
P80 Natural Essence Exerts Efficient Anti-HIV-1- as Well as Adjuvant Effects in DCs

 , , ,
, , ,  and
and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Longan Extract
2.2. Ethics Statement
2.3. Generation of Human Monocyte-Derived DCs
2.4. Multicolor FACS Analyses
2.5. DC Infection
2.6. Immunoblot Analyses of Phosphorylated Proteins
2.7. p24 ELISA
2.8. Virolysis
2.9. Statistical Analysis
3. Results
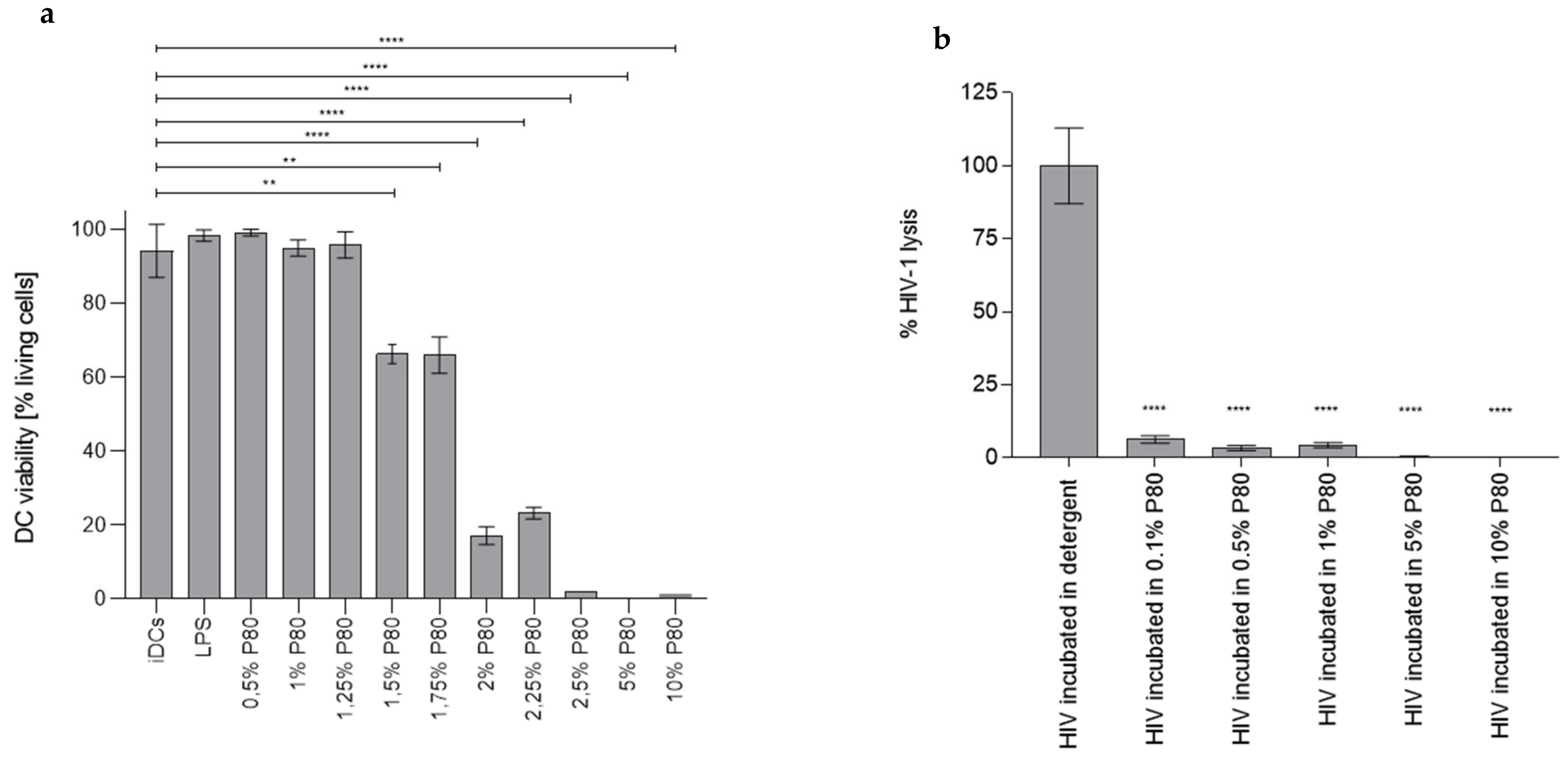
3.1. P80 Natural Essence Concentrations above 1% Result in Decreased DC Viability
3.2. P80 Natural Essence Does Not Induce Virolysis
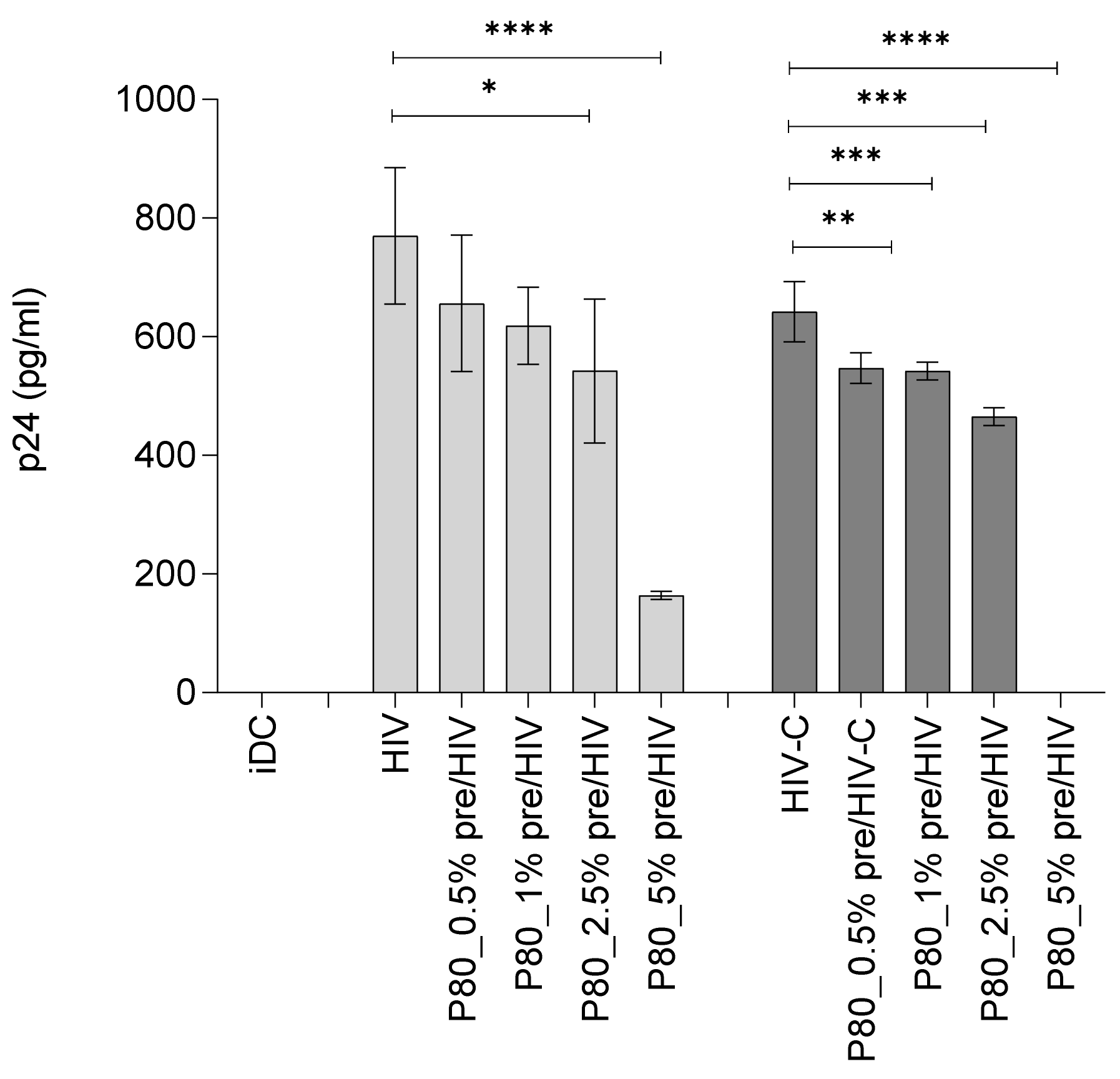
3.3. Binding of HIV Is Not as Affected by P80 Natural Essence as HIV-C attachment
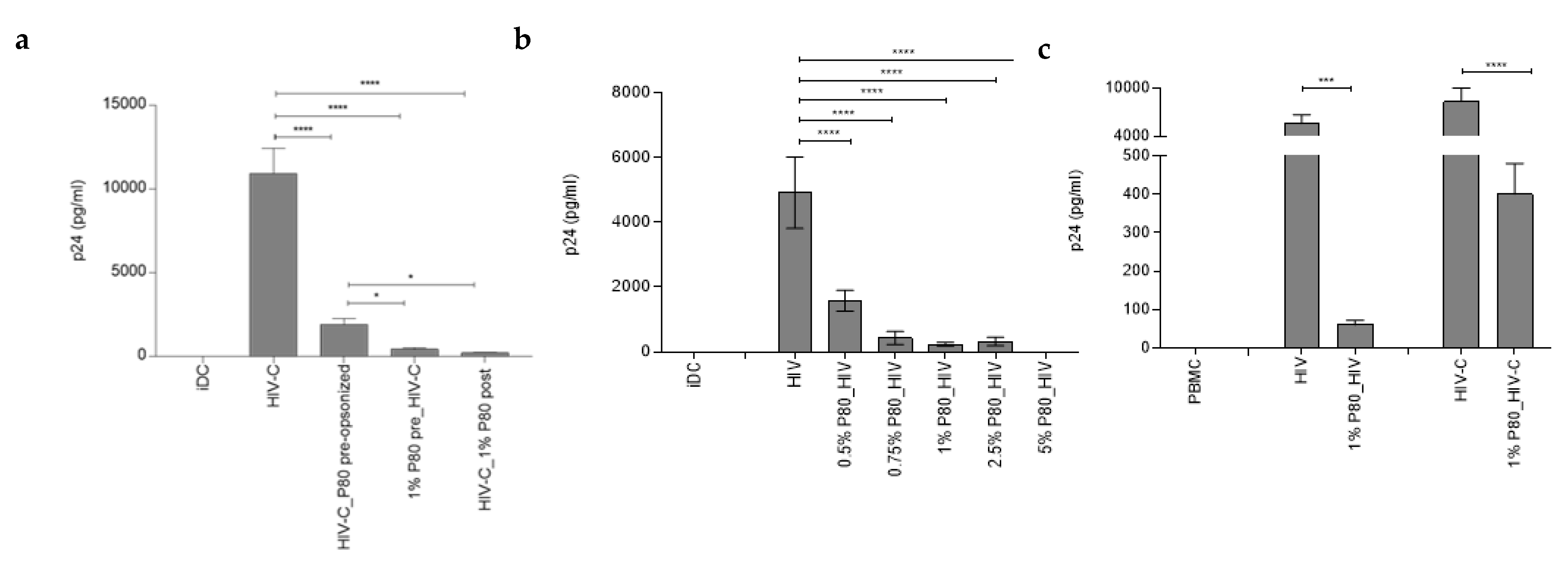
3.4. Productive Infection of DCs by Both, HIV and HIV-C, Is Abrogated by Pre-Incubation with 1% P80 Natural Essence
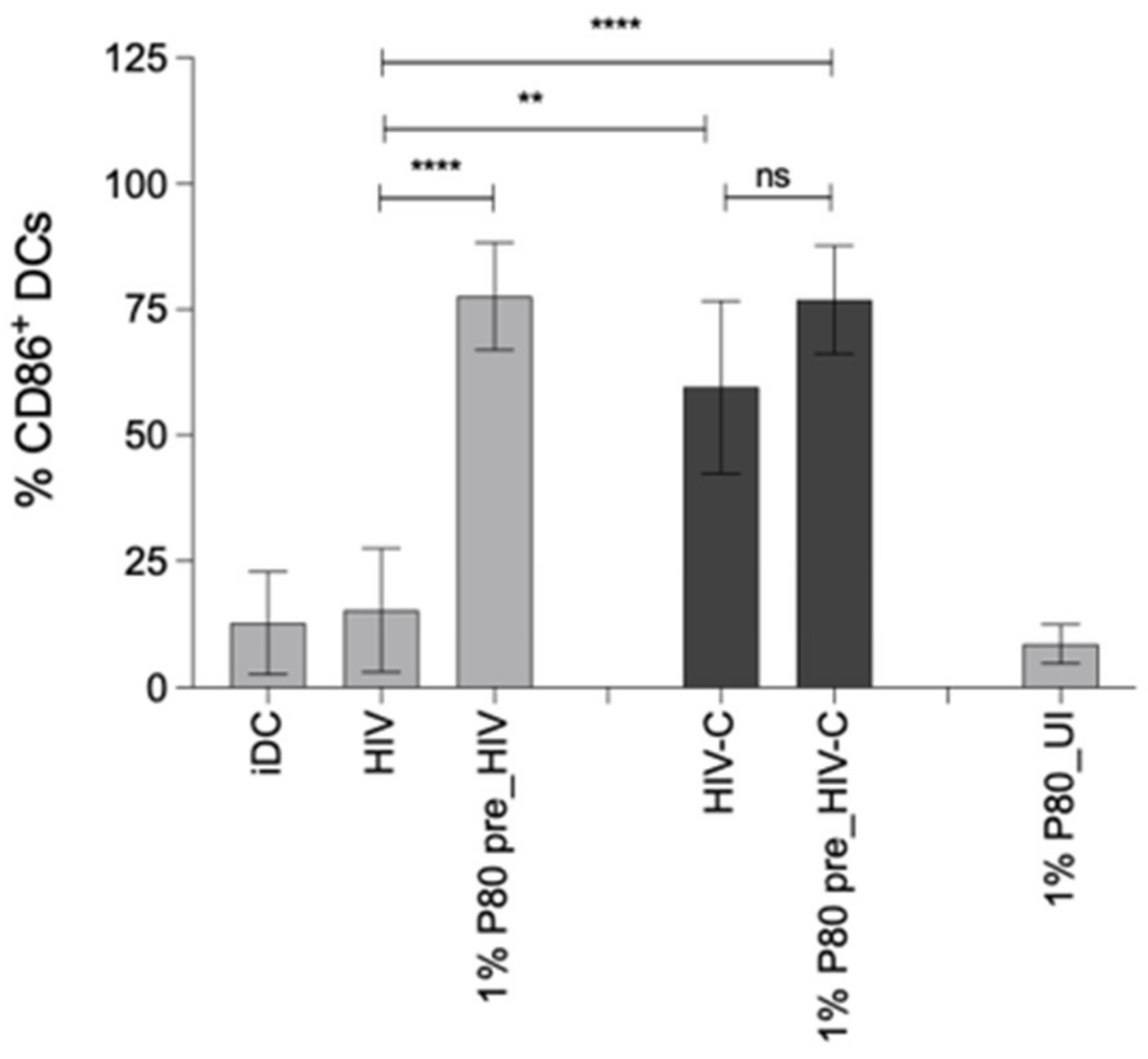
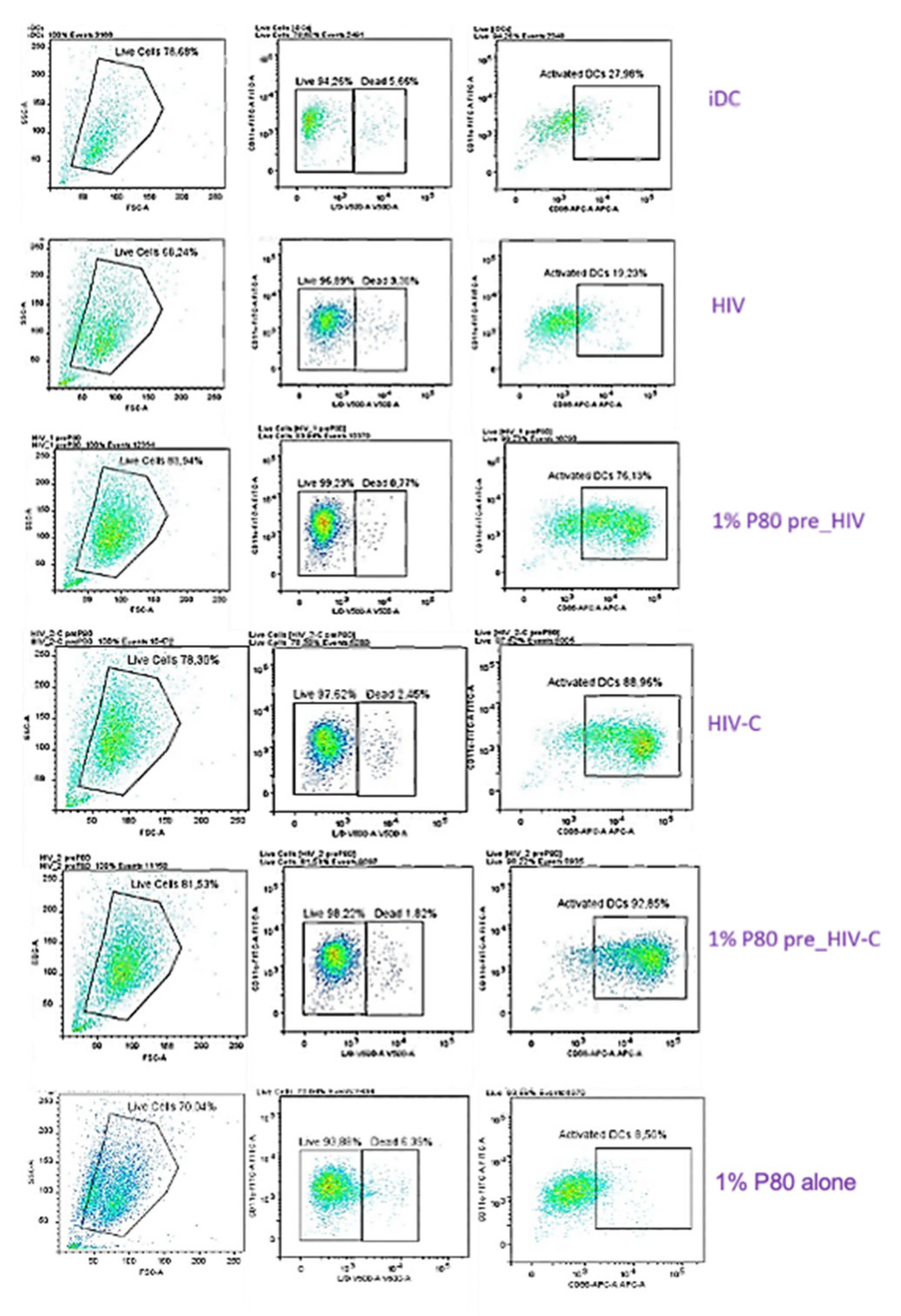
3.5. P80 Natural Essence Augments Co-Stimulatory Capacity and Maturation in HIV- and HIV-C-Exposed DCs
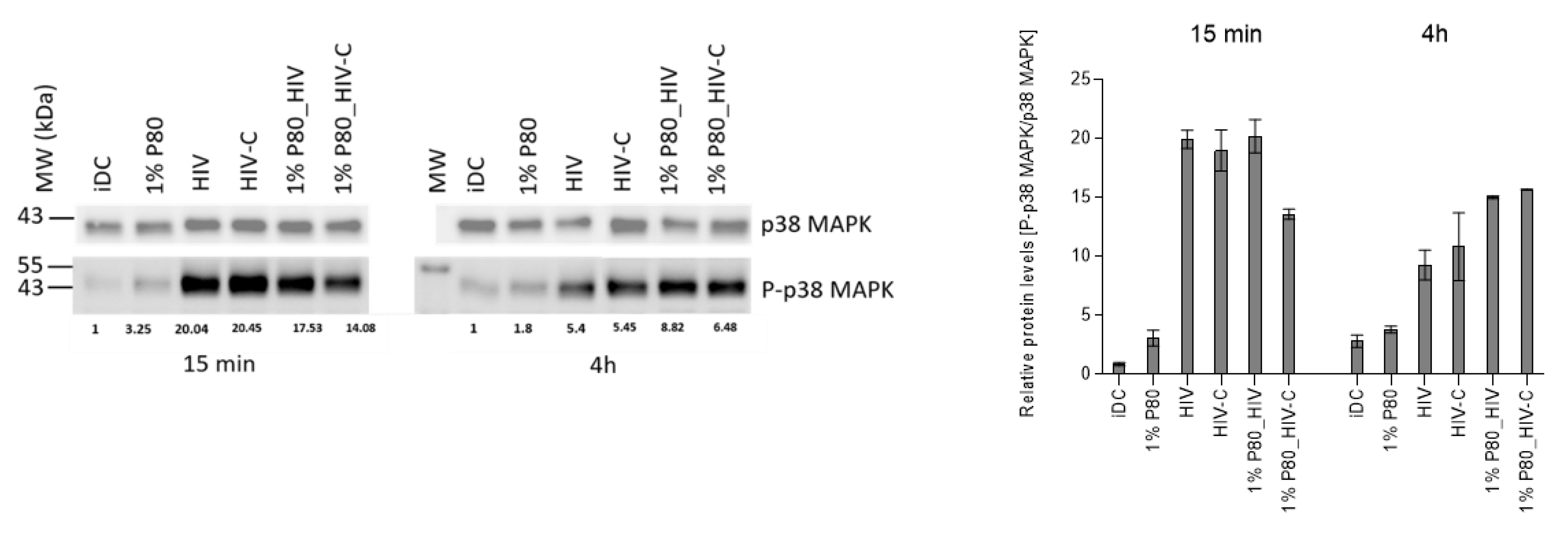
3.6. P80 Natural Essence Modifies MAPK Signaling in HIV- and HIV-C-DCs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Apriyanto, D.R.; Aoki, C.; Hartati, S.; Hanafi, M.; Kardono, L.B.S.; Arsianti, A.; Louisa, M.; Sudiro, T.M.; Dewi, B.E.; Sudarmono, P.; et al. Anti-Hepatitis C Virus Activity of a Crude Extract from Longan (Dimocarpus longan Lour.) Leaves. Jpn. J. Infect. Dis. 2016, 69, 213–220. [Google Scholar] [CrossRef]
- Yi, Y.; Liao, S.T.; Zhang, M.W.; Shi, J.; Zhang, R.F.; Deng, Y.Y.; Wei, Z.C. Physicochemical characteristics and immunomodulatory activities of three polysaccharide-protein complexes of longan pulp. Molecules 2011, 16, 6148–6164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.; Sun, J.; He, X.; Tang, Y.; Li, J.; Ling, D.; Li, C.; Li, L.; Zheng, F.; Sheng, J.; et al. Fermentation process optimization and chemical constituent analysis on longan (Dimocarpus longan Lour.) wine. Food Chem. 2018, 256, 268–279. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Gao, X.; Li, W.; Tan, S.; Zheng, Q. Phenolic content, antioxidant capacity, and alpha-amylase and alpha-glucosidase inhibitory activities of Dimocarpus longan Lour. Food Sci. Biotechnol. 2020, 29, 683–692. [Google Scholar] [CrossRef] [PubMed]
- Bai, Y.; Huang, F.; Zhang, R.; Dong, L.; Jia, X.; Liu, L.; Yi, Y.; Zhang, M. Longan pulp polysaccharides relieve intestinal injury in vivo and in vitro by promoting tight junction expression. Carbohydr. Polym. 2020, 229, 115475. [Google Scholar] [CrossRef]
- Rong, Y.; Yang, R.; Yang, Y.; Wen, Y.; Liu, S.; Li, C.; Hu, Z.; Cheng, X.; Li, W. Structural characterization of an active polysaccharide of longan and evaluation of immunological activity. Carbohydr. Polym. 2019, 213, 247–256. [Google Scholar] [CrossRef]
- Tam, J.C.H.; Bidgood, S.R.; McEwan, W.A.; James, L.C. Intracellular sensing of complement C3 activates cell autonomous immunity. Science 2014, 345, 1256070. [Google Scholar] [CrossRef] [Green Version]
- Posch, W.; Steger, M.; Knackmuss, U.; Blatzer, M.; Baldauf, H.M.; Doppler, W.; White, T.E.; Hörtnagl, P.; Diaz-Griffero, F.; Lass-Flörl, C.; et al. Complement-Opsonized HIV-1 Overcomes Restriction in Dendritic Cells. PLoS Pathog. 2015, 11, e1005005. [Google Scholar] [CrossRef]
- Wilflingseder, D.; Schroll, A.; Hackl, H.; Gallasch, R.; Frampton, D.; Lass-Flörl, C.; Pancino, G.; Saez-Cirion, A.; Lambotte, O.; Weiss, L.; et al. Immediate T-Helper 17 Polarization Upon Triggering CD11b/c on HIV-Exposed Dendritic Cells. J. Infect. Dis. 2015, 212, 44–56. [Google Scholar] [CrossRef]
- Liszewski, M.K.; Kemper, C. Complement in Motion: The Evolution of CD46 from a Complement Regulator to an Orchestrator of Normal Cell Physiology. J. Immunol. 2019, 203, 3–5. [Google Scholar] [CrossRef] [Green Version]
- Bánki, Z.; Posch, W.; Ejaz, A.; Oberhauser, V.; Willey, S.; Gassner, C.; Stoiber, H.; Dittmer, U.; Dierich, M.P.; Hasenkrug, K.J.; et al. Complement as an endogenous adjuvant for dendritic cell-mediated induction of retrovirus-specific CTLs. PLoS Pathog. 2010, 6, e1000891. [Google Scholar] [CrossRef]
- Fackler, O.T.; Alcover, A.; Schwartz, O. Modulation of the immunological synapse: A key to HIV-1 pathogenesis? Nat. Rev. Immunol. 2007, 7, 310–317. [Google Scholar] [CrossRef] [PubMed]
- Harman, A.N.; Wilkinson, J.; Bye, C.R.; Bosnjak, L.; Stern, J.L.; Nicholle, M.; Lai, J.; Cunningham, A.L. HIV induces maturation of monocyte-derived dendritic cells and Langerhans cells. J. Immunol. 2006, 177, 7103–7113. [Google Scholar] [CrossRef] [Green Version]
- Posch, W.; Cardinaud, S.; Hamimi, C.; Fletcher, A.; Mühlbacher, A.; Loacker, K.; Eichberger, P.; Dierich, M.P.; Pancino, G.; Lass-Flörl, C.; et al. Antibodies attenuate the capacity of dendritic cells to stimulate HIV-specific cytotoxic T lymphocytes. J. Allergy Clin. Immunol. 2012, 130, 1368–1374. [Google Scholar] [CrossRef] [Green Version]
- Wilflingseder, D.; Banki, Z.; Garcia, E.; Pruenster, M.; Pfister, G.; Muellauer, B.; Nikolic, D.S.; Gassner, C.; Ammann, C.G.; Dierich, M.P.; et al. IgG opsonization of HIV impedes provirus formation in and infection of dendritic cells and subsequent long-term transfer to T cells. J. Immunol. 2007, 178, 7840–7848. [Google Scholar] [CrossRef] [PubMed]
- Bánki, Z.; Stoiber, H.; Dierich, M.P. HIV and human complement: Inefficient virolysis and effective adherence. Immunol. Lett. 2005, 97, 209–214. [Google Scholar] [CrossRef]
- Aiba, S.; Manome, H.; Nakagawa, S.; Mollah, Z.U.; Mizuashi, M.; Ohtani, T.; Yoshino, Y.; Tagami, H. p38 Mitogen-activated protein kinase and extracellular signal-regulated kinases play distinct roles in the activation of dendritic cells by two representative haptens, NiCl2 and 2,4-dinitrochlorobenzene. J. Investig. Dermatol. 2003, 120, 390–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chinsembu, K.C. Chemical diversity and activity profiles of HIV-1 reverse transcriptase inhibitors from plants. Rev. Bras. Farmacogn. 2019, 29, 504–528. [Google Scholar] [CrossRef]
- Steinman, R.M. Decisions about dendritic cells: Past, present, and future. Annu. Rev. Immunol. 2012, 30, 1–22. [Google Scholar] [CrossRef] [Green Version]
- Manel, N.; Hogstad, B.; Wang, Y.; Levy, D.E.; Unutmaz, D.; Littman, D.R. A cryptic sensor for HIV-1 activates antiviral innate immunity in dendritic cells. Nature 2010, 467, 214–217. [Google Scholar] [CrossRef]
- White, T.E.; Brandariz-Nunez, A.; Valle-Casuso, J.C.; Amie, S.; Nguyen, L.; Kim, B.; Brojatsch, J.; Diaz-Griffero, F. Contribution of SAM and HD domains to retroviral restriction mediated by human SAMHD1. Virology 2013, 436, 81–90. [Google Scholar] [CrossRef] [Green Version]
- White, T.E.; Brandariz-Nuñez, A.; Valle-Casuso, J.C.; Amie, S.; Nguyen, L.A.; Kim, B.; Tuzova, M.; Diaz-Griffero, F. The retroviral restriction ability of SAMHD1, but not its deoxynucleotide triphosphohydrolase activity, is regulated by phosphorylation. Cell Host Microbe 2013, 13, 441–451. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rescigno, M.; Martino, M.; Sutherland, C.L.; Gold, M.R.; Ricciardi-Castagnoli, P. Dendritic cell survival and maturation are regulated by different signaling pathways. J. Exp. Med. 1998, 188, 2175–2180. [Google Scholar] [CrossRef] [PubMed]
- Ardeshna, K.M.; Pizzey, A.R.; Devereux, S.; Khwaja, A. The PI3 kinase, p38 SAP kinase, and NF-kappaB signal transduction pathways are involved in the survival and maturation of lipopolysaccharide-stimulated human monocyte-derived dendritic cells. Blood. J. Am. Soc. Hematol. 2000, 96, 1039–1046. [Google Scholar]
- Nijmeijer, B.M.; Bermejo-Jambrina, M.; Kaptein, T.M.; Ribeiro, C.M.S.; Wilflingseder, D.; Geijtenbeek, T.B.H. HIV-1 subverts the complement system in semen to enhance viral transmission. Mucosal. Immunol. 2021, 14, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Mason, H.S.; Thuenemann, E.; Kiyono, H.; Kessans, S.; Matoba, N.; Mor, T. Mucosal Vaccines from Plant Biotechnology. Mucosal. Immunol. 2015, 2, 1271–1289. [Google Scholar] [CrossRef]
- Kessans, S.A.; Linhart, M.D.; Matoba, N.; Mor, T. Biological and biochemical characterization of HIV-1 Gag/dgp41 virus-like particles expressed in Nicotiana benthamiana. Plant Biotechnol. J. 2013, 11, 681–690. [Google Scholar] [CrossRef]
- Posch, W.; Lass-Flörl, C.; Wilflingseder, D. SARS-CoV-2-infected primary human airway epithelia illustrate mucus hypersecretion. J. Allergy Clin. Immunol. 2021. S0091-6749(21)01101-5. [Google Scholar] [CrossRef] [PubMed]
- Posch, W.; Vosper, J.; Zaderer, V.; Noureen, A.; Constant, S.; Bellmann-Weiler, R.; Lass-Flörl, C.; Wilflingseder, D. ColdZyme Maintains Integrity in SARS-CoV-2-Infected Airway Epithelia. mBio 2021, 12, e00904-21. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zaderer, V.; Posch, W.; Gstir, R.; Filipek, P.A.; Bonn, G.K.; Aramwit, P.; Huber, L.A.; Wilflingseder, D. P80 Natural Essence Exerts Efficient Anti-HIV-1- as Well as Adjuvant Effects in DCs. Vaccines 2021, 9, 976. https://doi.org/10.3390/vaccines9090976
Zaderer V, Posch W, Gstir R, Filipek PA, Bonn GK, Aramwit P, Huber LA, Wilflingseder D. P80 Natural Essence Exerts Efficient Anti-HIV-1- as Well as Adjuvant Effects in DCs. Vaccines. 2021; 9(9):976. https://doi.org/10.3390/vaccines9090976
Chicago/Turabian StyleZaderer, Viktoria, Wilfried Posch, Ronald Gstir, Przemyslaw A. Filipek, Günther K. Bonn, Pornanong Aramwit, Lukas A. Huber, and Doris Wilflingseder. 2021. "P80 Natural Essence Exerts Efficient Anti-HIV-1- as Well as Adjuvant Effects in DCs" Vaccines 9, no. 9: 976. https://doi.org/10.3390/vaccines9090976
APA StyleZaderer, V., Posch, W., Gstir, R., Filipek, P. A., Bonn, G. K., Aramwit, P., Huber, L. A., & Wilflingseder, D. (2021). P80 Natural Essence Exerts Efficient Anti-HIV-1- as Well as Adjuvant Effects in DCs. Vaccines, 9(9), 976. https://doi.org/10.3390/vaccines9090976