein-Induced Disruption of Lipid Bilayers. Biomacromolecules 2014, 15, 3643–3654. [Google Scholar] [CrossRef]
- Owen, D.M.; Williamson, D.J.; Magenau, A.; Gaus, K. Sub-resolution lipid domains exist in the plasma membrane and regulate protein diffusion and distribution. Nat. Commun. 2012, 3, 1256. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, S.; Sancataldo, G.; Foderà, V.; Vetri, V. α-casein micelles-membranes interaction: Flower-like lipid protein coaggregates formation. Biochim. Biophys. Acta (BBA)-Gen. Subj. 2022, 1866, 130196. [Google Scholar] [CrossRef] [PubMed]
- Gaus, K.; Gratton, E.; Kable, E.P.W.; Jones, A.S.; Gelissen, I.; Kritharides, L.; Jessup, W. Visualizing lipid structure and raft domains in living cells with two-photon microscopy. Proc. Natl. Acad. Sci. USA 2003, 100, 15554–15559. [Google Scholar] [CrossRef] [PubMed]
- Vetri, V.; Ossato, G.; Militello, V.; Digman, M.; Leone, M.; Gratton, E. Fluctuation Methods To Study Protein Aggregation in Live Cells: Concanavalin A Oligomers Formation. Biophys. J. 2011, 100, 774–783. [Google Scholar] [CrossRef] [PubMed]
- Traian, M.M.D.; Flecha, F.L.G.; Levi, V. Imaging lipid lateral organization in membranes with C-laurdan in a confocal microscope. J. Lipid Res. 2012, 53, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.M.; Rentero, C.; Magenau, A.; Abu-Siniyeh, A.; Gaus, K. Quantitative imaging of membrane lipid order in cells and organisms. Nat. Protoc. 2011, 7, 24–35. [Google Scholar] [CrossRef] [PubMed]
- Panconi, L.; Euchner, J.; Tashev, S.A.; Makarova, M.; Herten, D.-P.; Owen, D.M.; Nieves, D.J. Mapping membrane biophysical nano-environments. Nat. Commun. 2024, 15, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Amaro, M.; Reina, F.; Hof, M.; Eggeling, C.; Sezgin, E. Laurdan and Di-4-ANEPPDHQ probe different properties of the membrane. J. Phys. D Appl. Phys. 2017, 50, 134004. [Google Scholar] [CrossRef] [PubMed]
- Dinic, J.; Biverståhl, H.; Mäler, L.; Parmryd, I. Laurdan and di-4-ANEPPDHQ do not respond to membrane-inserted peptides and are good probes for lipid packing. Biochim. Biophys. Acta (BBA)-Biomembr. 2010, 1808, 298–306. [Google Scholar] [CrossRef]
- Mukherjee, S.; Raghuraman, H.; Chattopadhyay, A. Membrane localization and dynamics of Nile Red: Effect of cholesterol. Biochim. Biophys. Acta (BBA)-Biomembr. 2006, 1768, 59–66. [Google Scholar] [CrossRef] [PubMed]
- Sackett, D.L.; Wolff, J. Nile red as a polarity-sensitive fluorescent probe of hydrophobic protein surfaces. Anal. Biochem. 1987, 167, 228–234. [Google Scholar] [CrossRef] [PubMed]
- Bondelli, G.; Paternò, G.M.; Lanzani, G. (INVITED) Fluorescent probes for optical investigation of the plasma membrane. Opt. Mater. X 2021, 12. [Google Scholar] [CrossRef]
- Paez-Perez, M.; Kuimova, M.K. Molecular Rotors: Fluorescent Sensors for Microviscosity and Conformation of Biomolecules. Angew. Chem. Int. Ed. 2023, 63, e202311233. [Google Scholar] [CrossRef] [PubMed]
- Dent, M.R.; López-Duarte, I.; Dickson, C.J.; Geoghegan, N.D.; Cooper, J.M.; Gould, I.R.; Krams, R.; Bull, J.A.; Brooks, N.J.; Kuimova, M.K. Imaging phase separation in model lipid membranes through the use of BODIPY based molecular rotors. Phys. Chem. Chem. Phys. 2015, 17, 18393–18402. [Google Scholar] [CrossRef] [PubMed]
- López-Duarte, I.; Vu, T.T.; Izquierdo, M.A.; Bull, J.A.; Kuimova, M.K. A molecular rotor for measuring viscosity in plasma membranes of live cells. Chem. Commun. 2013, 50, 5282–5284. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, N.; Silswal, A.; Koner, A.L. Julolidine-based fluorescent molecular rotor: A versatile tool for sensing and diagnosis. Sens. Diagn. 2024, 3, 585–598. [Google Scholar] [CrossRef]
- Haidekker, M.A.; Theodorakis, E.A. Molecular rotors—fluorescent biosensors for viscosity and flow. Org. Biomol. Chem. 2007, 5, 1669–1678. [Google Scholar] [CrossRef] [PubMed]
- Levental, I.; Veatch, S.L. The Continuing Mystery of Lipid Rafts. J. Mol. Biol. 2016, 428, 4749–4764. [Google Scholar] [CrossRef]
- Samsonov, A.V.; Mihalyov, I.; Cohen, F.S. Characterization of Cholesterol-Sphingomyelin Domains and Their Dynamics in Bilayer Membranes. Biophys. J. 2001, 81, 1486–1500. [Google Scholar] [CrossRef] [PubMed]
- Sticker, M.; Elsässer, R.; Neumann, M.; Wolff, H. How to Get Better Fluorescence Images with Your Widefield Microscope: A Methodology Review. Microsc. Today 2020, 28, 36–43. [Google Scholar] [CrossRef]
- Schneckenburger, H.; Richter, V. Laser Scanning versus Wide-Field—Choosing the Appropriate Microscope in Life Sciences. Appl. Sci. 2021, 11, 733. [Google Scholar] [CrossRef]
- Wang, S.; Larina, I.V. High-Resolution Imaging Techniques in Tissue Engineering. Monit. Eval. Biomater. Their Perform. In Vivo 2017, 151–180. [Google Scholar] [CrossRef]
- Sanderson, M.J.; Smith, I.; Parker, I.; Bootman, M.D. Fluorescence Microscopy. Cold Spring Harb. Protoc. 2014, 2014, 1042–1065. [Google Scholar] [CrossRef] [PubMed]
- Elliott, A.D. Confocal Microscopy: Principles and Modern Practices. Curr. Protoc. Cytom. 2019, 92, e68. [Google Scholar] [CrossRef] [PubMed]
- Korlach, J.; Schwille, P.; Webb, W.W.; Feigenson, G.W. Characterization of lipid bilayer phases by confocal microscopy and fluorescence correlation spectroscopy. Proc. Natl. Acad. Sci. USA 1999, 96, 8461–8466. [Google Scholar] [CrossRef] [PubMed]
- Shen, Y.; Wei, L.; Min, W. Raman Imaging Reveals Insights into Membrane Phase Biophysics in Cells. J. Phys. Chem. B 2023, 127, 6233–6240. [Google Scholar] [CrossRef] [PubMed]
- Van Meer, G.; Voelker, D.R.; Feigenson, G.W. Membrane lipids: Where they are and how they behave. Nat. Rev. Mol. Cell Biol. 2008, 9, 112–124. [Google Scholar] [CrossRef]
- Hao, M.; Mukherjee, S.; Maxfield, F.R. Cholesterol depletion induces large scale domain segregation in living cell membranes. Proc. Natl. Acad. Sci. USA 2001, 98, 13072–13077. [Google Scholar] [CrossRef] [PubMed]
- Baumgart, T.; Hammond, A.T.; Sengupta, P.; Hess, S.T.; Holowka, D.A.; Baird, B.A.; Webb, W.W. Large-scale fluid/fluid phase separation of proteins and lipids in giant plasma membrane vesicles. Proc. Natl. Acad. Sci. USA 2007, 104, 3165–3170. [Google Scholar] [CrossRef] [PubMed]
- Kahya, N.; Scherfeld, D.; Bacia, K.; Poolman, B.; Schwille, P. Probing Lipid Mobility of Raft-exhibiting Model Membranes by Fluorescence Correlation Spectroscopy. J. Biol. Chem. 2003, 278, 28109–28115. [Google Scholar] [CrossRef] [PubMed]
- Svoboda, K.; Yasuda, R. Principles of Two-Photon Excitation Microscopy and Its Applications to Neuroscience. Neuron 2006, 50, 823–839. [Google Scholar] [CrossRef] [PubMed]
- Bagatolli, L.; Gratton, E. Two Photon Fluorescence Microscopy of Coexisting Lipid Domains in Giant Unilamellar Vesicles of Binary Phospholipid Mixtures. Biophys. J. 2000, 78, 290–305. [Google Scholar] [CrossRef] [PubMed]
- Dietrich, C.; Bagatolli, L.; Volovyk, Z.; Thompson, N.; Levi, M.; Jacobson, K.; Gratton, E. Lipid Rafts Reconstituted in Model Membranes. Biophys. J. 2001, 80, 1417–1428. [Google Scholar] [CrossRef]
- Kaiser, H.-J.; Lingwood, D.; Levental, I.; Sampaio, J.L.; Kalvodova, L.; Rajendran, L.; Simons, K. Order of lipid phases in model and plasma membranes. Proc. Natl. Acad. Sci. USA 2009, 106, 16645–16650. [Google Scholar] [CrossRef] [PubMed]
- Gasecka, A.; Han, T.-J.; Favard, C.; Cho, B.R.; Brasselet, S. Quantitative Imaging of Molecular Order in Lipid Membranes Using Two-Photon Fluorescence Polarimetry. Biophys. J. 2009, 97, 2854–2862. [Google Scholar] [CrossRef] [PubMed]
- Patterson, G.H.; Piston, D.W. Photobleaching in Two-Photon Excitation Microscopy. Biophys. J. 2000, 78, 2159–2162. [Google Scholar] [CrossRef] [PubMed]
- Drummond, D.R.; Carter, N.; Cross, R.A. Multiphoton versus confocal high resolution z-sectioning of enhanced green fluorescent microtubules: Increased multiphoton photobleaching within the focal plane can be compensated using a Pockels cell and dual widefield detectors. J. Microsc. 2002, 206, 161–169. [Google Scholar] [CrossRef]
- Diaspro, A.; Chirico, G.; Collini, M. Two-photon fluorescence excitation and related techniques in biological microscopy. Q. Rev. Biophys. 2005, 38, 97–166. [Google Scholar] [CrossRef]
- Lorén, N.; Hagman, J.; Jonasson, J.K.; Deschout, H.; Bernin, D.; Cella-Zanacchi, F.; Diaspro, A.; McNally, J.G.; Ameloot, M.; Smisdom, N.; et al. Fluorescence recovery after photobleaching in material and life sciences: Putting theory into practice. Q. Rev. Biophys. 2015, 48, 323–387. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, C.A.; James, P.S.; Mackie, A.R.; Ladha, S.; Jones, R. Regionalized lipid diffusion in the plasma membrane of mammalian spermatozoa. Biol. Reprod. 1998, 59, 1506–1514. [Google Scholar] [CrossRef] [PubMed]
- Niv, H.; Gutman, O.; Henis, Y.I.; Kloog, Y. Membrane Interactions of a Constitutively Active GFP-Ki-Ras 4B and Their Role in Signaling. J. Biol. Chem. 1999, 274, 1606–1613. [Google Scholar] [CrossRef] [PubMed]
- Roy, S.; Plowman, S.; Rotblat, B.; Prior, I.A.; Muncke, C.; Grainger, S.; Parton, R.G.; Henis, Y.I.; Kloog, Y.; Hancock, J.F. Individual Palmitoyl Residues Serve Distinct Roles in H-Ras Trafficking, Microlocalization, and Signaling. Mol. Cell. Biol. 2005, 25, 6722–6733. [Google Scholar] [CrossRef] [PubMed]
- Kenworthy, A.K.; Nichols, B.J.; Remmert, C.L.; Hendrix, G.M.; Kumar, M.; Zimmerberg, J.; Lippincott-Schwartz, J. Dynamics of putative raft-associated proteins at the cell surface. J. Cell Biol. 2004, 165, 735–746. [Google Scholar] [CrossRef] [PubMed]
- Meder, D.; Moreno, M.J.; Verkade, P.; Vaz, W.L.C.; Simons, K. Phase coexistence and connectivity in the apical membrane of polarized epithelial cells. Proc. Natl. Acad. Sci. USA 2006, 103, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Kenworthy, A.K. Complex Applications of Simple FRAP on Membranes. Biomembr. Front. 2009, 187–221. [Google Scholar] [CrossRef]
- Rao, M.; Mayor, S. Use of Forster’s resonance energy transfer microscopy to study lipid rafts. Biochim. Biophys. Acta (BBA)-Mol. Cell Res. 2005, 1746, 221–233. [Google Scholar] [CrossRef] [PubMed]
- Feigenson, G.W.; Buboltz, J.T. Ternary Phase Diagram of Dipalmitoyl-PC/Dilauroyl-PC/Cholesterol: Nanoscopic Domain Formation Driven by Cholesterol. Biophys. J. 2001, 80, 2775–2788. [Google Scholar] [CrossRef] [PubMed]
- Heberle, F.A.; Wu, J.; Goh, S.L.; Petruzielo, R.S.; Feigenson, G.W. Comparison of Three Ternary Lipid Bilayer Mixtures: FRET and ESR Reveal Nanodomains. Biophys. J. 2010, 99, 3309–3318. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Holowka, D.; Baird, B. Fluorescence Resonance Energy Transfer between Lipid Probes Detects Nanoscopic Heterogeneity in the Plasma Membrane of Live Cells. Biophys. J. 2007, 92, 3564–3574. [Google Scholar] [CrossRef] [PubMed]
- Berney, C.; Danuser, G. FRET or No FRET: A Quantitative Comparison. Biophys. J. 2003, 84, 3992–4010. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, A.; Sharma, A.; Chizhik, A.I.; Isbaner, S.; Ruhlandt, D.; Tsukanov, R.; Gregor, I.; Karedla, N.; Enderlein, J. Graphene-based metal-induced energy transfer for sub-nanometre optical localization. Nat. Photonics 2019, 13, 860–865. [Google Scholar] [CrossRef]
- Chatterjee, A.; Ruturaj; Chakraborty, M.P.; Nandi, S.; Purkayastha, P. Biocompatible and optically stable hydrophobic fluorescent carbon dots for isolation and imaging of lipid rafts in model membrane. Anal. Bioanal. Chem. 2022, 414, 6055–6067. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Ghosh, A.; Enderlein, J. Cholesterol-Induced Nanoscale Variations in the Thickness of Phospholipid Membranes. Nano Lett. 2023, 23, 2421–2426. [Google Scholar] [CrossRef] [PubMed]
- Raja, S.O.; Chizhik, A.I.; Schmidt, C.F.; Enderlein, J.; Ghosh, A. Mapping Activity-Dependent Quasi-stationary States of Mitochondrial Membranes with Graphene-Induced Energy Transfer Imaging. Nano Lett. 2021, 21, 8244–8249. [Google Scholar] [CrossRef] [PubMed]
- A Lauterbach, M. Finding, defining and breaking the diffraction barrier in microscopy—A historical perspective. Opt. Nanoscopy 2012, 1, 8. [Google Scholar] [CrossRef]
- Liu, S.; Hoess, P.; Ries, J. Super-Resolution Microscopy for Structural Cell Biology. Annu. Rev. Biophys. 2022, 51, 301–326. [Google Scholar] [CrossRef] [PubMed]
- Sydor, A.M.; Czymmek, K.J.; Puchner, E.M.; Mennella, V. Super-Resolution Microscopy: From Single Molecules to Supramolecular Assemblies. Trends Cell Biol. 2015, 25, 730–748. [Google Scholar] [CrossRef] [PubMed]
- Vicidomini, G.; Bianchini, P.; Diaspro, A. STED super-resolved microscopy. Nat. Methods 2018, 15, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Xu, X.; Xi, P. Stimulated emission depletion microscopy for biological imaging in four dimensions: A review. Microsc. Res. Tech. 2021, 84, 1947–1958. [Google Scholar] [CrossRef] [PubMed]
- Lorizate, M.; Terrones, O.; Nieto-Garai, J.A.; Rojo-Bartolomé, I.; Ciceri, D.; Morana, O.; Olazar-Intxausti, J.; Arboleya, A.; Martin, A.; Szynkiewicz, M.; et al. Super-Resolution Microscopy Using a Bioorthogonal-Based Cholesterol Probe Provides Unprecedented Capabilities for Imaging Nanoscale Lipid Heterogeneity in Living Cells. Small Methods 2021, 5, 2100430. [Google Scholar] [CrossRef] [PubMed]
- Lingwood, D.; Simons, K. Lipid Rafts As a Membrane-Organizing Principle. Science 2010, 327, 46–50. [Google Scholar] [CrossRef]
- Heintzmann, R.; Huser, T. Super-Resolution Structured Illumination Microscopy. Chem. Rev. 2017, 117, 13890–13908. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.C.N.; Oddos, S.; Dobbie, I.M.; Alakoskela, J.-M.; Parton, R.M.; Eissmann, P.; Neil, M.A.A.; Dunsby, C.; French, P.M.W.; Davis, I.; et al. Remodelling of Cortical Actin Where Lytic Granules Dock at Natural Killer Cell Immune Synapses Revealed by Super-Resolution Microscopy. PLoS Biol. 2011, 9, e1001152. [Google Scholar] [CrossRef] [PubMed]
- Manton, J.D. Answering some questions about structured illumination microscopy. Philos. Trans. R. Soc. A Math. Phys. Eng. Sci. 2022, 380, 20210109. [Google Scholar] [CrossRef] [PubMed]
- Lelek, M.; Gyparaki, M.T.; Beliu, G.; Schueder, F.; Griffié, J.; Manley, S.; Jungmann, R.; Sauer, M.; Lakadamyali, M.; Zimmer, C. Single-molecule localization microscopy. Nat. Rev. Methods Prim. 2021, 1, 1–27. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Ma, H.; Liu, Y. Stochastic Optical Reconstruction Microscopy (STORM). Curr. Protoc. Cytom. 2017, 81, 12.46.1–12.46.27. [Google Scholar] [CrossRef] [PubMed]
- Hugelier, S.; Colosi, P.; Lakadamyali, M. Quantitative Single-Molecule Localization Microscopy. Annu. Rev. Biophys. 2023, 52, 139–160. [Google Scholar] [CrossRef] [PubMed]
- Sengupta, P.; Jovanovic-Talisman, T.; Skoko, D.; Renz, M.; Veatch, S.L.; Lippincott-Schwartz, J. Probing protein heterogeneity in the plasma membrane using PALM and pair correlation analysis. Nat. Methods 2011, 8, 969–975. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.M.; Rentero, C.; Rossy, J.; Magenau, A.; Williamson, D.; Rodriguez, M.; Gaus, K. PALM imaging and cluster analysis of protein heterogeneity at the cell surface. J. Biophotonics 2010, 3, 446–454. [Google Scholar] [CrossRef] [PubMed]
- Winkelmann, H.; Richter, C.P.; Eising, J.; Piehler, J.; Kurre, R. Correlative single-molecule and structured illumination microscopy of fast dynamics at the plasma membrane. Nat. Commun. 2024, 15, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Sharonov, A.; Hochstrasser, R.M. Wide-field subdiffraction imaging by accumulated binding of diffusing probes. Proc. Natl. Acad. Sci. USA 2006, 103, 18911–18916. [Google Scholar] [CrossRef] [PubMed]
- Winckler, P.; Lartigue, L.; Giannone, G.; De Giorgi, F.; Ichas, F.; Sibarita, J.-B.; Lounis, B.; Cognet, L. Identification and super-resolution imaging of ligand-activated receptor dimers in live cells. Sci. Rep. 2013, 3, srep02387. [Google Scholar] [CrossRef]
- Balzarotti, F.; Eilers, Y.; Gwosch, K.C.; Gynnå, A.H.; Westphal, V.; Stefani, F.D.; Elf, J.; Hell, S.W. Nanometer resolution imaging and tracking of fluorescent molecules with minimal photon fluxes. Science 2017, 355, 606–612. [Google Scholar] [CrossRef] [PubMed]
- Eilers, Y.; Ta, H.; Gwosch, K.C.; Balzarotti, F.; Hell, S.W. MINFLUX monitors rapid molecular jumps with superior spatiotemporal resolution. Proc. Natl. Acad. Sci. USA 2018, 115, 6117–6122. [Google Scholar] [CrossRef] [PubMed]
- Eggeling, C.; Ringemann, C.; Medda, R.; Schwarzmann, G.; Sandhoff, K.; Polyakova, S.; Belov, V.N.; Hein, B.; von Middendorff, C.; Schönle, A.; et al. Direct observation of the nanoscale dynamics of membrane lipids in a living cell. Nature 2008, 457, 1159–1162. [Google Scholar] [CrossRef] [PubMed]
- Shen, H.; Tauzin, L.J.; Baiyasi, R.; Wang, W.; Moringo, N.; Shuang, B.; Landes, C.F. Single Particle Tracking: From Theory to Biophysical Applications. Chem. Rev. 2017, 117, 7331–7376. [Google Scholar] [CrossRef] [PubMed]
- Ries, J.; Schwille, P. Fluorescence correlation spectroscopy. BioEssays 2012, 34, 361–368. [Google Scholar] [CrossRef] [PubMed]
- Alcor, D.; Gouzer, G.; Triller, A. Single-particle tracking methods for the study of membrane receptors dynamics. Eur. J. Neurosci. 2009, 30, 987–997. [Google Scholar] [CrossRef] [PubMed]
- Bouzigues, C.; Morel, M.; Triller, A.; Dahan, M. Asymmetric redistribution of GABA receptors during GABA gradient sensing by nerve growth cones analyzed by single quantum dot imaging. Proc. Natl. Acad. Sci. USA 2007, 104, 11251–11256. [Google Scholar] [CrossRef] [PubMed]
- Dahan, M.; Lévi, S.; Luccardini, C.; Rostaing, P.; Riveau, B.; Triller, A. Diffusion Dynamics of Glycine Receptors Revealed by Single-Quantum Dot Tracking. Science 2003, 302, 442–445. [Google Scholar] [CrossRef]
- Mashanov, G.I.; Nenasheva, T.A.; Mashanova, A.; Lape, R.; Birdsall, N.J.M.; Sivilotti, L.; Molloy, J.E. Heterogeneity of cell membrane structure studied by single molecule tracking. Faraday Discuss. 2021, 232, 358–374. [Google Scholar] [CrossRef]
- Bacia, K.; Scherfeld, D.; Kahya, N.; Schwille, P. Fluorescence Correlation Spectroscopy Relates Rafts in Model and Native Membranes. Biophys. J. 2004, 87, 1034–1043. [Google Scholar] [CrossRef] [PubMed]
- He, H.-T.; Marguet, D. Detecting Nanodomains in Living Cell Membrane by Fluorescence Correlation Spectroscopy. Annu. Rev. Phys. Chem. 2011, 62, 417–436. [Google Scholar] [CrossRef]
- Brown, C.M.; Dalal, R.B.; Hebert, B.; Digman, M.A.; Horwitz, A.R.; Gratton, E. Raster image correlation spectroscopy (RICS) for measuring fast protein dynamics and concentrations with a commercial laser scanning confocal microscope. J. Microsc. 2007, 229, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Gielen, E.; Smisdom, N.; Vandeven, M.; De Clercq, B.; Gratton, E.; Digman, M.; Rigo, J.-M.; Hofkens, J.; Engelborghs, Y.; Ameloot, M. Measuring Diffusion of Lipid-like Probes in Artificial and Natural Membranes by Raster Image Correlation Spectroscopy (RICS): Use of a Commercial Laser-Scanning Microscope with Analog Detection. Langmuir 2009, 25, 5209–5218. [Google Scholar] [CrossRef]
- Stöckl, M.T.; Bizzarri, R.; Subramaniam, V. Studying Membrane Properties Using Fluorescence Lifetime Imaging Microscopy (FLIM). In Fluorescent Methods to Study Biological Membranes, Springer Series on Fluorescence (Methods and Applications), 1st ed.; Mély, Y., Duportail, G., Eds.; Springer: Berlin/Heidelberg, Germany, 2013; Volume 13, pp. 215–240. [Google Scholar] [CrossRef]
- Mueller, V.; Ringemann, C.; Honigmann, A.; Schwarzmann, G.; Medda, R.; Leutenegger, M.; Polyakova, S.; Belov, V.; Hell, S.; Eggeling, C. STED Nanoscopy Reveals Molecular Details of Cholesterol- and Cytoskeleton-Modulated Lipid Interactions in Living Cells. Biophys. J. 2011, 101, 1651–1660. [Google Scholar] [CrossRef] [PubMed]
- Sankaran, J.; Wohland, T. Fluorescence strategies for mapping cell membrane dynamics and structures. APL Bioeng. 2020, 4, 020901. [Google Scholar] [CrossRef] [PubMed]
- Fecchi, K.; Anticoli, S.; Peruzzu, D.; Iessi, E.; Gagliardi, M.C.; Matarrese, P.; Ruggieri, A. Coronavirus Interplay With Lipid Rafts and Autophagy Unveils Promising Therapeutic Targets. Front. Microbiol. 2020, 11, 1821. [Google Scholar] [CrossRef] [PubMed]
- Sviridov, D.; Miller, Y.I.; Ballout, R.A.; Remaley, A.T.; Bukrinsky, M. Targeting Lipid Rafts—A Potential Therapy for COVID-19. Front. Immunol. 2020, 11, 574508. [Google Scholar] [CrossRef] [PubMed]
- Cordy, J.M.; Hussain, I.; Dingwall, C.; Hooper, N.M.; Turner, A.J. Exclusively targeting β-secretase to lipid rafts by GPI-anchor addition up-regulates β-site processing of the amyloid precursor protein. Proc. Natl. Acad. Sci. USA 2003, 100, 11735–11740. [Google Scholar] [CrossRef] [PubMed]
- Cordy, J.M.; Hooper, N.M.; Turner, A.J. The involvement of lipid rafts in Alzheimer’s disease (Review). Mol. Membr. Biol. 2006, 23, 111–122. [Google Scholar] [CrossRef] [PubMed]
- Simons, K.; Ehehalt, R. Cholesterol, lipid rafts, and disease. J. Clin. Investig. 2002, 110, 597–603. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Kulas, J.A.; Wang, C.; Holtzman, D.M.; Ferris, H.A.; Hansen, S.B. Regulation of beta-amyloid production in neurons by astrocyte-derived cholesterol. Proc. Natl. Acad. Sci. USA 2021, 118, e2102191118. [Google Scholar] [CrossRef] [PubMed]
- Mesa-Herrera, F.; Taoro-González, L.; Valdés-Baizabal, C.; Diaz, M.; Marín, R. Lipid and Lipid Raft Alteration in Aging and Neurodegenerative Diseases: A Window for the Development of New Biomarkers. Int. J. Mol. Sci. 2019, 20, 3810. [Google Scholar] [CrossRef] [PubMed]
- Badawy, S.M.M.; Okada, T.; Kajimoto, T.; Hirase, M.; Matovelo, S.A.; Nakamura, S.; Yoshida, D.; Ijuin, T.; Nakamura, S.-I. Extracellular α-synuclein drives sphingosine 1-phosphate receptor subtype 1 out of lipid rafts, leading to impaired inhibitory G-protein signaling. J. Biol. Chem. 2018, 293, 8208–8216. [Google Scholar] [CrossRef]
- Handlin, L.J.; Dai, G. Direct regulation of the voltage sensor of HCN channels by membrane lipid compartmentalization. Nat. Commun. 2023, 14, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Dai, G. Neuronal KCNQ2/3 channels are recruited to lipid raft microdomains by palmitoylation of BACE1. J. Gen. Physiol. 2022, 154. [Google Scholar] [CrossRef]
- Chochina, S.; Avdulov, N.; Igbavboa, U.; Cleary, J.; O’Hare, E.; Wood, W. Amyloid β-peptide1-40 increases neuronal membrane fluidity: Role of cholesterol and brain region. J. Lipid Res. 2001, 42, 1292–1297. [Google Scholar] [CrossRef] [PubMed]
- Müller, W.E.; Koch, S.; Eckert, A.; Hartmann, H.; Scheuer, K. β-amyloid peptide decreases membrane fluidity. Brain Res. 1995, 674, 133–136. [Google Scholar] [CrossRef] [PubMed]
- Muller, W.E.; Eckert, G.P.; Scheuer, K.; Cairns, N.J.; Maras, A.; Gattaz, W.F. Effects of β-amyloid peptides on the fluidity of membranes from frontal and parietal lobes of human brain. High potencies of Aβ1-42 and Aβ1-43. Amyloid 1998, 5, 10–15. [Google Scholar] [CrossRef] [PubMed]
- Ng, J.; Kamm, R.D.; Wohland, T.; Kraut, R.S. Evidence from ITIR-FCS Diffusion Studies that the Amyloid-Beta (Aβ) Peptide Does Not Perturb Plasma Membrane Fluidity in Neuronal Cells. J. Mol. Biol. 2018, 430, 3439–3453. [Google Scholar] [CrossRef] [PubMed]
- Chafekar, S.M.; Baas, F.; Scheper, W. Oligomer-specific Aβ toxicity in cell models is mediated by selective uptake. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2008, 1782, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Nazere, K.; Takahashi, T.; Hara, N.; Muguruma, K.; Nakamori, M.; Yamazaki, Y.; Morino, H.; Maruyama, H. Amyloid Beta Is Internalized via Macropinocytosis, an HSPG- and Lipid Raft-Dependent and Rac1-Mediated Process. Front. Mol. Neurosci. 2022, 15, 804702. [Google Scholar] [CrossRef] [PubMed]
- Ikeda, K.; Yamaguchi, T.; Fukunaga, S.; Hoshino, M.; Matsuzaki, K. Mechanism of Amyloid β-Protein Aggregation Mediated by GM1 Ganglioside Clusters. Biochemistry 2011, 50, 6433–6440. [Google Scholar] [CrossRef]
- Staneva, G.; Puff, N.; Stanimirov, S.; Tochev, T.; Angelova, M.I.; Seigneuret, M. The Alzheimer’s disease amyloid-β peptide affects the size-dynamics of raft-mimicking Lo domains in GM1-containing lipid bilayers. Soft Matter 2018, 14, 9609–9618. [Google Scholar] [CrossRef] [PubMed]
- Varshney, P.; Yadav, V.; Saini, N. Lipid rafts in immune signalling: Current progress and future perspective. Immunology 2016, 149, 13–24. [Google Scholar] [CrossRef] [PubMed]
- Inamdar, K.; Floderer, C.; Favard, C.; Muriaux, D. Monitoring HIV-1 Assembly in Living Cells: Insights from Dynamic and Single Molecule Microscopy. Viruses 2019, 11, 72. [Google Scholar] [CrossRef] [PubMed]
- Lelek, M.; Di Nunzio, F.; Henriques, R.; Charneau, P.; Arhel, N.; Zimmer, C. Superresolution imaging of HIV in infected cells with FlAsH-PALM. Proc. Natl. Acad. Sci. USA 2012, 109, 8564–8569. [Google Scholar] [CrossRef]
- Pathak, P.; London, E. The Effect of Membrane Lipid Composition on the Formation of Lipid Ultrananodomains. Biophys. J. 2015, 109, 1630–1638. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Yuan, Z.; Pavel, M.A.; Jablonski, S.M.; Jablonski, J.; Hobson, R.; Valente, S.; Reddy, C.B.; Hansen, S.B. The role of high cholesterol in SARS-CoV-2 infectivity. J. Biol. Chem. 2023, 299, 104763. [Google Scholar] [CrossRef] [PubMed]
- Storti, B.; Quaranta, P.; Di Primio, C.; Clementi, N.; Mancini, N.; Criscuolo, E.; Spezia, P.G.; Carnicelli, V.; Lottini, G.; Paolini, E.; et al. A spatial multi-scale fluorescence microscopy toolbox discloses entry checkpoints of SARS-CoV-2 variants in Vero E6 cells. Comput. Struct. Biotechnol. J. 2021, 19, 6140–6156. [Google Scholar] [CrossRef] [PubMed]
- A Mir, M.; Mir, B.; Kumawat, M.; Alkhanani, M.; Jan, U. Manipulation and Exploitation Of Host Immune System by Pathogenic Mycobacterium Tuberculosis for its Advantage. Future Microbiol. 2022, 17, 1171–1198. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, Y.; Li, W.; Tang, H.; Zhou, Y.; You, D.; Chu, X.; Li, H.; Shang, J.; Qi, N.; et al. Mycobacterium tuberculosis Suppresses Inflammatory Responses in Host through Its Cholesterol Metabolites. ACS Infect. Dis. 2024, 10, 3650–3663. [Google Scholar] [CrossRef]
- Guo, Y.; Song, G.; Sun, M.; Wang, J.; Wang, Y. Prevalence and Therapies of Antibiotic-Resistance in Staphylococcus aureus. Front. Cell. Infect. Microbiol. 2020, 10, 107. [Google Scholar] [CrossRef] [PubMed]
- Müller, P.; Donnelly, M.J.; Ranson, H. Transcription profiling of a recently colonised pyrethroid resistant Anopheles gambiae strain from Ghana. BMC Genom. 2007, 8, 36. [Google Scholar] [CrossRef]
- Goldmann, O.; Lang, J.C.; Rohde, M.; May, T.; Molinari, G.; Medina, E. Alpha-hemolysin promotes internalization of Staphylococcus aureus into human lung epithelial cells via caveolin-1- and cholesterol-rich lipid rafts. Cell. Mol. Life Sci. 2024, 81, 1–16. [Google Scholar] [CrossRef]
- Roy, A.; Patra, S.K. Lipid Raft Facilitated Receptor Organization and Signaling: A Functional Rheostat in Embryonic Development, Stem Cell Biology and Cancer. Stem Cell Rev. Rep. 2022, 19, 2–25. [Google Scholar] [CrossRef]
- Bag, N.; Wagenknecht-Wiesner, A.; Lee, A.; Shi, S.M.; Holowka, D.A.; Baird, B.A. Lipid-based and protein-based interactions synergize transmembrane signaling stimulated by antigen clustering of IgE receptors. Proc. Natl. Acad. Sci. USA 2021, 118, e2026583118. [Google Scholar] [CrossRef]
- Hernaiz-Llorens, M.; Roselló-Busquets, C.; Durisic, N.; Filip, A.; Ulloa, F.; Martínez-Mármol, R.; Soriano, E. Growth cone repulsion to Netrin-1 depends on lipid raft microdomains enriched in UNC5 receptors. Cell. Mol. Life Sci. 2020, 78, 2797–2820. [Google Scholar] [CrossRef] [PubMed]
- Inestrosa, N.C.; Varela-Nallar, L. Wnt signalling in neuronal differentiation and development. Cell Tissue Res. 2014, 359, 215–223. [Google Scholar] [CrossRef]
- Ng, X.W.; Teh, C.; Korzh, V.; Wohland, T. The Secreted Signaling Protein Wnt3 Is Associated with Membrane Domains In Vivo: A SPIM-FCS Study. Biophys. J. 2016, 111, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Zha, L.-Y.; Mao, L.-M.; Lu, X.-C.; Deng, H.; Ye, J.-F.; Chu, X.-W.; Sun, S.-X.; Luo, H.-J. Anti-inflammatory effect of soyasaponins through suppressing nitric oxide production in LPS-stimulated RAW 264.7 cells by attenuation of NF-κB-mediated nitric oxide synthase expression. Bioorg. Med. Chem. Lett. 2011, 21, 2415–2418. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Ullah, H.; Zheng, Z.; Gu, X.; Su, C.; Xiao, L.; Wu, X.; Xiong, F.; Li, Q.; Zha, L. Soyasaponins reduce inflammation by downregulating MyD88 expression and suppressing the recruitments of TLR4 and MyD88 into lipid rafts. BMC Complement. Med. Ther. 2020, 20, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.-A.; Park, Y.-J.; Joh, E.-H.; Kim, D.-H. Soyasaponin Ab Ameliorates Colitis by Inhibiting the Binding of Lipopolysaccharide (LPS) to Toll-like Receptor (TLR)4 on Macrophages. J. Agric. Food Chem. 2011, 59, 13165–13172. [Google Scholar] [CrossRef]
- Gu, X.; Chen, J.; Zheng, Z.; Xiao, L.; Su, C.; Xiong, F.; Zha, L. Soyasaponin A1 Inhibits the Lipid Raft Recruitment and Dimerization of TLR4, MyD88 and TRIF in Palmitate-Stimulated Inflammatory Macrophages. Curr. Dev. Nutr. 2020, 4, nzaa068_007. [Google Scholar] [CrossRef]
- Virk, R.; Cook, K.; Cavazos, A.; Wassall, S.R.; Gowdy, K.M.; Shaikh, S.R. How membrane phospholipids containing long chain polyunsaturated fatty acids and their oxidation products orchestrate lipid raft dynamics to control inflammation. J. Nutr. 2024, 154, 2862–2870. [Google Scholar] [CrossRef]
- Tsuchiya, H.; Mizogami, M. Interaction of drugs with lipid raft membrane domains as a possible target. Drug Target Insights 2020, 14, 34–47. [Google Scholar] [CrossRef]
- Zapata-Morin, P.A.; Sierra-Valdez, F.; Ruiz-Suárez, J. The cut-off effect of n-alcohols in lipid rafts: A lipid-dependent phenomenon. J. Mol. Graph. Model. 2020, 101, 107732. [Google Scholar] [CrossRef]
- Dai, Q.; Pruett, S.B. Ethanol Suppresses LPS-induced Toll-like Receptor 4 Clustering, Reorganization of the Actin Cytoskeleton, and Associated TNF-α Production. Alcohol. Clin. Exp. Res. 2006, 30, 1436–1444. [Google Scholar] [CrossRef]
- Tobin, S.J.; Cacao, E.E.; Hong, D.W.W.; Terenius, L.; Vukojevic, V.; Jovanovic-Talisman, T. Nanoscale Effects of Ethanol and Naltrexone on Protein Organization in the Plasma Membrane Studied by Photoactivated Localization Microscopy (PALM). PLoS ONE 2014, 9, e87225. [Google Scholar] [CrossRef] [PubMed]
- Cutone, A.; Rosa, L.; Ianiro, G.; Lepanto, M.S.; Di Patti, M.C.B.; Valenti, P.; Musci, G. Lactoferrin’s Anti-Cancer Properties: Safety, Selectivity, and Wide Range of Action. Biomolecules 2020, 10, 456. [Google Scholar] [CrossRef] [PubMed]
- Santos-Pereira, C.; Guedes, J.P.; Ferreira, D.; Rodrigues, L.R.; Côrte-Real, M. Lactoferrin perturbs intracellular trafficking, disrupts cholesterol-rich lipid rafts and inhibits glycolysis of highly metastatic cancer cells harbouring plasmalemmal V-ATPase. Int. J. Biol. Macromol. 2022, 220, 1589–1604. [Google Scholar] [CrossRef] [PubMed]
- Verma, A.K.; Noumani, A.; Yadav, A.K.; Solanki, P.R. FRET Based Biosensor: Principle Applications Recent Advances and Challenges. Diagnostics 2023, 13, 1375. [Google Scholar] [CrossRef] [PubMed]
- Seong, J. Lipid rafts signaling detected by FRET-based molecular biosensors. J. Korean Soc. Appl. Biol. Chem. 2015, 58, 629–636. [Google Scholar] [CrossRef]
- Seong, J.; Huang, M.; Sim, K.M.; Kim, H.; Wang, Y. FRET-based Visualization of PDGF Receptor Activation at Membrane Microdomains. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Demoulin, J.-B.; Essaghir, A. PDGF receptor signaling networks in normal and cancer cells. Cytokine Growth Factor Rev. 2014, 25, 273–283. [Google Scholar] [CrossRef] [PubMed]
- Papadopoulos, N.; Lennartsson, J. The PDGF/PDGFR pathway as a drug target. Mol. Asp. Med. 2018, 62, 75–88. [Google Scholar] [CrossRef] [PubMed]
- Kim, T.; Lei, L.; Seong, J.; Suh, J.; Jang, Y.; Jung, S.H.; Sun, J.; Kim, D.; Wang, Y. Matrix Rigidity-Dependent Regulation of Ca2+ at Plasma Membrane Microdomains by FAK Visualized by Fluorescence Resonance Energy Transfer. Adv. Sci. 2018, 6, 1801290. [Google Scholar] [CrossRef] [PubMed]

| Name | Partitioning | Reference |
|---|---|---|
| Probes for lipid components | ||
| Cholera toxin B (CT-B) | - | [52,53] |
| BODIPY-GM1 | Lo | [54] |
| Filipin | - | [55,56,57] |
| Cholestatrienol (CTL) | Lo | [58,59] |
| Dehydroergosterol (DHE) | - | [58,59,60] |
| (NBD)-cholesterol | Ld | [61] |
| TopFluor-Cholesterol (TF-Chol) | Lo | [62] |
| BODIPY-SM | Lo | [63] |
| ATTO488-SM | Ld | [64] |
| ATTO594-SM | Ld | [64] |
| Probes Selective to Lo Phase | ||
| Terrylene | Lo | [65] |
| Naphthopyrene (NAP) | Lo | [65] |
| Solvatochromic probes | ||
| Laurdan | Lo/Ld | [66,67] |
| Prodan | Lo/Ld | [68,69] |
| C-Laurdan | Lo/Ld | [70] |
| di-4-ANEPPDHQ | Lo/Ld | [71] |
| Nile Red | Lo/Ld | [72] |
| NR12S | Lo/Ld | [73] |
| F2N12S | Lo/Ld | [74] |
| Molecular rotors | ||
| C-Laurdan-2 | Lo/Ld | [75] |
| S-Laurdan-2 | Lo/Ld | [76] |
| FCVJ | - | [77] |
| BODIPY C10 | Ld | [78] |
| BODIPY-PM | Lo/Ld | [79] |
| CCVJ | - | [80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Anselmo, S.; Bonaccorso, E.; Gangemi, C.; Sancataldo, G.; Conti Nibali, V.; D’Angelo, G. Lipid Rafts in Signalling, Diseases, and Infections: What Can Be Learned from Fluorescence Techniques? Membranes 2025, 15, 6. https://doi.org/10.3390/membranes15010006
Anselmo S, Bonaccorso E, Gangemi C, Sancataldo G, Conti Nibali V, D’Angelo G. Lipid Rafts in Signalling, Diseases, and Infections: What Can Be Learned from Fluorescence Techniques? Membranes. 2025; 15(1):6. https://doi.org/10.3390/membranes15010006
Chicago/Turabian StyleAnselmo, Sara, Elisa Bonaccorso, Chiara Gangemi, Giuseppe Sancataldo, Valeria Conti Nibali, and Giovanna D’Angelo. 2025. "Lipid Rafts in Signalling, Diseases, and Infections: What Can Be Learned from Fluorescence Techniques?" Membranes 15, no. 1: 6. https://doi.org/10.3390/membranes15010006
APA StyleAnselmo, S., Bonaccorso, E., Gangemi, C., Sancataldo, G., Conti Nibali, V., & D’Angelo, G. (2025). Lipid Rafts in Signalling, Diseases, and Infections: What Can Be Learned from Fluorescence Techniques? Membranes, 15(1), 6. https://doi.org/10.3390/membranes15010006