
Dysfunction of EAAT3 Aggravates LPS-Induced Post-Operative Cognitive Dysfunction

 , and
, and {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs
2.3. Experiment Design and Groups
2.4. Experimental Methods
2.5. Statistical Analysis
3. Results
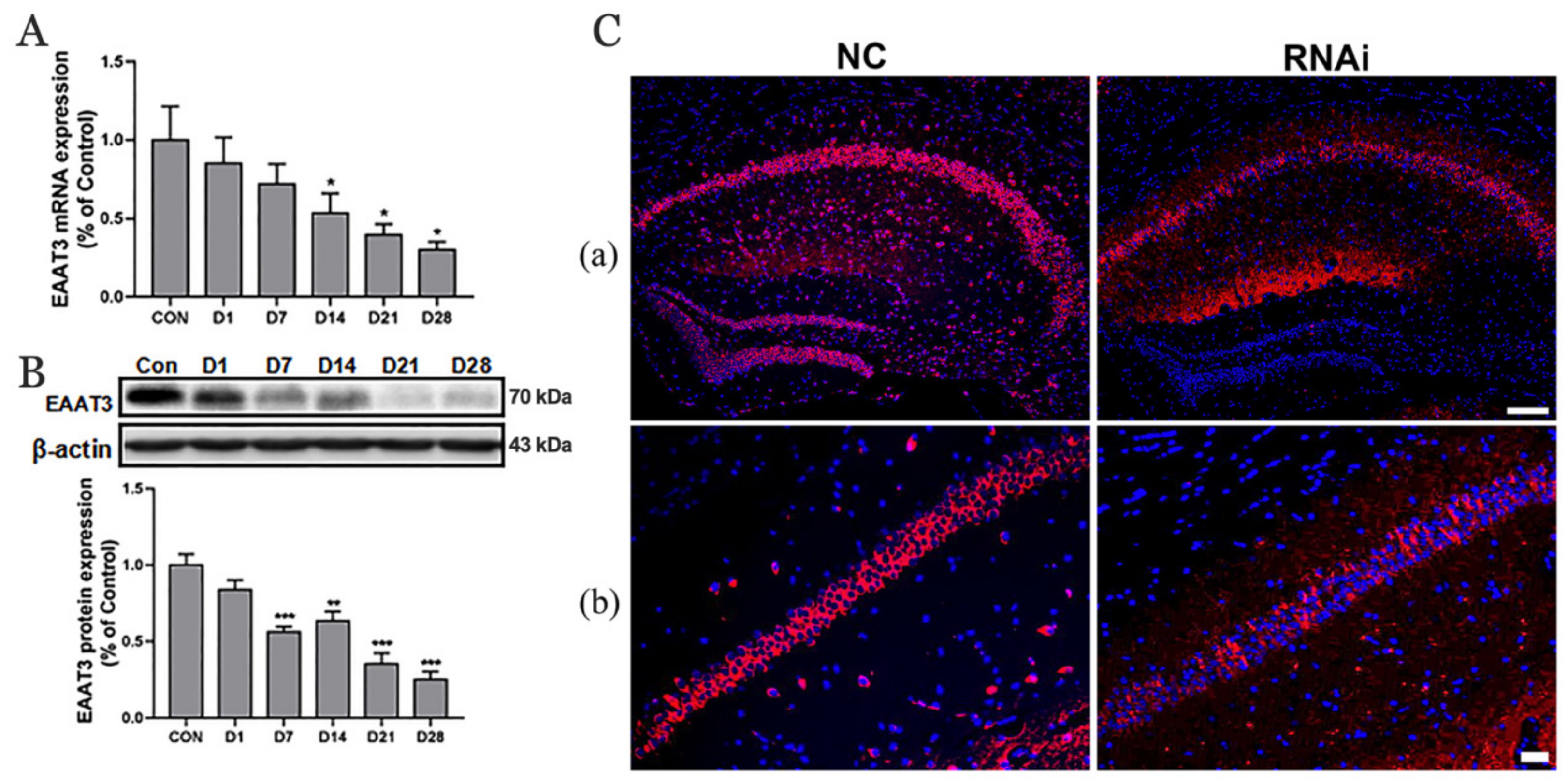
3.1. Expression of EAAT3 in the Hippocampus of Adult Mice Significantly Decreased 21 Days after rAAV-RNAi Microinjection
3.2. SLC1A1/EAAT3 Knockdown in the Hippocampus Had No Significant Effect on the Learning and Memory of Adult Mice
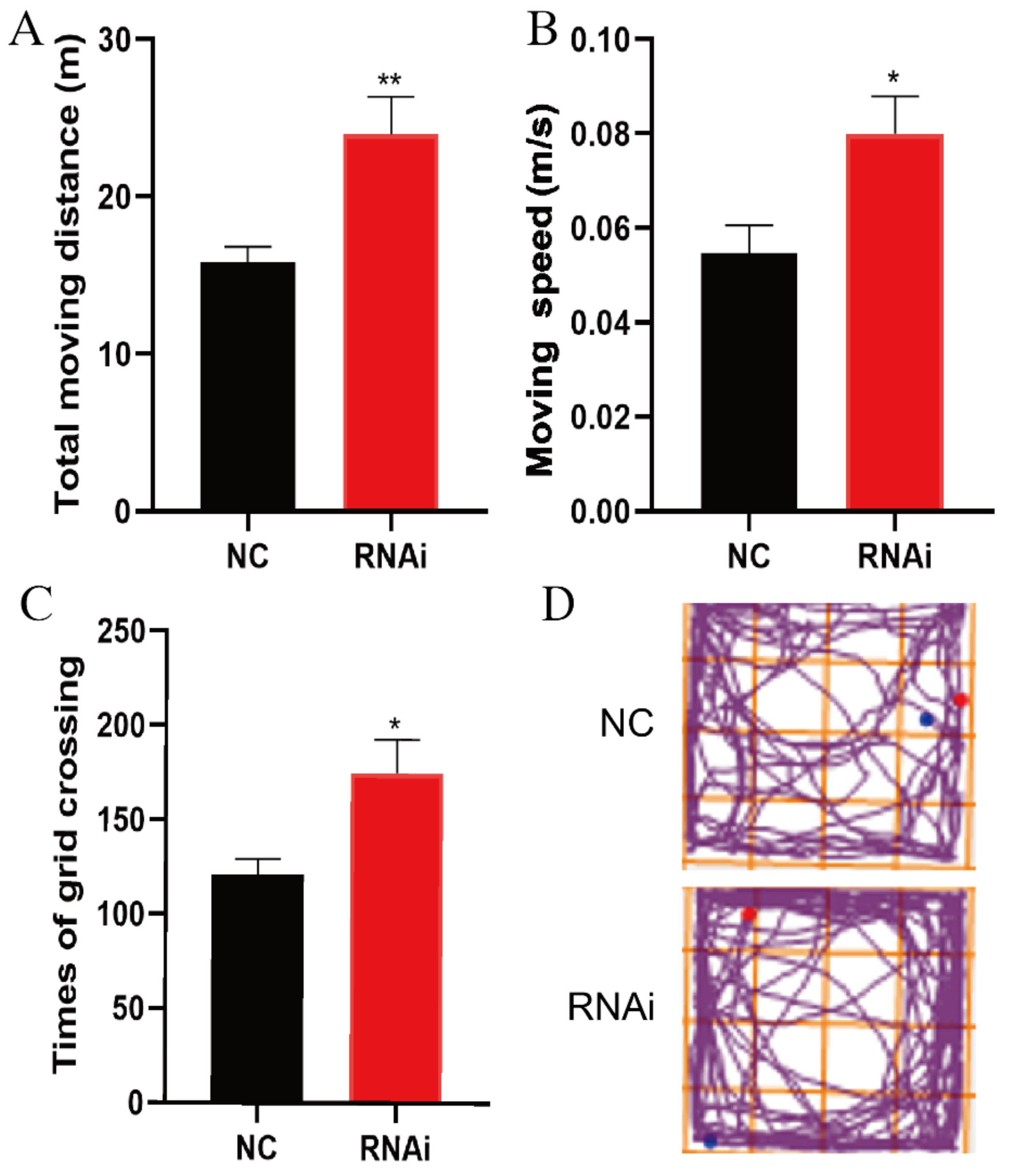
3.3. SLC1A1/EAAT3 Knockdown in the Hippocampus Significantly Increased the Motility of Adult Mice
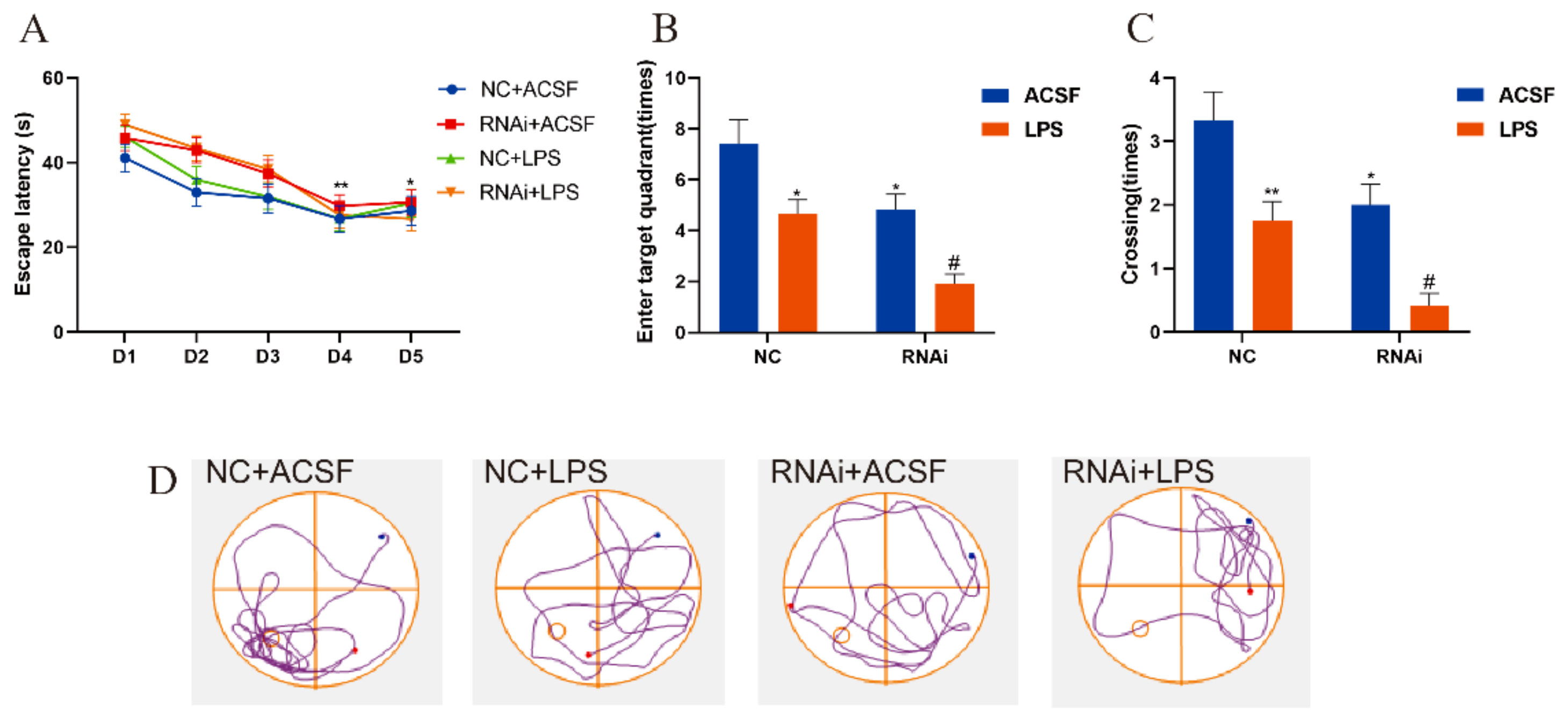
3.4. Hippocampal SLC1A1/EAAT3 Knockdown Significantly Aggravated LPS-Induced Learning and Memory Deficit in Adult Mice
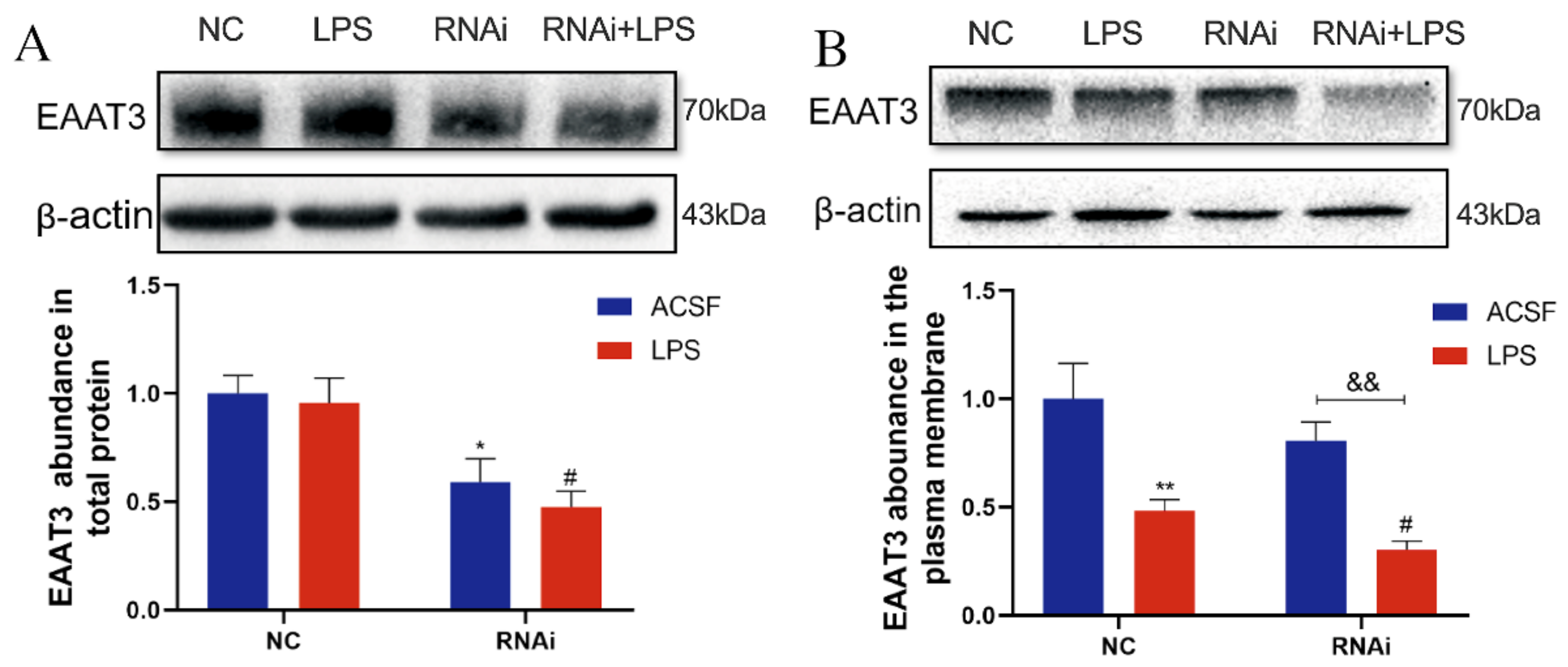
3.5. LPS Significantly Decreased Plasma Membrane Protein Level of EAAT3 in Adult Mice
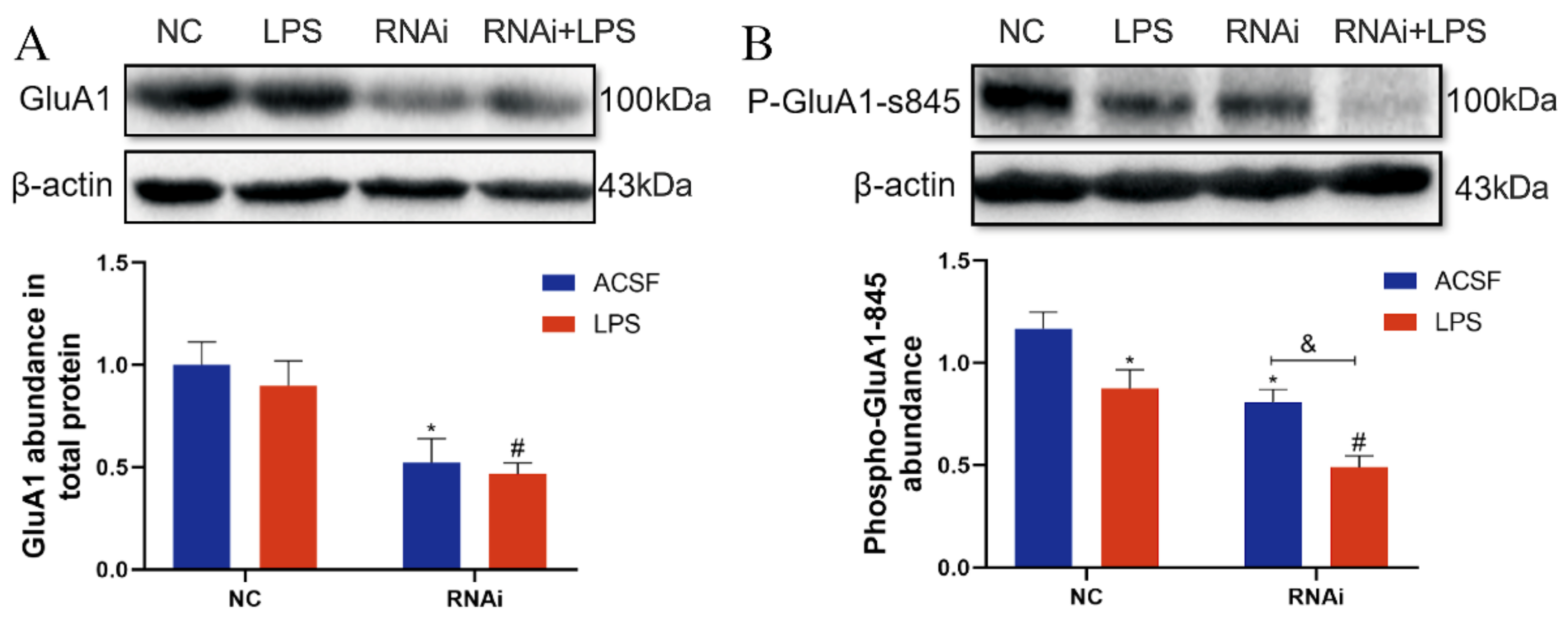
3.6. Expression and Phosphorylation of GluA1 Protein Were Inhibited by LPS in the Hippocampus of EAAT3 Knockdown Mice
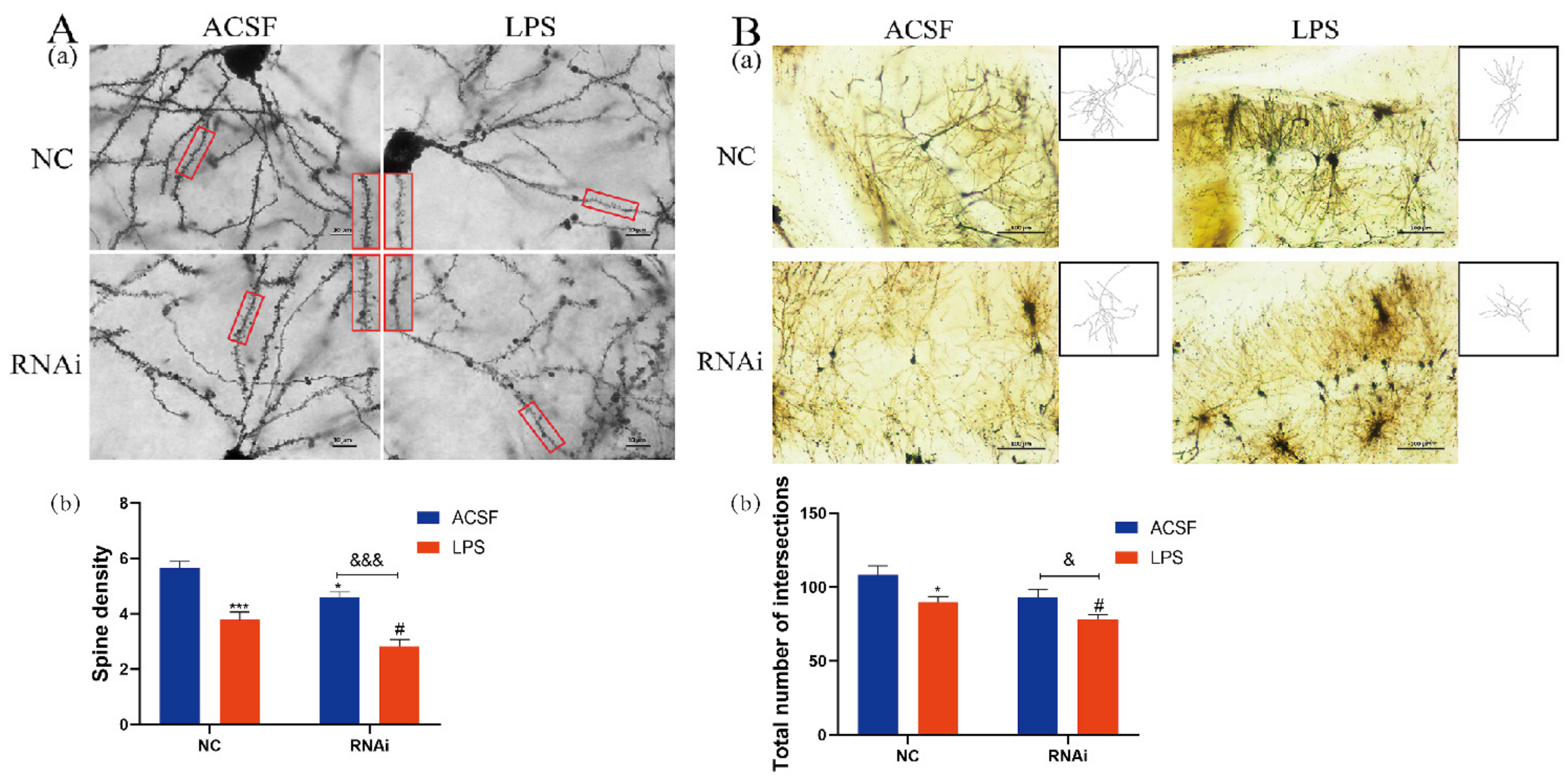
3.7. LPS Aggravated the Decreases of Dendritic Complexity in the Hippocampus of EAAT3 Knockdown Mice
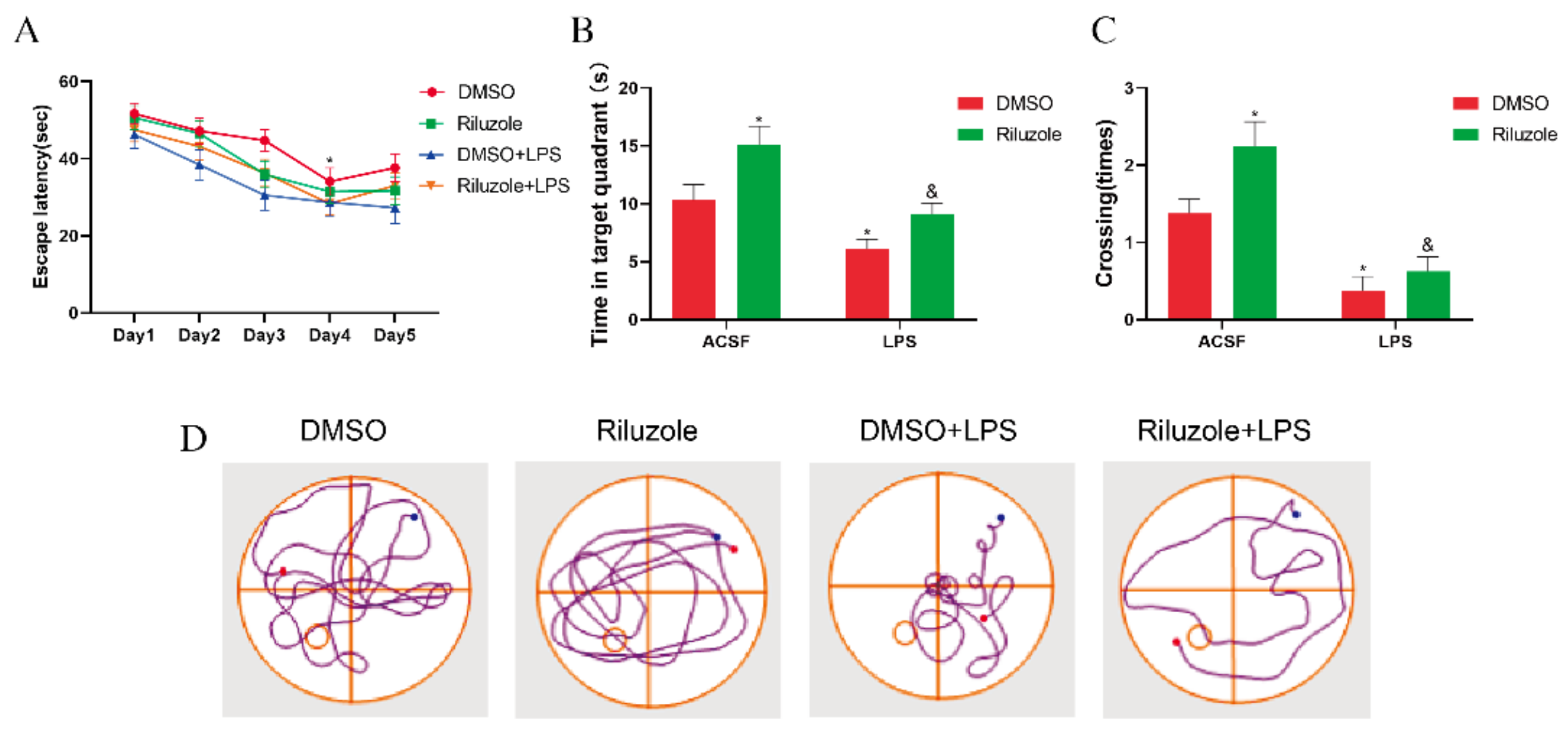
3.8. Riluzole Improved Learning and Memory Ability and Ameliorated LPS-Induced Cognitive Impairment in Old Mice
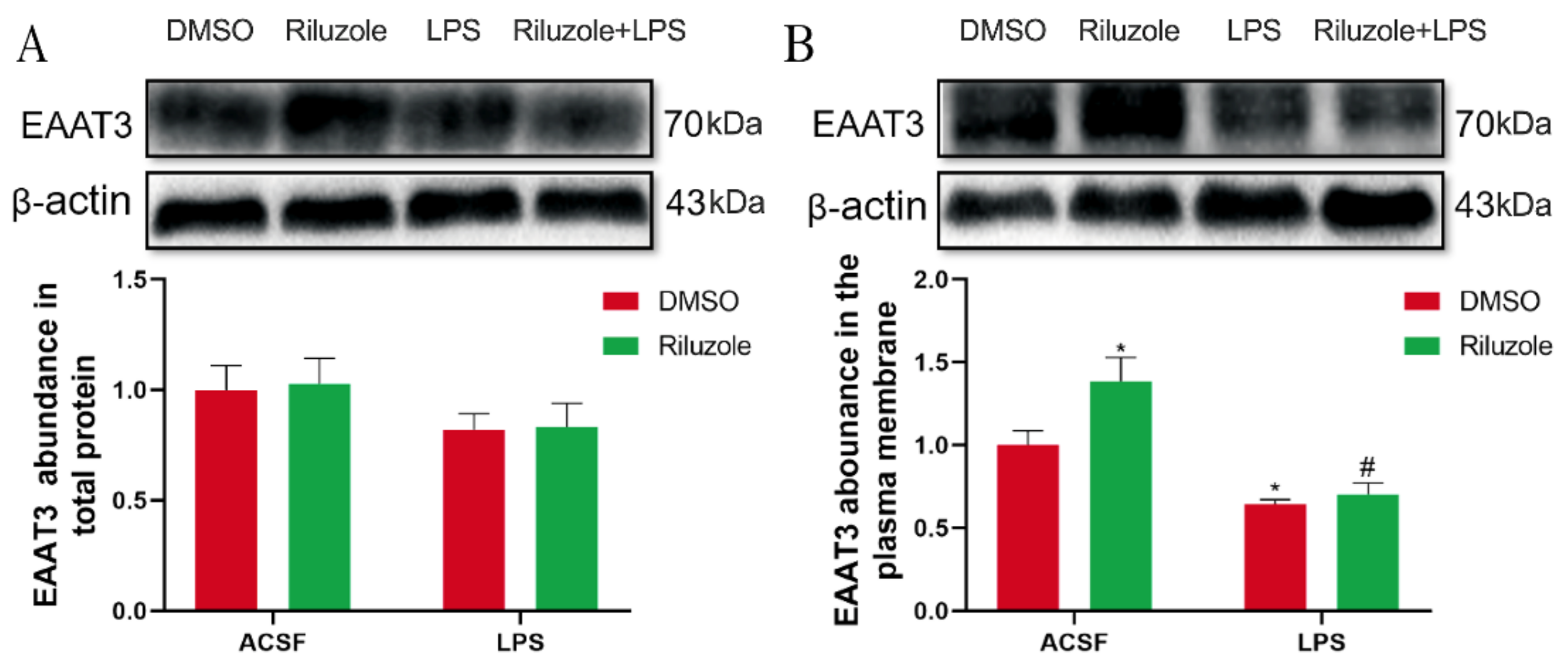
3.9. Riluzole Significantly Increased the Expression of EAAT3 Membrane Protein in the Hippocampus of Old Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
References
- Benson, R.A.; Ozdemir, B.A.; Matthews, D.; Loftus, I.M. A systematic review of postoperative cognitive decline following open and endovascular aortic aneurysm surgery. Ann. R. Coll. Surg. Engl. 2017, 99, 97–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Safavynia, S.A.; Goldstein, P.A. The Role of Neuroinflammation in Postoperative Cognitive Dysfunction: Moving from Hypothesis to Treatment. Front. Psychiatry 2018, 9, 752. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Dong, Y.; Wang, H.; Culley, D.J.; Marcantonio, E.R.; Crosby, G.; Tanzi, R.E.; Zhang, Y.; Xie, Z. Age-dependent postoperative cognitive impairment and Alzheimer-related neuropathology in mice. Sci. Rep. 2014, 4, 3766. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bjørn-Yoshimoto, W.E.; Underhill, S.M. The importance of the excitatory amino acid transporter 3 (EAAT3). Neurochem. Int. 2016, 98, 4–18. [Google Scholar] [CrossRef] [Green Version]
- Sheldon, A.L.; González, M.I.; Robinson, M.B. A carboxyl-terminal determinant of the neuronal glutamate transporter, EAAC1, is required for platelet-derived growth factor-dependent trafficking. J. Biol. Chem. 2006, 281, 4876–4886. [Google Scholar] [CrossRef] [Green Version]
- Rothstein, J.D.; Martin, L.; Levey, A.I.; Dykes-Hoberg, M.; Jin, L.; Wu, D.; Nash, N.; Kuncl, R.W. Localization of neuronal and glial glutamate transporters. Neuron 1994, 13, 713–725. [Google Scholar] [CrossRef]
- Sims, K.D.; Straff, D.J.; Robinson, M.B. Platelet-derived growth factor rapidly increases activity and cell surface expression of the EAAC1 subtype of glutamate transporter through activation of phosphatidylinositol 3-kinase. J. Biol. Chem. 2000, 275, 5228–5237. [Google Scholar] [CrossRef] [Green Version]
- Hodgson, N.; Trivedi, M.; Muratore, C.; Li, S.; Deth, R. Soluble oligomers of amyloid-β cause changes in redox state, DNA methylation, and gene transcription by inhibiting EAAT3 mediated cysteine uptake. J. Alzheimer’s Dis. 2013, 36, 197–209. [Google Scholar] [CrossRef]
- Duerson, K.; Woltjer, R.L.; Mookherjee, P.; Leverenz, J.B.; Montine, T.J.; Bird, T.D.; Pow, D.V.; Rauen, T.; Cool, D.G. Detergent-insoluble EAAC1/EAAT3 aberrantly accumulates in hippocampal neurons of Alzheimer’s disease patients. Brain Pathol. 2009, 19, 267–278. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.C.; Jackson, J.G.; Krizman, E.N.; Rothstein, J.D.; Porter, B.E.; Robinson, M.B. Genetic deletion of the neuronal glutamate transporter, EAAC1, results in decreased neuronal death after pilocarpine-induced status epilepticus. Neurochem. Int. 2014, 73, 152–158. [Google Scholar] [CrossRef] [Green Version]
- Sala, G.; Beretta, S.; Ceresa, C.; Mattavelli, L.; Zoia, C.; Tremolizzo, L.; Ferri, A.; Carrì, M.T.; Ferrarese, C. Impairment of glutamate transport and increased vulnerability to oxidative stress in neuroblastoma SH-SY5Y cells expressing a Cu, Zn superoxide dismutase typical of familial amyotrophic lateral sclerosis. Neurochem. Int. 2005, 46, 227–234. [Google Scholar] [CrossRef]
- McCullumsmith, R.E.; Meador-Woodruff, J.H. Striatal excitatory amino acid transporter transcript expression in schizophrenia, bipolar disorder, and major depressive disorder. Neuropsychopharmacology 2002, 26, 368–375. [Google Scholar] [CrossRef] [Green Version]
- Gil, Y.S.; Kim, J.H.; Kim, C.H.; Han, J.I.; Zuo, Z.; Baik, H.J. Gabapentin inhibits the activity of the rat excitatory glutamate transporter 3 expressed in Xenopus oocytes. Eur. J. Pharmacol. 2015, 762, 112–117. [Google Scholar] [CrossRef]
- Chohan, M.O.; Kopelman, J.M.; Yueh, H.; Fazlali, Z.; Greene, N.; Harris, A.Z.; Balsam, P.D.; Leonardo, E.D.; Kramer, E.R.; Veenstra-VanderWeele, J.; et al. Developmental impact of glutamate transporter overexpression on dopaminergic neuron activity and stereotypic behavior. Mol Psychiatry. 2022. [Google Scholar] [CrossRef]
- Escobar, A.P.; Martínez-Pinto, J.; Silva-Olivares, F.; Sotomayor-Zárate, R.; Moya, P.R. Altered Grooming Syntax and Amphetamine-Induced Dopamine Release in EAAT3 Overexpressing Mice. Front. Cell Neurosci. 2021, 15, 661478. [Google Scholar] [CrossRef]
- Shi, C.; Yi, D.; Li, Z.; Zhou, Y.; Cao, Y.; Sun, Y.; Chui, D.; Guo, X. Anti-RAGE antibody attenuates isoflurane-induced cognitive dysfunction in aged rats. Behav. Brain Res. 2017, 322, 167–176. [Google Scholar] [CrossRef]
- Banoub, J.H.; El Aneed, A.; Cohen, A.M.; Joly, N. Structural investigation of bacterial lipopolysaccharides by mass spectrometry and tandem mass spectrometry. Mass Spectrom. Rev. 2010, 29, 606–650. [Google Scholar] [CrossRef]
- Bossù, P.; Cutuli, D.; Palladino, I.; Caporali, P.; Angelucci, F.; Laricchiuta, D.; Gelfo, F.; De Bartolo, P.; Caltagirone, C.; Petrosini, L. A single intraperitoneal injection of endotoxin in rats induces long-lasting modifications in behavior and brain protein levels of TNF-α and IL-18. J. Neuroinflamm. 2012, 9, 101. [Google Scholar] [CrossRef]
- Zhang, X.Y.; Cao, J.B.; Zhang, L.M.; Li, Y.F.; Mi, W.D. Deferoxamine attenuates lipopolysaccharide-induced neuroinflammation and memory impairment in mice. J. Neuroinflamm. 2015, 12, 20. [Google Scholar] [CrossRef] [Green Version]
- Zhao, W.X.; Zhang, J.H.; Cao, J.B.; Wang, W.; Wang, D.X.; Zhang, X.Y.; Yu, J.; Zhang, Y.-Y.; Zhang, Y.-Z.; Mi, W.-D. Acetaminophen attenuates lipopolysaccharide-induced cognitive impairment through antioxidant activity. J. Neuroinflamm. 2017, 14, 17. [Google Scholar] [CrossRef] [Green Version]
- Carbone, M.; Duty, S.; Rattray, M. Riluzole neuroprotection in a Parkinson’s disease model involves suppression of reactive astrocytosis but not GLT-1 regulation. BMC Neurosci. 2012, 13, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorhees, C.V.; Williams, M.T. Morris water maze: Procedures for assessing spatial and related forms of learning and memory. Nat. Protoc. 2006, 1, 848–858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, F. Golgi-Cox Staining of Neuronal Dendrites and Dendritic Spines with FD Rapid GolgiStain™ Kit. Curr. Protoc. Neurosci. 2019, 88, e69. [Google Scholar] [CrossRef] [PubMed]
- Donzanti, B.A.; Hite, J.F.; Yamamoto, B.K. Extracellular glutamate levels increase with age in the lateral striatum: Potential involvement of presynaptic D-2 receptors. Synapse 1993, 13, 376–382. [Google Scholar] [CrossRef]
- Massieu, L.; Tapia, R. Glutamate uptake impairment and neuronal damage in young and aged rats in vivo. J. Neurochem. 1997, 69, 1151–1160. [Google Scholar] [CrossRef]
- Brimson, J.M.; Brimson, S.J.; Brimson, C.A.; Rakkhitawatthana, V.; Tencomnao, T. Rhinacanthus nasutus extracts prevent glutamate and amyloid-β neurotoxicity in HT-22 mouse hippocampal cells: Possible active compounds include lupeol, stigmasterol and β-sitosterol. Int. J. Mol. Sci. 2012, 13, 5074–5097. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Liang, L.; Liu, G.; Liu, Y.; Zheng, H.; Dai, L. The effects of short time hyperoxia on glutamate concentration and glutamate transporters expressions in brain of neonatal rats. Neurosci. Lett. 2021, 758, 136013. [Google Scholar] [CrossRef]
- Wang, Z.; Park, S.H.; Zhao, H.; Peng, S.; Zuo, Z. A critical role of glutamate transporter type 3 in the learning and memory of mice. Neurobiol. Learn. Mem. 2014, 114, 70–80. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Wang, Z.; Mi, W.; Zuo, Z. Isoflurane unveils a critical role of glutamate transporter type 3 in regulating hippocampal GluR1 trafficking and context-related learning and memory in mice. Neuroscience 2014, 272, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Holmseth, S.; Dehnes, Y.; Huang, Y.H.; Follin-Arbelet, V.V.; Grutle, N.J.; Mylonakou, M.N.; Plachez, C.; Zhou, Y.; Furness, D.N.; Bergles, D.E.; et al. The density of EAAC1 (EAAT3) glutamate transporters expressed by neurons in the mammalian CNS. J. Neurosci. 2012, 32, 6000–6013. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.; Park, S.H.; Zuo, Z. Effects of isoflurane on learning and memory functions of wild-type and glutamate transporter type 3 knockout mice. J. Pharm. Pharmacol. 2012, 64, 302–307. [Google Scholar] [CrossRef]
- Lee, S.N.; Li, L.; Zuo, Z. Glutamate transporter type 3 knockout mice have a decreased isoflurane requirement to induce loss of righting reflex. Neuroscience 2010, 171, 788–793. [Google Scholar] [CrossRef] [Green Version]
- González, L.F.; Henríquez-Belmar, F.; Delgado-Acevedo, C.; Cisternas-Olmedo, M.; Arriagada, G.; Sotomayor-Zárate, R.; Murphy, D.L.; Moya, P.R. Neurochemical and behavioral characterization of neuronal glutamate transporter EAAT3 heterozygous mice. Biol. Res. 2017, 50, 29. [Google Scholar] [CrossRef] [Green Version]
- Van Zandt, M.; Weiss, E.; Almyasheva, A.; Lipior, S.; Maisel, S.; Naegele, J.R. Adeno-associated viral overexpression of neuroligin 2 in the mouse hippocampus enhances GABAergic synapses and impairs hippocampal-dependent behaviors. Behav. Brain Res. 2019, 362, 7–20. [Google Scholar] [CrossRef]
- Jang, M.; Lee, S.E.; Cho, I.H. Adeno-Associated Viral Vector Serotype DJ-Mediated Overexpression of N171-82Q-Mutant Huntingtin in the Striatum of Juvenile Mice Is a New Model for Huntington’s Disease. Front. Cell Neurosci. 2018, 12, 157. [Google Scholar] [CrossRef]
- Woldbye, D.P.; Angehagen, M.; Gøtzsche, C.R.; Elbrønd-Bek, H.; Sørensen, A.T.; Christiansen, S.H.; Olesen, M.V.; Nikitidou, L.; Hansen, T.v.O.; Kanter-Schlifke, I.; et al. Adeno-associated viral vector-induced overexpression of neuropeptide Y Y2 receptors in the hippocampus suppresses seizures. Brain 2010, 133, 2778–2788. [Google Scholar] [CrossRef]
- Minami, T.; Matsumura, S.; Nishizawa, M.; Sasaguri, Y.; Hamanaka, N.; Ito, S. Acute and late effects on induction of allodynia by acromelic acid, a mushroom poison related structurally to kainic acid. Br. J. Pharmacol. 2004, 142, 679–688. [Google Scholar] [CrossRef] [Green Version]
- Lewerenz, J.; Maher, P. Chronic Glutamate Toxicity in Neurodegenerative Diseases-What is the Evidence? Front. Neurosci. 2015, 9, 469. [Google Scholar] [CrossRef]
- Zhao, W.; Xu, Z.; Cao, J.; Fu, Q.; Wu, Y.; Zhang, X.; Long, Y.; Zhang, X.; Yang, Y.; Li, Y.; et al. Elamipretide (SS-31) improves mitochondrial dysfunction, synaptic and memory impairment induced by lipopolysaccharide in mice. J. Neuroinflamm. 2019, 16, 230. [Google Scholar] [CrossRef]
- Nieoullon, A.; Canolle, B.; Masmejean, F.; Guillet, B.; Pisano, P.; Lortet, S. The neuronal excitatory amino acid transporter EAAC1/EAAT3, does it represent a major actor at the brain excitatory synapse? J. Neurochem. 2006, 98, 1007–1018. [Google Scholar] [CrossRef]
- Underhill, S.M.; Ingram, S.L.; Ahmari, S.E.; Veenstra-VanderWeele, J.; Amara, S.G. Neuronal excitatory amino acid transporter EAAT3, Emerging functions in health and disease. Neurochem. Int. 2019, 123, 69–76. [Google Scholar] [CrossRef]
- Potenza, N.; Mosca, N.; Mondola, P.; Damiano, S.; Russo, A.; De Felice, B. Human miR-26a-5p regulates the glutamate transporter SLC1A1 (EAAT3) expression. Relev. Mult. Sclerosis. Biochim. Biophys. Acta Mol. Basis Dis. 2018, 1864, 317–323. [Google Scholar] [CrossRef]
- Rumpel, S.; LeDoux, J.; Zador, A.; Malinow, R. Postsynaptic receptor trafficking underlying a form of associative learning. Science 2005, 308, 83–88. [Google Scholar] [CrossRef] [Green Version]
- Man, H.Y.; Sekine-Aizawa, Y.; Huganir, R.L. Regulation of {alpha}-amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid receptor trafficking through PKA phosphorylation of the Glu receptor 1 subunit. Proc. Natl. Acad. Sci. USA 2007, 104, 3579–3584. [Google Scholar] [CrossRef] [Green Version]
- Whitlock, J.R.; Heynen, A.J.; Shuler, M.G.; Bear, M.F. Learning induces long-term potentiation in the hippocampus. Science 2006, 313, 1093–1097. [Google Scholar] [CrossRef] [Green Version]
- Esteban, J.A.; Shi, S.H.; Wilson, C.; Nuriya, M.; Huganir, R.L.; Malinow, R. PKA phosphorylation of AMPA receptor subunits controls synaptic trafficking underlying plasticity. Nat. Neurosci. 2003, 6, 136–143. [Google Scholar] [CrossRef]
- Cao, J.; Tan, H.; Mi, W.; Zuo, Z. Glutamate transporter type 3 regulates mouse hippocampal GluR1 trafficking. Biochim. Biophys. Acta 2014, 1840, 1640–1645. [Google Scholar] [CrossRef] [Green Version]
- Caffino, L.; Messa, G.; Fumagalli, F. A single cocaine administration alters dendritic spine morphology and impairs glutamate receptor synaptic retention in the medial prefrontal cortex of adolescent rats. Neuropharmacology 2018, 140, 209–216. [Google Scholar] [CrossRef] [Green Version]
- Borczyk, M.; Śliwińska, M.A.; Caly, A.; Bernas, T.; Radwanska, K. Neuronal plasticity affects correlation between the size of dendritic spine and its postsynaptic density. Sci. Rep. 2019, 9, 1693. [Google Scholar] [CrossRef]
- Zhang, J.C.; Wu, J.; Fujita, Y.; Yao, W.; Ren, Q.; Yang, C.; Li, S.; Shirayama, Y.; Hashimoto, K. Antidepressant effects of TrkB ligands on depression-like behavior and dendritic changes in mice after inflammation. Int J Neuropsychopharmacol. 2014, 18, pyu077. [Google Scholar] [CrossRef]
- Maysinger, D.; Gröger, D.; Lake, A.; Licha, K.; Weinhart, M.; Chang, P.K.; Mulvey, R.; Haag, R.; Mckinney, R.A. Dendritic Polyglycerol Sulfate Inhibits Microglial Activation and Reduces Hippocampal CA1 Dendritic Spine Morphology Deficits. Biomacromolecules 2015, 16, 3073–3082. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, H.; Okada, Y. Roles of volume-sensitive chloride channel in excitotoxic neuronal injury. J. Neurosci. 2007, 27, 1445–1455. [Google Scholar] [CrossRef] [PubMed]
- Swiatkowski, P.; Nikolaeva, I.; Kumar, G.; Zucco, A.; Akum, B.F.; Patel, M.V.; D’Arcangelo, G.; Firestein, B.L. Role of Akt-independent mTORC1 and GSK3β signaling in sublethal NMDA-induced injury and the recovery of neuronal electrophysiology and survival. Sci. Rep. 2017, 7, 1539. [Google Scholar] [CrossRef] [PubMed] [Green Version]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, X.-Y.; Liu, W.-G.; Hou, A.-S.; Song, Y.-X.; Ma, Y.-L.; Wu, X.-D.; Cao, J.-B.; Mi, W.-D. Dysfunction of EAAT3 Aggravates LPS-Induced Post-Operative Cognitive Dysfunction. Membranes 2022, 12, 317. https://doi.org/10.3390/membranes12030317
Wang X-Y, Liu W-G, Hou A-S, Song Y-X, Ma Y-L, Wu X-D, Cao J-B, Mi W-D. Dysfunction of EAAT3 Aggravates LPS-Induced Post-Operative Cognitive Dysfunction. Membranes. 2022; 12(3):317. https://doi.org/10.3390/membranes12030317
Chicago/Turabian StyleWang, Xiao-Yan, Wen-Gang Liu, Ai-Sheng Hou, Yu-Xiang Song, Yu-Long Ma, Xiao-Dong Wu, Jiang-Bei Cao, and Wei-Dong Mi. 2022. "Dysfunction of EAAT3 Aggravates LPS-Induced Post-Operative Cognitive Dysfunction" Membranes 12, no. 3: 317. https://doi.org/10.3390/membranes12030317
APA StyleWang, X.-Y., Liu, W.-G., Hou, A.-S., Song, Y.-X., Ma, Y.-L., Wu, X.-D., Cao, J.-B., & Mi, W.-D. (2022). Dysfunction of EAAT3 Aggravates LPS-Induced Post-Operative Cognitive Dysfunction. Membranes, 12(3), 317. https://doi.org/10.3390/membranes12030317