
The Human Antibody Response to the Influenza Virus Neuraminidase Following Infection or Vaccination



Abstract
1. Introduction
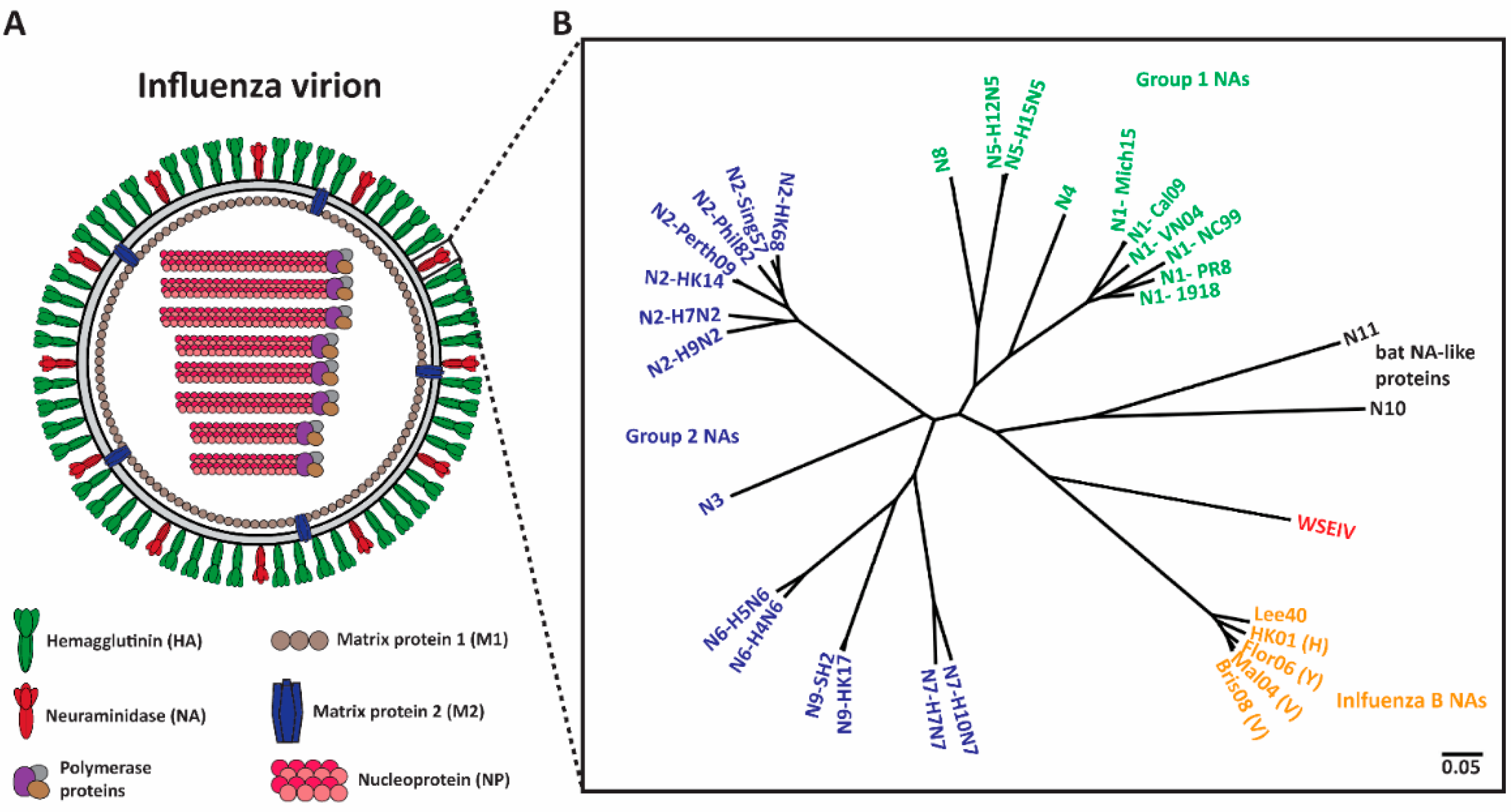
1.1. NA-Based Immunity
1.2. Human mAbs That Target NA
1.2.1. Group 1 and 2 mAbs
1.2.2. Influenza B mAbs
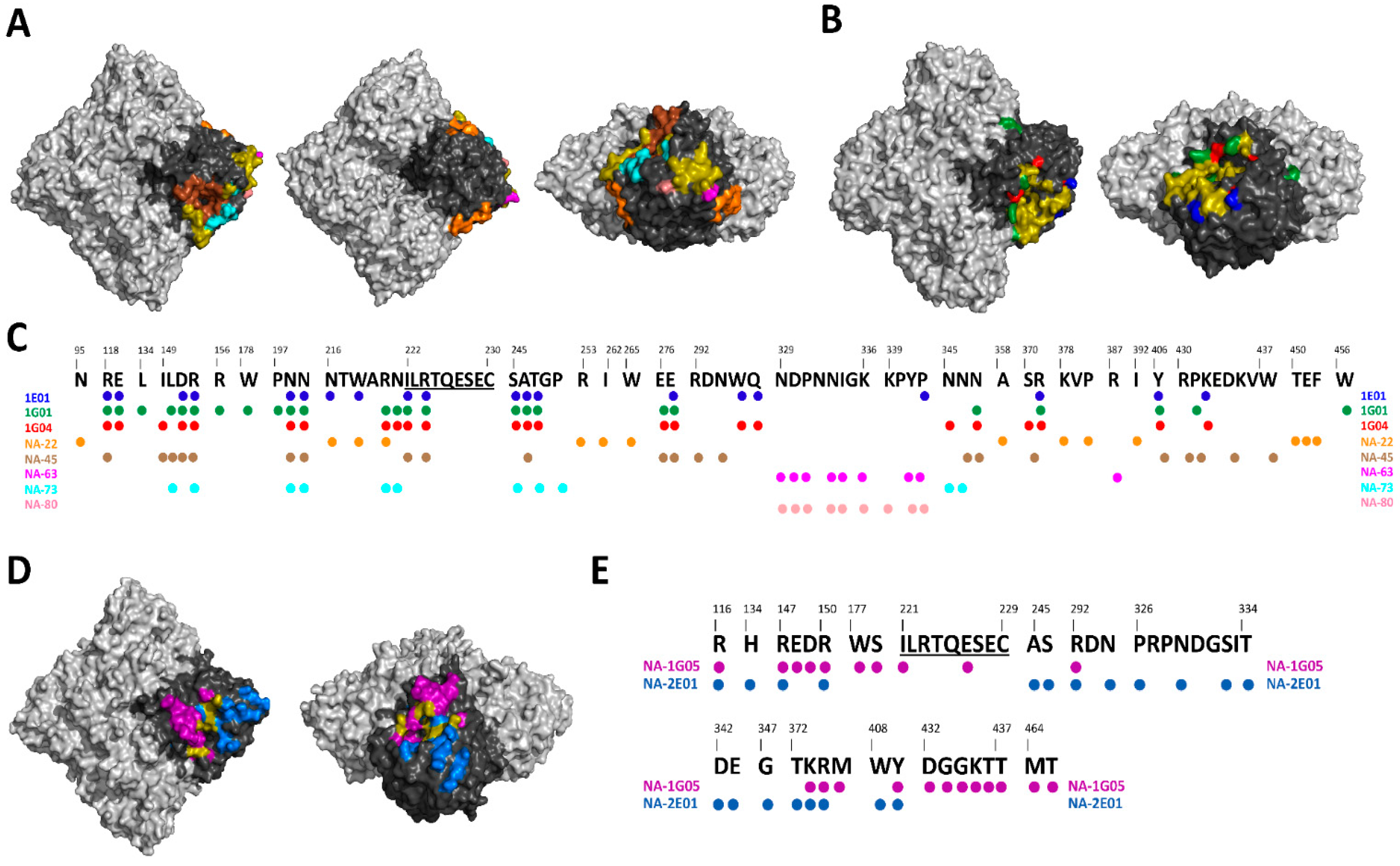
1.2.3. Pan NA mAbs
1.3. NA Human mAbs Inform Vaccine Design
1.4. Emerging Platforms for the Development of NA-Based Vaccines
1.4.1. Modified Inactivated Vaccines
1.4.2. Recombinant NA Vaccines
1.4.3. Virus Like Particles (VLPs)
1.4.4. Viral Replicon Particles (VRPs)
1.4.5. Viral Vector Vaccines
1.4.6. DNA Vaccines
1.4.7. RNA Vaccines
2. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Bernstein, D.I.; Guptill, J.; Naficy, A.; Nachbagauer, R.; Berlanda-Scorza, F.; Feser, J.; Wilson, P.C.; Solorzano, A.; Van der Wielen, M.; Walter, E.B.; et al. Immunogenicity of chimeric haemagglutinin-based, universal influenza virus vaccine candidates: Interim results of a randomised, placebo-controlled, phase 1 clinical trial. Lancet Infect. Dis. 2020, 20, 80–91. [Google Scholar] [CrossRef]
- CDC. Vaccine Effectiveness: How Well Do the Flu Vaccines Work? 2020. Available online: https://www.cdc.gov/flu/vaccines-work/vaccineeffect.htm (accessed on 10 February 2021).
- CDC. Who Is at High Risk for Flu Complications. 2018. Available online: https://www.cdc.gov/flu/highrisk/index.htm (accessed on 1 January 2021).
- WHO. Influenza (Seasonal). 2018. Available online: https://www.who.int/en/news-room/fact-sheets/detail/influenza-(seasonal) (accessed on 22 January 2021).
- Nachbagauer, R.; Liu, W.C.; Choi, A.; Wohlbold, T.J.; Atlas, T.; Rajendran, M.; Solorzano, A.; Berlanda-Scorza, F.; Garcia-Sastre, A.; Palese, P.; et al. A universal influenza virus vaccine candidate confers protection against pandemic H1N1 infection in preclinical ferret studies. NPJ Vaccines 2017, 2, 26. [Google Scholar] [CrossRef]
- Shaw, M.L.; Palese, P. Orthomyxoviridae: The Viruses and Their Replication. In Fields Virology, 6th ed.; Knipe, D.M., Howley, P.M., Eds.; Lippincott Williams & Wilkins: Philadelphia, PA, USA, 2013; pp. 1691–1740. [Google Scholar]
- Bouvier, N.M.; Palese, P. The Biology of Influenza Viruses. Vaccine 2008, 26 (Suppl. S4), D49–D53. [Google Scholar] [CrossRef]
- Heaton, N.S.; Sachs, D.; Chen, C.J.; Hai, R.; Palese, P. Genome-wide mutagenesis of influenza virus reveals unique plasticity of the hemagglutinin and NS1 proteins. Proc. Natl. Acad. Sci. USA 2013, 110, 20248–20253. [Google Scholar] [CrossRef] [PubMed]
- Xie, H.; Wan, X.F.; Ye, Z.; Plant, E.P.; Zhao, Y.; Xu, Y.; Li, X.; Finch, C.; Zhao, N.; Kawano, T.; et al. H3N2 Mismatch of 2014-15 Northern Hemisphere Influenza Vaccines and Head-to-head Comparison between Human and Ferret Antisera derived Antigenic Maps. Sci. Rep. 2015, 5, 15279. [Google Scholar] [CrossRef]
- De Jong, J.C.; Beyer, W.E.; Palache, A.M.; Rimmelzwaan, G.F.; Osterhaus, A.D. Mismatch between the 1997/1998 influenza vaccine and the major epidemic A(H3N2) virus strain as the cause of an inadequate vaccine-induced antibody response to this strain in the elderly. J. Med. Virol. 2000, 61, 94–99. [Google Scholar] [CrossRef]
- CDC. Past Pandemics. 2018. Available online: https://www.cdc.gov/flu/pandemic-resources/basics/past-pandemics.html (accessed on 21 January 2021).
- Krammer, F.; Palese, P. Advances in the development of influenza virus vaccines. Nat. Rev. Drug. Discov. 2015, 14, 167–182. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.J.; Krammer, F. In the shadow of hemagglutinin: A growing interest in influenza viral neuraminidase and its role as a vaccine antigen. Viruses 2014, 6, 2465–2494. [Google Scholar] [CrossRef]
- Eichelberger, M.C.; Wan, H. Influenza neuraminidase as a vaccine antigen. Curr. Top. Microbiol. Immunol. 2015, 386, 275–299. [Google Scholar]
- Matrosovich, M.N.; Matrosovich, T.Y.; Gray, T.; Roberts, N.A.; Klenk, H.D. Neuraminidase is important for the initiation of influenza virus infection in human airway epithelium. J. Virol. 2004, 78, 12665–12667. [Google Scholar] [CrossRef]
- Cohen, M.; Zhang, X.Q.; Senaati, H.P.; Chen, H.W.; Varki, N.M.; Schooley, R.T.; Gagneux, P. Influenza A penetrates host mucus by cleaving sialic acids with neuraminidase. Virol. J. 2013, 10, 321. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Rubin, B.K.; Voynow, J.A. Mucins, Mucus, and Goblet Cells. Chest 2018, 154, 169–176. [Google Scholar] [CrossRef]
- McAuley, J.L.; Gilbertson, B.P.; Trifkovic, S.; Brown, L.E.; McKimm-Breschkin, J.L. Influenza Virus Neuraminidase Structure and Functions. Front. Microbiol. 2019, 10, 39. [Google Scholar] [CrossRef]
- Shtyrya, Y.A.; Mochalova, L.V.; Bovin, N.V. Influenza virus neuraminidase: Structure and function. Acta Nat. 2009, 1, 26–32. [Google Scholar] [CrossRef]
- Krammer, F.; Fouchier, R.A.M.; Eichelberger, M.C.; Webby, R.J.; Shaw-Saliba, K.; Wan, H.; Wilson, P.C.; Compans, R.W.; Skountzou, I.; Monto, A.S. NAction! How Can Neuraminidase-Based Immunity Contribute to Better Influenza Virus Vaccines? mBio 2018, 9, e02332-17. [Google Scholar] [CrossRef]
- Su, B.; Wurtzer, S.; Rameix-Welti, M.A.; Dwyer, D.; van der Werf, S.; Naffakh, N.; Clavel, F.; Labrosse, B. Enhancement of the influenza A hemagglutinin (HA)-mediated cell-cell fusion and virus entry by the viral neuraminidase (NA). PLoS ONE 2009, 4, e8495. [Google Scholar] [CrossRef]
- Kilbourne, E.D.; Johansson, B.E.; Grajower, B. Independent and disparate evolution in nature of influenza A virus hemagglutinin and neuraminidase glycoproteins. Proc. Natl. Acad. Sci. USA 1990, 87, 786–790. [Google Scholar] [CrossRef] [PubMed]
- Westgeest, K.B.; de Graaf, M.; Fourment, M.; Bestebroer, T.M.; van Beek, R.; Spronken, M.I.; de Jong, J.C.; Rimmelzwaan, G.F.; Russell, C.A.; Osterhaus, A.D.; et al. Genetic evolution of the neuraminidase of influenza A (H3N2) viruses from 1968 to 2009 and its correspondence to haemagglutinin evolution. J. Gen. Virol. 2012, 93, 1996–2007. [Google Scholar] [CrossRef]
- Schulman, J.L.; Kilbourne, E.D. Independent variation in nature of hemagglutinin and neuraminidase antigens of influenza virus: Distinctiveness of hemagglutinin antigen of Hong Kong-68 virus. Proc. Natl. Acad. Sci. USA 1969, 63, 326–333. [Google Scholar] [CrossRef] [PubMed]
- Couzens, L.; Gao, J.; Westgeest, K.; Sandbulte, M.; Lugovtsev, V.; Fouchier, R.; Eichelberger, M. An optimized enzyme-linked lectin assay to measure influenza A virus neuraminidase inhibition antibody titers in human sera. J. Virol. Methods 2014, 210, 7–14. [Google Scholar] [CrossRef]
- Schulman, J.L.; Khakpour, M.; Kilbourne, E.D. Protective effects of specific immunity to viral neuraminidase on influenza virus infection of mice. J. Virol. 1968, 2, 778–786. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.J.; Nachbagauer, R.; Xu, H.; Tan, G.S.; Hirsh, A.; Brokstad, K.A.; Cox, R.J.; Palese, P.; Krammer, F. Vaccination with adjuvanted recombinant neuraminidase induces broad heterologous, but not heterosubtypic, cross-protection against influenza virus infection in mice. mBio 2015, 6, e02556. [Google Scholar] [CrossRef]
- Smith, G.E.; Sun, X.; Bai, Y.; Liu, Y.V.; Massare, M.J.; Pearce, M.B.; Belser, J.A.; Maines, T.R.; Creager, H.M.; Glenn, G.M.; et al. Neuraminidase-based recombinant virus-like particles protect against lethal avian influenza A(H5N1) virus infection in ferrets. Virology 2017, 509, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Couch, R.B.; Kasel, J.A.; Gerin, J.L.; Schulman, J.L.; Kilbourne, E.D. Induction of partial immunity to influenza by a neuraminidase-specific vaccine. J. Infect. Dis. 1974, 129, 411–420. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.R.; Kasel, J.A.; Chanock, R.M. Association of serum anti-neuraminidase antibody with resistance to influenza in man. N. Engl. J. Med. 1972, 286, 1329–1332. [Google Scholar] [CrossRef]
- Memoli, M.J.; Shaw, P.A.; Han, A.; Czajkowski, L.; Reed, S.; Athota, R.; Bristol, T.; Fargis, S.; Risos, K.; Powers, J.H.; et al. Evaluation of Antihemagglutinin and Antineuraminidase Antibodies as Correlates of Protection in an Influenza A/H1N1 Virus Healthy Human Challenge Model. mBio 2016, 7, e00417-16. [Google Scholar] [CrossRef]
- Maier, H.E.; Nachbagauer, R.; Kuan, G.; Ng, S.; Lopez, R.; Sanchez, N.; Stadlbauer, D.; Gresh, L.; Schiller, A.; Rajabhathor, A.; et al. Pre-existing anti-neuraminidase antibodies are associated with shortened duration of influenza A (H1N1)pdm virus shedding and illness in naturally infected adults. Clin. Infect. Dis. 2019, 70, 2290–2297. [Google Scholar] [CrossRef]
- Couch, R.B.; Atmar, R.L.; Franco, L.M.; Quarles, J.M.; Wells, J.; Arden, N.; Nino, D.; Belmont, J.W. Antibody correlates and predictors of immunity to naturally occurring influenza in humans and the importance of antibody to the neuraminidase. J. Infect. Dis. 2013, 207, 974–981. [Google Scholar] [CrossRef]
- Monto, A.S.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Liu, M.; Zhong, W.; Levine, M.; Katz, J.M.; Ohmit, S.E. Antibody to Influenza Virus Neuraminidase: An Independent Correlate of Protection. J. Infect. Dis. 2015, 212, 1191–1199. [Google Scholar] [CrossRef]
- Smith, W.; Andrews, C.H.; Laidlaw, P.P. A virus obtained from influenza patients. Lancet 1933, 222, 66–68. [Google Scholar] [CrossRef]
- Johansson, B.E.; Moran, T.M.; Bona, C.A.; Popple, S.W.; Kilbourne, E.D. Immunologic response to influenza virus neuraminidase is influenced by prior experience with the associated viral hemagglutinin. II. Sequential infection of mice simulates human experience. J. Immunol. 1987, 139, 2010–2014. [Google Scholar]
- Chen, Y.Q.; Wohlbold, T.J.; Zheng, N.Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies. Cell 2018, 173, 417–429.e10. [Google Scholar] [CrossRef] [PubMed]
- Nachbagauer, R.; Choi, A.; Hirsh, A.; Margine, I.; Iida, S.; Barrera, A.; Ferres, M.; Albrecht, R.A.; Garcia-Sastre, A.; Bouvier, N.M.; et al. Defining the antibody cross-reactome directed against the influenza virus surface glycoproteins. Nat. Immunol. 2017, 18, 464–473. [Google Scholar] [CrossRef] [PubMed]
- Changsom, D.; Jiang, L.; Lerdsamran, H.; Iamsirithaworn, S.; Kitphati, R.; Pooruk, P.; Auewarakul, P.; Puthavathana, P. Kinetics, Longevity, and Cross-Reactivity of Antineuraminidase Antibody after Natural Infection with Influenza A Viruses. Clin. Vaccine Immunol. 2017, 24, e00248-17. [Google Scholar] [CrossRef]
- Smith, A.J.; Davies, J.R. Natural infection with influenza A (H3N2). The development, persistance and effect of antibodies to the surface antigens. Epidemiol. Infect. 1976, 77, 271–282. [Google Scholar] [CrossRef] [PubMed]
- Schild, G.C. Antibody against influenza A2 virus neuraminidase in human sera. J. Hyg. 1969, 67, 353–365. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Rajendran, M.; Nachbagauer, R.; Ermler, M.E.; Bunduc, P.; Amanat, F.; Izikson, R.; Cox, M.; Palese, P.; Eichelberger, M.; Krammer, F. Analysis of Anti-Influenza Virus Neuraminidase Antibodies in Children, Adults, and the Elderly by ELISA and Enzyme Inhibition: Evidence for Original Antigenic Sin. mBio 2017, 8. [Google Scholar] [CrossRef]
- Gao, Z.; Robinson, K.; Skowronski, D.M.; De Serres, G.; Withers, S.G. Quantification of the total neuraminidase content of recent commercially-available influenza vaccines: Introducing a neuraminidase titration reagent. Vaccine 2020, 38, 715–718. [Google Scholar] [CrossRef]
- Petrie, J.G.; Ohmit, S.E.; Johnson, E.; Truscon, R.; Monto, A.S. Persistence of Antibodies to Influenza Hemagglutinin and Neuraminidase Following One or Two Years of Influenza Vaccination. J. Infect. Dis. 2015, 212, 1914–1922. [Google Scholar] [CrossRef]
- Ehrlich, H.J.; Muller, M.; Kollaritsch, H.; Pinl, F.; Schmitt, B.; Zeitlinger, M.; Loew-Baselli, A.; Kreil, T.R.; Kistner, O.; Portsmouth, D.; et al. Pre-vaccination immunity and immune responses to a cell culture-derived whole-virus H1N1 vaccine are similar to a seasonal influenza vaccine. Vaccine 2012, 30, 4543–4551. [Google Scholar] [CrossRef]
- Couch, R.B.; Atmar, R.L.; Keitel, W.A.; Quarles, J.M.; Wells, J.; Arden, N.; Nino, D. Randomized comparative study of the serum antihemagglutinin and antineuraminidase antibody responses to six licensed trivalent influenza vaccines. Vaccine 2012, 31, 190–195. [Google Scholar] [CrossRef]
- Laguio-Vila, M.R.; Thompson, M.G.; Reynolds, S.; Spencer, S.M.; Gaglani, M.; Naleway, A.; Ball, S.; Bozeman, S.; Baker, S.; Martinez-Sobrido, L.; et al. Comparison of serum hemagglutinin and neuraminidase inhibition antibodies after 2010-2011 trivalent inactivated influenza vaccination in healthcare personnel. Open Forum Infect. Dis. 2015, 2, ofu115. [Google Scholar] [CrossRef]
- Gross, P.A.; Russo, C.; Dran, S.; Cataruozolo, P.; Munk, G.; Lancey, S.C. Time to earliest peak serum antibody response to influenza vaccine in the elderly. Clin. Diagn. Lab. Immunol. 1997, 4, 491–492. [Google Scholar] [CrossRef] [PubMed]
- Gross, P.A.; Russo, C.; Teplitzky, M.; Dran, S.; Cataruozolo, P.; Munk, G. Time to peak serum antibody response to influenza vaccine in the elderly. Clin. Diagn. Lab. Immunol. 1996, 3, 361–362. [Google Scholar] [CrossRef]
- Rastogi, S.; Gross, P.A.; Bonelli, J.; Dran, S.; Levandowski, R.A.; Russo, C.; Weksler, M.E.; Kaye, D.; Levison, M.; Abrutyn, E.; et al. Time to peak serum antibody response to influenza vaccine. Clin. Diagn. Lab. Immunol. 1995, 2, 120–121. [Google Scholar] [CrossRef]
- Kositanont, U.; Assantachai, P.; Wasi, C.; Puthavathana, P.; Praditsuwan, R. Kinetics of the antibody response to seasonal influenza vaccination among the elderly. Viral Immunol. 2012, 25, 471–476. [Google Scholar] [CrossRef]
- Mohn, K.G.; Smith, I.; Sjursen, H.; Cox, R.J. Immune responses after live attenuated influenza vaccination. Hum. Vaccin. Immunother. 2018, 14, 571–578. [Google Scholar] [CrossRef] [PubMed]
- Ghendon, Y. The immune response to influenza vaccines. Acta Virol. 1990, 34, 295–304. [Google Scholar]
- Belongia, E.A.; Sundaram, M.E.; McClure, D.L.; Meece, J.K.; Ferdinands, J.; VanWormer, J.J. Waning vaccine protection against influenza A (H3N2) illness in children and older adults during a single season. Vaccine 2015, 33, 246–251. [Google Scholar] [CrossRef]
- Puig-Barbera, J.; Mira-Iglesias, A.; Tortajada-Girbes, M.; Lopez-Labrador, F.X.; Librero-Lopez, J.; Diez-Domingo, J.; Carballido-Fernandez, M.; Carratala-Munuera, C.; Correcher-Medina, P.; Gil-Guillen, V.; et al. Waning protection of influenza vaccination during four influenza seasons, 2011/2012 to 2014/2015. Vaccine 2017, 35, 5799–5807. [Google Scholar] [CrossRef] [PubMed]
- Kirkpatrick, E.; Qiu, X.; Wilson, P.C.; Bahl, J.; Krammer, F. The influenza virus hemagglutinin head evolves faster than the stalk domain. Sci. Rep. 2018, 8, 10432. [Google Scholar] [CrossRef]
- Bangaru, S.; Lang, S.; Schotsaert, M.; Vanderven, H.A.; Zhu, X.; Kose, N.; Bombardi, R.; Finn, J.A.; Kent, S.J.; Gilchuk, P.; et al. A Site of Vulnerability on the Influenza Virus Hemagglutinin Head Domain Trimer Interface. Cell 2019, 177, 1136–1152.e18. [Google Scholar] [CrossRef] [PubMed]
- Corti, D.; Cameroni, E.; Guarino, B.; Kallewaard, N.L.; Zhu, Q.; Lanzavecchia, A. Tackling influenza with broadly neutralizing antibodies. Curr. Opin. Virol. 2017, 24, 60–69. [Google Scholar] [CrossRef]
- Tay, M.Z.; Wiehe, K.; Pollara, J. Antibody-Dependent Cellular Phagocytosis in Antiviral Immune Responses. Front. Immunol. 2019, 10, 332. [Google Scholar] [CrossRef] [PubMed]
- Jegaskanda, S. The Potential Role of Fc-Receptor Functions in the Development of a Universal Influenza Vaccine. Vaccines 2018, 6, 27. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Tan, H.X.; Koutsakos, M.; Jegaskanda, S.; Esterbauer, R.; Tilmanis, D.; Aban, M.; Kedzierska, K.; Hurt, A.C.; Kent, S.J.; et al. Cross-lineage protection by human antibodies binding the influenza B hemagglutinin. Nat. Commun. 2019, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Whittle, J.R.; Zhang, R.; Khurana, S.; King, L.R.; Manischewitz, J.; Golding, H.; Dormitzer, P.R.; Haynes, B.F.; Walter, E.B.; Moody, M.A. Broadly neutralizing human antibody that recognizes the receptor-binding pocket of influenza virus hemagglutinin. Proc. Natl. Acad. Sci. USA 2011, 108, 14216–14221. [Google Scholar] [CrossRef]
- Zost, S.J.; Lee, J.; Gumina, M.E.; Parkhouse, K.; Henry, C.; Wu, N.C.; Lee, C.D.; Wilson, I.A.; Wilson, P.C.; Bloom, J.D.; et al. Identification of Antibodies Targeting the H3N2 Hemagglutinin Receptor Binding Site following Vaccination of Humans. Cell 2019, 29, 4460–4470.e8. [Google Scholar]
- Shen, C.; Zhang, M.; Chen, Y.; Zhang, L.; Wang, G.; Chen, J.; Chen, S.; Li, Z.; Wei, F.; Chen, J.; et al. An IgM antibody targeting the receptor binding site of influenza B blocks viral infection with great breadth and potency. Theranostics 2019, 9, 210–231. [Google Scholar] [CrossRef]
- Krammer, F.; Palese, P. Influenza virus hemagglutinin stalk-based antibodies and vaccines. Curr. Opin. Virol. 2013, 3, 521–530. [Google Scholar] [CrossRef]
- Rajendran, M.; Sun, W.; Comella, P.; Nachbagauer, R.; Wohlbold, T.J.; Amanat, F.; Kirkpatrick, E.; Palese, P.; Krammer, F. An immuno-assay to quantify influenza virus hemagglutinin with correctly folded stalk domains in vaccine preparations. PLoS ONE 2018, 13, e0194830. [Google Scholar] [CrossRef] [PubMed]
- Sui, J.; Hwang, W.C.; Perez, S.; Wei, G.; Aird, D.; Chen, L.M.; Santelli, E.; Stec, B.; Cadwell, G.; Ali, M.; et al. Structural and functional bases for broad-spectrum neutralization of avian and human influenza A viruses. Nat. Struct. Mol. Biol. 2009, 16, 265–273. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, D.C.; Bhabha, G.; Elsliger, M.A.; Friesen, R.H.; Jongeneelen, M.; Throsby, M.; Goudsmit, J.; Wilson, I.A. Antibody recognition of a highly conserved influenza virus epitope. Science 2009, 324, 246–251. [Google Scholar] [CrossRef] [PubMed]
- Ekiert, D.C.; Friesen, R.H.; Bhabha, G.; Kwaks, T.; Jongeneelen, M.; Yu, W.; Ophorst, C.; Cox, F.; Korse, H.J.; Brandenburg, B.; et al. A highly conserved neutralizing epitope on group 2 influenza A viruses. Science 2011, 333, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Gilchuk, I.M.; Bangaru, S.; Gilchuk, P.; Irving, R.P.; Kose, N.; Bombardi, R.G.; Thornburg, N.J.; Creech, C.B.; Edwards, K.M.; Li, S.; et al. Influenza H7N9 Virus Neuraminidase-Specific Human Monoclonal Antibodies Inhibit Viral Egress and Protect from Lethal Influenza Infection in Mice. Cell Host Microbe 2019, 26, 715–728.e8. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Fantoni, G.; Couzens, L.; Gao, J.; Plant, E.; Ye, Z.; Eichelberger, M.C.; Wan, H. Comparative Efficacy of Monoclonal Antibodies That Bind to Different Epitopes of the 2009 Pandemic H1N1 Influenza Virus Neuraminidase. J. Virol. 2016, 90, 117–128. [Google Scholar] [CrossRef]
- Wan, H.; Gao, J.; Xu, K.; Chen, H.; Couzens, L.K.; Rivers, K.H.; Easterbrook, J.D.; Yang, K.; Zhong, L.; Rajabi, M.; et al. Molecular basis for broad neuraminidase immunity: Conserved epitopes in seasonal and pandemic H1N1 as well as H5N1 influenza viruses. J. Virol. 2013, 87, 9290–9300. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.J.; Chromikova, V.; Tan, G.S.; Meade, P.; Amanat, F.; Comella, P.; Hirsh, A.; Krammer, F. Hemagglutinin Stalk- and Neuraminidase-Specific Monoclonal Antibodies Protect against Lethal H10N8 Influenza Virus Infection in Mice. J. Virol. 2016, 90, 851–861. [Google Scholar] [CrossRef]
- Shcherbik, S.; Carney, P.; Pearce, N.; Stevens, J.; Dugan, V.G.; Wentworth, D.E.; Bousse, T. Monoclonal antibody against N2 neuraminidase of cold adapted A/Leningrad/134/17/57 (H2N2) enables efficient generation of live attenuated influenza vaccines. Virology 2018, 522, 65–72. [Google Scholar] [CrossRef]
- Job, E.R.; Ysenbaert, T.; Smet, A.; Van Hecke, A.; Meuris, L.; Kleanthous, H.; Saelens, X.; Vogel, T.U. Fcgamma Receptors Contribute to the Antiviral Properties of Influenza Virus Neuraminidase-Specific Antibodies. mBio 2019, 10, e01667-19. [Google Scholar] [CrossRef]
- Von Holle, T.A.; Moody, M.A. Influenza and Antibody-Dependent Cellular Cytotoxicity. Front. Immunol. 2019, 10, 1457. [Google Scholar] [CrossRef]
- Jegaskanda, S.; Reading, P.C.; Kent, S.J. Influenza-specific antibody-dependent cellular cytotoxicity: Toward a universal influenza vaccine. J. Immunol. 2014, 193, 469–475. [Google Scholar] [CrossRef]
- Jegaskanda, S.; Weinfurter, J.T.; Friedrich, T.C.; Kent, S.J. Antibody-dependent cellular cytotoxicity is associated with control of pandemic H1N1 influenza virus infection of macaques. J. Virol. 2013, 87, 5512–5522. [Google Scholar] [CrossRef]
- Valkenburg, S.A.; Fang, V.J.; Leung, N.H.; Chu, D.K.; Ip, D.K.; Perera, R.A.; Wang, Y.; Li, A.P.; Peiris, J.M.; Cowling, B.J.; et al. Cross-reactive antibody-dependent cellular cytotoxicity antibodies are increased by recent infection in a household study of influenza transmission. Clin. Transl. Immunol. 2019, 8, e1092. [Google Scholar] [CrossRef]
- Rijal, P.; Wang, B.B.; Tan, T.K.; Schimanski, L.; Janesch, P.; Dong, T.; McCauley, J.W.; Daniels, R.S.; Townsend, A.R.; Huang, K.A. Broadly Inhibiting Antineuraminidase Monoclonal Antibodies Induced by Trivalent Influenza Vaccine and H7N9 Infection in Humans. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.; Turner, H.L.; Lang, S.; McBride, R.; Bangaru, S.; Gilchuk, I.M.; Yu, W.; Paulson, J.C.; Crowe, J.E., Jr.; Ward, A.B.; et al. Structural Basis of Protection against H7N9 Influenza Virus by Human Anti-N9 Neuraminidase Antibodies. Cell Host Microbe 2019, 26, 729–738.e4. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Li, L.; Wilson, P.C. Emerging from the Shadow of Hemagglutinin: Neuraminidase Is an Important Target for Influenza Vaccination. Cell Host Microbe 2019, 26, 712–713. [Google Scholar] [CrossRef] [PubMed]
- Madsen, A.; Dai, Y.N.; McMahon, M.; Schmitz, A.J.; Turner, J.S.; Tan, J.; Lei, T.; Alsoussi, W.B.; Strohmeier, S.; Amor, M.; et al. Human Antibodies Targeting Influenza B Virus Neuraminidase Active Site Are Broadly Protective. Immunity 2020, 53, 852–863.e7. [Google Scholar] [CrossRef] [PubMed]
- Piepenbrink, M.S.; Nogales, A.; Basu, M.; Fucile, C.F.; Liesveld, J.L.; Keefer, M.C.; Rosenberg, A.F.; Martinez-Sobrido, L.; Kobie, J.J. Broad and Protective Influenza B Virus Neuraminidase Antibodies in Humans after Vaccination and their Clonal Persistence as Plasma Cells. mBio 2019, 10, e00066-19. [Google Scholar] [CrossRef]
- Stadlbauer, D.; Zhu, X.; McMahon, M.; Turner, J.S.; Wohlbold, T.J.; Schmitz, A.J.; Strohmeier, S.; Yu, W.; Nachbagauer, R.; Mudd, P.A.; et al. Broadly protective human antibodies that target the active site of influenza virus neuraminidase. Science 2019, 366, 499–504. [Google Scholar] [CrossRef] [PubMed]
- Arunkumar, G.A.; Strohmeier, S.; Li, T.; Bhavsar, D.; Chromikova, V.; Amanat, F.; Bunyatov, M.; Wilson, P.C.; Ellebedy, A.H.; Boons, G.-J.; et al. Reactions of antibodies with surface antigens of influenza virus. J. Gen. Virol. 1968, 3, 315–326. [Google Scholar]
- Webster, R.G.; Laver, W.G.; Kilbourne, E.D. Reactions of Antibodies with Surface Antigens of Influenza Virus. J. Gen. Virol. 1968, 3, 315–326. [Google Scholar] [CrossRef] [PubMed]
- Westgeest, K.B.; Russell, C.A.; Lin, X.; Spronken, M.I.; Bestebroer, T.M.; Bahl, J.; van Beek, R.; Skepner, E.; Halpin, R.A.; de Jong, J.C.; et al. Genomewide analysis of reassortment and evolution of human influenza A(H3N2) viruses circulating between 1968 and 2011. J. Virol. 2014, 88, 2844–2857. [Google Scholar] [CrossRef] [PubMed]
- Hensley, S.E.; Das, S.R.; Gibbs, J.S.; Bailey, A.L.; Schmidt, L.M.; Bennink, J.R.; Yewdell, J.W. Influenza A virus hemagglutinin antibody escape promotes neuraminidase antigenic variation and drug resistance. PLoS ONE 2011, 6, e15190. [Google Scholar] [CrossRef] [PubMed]
- Johansson, B.E.; Bucher, D.J.; Kilbourne, E.D. Purified influenza virus hemagglutinin and neuraminidase are equivalent in stimulation of antibody response but induce contrasting types of immunity to infection. J. Virol. 1989, 63, 1239–1246. [Google Scholar] [CrossRef]
- Matthew, J.; Syle, D.L.S. Influenza Neuraminidase as a Vaccine Antigen. In Vaccines for Pandemic Influenza; Richard, W., Compans, W.A.O., Eds.; Springer: Berlin, Germany, 2009; Volume 333, p. 512. [Google Scholar]
- Jagadesh, A.; Salam, A.A.; Mudgal, P.P.; Arunkumar, G. Influenza virus neuraminidase (NA): A target for antivirals and vaccines. Arch. Virol. 2016, 161, 2087–2094. [Google Scholar] [CrossRef]
- Getie-Kebtie, M.; Sultana, I.; Eichelberger, M.; Alterman, M. Label-free mass spectrometry-based quantification of hemagglutinin and neuraminidase in influenza virus preparations and vaccines. Influenza Other Respir. Viruses 2013, 7, 521–530. [Google Scholar] [CrossRef]
- Williams, T.L.; Pirkle, J.L.; Barr, J.R. Simultaneous quantification of hemagglutinin and neuraminidase of influenza virus using isotope dilution mass spectrometry. Vaccine 2012, 30, 2475–2482. [Google Scholar] [CrossRef]
- Wan, H.; Sultana, I.; Couzens, L.K.; Mindaye, S.; Eichelberger, M.C. Assessment of influenza A neuraminidase (subtype N1) potency by ELISA. J. Virol. Methods 2017, 244, 23–28. [Google Scholar] [CrossRef]
- Broecker, F.; Zheng, A.; Suntronwong, N.; Sun, W.; Bailey, M.J.; Krammer, F.; Palese, P. Extending the Stalk Enhances Immunogenicity of the Influenza Virus Neuraminidase. J. Virol. 2019, 93. [Google Scholar] [CrossRef]
- Kilbourne, E.D.; Couch, R.B.; Kasel, J.A.; Keitel, W.A.; Cate, T.R.; Quarles, J.H.; Grajower, B.; Pokorny, B.A.; Johansson, B.E. Purified influenza A virus N2 neuraminidase vaccine is immunogenic and non-toxic in humans. Vaccine 1995, 13, 1799–1803. [Google Scholar] [CrossRef]
- Liu, W.C.; Lin, C.Y.; Tsou, Y.T.; Jan, J.T.; Wu, S.C. Cross-Reactive Neuraminidase-Inhibiting Antibodies Elicited by Immunization with Recombinant Neuraminidase Proteins of H5N1 and Pandemic H1N1 Influenza A Viruses. J. Virol. 2015, 89, 7224–7234. [Google Scholar] [CrossRef] [PubMed]
- Job, E.R.; Ysenbaert, T.; Smet, A.; Christopoulou, I.; Strugnell, T.; Oloo, E.O.; Oomen, R.P.; Kleanthous, H.; Vogel, T.U.; Saelens, X. Broadened immunity against influenza by vaccination with computationally designed influenza virus N1 neuraminidase constructs. NPJ Vaccines 2018, 3, 55. [Google Scholar] [CrossRef] [PubMed]
- Brett, I.C.; Johansson, B.E. Immunization against influenza A virus: Comparison of conventional inactivated, live-attenuated and recombinant baculovirus produced purified hemagglutinin and neuraminidase vaccines in a murine model system. Virology 2005, 339, 273–280. [Google Scholar] [CrossRef] [PubMed]
- Johansson, B.E.; Brett, I.C. Recombinant influenza B virus HA and NA antigens administered in equivalent amounts are immunogenically equivalent and induce equivalent homotypic and broader heterovariant protection in mice than conventional and live influenza vaccines. Hum. Vaccin. 2008, 4, 420–424. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.; Kirkpatrick, E.; Stadlbauer, D.; Strohmeier, S.; Bouvier, N.M.; Krammer, F. Mucosal Immunity against Neuraminidase Prevents Influenza B Virus Transmission in Guinea Pigs. mBio 2019, 10. [Google Scholar] [CrossRef]
- Kim, K.H.; Lee, Y.T.; Park, S.; Jung, Y.J.; Lee, Y.; Ko, E.J.; Kim, Y.J.; Li, X.; Kang, S.M. Neuraminidase expressing virus-like particle vaccine provides effective cross protection against influenza virus. Virology 2019, 535, 179–188. [Google Scholar] [CrossRef] [PubMed]
- Easterbrook, J.D.; Schwartzman, L.M.; Gao, J.; Kash, J.C.; Morens, D.M.; Couzens, L.; Wan, H.; Eichelberger, M.C.; Taubenberger, J.K. Protection against a lethal H5N1 influenza challenge by intranasal immunization with virus-like particles containing 2009 pandemic H1N1 neuraminidase in mice. Virology 2012, 432, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Halbherr, S.J.; Ludersdorfer, T.H.; Ricklin, M.; Locher, S.; Berger Rentsch, M.; Summerfield, A.; Zimmer, G. Biological and protective properties of immune sera directed to the influenza virus neuraminidase. J. Virol. 2015, 89, 1550–1563. [Google Scholar] [CrossRef]
- Mooney, A.J.; Gabbard, J.D.; Li, Z.; Dlugolenski, D.A.; Johnson, S.K.; Tripp, R.A.; He, B.; Tompkins, S.M. Vaccination with Recombinant Parainfluenza Virus 5 Expressing Neuraminidase Protects against Homologous and Heterologous Influenza Virus Challenge. J. Virol. 2017, 91. [Google Scholar] [CrossRef] [PubMed]
- Meseda, C.A.; Atukorale, V.; Soto, J.; Eichelberger, M.C.; Gao, J.; Wang, W.; Weiss, C.D.; Weir, J.P. Immunogenicity and Protection Against Influenza H7N3 in Mice by Modified Vaccinia Virus Ankara Vectors Expressing Influenza Virus Hemagglutinin or Neuraminidase. Sci. Rep. 2018, 8, 5364. [Google Scholar] [CrossRef] [PubMed]
- Sandbulte, M.R.; Jimenez, G.S.; Boon, A.C.; Smith, L.R.; Treanor, J.J.; Webby, R.J. Cross-reactive neuraminidase antibodies afford partial protection against H5N1 in mice and are present in unexposed humans. PLoS Med. 2007, 4, e59. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Kadowaki, S.; Hagiwara, Y.; Yoshikawa, T.; Matsuo, K.; Kurata, T.; Tamura, S. Cross-protection against a lethal influenza virus infection by DNA vaccine to neuraminidase. Vaccine 2000, 18, 3214–3222. [Google Scholar] [CrossRef]
- Freyn, A.W.; Ramos da Silva, J.; Rosado, V.C.; Bliss, C.M.; Pine, M.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; de Souza Ferreira, L.C.; Weissman, D.; et al. A Multi-Targeting, Nucleoside-Modified mRNA Influenza Virus Vaccine Provides Broad Protection in Mice. Mol. Ther. 2020, 28, 1569–1584. [Google Scholar] [CrossRef] [PubMed]
- Nicholson, K.G.; Webster, R.G.; Hay, A. Standardization of Inactivated Influenza Vaccines. In Textbook of Influenza; Woods, J.M., Ed.; Wiley-Blackwell: Hoboken, NJ, USA, 1998; pp. 333–345. [Google Scholar]
- Van Deusen, R.A.; Hinshaw, V.S.; Senne, D.A.; Pellacani, D. Micro neuraminidase-inhibition assay for classification of influenza A virus neuraminidases. Avian Dis. 1983, 27, 745–750. [Google Scholar] [CrossRef]
- Aminoff, D. Methods for the quantitative estimation of N-acetylneuraminic acid and their application to hydrolysates of sialomucoids. Biochem. J. 1961, 81, 384–392. [Google Scholar] [CrossRef]
- Eichelberger, M.C.; Monto, A.S. Neuraminidase, the Forgotten Surface Antigen, Emerges as an Influenza Vaccine Target for Broadened Protection. J. Infect. Dis. 2019, 219, S75–S80. [Google Scholar] [CrossRef]
- Gomez Lorenzo, M.M.; Fenton, M.J. Immunobiology of influenza vaccines. Chest 2013, 143, 502–510. [Google Scholar] [CrossRef] [PubMed]
- Sultana, I.; Yang, K.; Getie-Kebtie, M.; Couzens, L.; Markoff, L.; Alterman, M.; Eichelberger, M.C. Stability of neuraminidase in inactivated influenza vaccines. Vaccine 2014, 32, 2225–2230. [Google Scholar] [CrossRef]
- Zheng, A.; Sun, W.; Xiong, X.; Freyn, A.W.; Peukes, J.; Strohmeier, S.; Nachbagauer, R.; Briggs, J.A.G.; Krammer, F.; Palese, P. Enhancing Neuraminidase Immunogenicity of Influenza A Viruses by Rewiring RNA Packaging Signals. J. Virol. 2020, 94. [Google Scholar] [CrossRef]
- Roldao, A.; Mellado, M.C.; Castilho, L.R.; Carrondo, M.J.; Alves, P.M. Virus-like particles in vaccine development. Expert Rev. Vaccines 2010, 9, 1149–1176. [Google Scholar] [CrossRef] [PubMed]
- Jegerlehner, A.; Zabel, F.; Langer, A.; Dietmeier, K.; Jennings, G.T.; Saudan, P.; Bachmann, M.F. Bacterially produced recombinant influenza vaccines based on virus-like particles. PLoS ONE 2013, 8, e78947. [Google Scholar] [CrossRef]
- Zhao, Q.; Li, S.; Yu, H.; Xia, N.; Modis, Y. Virus-like particle-based human vaccines: Quality assessment based on structural and functional properties. Trends Biotechnol. 2013, 31, 654–663. [Google Scholar] [CrossRef] [PubMed]
- Lundstrom, K. Replicon RNA Viral Vectors as Vaccines. Vaccines 2016, 4, 39. [Google Scholar] [CrossRef]
- Zimmer, G. RNA replicons—A new approach for influenza virus immunoprophylaxis. Viruses 2010, 2, 413–434. [Google Scholar] [CrossRef]
- Hoelscher, M.A.; Garg, S.; Bangari, D.S.; Belser, J.A.; Lu, X.; Stephenson, I.; Bright, R.A.; Katz, J.M.; Mittal, S.K.; Sambhara, S. Development of adenoviral-vector-based pandemic influenza vaccine against antigenically distinct human H5N1 strains in mice. Lancet 2006, 367, 475–481. [Google Scholar] [CrossRef]
- Yang, S.G.; Wo, J.E.; Li, M.W.; Mi, F.F.; Yu, C.B.; Lv, G.L.; Cao, H.C.; Lu, H.F.; Wang, B.H.; Zhu, H.; et al. Construction and cellular immune response induction of HA-based alphavirus replicon vaccines against human-avian influenza (H5N1). Vaccine 2009, 27, 7451–7458. [Google Scholar] [CrossRef]
- Choi, Y.; Chang, J. Viral vectors for vaccine applications. Clin. Exp. Vaccine Res. 2013, 2, 97–105. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Jacob, J. DNA vaccines against influenza viruses. Curr. Top. Microbiol. Immunol. 2009, 333, 197–210. [Google Scholar]
- Wolff, J.A.; Malone, R.W.; Williams, P.; Chong, W.; Acsadi, G.; Jani, A.; Felgner, P.L. Direct gene transfer into mouse muscle in vivo. Science 1990, 247, 1465–1468. [Google Scholar] [CrossRef] [PubMed]
- Ulmer, J.B.; Mason, P.W.; Geall, A.; Mandl, C.W. RNA-based vaccines. Vaccine 2012, 30, 4414–4418. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez-Gascon, A.; del Pozo-Rodriguez, A.; Solinis, M.A. Development of nucleic acid vaccines: Use of self-amplifying RNA in lipid nanoparticles. Int. J. Nanomed. 2014, 9, 1833–1843. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Reactivity | Ref. | mAb Name | Induced after |
|---|---|---|---|
| Group 1 NA | [37] | 1000-3C05, 1000-2E06, 1000-3B04, 1000-3B06, EM-2E01, 1000-1D05, 1000-1E02, 1000-1H01, 294-G-1F01, 294-A-1C02, 295-G-2F04, 300-G-2A04, 300-G-2F04, 294-A-1C06 | H1N1 infection |
| [80] | AG7C, AF9C | Seasonal trivalent inactivated vaccine | |
| Group 2 NA | [37] | 229-1D05, 235-1C02, 235-1E06, 294-1A02, 228-1B03, 228-3F04, 2291B05, 229-1F06, 229-1G03, 229-2B04, 229-2C06, 229-2E02 | H3N2 infection |
| [70,81,82] | NA-97 | A/British Columbia/1/2015 (H7N9) natural infection | |
| [70,81,82] | NA-22, NA-45, NA-63, NA-73, NA-80 | A/Shanghai/2/2013 (H7N9) monovalent inactivated influenza vaccine | |
| Influenza B NA | [83] | NA-1A03, NA-1G05, NA-2D10, NA-2E01, NA-2H09, NA-3C01 | Influenza B infection |
| [84] | 1086C12, 1092D4, 1092E10, 1122C7 | Quadrivalent inactivated influenza vaccine | |
| Pan NA | 1G01, 1E01, 1G04 | H3N2 infection |
| Platform | NA Antigen Subtype | Animal Model | Immunogenicity | Protection | Ref. |
|---|---|---|---|---|---|
| Inactivated vaccine | 30 AA insertion in seasonal N1 15 AA insertion in N2 | Mice | +++ +++ | N.D. | [96] |
| Recombinant NA vaccine | N2 | Human | + | N.D. | [97] |
| Seasonal N1 N2 B/Yamagata/16/88-like B-NA | Mice | +++ +++ +++ | Homologous Heterologous | [27] | |
| Avian N1 Pandemic N1 | Mice | + | Homologous Partial heterologous | [98] | |
| N1 | Mice | +++ | Homologous | [99] | |
| N2 | Mice | +++ | Homologous Partial heterologous | [100] | |
| B-NA | Mice Guinea pigs | + | Homologous Heterologous | [101] | |
| B-NA | Guinea pigs | +++ | Homologous Partial heterologous | [102] | |
| Virus like particles | Avian N1 | Ferrets | +++ | Homologous | [28] |
| Pandemic N1 | Mice | + | Homologous Heterologous | [103] | |
| Avian N1 Seasonal N1 | Mice | +++ | Homologous Heterologous | [104] | |
| Viral replicon particles | Avian N1 | Chicken | +++ | N.D. | [105] |
| Viral Vector vaccines | Avian N1 Pandemic N1 | Mice | +++ | Homologous Heterologous Heterosubtypic | [106] |
| N3 N9 | Mice | +++ | Homologous | [107] | |
| Nucleic Acid-DNA | Seasonal N1 | Mice | + | Homologous Partial heterologous | [108] |
| N2 | Mice | + | Homologous Partial heterologous | [109] | |
| Nucleic Acid-RNA | Seasonal N1 | Mice | +++ | Homologous | [110] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajendran, M.; Krammer, F.; McMahon, M. The Human Antibody Response to the Influenza Virus Neuraminidase Following Infection or Vaccination. Vaccines 2021, 9, 846. https://doi.org/10.3390/vaccines9080846
Rajendran M, Krammer F, McMahon M. The Human Antibody Response to the Influenza Virus Neuraminidase Following Infection or Vaccination. Vaccines. 2021; 9(8):846. https://doi.org/10.3390/vaccines9080846
Chicago/Turabian StyleRajendran, Madhusudan, Florian Krammer, and Meagan McMahon. 2021. "The Human Antibody Response to the Influenza Virus Neuraminidase Following Infection or Vaccination" Vaccines 9, no. 8: 846. https://doi.org/10.3390/vaccines9080846
APA StyleRajendran, M., Krammer, F., & McMahon, M. (2021). The Human Antibody Response to the Influenza Virus Neuraminidase Following Infection or Vaccination. Vaccines, 9(8), 846. https://doi.org/10.3390/vaccines9080846