
Animal Models Utilized for the Development of Influenza Virus Vaccines


Abstract
1. Introduction
2. Seasonal Influenza Virus Vaccines
3. Universal Influenza Virus Vaccines
4. Animal Models Used in Influenza Virus Vaccine Studies
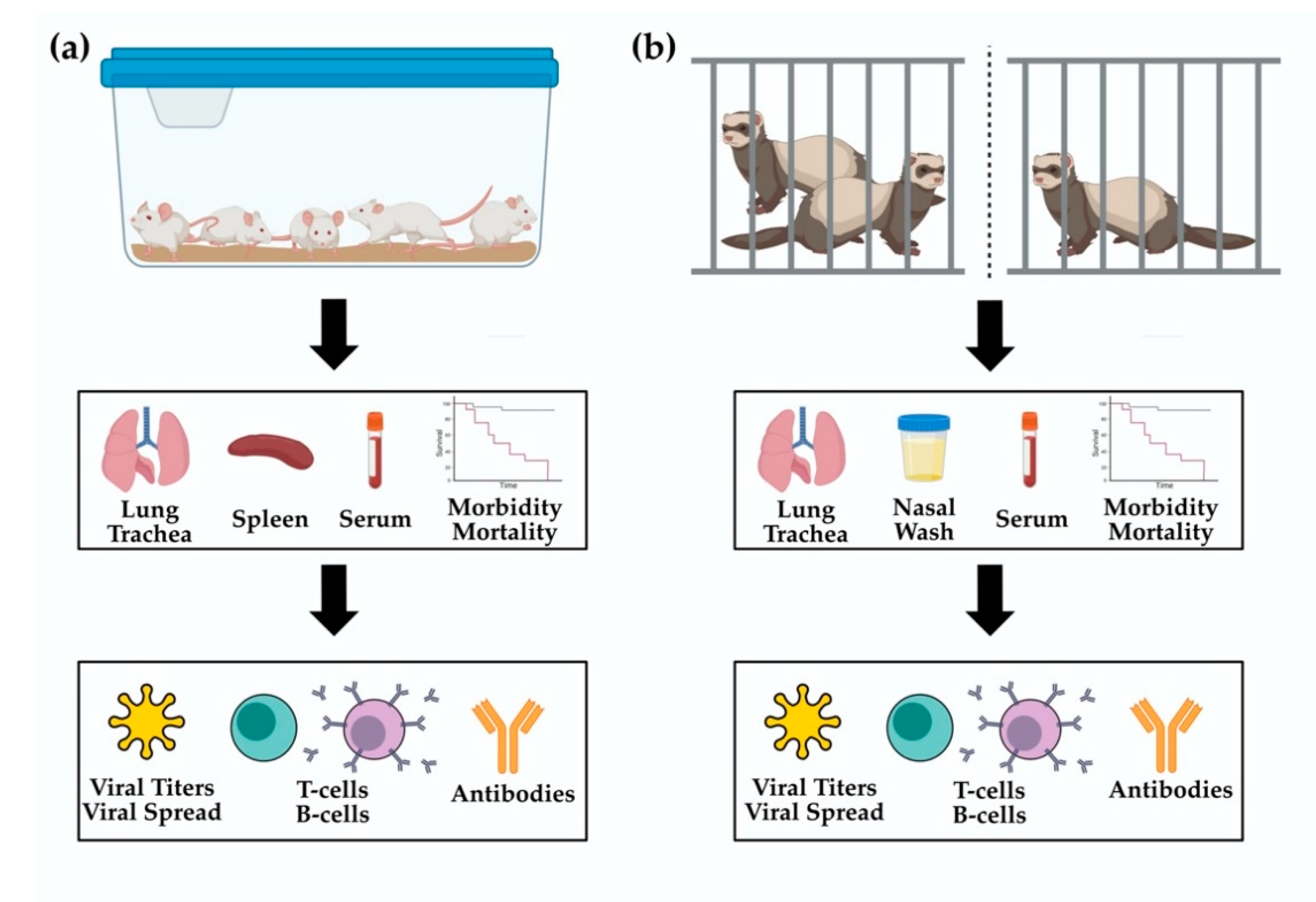
4.1. Mice
4.2. Ferrets
4.3. Guinea Pigs
4.4. Cotton Rats
4.5. Hamsters
4.6. Swine
4.7. Non-Human Primates
5. Pre-Clinical Animal Models of High-Risk Populations
5.1. Mice
5.2. Ferrets
5.3. Non-Human Primates
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Barberis, I.; Myles, P.; Ault, S.K.; Bragazzi, N.L.; Martini, M. History and evolution of influenza control through vaccination: From the first monovalent vaccine to universal vaccines. J. Prev. Med. Hyg. 2016, 57, E115–E120. [Google Scholar]
- Smith, W.; Andrewes, C.H.; Laidlaw, P.P. A virus obtained from influenza patients. Lancet 1933, 222, 66–68. [Google Scholar] [CrossRef]
- Francis, T. A new type of virus from epidemic influenza. Science 1940, 92, 405. [Google Scholar] [CrossRef]
- Flu Activity & Surveillance. Available online: https://www.cdc.gov/flu/weekly/fluactivitysurv.htm (accessed on 1 February 2021).
- Molinari, N.-A.M.; Ortega-Sanchez, I.R.; Messonnier, M.L.; Thompson, W.W.; Wortley, P.M.; Weintraub, E.; Bridges, C.B. The annual impact of seasonal influenza in the US: Measuring disease burden and costs. Vaccine 2007, 25, 5086–5096. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Palese, P. The biology of influenza viruses. Vaccine 2008, 26 (Suppl. 4), D49–D53. [Google Scholar] [CrossRef]
- Hay, A.J.; Gregory, V.; Douglas, A.R.; Lin, Y.P. The evolution of human influenza viruses. Philos. Trans. R Soc. Lond. B Biol. Sci. 2001, 356, 1861–1870. [Google Scholar] [CrossRef]
- Parrish, C.R.; Murcia, P.R.; Holmes, E.C. Influenza Virus Reservoirs and Intermediate Hosts: Dogs, Horses, and New Possibilities for Influenza Virus Exposure of Humans. J. Virol. 2015, 89, 2990. [Google Scholar] [CrossRef] [PubMed]
- Webster, R.G.; Yoon, S.; Webby, R.J. Evolution and ecology of influenza a viruses. Microbiol. Rev. 1992, 56, 152. [Google Scholar] [CrossRef] [PubMed]
- Reperant, L.A.; Kuiken, T.; Osterhaus, A.D.M.E. Influenza viruses. Hum. Vaccines Immunother. 2012, 8, 7–16. [Google Scholar] [CrossRef] [PubMed]
- Bodewes, R.; Morick, D.; de Mutsert, G.; Osinga, N.; Bestebroer, T.; van der Vliet, S.; Smits, S.L.; Kuiken, T.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; et al. Recurring influenza B virus infections in seals. Emerg. Infect. Dis. 2013, 19, 511–512. [Google Scholar] [CrossRef]
- Osterhaus, A.D.; Rimmelzwaan, G.F.; Martina, B.E.; Bestebroer, T.M.; Fouchier, R.A. Influenza B virus in seals. Science 2000, 288, 1051–1053. [Google Scholar] [CrossRef] [PubMed]
- Hulse-Post, D.J.; Sturm-Ramirez, K.M.; Humberd, J.; Seiler, P.; Govorkova, E.A.; Krauss, S.; Scholtissek, C.; Puthavathana, P.; Buranathai, C.; Nguyen, T.D.; et al. Role of domestic ducks in the propagation and biological evolution of highly pathogenic H5N1 influenza viruses in Asia. Proc. Natl. Acad. Sci. USA 2005, 102, 10682–10687. [Google Scholar] [CrossRef]
- Hui, D.S. Review of clinical symptoms and spectrum in humans with influenza A/H5N1 infection. Respirology 2008, 13 (Suppl. 1), S10–S13. [Google Scholar] [CrossRef] [PubMed]
- De Vries, R.D.; Herfst, S.; Richard, M. Avian Influenza A Virus Pandemic Preparedness and Vaccine Development. Vaccines 2018, 6, 46. [Google Scholar] [CrossRef] [PubMed]
- Hampson, A.W. Vaccines for pandemic influenza. The history of our current vaccines, their limitations and the requirements to deal with a pandemic threat. Ann. Acad. Med. Singap. 2008, 37, 510–517. [Google Scholar]
- Soema, P.C.; Kompier, R.; Amorij, J.P.; Kersten, G.F. Current and next generation influenza vaccines: Formulation and production strategies. Eur. J. Pharm. Biopharm. 2015, 94, 251–263. [Google Scholar] [CrossRef]
- Flannery, B.; Kondor, R.J.G.; Chung, J.R.; Gaglani, M.; Reis, M.; Zimmerman, R.K.; Nowalk, M.P.; Jackson, M.L.; Jackson, L.A.; Monto, A.S.; et al. Spread of Antigenically Drifted Influenza A(H3N2) Viruses and Vaccine Effectiveness in the United States During the 2018–2019 Season. J. Infect. Dis. 2019, 221, 8–15. [Google Scholar] [CrossRef]
- Francis, T.; Salk, J.E.; Pearson, H.E.; Brown, P.N. Protective effect of vaccination against induced influenza A. J. Clin. Investig. 1945, 24, 536–546. [Google Scholar] [CrossRef]
- Francis, T., Jr.; Salk, J.E.; Brace, W.M. The protective effect of vaccination against epidemic influenza B. J. Am. Med. Assoc. 1946, 131, 275–278. [Google Scholar] [CrossRef]
- Rota, P.A.; Wallis, T.R.; Harmon, M.W.; Rota, J.S.; Kendal, A.P.; Nerome, K. Cocirculation of two distinct evolutionary lineages of influenza type B virus since 1983. Virology 1990, 175, 59–68. [Google Scholar] [CrossRef]
- Ambrose, C.S.; Levin, M.J. The rationale for quadrivalent influenza vaccines. Hum. Vaccines Immunother. 2012, 8, 81–88. [Google Scholar] [CrossRef]
- Belshe, R.B. The need for quadrivalent vaccine against seasonal influenza. Vaccine 2010, 28, D45–D53. [Google Scholar] [CrossRef] [PubMed]
- Boyoglu-Barnum, S.; Ellis, D.; Gillespie, R.A.; Hutchinson, G.B.; Park, Y.-J.; Moin, S.M.; Acton, O.J.; Ravichandran, R.; Murphy, M.; Pettie, D.; et al. Quadrivalent influenza nanoparticle vaccines induce broad protection. Nature 2021, 592, 623–628. [Google Scholar] [CrossRef]
- Lee, B.Y.; Bartsch, S.M.; Willig, A.M. The economic value of a quadrivalent versus trivalent influenza vaccine. Vaccine 2012, 30, 7443–7446. [Google Scholar] [CrossRef]
- Gianchecchi, E.; Trombetta, C.; Piccirella, S.; Montomoli, E. Evaluating influenza vaccines: Progress and perspectives. Future Virol. 2016, 11, 379–393. [Google Scholar] [CrossRef]
- Influenza Virus Vaccine Safety & Availability. 18 September 2018. Available online: https://www.fda.gov/vaccines-blood-biologics/safety-availability-biologics/influenza-virus-vaccine-safety-availability (accessed on 13 July 2021).
- Weir, J.P.; Gruber, M.F. An overview of the regulation of influenza vaccines in the United States. Influenza Other Respir. Viruses 2016, 10, 354–360. [Google Scholar] [CrossRef]
- Schultz-Cherry, S.; Jones, J.C. Influenza vaccines: The good, the bad, and the eggs. Adv. Virus Res. 2010, 77, 63–84. [Google Scholar] [CrossRef] [PubMed]
- Stöhr, K.; Bucher, D.; Colgate, T.; Wood, J. Influenza virus surveillance, vaccine strain selection, and manufacture. Methods Mol. Biol. 2012, 865, 147–162. [Google Scholar] [CrossRef] [PubMed]
- Brauer, R.; Chen, P. Influenza virus propagation in embryonated chicken eggs. JoVE 2015, 97, e52421. [Google Scholar] [CrossRef]
- Zost, S.J.; Parkhouse, K.; Gumina, M.E.; Kim, K.; Diaz Perez, S.; Wilson, P.C.; Treanor, J.J.; Sant, A.J.; Cobey, S.; Hensley, S.E. Contemporary H3N2 influenza viruses have a glycosylation site that alters binding of antibodies elicited by egg-adapted vaccine strains. Proc. Natl. Acad. Sci. USA 2017, 114, 12578–12583. [Google Scholar] [CrossRef]
- Gouma, S.; Zost, S.J.; Parkhouse, K.; Branche, A.; Topham, D.J.; Cobey, S.; Hensley, S.E. Comparison of Human H3N2 Antibody Responses Elicited by Egg-Based, Cell-Based, and Recombinant Protein-Based Influenza Vaccines During the 2017–2018 Season. Clin. Infect. Dis. 2020, 71, 1447–1453. [Google Scholar] [CrossRef] [PubMed]
- van Riel, D.; Munster, V.J.; de Wit, E.; Rimmelzwaan, G.F.; Fouchier, R.A.M.; Osterhaus, A.D.M.E.; Kuiken, T. Human and avian influenza viruses target different cells in the lower respiratory tract of humans and other mammals. Am. J. Pathol. 2007, 171, 1215–1223. [Google Scholar] [CrossRef] [PubMed]
- Wong, S.S.; Webby, R.J. Traditional and new influenza vaccines. Clin. Microbiol. Rev. 2013, 26, 476–492. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Russell, R.F.; Kinnear, E. Adjuvanted influenza vaccines. Hum. Vaccines Immunother. 2018, 14, 550–564. [Google Scholar] [CrossRef]
- Ending the Pandemic Threat: A Grand Challenge for Universal Influenza Vaccine Development. Available online: https://gcgh.grandchallenges.org/challenge/ending-pandemic-threat-grand-challenge-universal-influenza-vaccine-development (accessed on 14 July 2021).
- WHO Preferred Product Characteristics for Next Generation Influenza Vaccines; World Health Organization: Geneva, Switzerland, 2017.
- Erbelding, E.J.; Post, D.J.; Stemmy, E.J.; Roberts, P.C.; Augustine, A.D.; Ferguson, S.; Paules, C.I.; Graham, B.S.; Fauci, A.S. A Universal Influenza Vaccine: The Strategic Plan for the National Institute of Allergy and Infectious Diseases. J. Infect. Dis. 2018, 218, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Hashem, A.M. Prospects of HA-based universal influenza vaccine. Biomed. Res. Int. 2015, 2015, 414637. [Google Scholar] [CrossRef]
- Henry, C.; Palm, A.E.; Krammer, F.; Wilson, P.C. From Original Antigenic Sin to the Universal Influenza Virus Vaccine. Trends Immunol. 2018, 39, 70–79. [Google Scholar] [CrossRef]
- Lee, Y.-T.; Kim, K.-H.; Ko, E.-J.; Lee, Y.-N.; Kim, M.-C.; Kwon, Y.-M.; Tang, Y.; Cho, M.-K.; Lee, Y.-J.; Kang, S.-M. New vaccines against influenza virus. Clin. Exp. Vaccine Res. 2014, 3, 12–28. [Google Scholar] [CrossRef]
- Neu, K.E.; Henry Dunand, C.J.; Wilson, P.C. Heads, stalks and everything else: How can antibodies eradicate influenza as a human disease? Curr. Opin. Immunol. 2016, 42, 48–55. [Google Scholar] [CrossRef]
- Ostrowsky, J.; Arpey, M.; Moore, K.; Osterholm, M.; Friede, M.; Gordon, J.; Higgins, D.; Molto-Lopez, J.; Seals, J.; Bresee, J. Tracking progress in universal influenza vaccine development. Curr. Opin. Virol. 2020, 40, 28–36. [Google Scholar] [CrossRef]
- Sangesland, M.; Lingwood, D. Antibody Focusing to Conserved Sites of Vulnerability: The Immunological Pathways for ‘Universal’ Influenza Vaccines. Vaccines 2021, 9, 125. [Google Scholar] [CrossRef] [PubMed]
- Sautto, G.A.; Kirchenbaum, G.A.; Ross, T.M. Towards a universal influenza vaccine: Different approaches for one goal. Virol. J. 2018, 15, 17. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Deng, L.; Kang, S.M.; Wang, B.Z. Universal influenza vaccines: From viruses to nanoparticles. Expert Rev. Vaccines 2018, 17, 967–976. [Google Scholar] [CrossRef] [PubMed]
- Hobson, D.; Curry, R.L.; Beare, A.S.; Ward-Gardner, A. The role of serum haemagglutination-inhibiting antibody in protection against challenge infection with influenza A2 and B viruses. Epidemiol. Infect. 1972, 70, 767–777. [Google Scholar] [CrossRef] [PubMed]
- Potter, C.W.; Oxford, J.S. Determinants of immunity to influenza infection in man. Br. Med. Bull. 1979, 35, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Ohmit, S.E.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Monto, A.S. Influenza Hemagglutination-Inhibition Antibody Titer as a Correlate of Vaccine-Induced Protection. J. Infect. Dis. 2011, 204, 1879–1885. [Google Scholar] [CrossRef]
- Chen, Y.Q.; Wohlbold, T.J.; Zheng, N.Y.; Huang, M.; Huang, Y.; Neu, K.E.; Lee, J.; Wan, H.; Rojas, K.T.; Kirkpatrick, E.; et al. Influenza Infection in Humans Induces Broadly Cross-Reactive and Protective Neuraminidase-Reactive Antibodies. Cell 2018, 173, 417–429.e410. [Google Scholar] [CrossRef]
- Monto, A.S.; Petrie, J.G.; Cross, R.T.; Johnson, E.; Liu, M.; Zhong, W.; Levine, M.; Katz, J.M.; Ohmit, S.E. Antibody to Influenza Virus Neuraminidase: An Independent Correlate of Protection. J. Infect. Dis. 2015, 212, 1191–1199. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Choi, A.; Hirsh, A.; Margine, I.; Iida, S.; Barrera, A.; Ferres, M.; Albrecht, R.A.; García-Sastre, A.; Bouvier, N.M.; et al. Defining the antibody cross-reactome directed against the influenza virus surface glycoproteins. Nat. Immunol. 2017, 18, 464–473. [Google Scholar] [CrossRef]
- Ng, S.; Nachbagauer, R.; Balmaseda, A.; Stadlbauer, D.; Ojeda, S.; Patel, M.; Rajabhathor, A.; Lopez, R.; Guglia, A.F.; Sanchez, N.; et al. Novel correlates of protection against pandemic H1N1 influenza A virus infection. Nat. Med. 2019, 25, 962–967. [Google Scholar] [CrossRef]
- Freyn, A.W.; Ramos da Silva, J.; Rosado, V.C.; Bliss, C.M.; Pine, M.; Mui, B.L.; Tam, Y.K.; Madden, T.D.; de Souza Ferreira, L.C.; Weissman, D.; et al. A Multi-Targeting, Nucleoside-Modified mRNA Influenza Virus Vaccine Provides Broad Protection in Mice. Mol. Ther. 2020, 28, 1569–1584. [Google Scholar] [CrossRef]
- Pardi, N.; Parkhouse, K.; Kirkpatrick, E.; McMahon, M.; Zost, S.J.; Mui, B.L.; Tam, Y.K.; Karikó, K.; Barbosa, C.J.; Madden, T.D.; et al. Nucleoside-modified mRNA immunization elicits influenza virus hemagglutinin stalk-specific antibodies. Nat. Commun. 2018, 9, 3361. [Google Scholar] [CrossRef]
- Liang, F.; Lindgren, G.; Lin, A.; Thompson, E.A.; Ols, S.; Röhss, J.; John, S.; Hassett, K.; Yuzhakov, O.; Bahl, K.; et al. Efficient Targeting and Activation of Antigen-Presenting Cells In Vivo after Modified mRNA Vaccine Administration in Rhesus Macaques. Mol. Ther. 2017, 25, 2635–2647. [Google Scholar] [CrossRef]
- Wong, S.S.; Webby, R.J. An mRNA vaccine for influenza. Nat. Biotechnol. 2012, 30, 1202–1204. [Google Scholar] [CrossRef]
- Krammer, F.; Pica, N.; Hai, R.; Margine, I.; Palese, P. Chimeric Hemagglutinin Influenza Virus Vaccine Constructs Elicit Broadly Protective Stalk-Specific Antibodies. J. Virol. 2013, 87, 6542. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Feser, J.; Naficy, A.; Bernstein, D.I.; Guptill, J.; Walter, E.B.; Berlanda-Scorza, F.; Stadlbauer, D.; Wilson, P.C.; Aydillo, T.; et al. A chimeric hemagglutinin-based universal influenza virus vaccine approach induces broad and long-lasting immunity in a randomized, placebo-controlled phase I trial. Nat. Med. 2021, 27, 106–114. [Google Scholar] [CrossRef] [PubMed]
- Margine, I.; Krammer, F.; Hai, R.; Heaton, N.S.; Tan, G.S.; Andrews, S.A.; Runstadler, J.A.; Wilson, P.C.; Albrecht, R.A.; García-Sastre, A.; et al. Hemagglutinin stalk-based universal vaccine constructs protect against group 2 influenza A viruses. J. Virol. 2013, 87, 10435–10446. [Google Scholar] [CrossRef] [PubMed]
- Ermler, M.E.; Kirkpatrick, E.; Sun, W.; Hai, R.; Amanat, F.; Chromikova, V.; Palese, P.; Krammer, F. Chimeric Hemagglutinin Constructs Induce Broad Protection against Influenza B Virus Challenge in the Mouse Model. J. Virol. 2017, 91, e00286-17. [Google Scholar] [CrossRef] [PubMed]
- Steel, J.; Lowen, A.C.; Wang, T.T.; Yondola, M.; Gao, Q.; Haye, K.; García-Sastre, A.; Palese, P. Influenza Virus Vaccine Based on the Conserved Hemagglutinin Stalk Domain. mBio 2010, 1, e00018-10. [Google Scholar] [CrossRef]
- van der Lubbe, J.E.M.; Verspuij, J.W.A.; Huizingh, J.; Schmit-Tillemans, S.P.R.; Tolboom, J.; Dekking, L.; Kwaks, T.; Brandenburg, B.; Meijberg, W.; Zahn, R.C.; et al. Mini-HA Is Superior to Full Length Hemagglutinin Immunization in Inducing Stem-Specific Antibodies and Protection Against Group 1 Influenza Virus Challenges in Mice. Front. Immunol. 2018, 9, 2350. [Google Scholar] [CrossRef]
- Sun, W.; Kirkpatrick, E.; Ermler, M.; Nachbagauer, R.; Broecker, F.; Krammer, F.; Palese, P. Development of Influenza B Universal Vaccine Candidates Using the “Mosaic” Hemagglutinin Approach. J. Virol. 2019, 93, e00333-19. [Google Scholar] [CrossRef] [PubMed]
- Broecker, F.; Liu, S.T.H.; Suntronwong, N.; Sun, W.; Bailey, M.J.; Nachbagauer, R.; Krammer, F.; Palese, P. A mosaic hemagglutinin-based influenza virus vaccine candidate protects mice from challenge with divergent H3N2 strains. NPJ Vaccines 2019, 4, 31. [Google Scholar] [CrossRef] [PubMed]
- Carter, D.M.; Darby, C.A.; Lefoley, B.C.; Crevar, C.J.; Alefantis, T.; Oomen, R.; Anderson, S.F.; Strugnell, T.; Cortés-Garcia, G.; Vogel, T.U.; et al. Design and Characterization of a Computationally Optimized Broadly Reactive Hemagglutinin Vaccine for H1N1 Influenza Viruses. J. Virol. 2016, 90, 4720. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Ross, T.M. Next generation methodology for updating HA vaccines against emerging human seasonal influenza A(H3N2) viruses. Sci. Rep. 2021, 11, 4554. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.D.; Ray, S.; Ross, T.M. Split inactivated COBRA vaccine elicits protective antibodies against H1N1 and H3N2 influenza viruses. PLoS ONE 2018, 13, e0204284. [Google Scholar] [CrossRef]
- Crevar, C.J.; Carter, D.M.; Lee, K.Y.; Ross, T.M. Cocktail of H5N1 COBRA HA vaccines elicit protective antibodies against H5N1 viruses from multiple clades. Hum. Vaccines Immunother. 2015, 11, 572–583. [Google Scholar] [CrossRef] [PubMed]
- Wohlbold, T.J.; Nachbagauer, R.; Xu, H.; Tan, G.S.; Hirsh, A.; Brokstad, K.A.; Cox, R.J.; Palese, P.; Krammer, F. Vaccination with Adjuvanted Recombinant Neuraminidase Induces Broad Heterologous, but Not Heterosubtypic, Cross-Protection against Influenza Virus Infection in Mice. mBio 2015, 6, e02556-14. [Google Scholar] [CrossRef]
- McMahon, M.; Kirkpatrick, E.; Stadlbauer, D.; Strohmeier, S.; Bouvier, N.M.; Krammer, F. Mucosal Immunity against Neuraminidase Prevents Influenza B Virus Transmission in Guinea Pigs. mBio 2019, 10, e00560-19. [Google Scholar] [CrossRef]
- Job, E.R.; Ysenbaert, T.; Smet, A.; Christopoulou, I.; Strugnell, T.; Oloo, E.O.; Oomen, R.P.; Kleanthous, H.; Vogel, T.U.; Saelens, X. Broadened immunity against influenza by vaccination with computationally designed influenza virus N1 neuraminidase constructs. NPJ Vaccines 2018, 3, 55. [Google Scholar] [CrossRef]
- Eichelberger, M.C.; Wan, H. Influenza neuraminidase as a vaccine antigen. Curr. Top. Microbiol. Immunol. 2015, 386, 275–299. [Google Scholar] [CrossRef] [PubMed]
- Fiers, W.; De Filette, M.; Bakkouri, K.E.; Schepens, B.; Roose, K.; Schotsaert, M.; Birkett, A.; Saelens, X. M2e-based universal influenza A vaccine. Vaccine 2009, 27, 6280–6283. [Google Scholar] [CrossRef]
- Saelens, X. The Role of Matrix Protein 2 Ectodomain in the Development of Universal Influenza Vaccines. J. Infect. Dis. 2019, 219, S68–S74. [Google Scholar] [CrossRef]
- Mezhenskaya, D.; Isakova-Sivak, I.; Rudenko, L. M2e-based universal influenza vaccines: A historical overview and new approaches to development. J. Biomed. Sci. 2019, 26, 76. [Google Scholar] [CrossRef] [PubMed]
- Watkins, H.C.; Rappazzo, C.G.; Higgins, J.S.; Sun, X.; Brock, N.; Chau, A.; Misra, A.; Cannizzo, J.P.B.; King, M.R.; Maines, T.R.; et al. Safe Recombinant Outer Membrane Vesicles that Display M2e Elicit Heterologous Influenza Protection. Mol. Ther. 2017, 25, 989–1002. [Google Scholar] [CrossRef] [PubMed]
- Coughlan, L.; Sridhar, S.; Payne, R.; Edmans, M.; Milicic, A.; Venkatraman, N.; Lugonja, B.; Clifton, L.; Qi, C.; Folegatti, P.M.; et al. Heterologous Two-Dose Vaccination with Simian Adenovirus and Poxvirus Vectors Elicits Long-Lasting Cellular Immunity to Influenza Virus A in Healthy Adults. EBioMedicine 2018, 29, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Antrobus, R.D.; Coughlan, L.; Berthoud, T.K.; Dicks, M.D.; Hill, A.V.; Lambe, T.; Gilbert, S.C. Clinical assessment of a novel recombinant simian adenovirus ChAdOx1 as a vectored vaccine expressing conserved Influenza A antigens. Mol. Ther. 2014, 22, 668–674. [Google Scholar] [CrossRef] [PubMed]
- Pillet, S.; Aubin, É.; Trépanier, S.; Bussière, D.; Dargis, M.; Poulin, J.-F.; Yassine-Diab, B.; Ward, B.J.; Landry, N. A plant-derived quadrivalent virus like particle influenza vaccine induces cross-reactive antibody and T cell response in healthy adults. Clin. Immunol. 2016, 168, 72–87. [Google Scholar] [CrossRef]
- Albrecht, R.A.; Liu, W.-C.; Sant, A.J.; Tompkins, S.M.; Pekosz, A.; Meliopoulos, V.; Cherry, S.; Thomas, P.G.; Schultz-Cherry, S. Moving Forward: Recent Developments for the Ferret Biomedical Research Model. mBio 2018, 9, e01113-18. [Google Scholar] [CrossRef]
- Belser, J.A.; Barclay, W.; Barr, I.; Fouchier, R.A.M.; Matsuyama, R.; Nishiura, H.; Peiris, M.; Russell, C.J.; Subbarao, K.; Zhu, H.; et al. Ferrets as Models for Influenza Virus Transmission Studies and Pandemic Risk Assessments. Emerg. Infect. Dis. 2018, 24, 965–971. [Google Scholar] [CrossRef]
- Belser, J.A.; Eckert, A.M.; Huynh, T.; Gary, J.M.; Ritter, J.M.; Tumpey, T.M.; Maines, T.R. A Guide for the Use of the Ferret Model for Influenza Virus Infection. Am. J. Pathol. 2020, 190, 11–24. [Google Scholar] [CrossRef]
- Belser, J.A.; Eckert, A.M.; Tumpey, T.M.; Maines, T.R. Complexities in Ferret Influenza Virus Pathogenesis and Transmission Models. Microbiol. Mol. Biol. Rev. 2016, 80, 733–744. [Google Scholar] [CrossRef]
- Belser, J.A.; Katz, J.M.; Tumpey, T.M. The ferret as a model organism to study influenza A virus infection. Dis. Models Mech. 2011, 4, 575–579. [Google Scholar] [CrossRef]
- Berendt, R.F. Simian model for the evaluation of immunity to influenza. Infect. Immun. 1974, 9, 101–105. [Google Scholar] [CrossRef]
- Bodewes, R.; Rimmelzwaan, G.F.; Osterhaus, A.D. Animal models for the preclinical evaluation of candidate influenza vaccines. Expert Rev. Vaccines 2010, 9, 59–72. [Google Scholar] [CrossRef] [PubMed]
- Boukhvalova, M.S.; Prince, G.A.; Blanco, J.C. The cotton rat model of respiratory viral infections. Biologicals 2009, 37, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Eichelberger, M.C. The cotton rat as a model to study influenza pathogenesis and immunity. Viral Immunol. 2007, 20, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Eichelberger, M.C.; Green, M.D. Animal models to assess the toxicity, immunogenicity and effectiveness of candidate influenza vaccines. Expert Opin. Drug Metab. Toxicol. 2011, 7, 1117–1127. [Google Scholar] [CrossRef]
- Fullen, D.J.; Noulin, N.; Catchpole, A.; Fathi, H.; Murray, E.J.; Mann, A.; Eze, K.; Balaratnam, G.; Borley, D.W.; Gilbert, A.; et al. Accelerating Influenza Research: Vaccines, Antivirals, Immunomodulators and Monoclonal Antibodies. The Manufacture of a New Wild-Type H3N2 Virus for the Human Viral Challenge Model. PLoS ONE 2016, 11, e0145902. [Google Scholar] [CrossRef]
- Groves, H.T.; McDonald, J.U.; Langat, P.; Kinnear, E.; Kellam, P.; McCauley, J.; Ellis, J.; Thompson, C.; Elderfield, R.; Parker, L.; et al. Mouse Models of Influenza Infection with Circulating Strains to Test Seasonal Vaccine Efficacy. Front. Immunol. 2018, 9, 126. [Google Scholar] [CrossRef] [PubMed]
- Honce, R.; Wohlgemuth, N.; Meliopoulos, V.A.; Short, K.R.; Schultz-Cherry, S. Influenza in High-Risk Hosts-Lessons Learned from Animal Models. Cold Spring Harb. Perspect. Med. 2020, 10. [Google Scholar] [CrossRef]
- Iwatsuki-Horimoto, K.; Nakajima, N.; Shibata, M.; Takahashi, K.; Sato, Y.; Kiso, M.; Yamayoshi, S.; Ito, M.; Enya, S.; Otake, M.; et al. The Microminipig as an Animal Model for Influenza A Virus Infection. J. Virol. 2017, 91. [Google Scholar] [CrossRef]
- Kollmus, H.; Pilzner, C.; Leist, S.R.; Heise, M.; Geffers, R.; Schughart, K. Of mice and men: The host response to influenza virus infection. Mamm. Genome 2018, 29, 446–470. [Google Scholar] [CrossRef]
- Lowen, A.C.; Bouvier, N.M.; Steel, J. Transmission in the guinea pig model. Curr. Top. Microbiol. Immunol. 2014, 385, 157–183. [Google Scholar] [CrossRef]
- Lutz, T.A.; Woods, S.C. Overview of animal models of obesity. Curr. Protoc. Pharmacol. 2012, 5, 61. [Google Scholar] [CrossRef] [PubMed]
- Maher, J.A.; DeStefano, J. The Ferret: An Animal Model to Study Influenza Virus. Lab. Animal 2004, 33, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Ottolini, M.G.; Blanco, J.C.G.; Eichelberger, M.C.; Porter, D.D.; Pletneva, L.; Richardson, J.Y.; Prince, G.A. The cotton rat provides a useful small-animal model for the study of influenza virus pathogenesis. J. Gen. Virol. 2005, 86, 2823–2830. [Google Scholar] [CrossRef] [PubMed]
- Rajao, D.S.; Vincent, A.L. Swine as a model for influenza A virus infection and immunity. Ilar. J. 2015, 56, 44–52. [Google Scholar] [CrossRef]
- Rimmelzwaan, G.F.; Baars, M.; van Beek, R.; van Amerongen, G.; Lövgren-Bengtsson, K.; Claas, E.C.; Osterhaus, A.D. Induction of protective immunity against influenza virus in a macaque model: Comparison of conventional and iscom vaccines. J. Gen. Virol. 1997, 78 Pt 4, 757–765. [Google Scholar] [CrossRef]
- Sun, Y.; Bi, Y.; Pu, J.; Hu, Y.; Wang, J.; Gao, H.; Liu, L.; Xu, Q.; Tan, Y.; Liu, M.; et al. Guinea pig model for evaluating the potential public health risk of swine and avian influenza viruses. PLoS ONE 2010, 5, e15537. [Google Scholar] [CrossRef]
- Taylor, R.M. Experimental infection with influenza a virus in mice: The increase in intrapulmonary virus after inoculation and the influence of various factors thereon. J. Exp. Med. 1941, 73, 43–55. [Google Scholar] [CrossRef]
- Francis, T. Quantitative relationships between the immunizing dose of epidemic influenza virus and the resultant immunity. J. Exp. Med. 1939, 69, 283–300. [Google Scholar] [CrossRef]
- Pearce, M.B.; Belser, J.A.; Houser, K.V.; Katz, J.M.; Tumpey, T.M. Efficacy of seasonal live attenuated influenza vaccine against virus replication and transmission of a pandemic 2009 H1N1 virus in ferrets. Vaccine 2011, 29, 2887–2894. [Google Scholar] [CrossRef] [PubMed]
- Yim, K.; Miles, B.; Zinsou, R.; Prince, G.; Boukhvalova, M. Efficacy of trivalent inactivated influenza vaccines in the cotton rat Sigmodon hispidus model. Vaccine 2012, 30, 1291–1296. [Google Scholar] [CrossRef] [PubMed]
- Hamouda, T.; Sutcliffe, J.A.; Ciotti, S.; Baker, J.R. Intranasal Immunization of Ferrets with Commercial Trivalent Influenza Vaccines Formulated in a Nanoemulsion-Based Adjuvant. Clin. Vaccine Immunol. 2011, 18, 1167. [Google Scholar] [CrossRef]
- Ellebedy, A.H.; Ducatez, M.F.; Duan, S.; Stigger-Rosser, E.; Rubrum, A.M.; Govorkova, E.A.; Webster, R.G.; Webby, R.J. Impact of prior seasonal influenza vaccination and infection on pandemic A (H1N1) influenza virus replication in ferrets. Vaccine 2011, 29, 3335–3339. [Google Scholar] [CrossRef]
- Iwasaki, A.; Pillai, P.S. Innate immunity to influenza virus infection. Nat. Rev. Immunol. 2014, 14, 315–328. [Google Scholar] [CrossRef] [PubMed]
- L’Huillier, A.G.; Ferreira, V.H.; Hirzel, C.; Nellimarla, S.; Ku, T.; Natori, Y.; Humar, A.; Kumar, D. T-cell responses following Natural Influenza Infection or Vaccination in Solid Organ Transplant Recipients. Sci. Rep. 2020, 10, 10104. [Google Scholar] [CrossRef]
- Ning, Z.Y.; Luo, M.Y.; Qi, W.B.; Yu, B.; Jiao, P.R.; Liao, M. Detection of expression of influenza virus receptors in tissues of BALB/c mice by histochemistry. Vet. Res. Commun. 2009, 33, 895–903. [Google Scholar] [CrossRef]
- Jia, N.; Barclay, W.S.; Roberts, K.; Yen, H.-L.; Chan, R.W.Y.; Lam, A.K.Y.; Air, G.; Peiris, J.S.M.; Dell, A.; Nicholls, J.M.; et al. Glycomic characterization of respiratory tract tissues of ferrets: Implications for its use in influenza virus infection studies. J. Biol. Chem. 2014, 289, 28489–28504. [Google Scholar] [CrossRef]
- Clemens, E.B.; van de Sandt, C.; Wong, S.S.; Wakim, L.M.; Valkenburg, S.A. Harnessing the Power of T Cells: The Promising Hope for a Universal Influenza Vaccine. Vaccines 2018, 6, 18. [Google Scholar] [CrossRef]
- Krammer, F. The human antibody response to influenza A virus infection and vaccination. Nat. Rev. Immunol. 2019, 19, 383–397. [Google Scholar] [CrossRef]
- Chen, X.; Liu, S.; Goraya, M.U.; Maarouf, M.; Huang, S.; Chen, J.-L. Host Immune Response to Influenza A Virus Infection. Front. Immunol. 2018, 9, 320. [Google Scholar] [CrossRef] [PubMed]
- Hannoun, C. The evolving history of influenza viruses and influenza vaccines. Expert Rev. Vaccines 2013, 12, 1085–1094. [Google Scholar] [CrossRef] [PubMed]
- Francis, T., Jr.; Pearson, H.E.; Sullivan, E.R.; Brown, P.M. The Effect of Subcutaneous Vaccination with Influenza Virus upon the Virus.Inactivating Capacity of Nasal Secretions. Am. J. Hyg. 1943, 37, 294–300. [Google Scholar]
- Francis, T., Jr. Vaccination against influenza. Bull. World Health Organ. 1953, 8, 725–741. [Google Scholar]
- Margine, I.; Krammer, F. Animal models for influenza viruses: Implications for universal vaccine development. Pathogens 2014, 3, 845–874. [Google Scholar] [CrossRef]
- Xu, L.; Bao, L.; Li, F.; Lv, Q.; Ma, Y.; Zhou, J.; Xu, Y.; Deng, W.; Zhan, L.; Zhu, H.; et al. Adaption of seasonal H1N1 influenza virus in mice. PLoS ONE 2011, 6, e28901. [Google Scholar] [CrossRef]
- Hu, Z.; Zhang, Y.; Wang, Z.; Wang, J.; Tong, Q.; Wang, M.; Sun, H.; Pu, J.; Liu, C.; Liu, J.; et al. Mouse-adapted H9N2 avian influenza virus causes systemic infection in mice. Virol. J. 2019, 16, 135. [Google Scholar] [CrossRef]
- Narasaraju, T.; Sim, M.K.; Ng, H.H.; Phoon, M.C.; Shanker, N.; Lal, S.K.; Chow, V.T. Adaptation of human influenza H3N2 virus in a mouse pneumonitis model: Insights into viral virulence, tissue tropism and host pathogenesis. Microbes. Infect. 2009, 11, 2–11. [Google Scholar] [CrossRef]
- Prokopyeva, E.; Kurskaya, O.; Sobolev, I.; Solomatina, M.; Murashkina, T.; Suvorova, A.; Alekseev, A.; Danilenko, D.; Komissarov, A.; Fadeev, A.; et al. Experimental Infection Using Mouse-Adapted Influenza B Virus in a Mouse Model. Viruses 2020, 12, 470. [Google Scholar] [CrossRef]
- Prokopyeva, E.A.; Sobolev, I.A.; Prokopyev, M.V.; Shestopalov, A.M. Adaptation of influenza A(H1N1)pdm09 virus in experimental mouse models. Infect. Genet. Evol. 2016, 39, 265–271. [Google Scholar] [CrossRef]
- Baz, M.; M’Hamdi, Z.; Carbonneau, J.; Lavigne, S.; Couture, C.; Abed, Y.; Boivin, G. Synergistic PA and HA mutations confer mouse adaptation of a contemporary A/H3N2 influenza virus. Sci. Rep. 2019, 9, 16616. [Google Scholar] [CrossRef]
- Qin, J.; Peng, O.; Shen, X.; Gong, L.; Xue, C.; Cao, Y. Multiple amino acid substitutions involved in the adaption of three avian-origin H7N9 influenza viruses in mice. Virol. J. 2019, 16, 3. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Zhang, H.; Yang, K.; Liang, W.; Xiong, Z.; Liu, Z.; Yang, X.; Shao, H.; Zheng, X.; Chen, M.; et al. Multiple amino acid substitutions are involved in the adaptation of H9N2 avian influenza virus to mice. Vet. Microbiol. 2009, 138, 85–91. [Google Scholar] [CrossRef]
- Matsuoka, Y.; Lamirande, E.W.; Subbarao, K. The Mouse Model for Influenza. Curr. Protoc. Microbiol. 2009, 13, 15G.3.1–15G.3.30. [Google Scholar] [CrossRef] [PubMed]
- Mestas, J.; Hughes, C.C. Of mice and not men: Differences between mouse and human immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [PubMed]
- Brehm, M.A.; Shultz, L.D.; Greiner, D.L. Humanized mouse models to study human diseases. Curr. Opin. Endocrinol. Diabetes Obes. 2010, 17, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, E.; Momose, H.; Hiradate, Y.; Furuhata, K.; Mizukami, T.; Hamaguchi, I. Development of a preclinical humanized mouse model to evaluate acute toxicity of an influenza vaccine. Oncotarget 2018, 9, 25751–25763. [Google Scholar] [CrossRef] [PubMed]
- Yu, C.I.; Gallegos, M.; Marches, F.; Zurawski, G.; Ramilo, O.; García-Sastre, A.; Banchereau, J.; Palucka, A.K. Broad influenza-specific CD8+ T-cell responses in humanized mice vaccinated with influenza virus vaccines. Blood 2008, 112, 3671–3678. [Google Scholar] [CrossRef]
- Dengler, L.; May, M.; Wilk, E.; Bahgat, M.M.; Schughart, K. Immunization with live virus vaccine protects highly susceptible DBA/2J mice from lethal influenza A H1N1 infection. Virol. J. 2012, 9, 212. [Google Scholar] [CrossRef]
- Pica, N.; Iyer, A.; Ramos, I.; Bouvier, N.M.; Fernandez-Sesma, A.; García-Sastre, A.; Lowen, A.C.; Palese, P.; Steel, J. The DBA.2 mouse is susceptible to disease following infection with a broad, but limited, range of influenza A and B viruses. J. Virol. 2011, 85, 12825–12829. [Google Scholar] [CrossRef]
- Jeong, J.H.; Kim, E.H.; Lloren, K.K.S.; Kwon, J.J.; Kwon, H.I.; Ahn, S.J.; Kim, Y.I.; Choi, W.S.; Si, Y.J.; Lee, O.J.; et al. Preclinical evaluation of the efficacy of an H5N8 vaccine candidate (IDCDC-RG43A) in mouse and ferret models for pandemic preparedness. Vaccine 2019, 37, 484–493. [Google Scholar] [CrossRef]
- Jiang, W.; Wong, J.; Tan, H.X.; Kelly, H.G.; Whitney, P.G.; Barr, I.; Layton, D.S.; Kent, S.J.; Wheatley, A.K.; Juno, J.A. Screening and development of monoclonal antibodies for identification of ferret T follicular helper cells. Sci. Rep. 2021, 11, 1864. [Google Scholar] [CrossRef]
- Jayaraman, A.; Chandrasekaran, A.; Viswanathan, K.; Raman, R.; Fox, J.G.; Sasisekharan, R. Decoding the distribution of glycan receptors for human-adapted influenza A viruses in ferret respiratory tract. PLoS ONE 2012, 7, e27517. [Google Scholar] [CrossRef]
- Cherukuri, A.; Servat, E.; Woo, J. Vaccine-specific antibody secreting cells are a robust early marker of LAIV-induced B-cell response in ferrets. Vaccine 2012, 30, 237–246. [Google Scholar] [CrossRef]
- Cox, R.J.; Major, D.; Hauge, S.; Madhun, A.S.; Brokstad, K.A.; Kuhne, M.; Smith, J.; Vogel, F.R.; Zambon, M.; Haaheim, L.R.; et al. A cell-based H7N1 split influenza virion vaccine confers protection in mouse and ferret challenge models. Influenza Other Respir. Viruses 2009, 3, 107–117. [Google Scholar] [CrossRef] [PubMed]
- Fan, J.; Liang, X.; Horton, M.S.; Perry, H.C.; Citron, M.P.; Heidecker, G.J.; Fu, T.M.; Joyce, J.; Przysiecki, C.T.; Keller, P.M.; et al. Preclinical study of influenza virus A M2 peptide conjugate vaccines in mice, ferrets, and rhesus monkeys. Vaccine 2004, 22, 2993–3003. [Google Scholar] [CrossRef] [PubMed]
- Huber, V.C.; McCullers, J.A. Live attenuated influenza vaccine is safe and immunogenic in immunocompromised ferrets. J. Infect. Dis. 2006, 193, 677–684. [Google Scholar] [CrossRef] [PubMed]
- Isakova-Sivak, I.; Matyushenko, V.; Kotomina, T.; Kiseleva, I.; Krutikova, E.; Donina, S.; Rekstin, A.; Larionova, N.; Mezhenskaya, D.; Sivak, K.; et al. Sequential Immunization with Universal Live Attenuated Influenza Vaccine Candidates Protects Ferrets against a High-Dose Heterologous Virus Challenge. Vaccines 2019, 7, 61. [Google Scholar] [CrossRef] [PubMed]
- Krammer, F.; Hai, R.; Yondola, M.; Tan, G.S.; Leyva-Grado, V.H.; Ryder, A.B.; Miller, M.S.; Rose, J.K.; Palese, P.; García-Sastre, A.; et al. Assessment of influenza virus hemagglutinin stalk-based immunity in ferrets. J. Virol. 2014, 88, 3432–3442. [Google Scholar] [CrossRef]
- Liu, W.C.; Nachbagauer, R.; Stadlbauer, D.; Solórzano, A.; Berlanda-Scorza, F.; García-Sastre, A.; Palese, P.; Krammer, F.; Albrecht, R.A. Sequential Immunization With Live-Attenuated Chimeric Hemagglutinin-Based Vaccines Confers Heterosubtypic Immunity Against Influenza A Viruses in a Preclinical Ferret Model. Front. Immunol. 2019, 10, 756. [Google Scholar] [CrossRef]
- Liu, W.C.; Nachbagauer, R.; Stadlbauer, D.; Strohmeier, S.; Solórzano, A.; Berlanda-Scorza, F.; Innis, B.L.; García-Sastre, A.; Palese, P.; Krammer, F.; et al. Chimeric Hemagglutinin-Based Live-Attenuated Vaccines Confer Durable Protective Immunity against Influenza A Viruses in a Preclinical Ferret Model. Vaccines 2021, 9, 40. [Google Scholar] [CrossRef] [PubMed]
- Ljungberg, K.; Kolmskog, C.; Wahren, B.; van Amerongen, G.; Baars, M.; Osterhaus, A.; Linde, A.; Rimmelzwaan, G. DNA vaccination of ferrets with chimeric influenza A virus hemagglutinin (H3) genes. Vaccine 2002, 20, 2045–2052. [Google Scholar] [CrossRef]
- Marriott, A.C.; Dove, B.K.; Whittaker, C.J.; Bruce, C.; Ryan, K.A.; Bean, T.J.; Rayner, E.; Pearson, G.; Taylor, I.; Dowall, S.; et al. Low dose influenza virus challenge in the ferret leads to increased virus shedding and greater sensitivity to oseltamivir. PLoS ONE 2014, 9, e94090. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.; Asthagiri Arunkumar, G.; Liu, W.C.; Stadlbauer, D.; Albrecht, R.A.; Pavot, V.; Aramouni, M.; Lambe, T.; Gilbert, S.C.; Krammer, F. Vaccination With Viral Vectors Expressing Chimeric Hemagglutinin, NP and M1 Antigens Protects Ferrets Against Influenza Virus Challenge. Front. Immunol. 2019, 10, 2005. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Krammer, F.; Albrecht, R.A. A Live-Attenuated Prime, Inactivated Boost Vaccination Strategy with Chimeric Hemagglutinin-Based Universal Influenza Virus Vaccines Provides Protection in Ferrets: A Confirmatory Study. Vaccines 2018, 6, 47. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Liu, W.C.; Choi, A.; Wohlbold, T.J.; Atlas, T.; Rajendran, M.; Solórzano, A.; Berlanda-Scorza, F.; García-Sastre, A.; Palese, P.; et al. A universal influenza virus vaccine candidate confers protection against pandemic H1N1 infection in preclinical ferret studies. NPJ Vaccines 2017, 2, 26. [Google Scholar] [CrossRef]
- Nachbagauer, R.; Miller, M.S.; Hai, R.; Ryder, A.B.; Rose, J.K.; Palese, P.; García-Sastre, A.; Krammer, F.; Albrecht, R.A. Hemagglutinin Stalk Immunity Reduces Influenza Virus Replication and Transmission in Ferrets. J. Virol. 2015, 90, 3268–3273. [Google Scholar] [CrossRef]
- Ou, H.; Yao, W.; Wu, N.; Wang, F.X.; Weng, T.; Han, C.; Lu, X.; Yu, D.; Wu, H.; Cheng, L.; et al. Preclinical evaluation of the safety and pathogenicity of a live attenuated recombinant influenza A/H7N9 seed strain and corresponding MF59-adjuvanted split vaccine. Oncotarget 2016, 7, 81012–81025. [Google Scholar] [CrossRef][Green Version]
- Romanova, J.; Krenn, B.M.; Wolschek, M.; Ferko, B.; Romanovskaja-Romanko, E.; Morokutti, A.; Shurygina, A.P.; Nakowitsch, S.; Ruthsatz, T.; Kiefmann, B.; et al. Preclinical evaluation of a replication-deficient intranasal DeltaNS1 H5N1 influenza vaccine. PLoS ONE 2009, 4, e5984. [Google Scholar] [CrossRef]
- Vidaña, B.; Brookes, S.M.; Everett, H.E.; Garcon, F.; Nuñez, A.; Engelhardt, O.; Major, D.; Hoschler, K.; Brown, I.H.; Zambon, M. Inactivated pandemic 2009 H1N1 influenza A virus human vaccines have different efficacy after homologous challenge in the ferret model. Influenza Other Respir. Viruses 2021, 15, 142–153. [Google Scholar] [CrossRef]
- Bushnell, R.V.; Tobin, J.K.; Long, J.; Schultz-Cherry, S.; Chaudhuri, A.R.; Nara, P.L.; Tobin, G.J. Serological characterization of guinea pigs infected with H3N2 human influenza or immunized with hemagglutinin protein. Virol. J. 2010, 7, 200. [Google Scholar] [CrossRef]
- Gustin, K.M.; Belser, J.A.; Veguilla, V.; Zeng, H.; Katz, J.M.; Tumpey, T.M.; Maines, T.R. Environmental Conditions Affect Exhalation of H3N2 Seasonal and Variant Influenza Viruses and Respiratory Droplet Transmission in Ferrets. PLoS ONE 2015, 10, e0125874. [Google Scholar] [CrossRef]
- O’Donnell, C.D.; Wright, A.; Vogel, L.; Boonnak, K.; Treanor, J.J.; Subbarao, K. Humans and ferrets with prior H1N1 influenza virus infections do not exhibit evidence of original antigenic sin after infection or vaccination with the 2009 pandemic H1N1 influenza virus. Clin. Vaccine Immunol. 2014, 21, 737–746. [Google Scholar] [CrossRef] [PubMed]
- Rutigliano, J.A.; Doherty, P.C.; Franks, J.; Morris, M.Y.; Reynolds, C.; Thomas, P.G. Screening monoclonal antibodies for cross-reactivity in the ferret model of influenza infection. J. Immunol. Methods 2008, 336, 71–77. [Google Scholar] [CrossRef] [PubMed]
- Peng, X.; Alföldi, J.; Gori, K.; Eisfeld, A.J.; Tyler, S.R.; Tisoncik-Go, J.; Brawand, D.; Law, G.L.; Skunca, N.; Hatta, M.; et al. The draft genome sequence of the ferret (Mustela putorius furo) facilitates study of human respiratory disease. Nat. Biotechnol. 2014, 32, 1250–1255. [Google Scholar] [CrossRef] [PubMed]
- Nakata, M.; Itou, T.; Sakai, T. Molecular cloning and phylogenetic analysis of inflammatory cytokines of the ferret (Mustela putorius furo). J. Vet. Med. Sci. 2008, 70, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Ochi, A.; Danesh, A.; Seneviratne, C.; Banner, D.; Devries, M.E.; Rowe, T.; Xu, L.; Ran, L.; Czub, M.; Bosinger, S.E.; et al. Cloning, expression and immunoassay detection of ferret IFN-gamma. Dev. Comp. Immunol. 2008, 32, 890–897. [Google Scholar] [CrossRef]
- Danesh, A.; Seneviratne, C.; Cameron, C.M.; Banner, D.; Devries, M.E.; Kelvin, A.A.; Xu, L.; Ran, L.; Bosinger, S.E.; Rowe, T.; et al. Cloning, expression and characterization of ferret CXCL10. Mol. Immunol. 2008, 45, 1288–1297. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.J.; Teh, C.Z.; Jennings, R.; Potter, C.W. Transmissibility of influenza viruses in hamsters. Arch. Virol. 1982, 72, 187–197. [Google Scholar] [CrossRef]
- Bouvier, N.M.; Lowen, A.C. Animal Models for Influenza Virus Pathogenesis and Transmission. Viruses 2010, 2, 1530–1563. [Google Scholar] [CrossRef]
- DiPiazza, A.; Richards, K.; Batarse, F.; Lockard, L.; Zeng, H.; García-Sastre, A.; Albrecht, R.A.; Sant, A.J. Flow Cytometric and Cytokine ELISpot Approaches To Characterize the Cell-Mediated Immune Response in Ferrets following Influenza Virus Infection. J. Virol. 2016, 90, 7991. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Miao, J.; Fan, Z.; Song, S.; Kong, I.K.; Wang, Y.; Wang, Z. Production of Genetically Engineered Golden Syrian Hamsters by Pronuclear Injection of the CRISPR/Cas9 Complex. J. Vis. Exp. 2018, 56263. [Google Scholar] [CrossRef] [PubMed]
- Miao, J.; Chard, L.S.; Wang, Z.; Wang, Y. Syrian Hamster as an Animal Model for the Study on Infectious Diseases. Front. Immunol. 2019, 10, 2329. [Google Scholar] [CrossRef] [PubMed]
- Lange, E.; Kalthoff, D.; Blohm, U.; Teifke, J.P.; Breithaupt, A.; Maresch, C.; Starick, E.; Fereidouni, S.; Hoffmann, B.; Mettenleiter, T.C.; et al. Pathogenesis and transmission of the novel swine-origin influenza virus A/H1N1 after experimental infection of pigs. J. Gen. Virol. 2009, 90, 2119–2123. [Google Scholar] [CrossRef] [PubMed]
- Bhide, Y.; Dong, W.; Meijerhof, T.; de Vries-Idema, J.; Niesters, H.G.; Huckriede, A. Characterization of humoral immune responses and degree of protection induced by influenza vaccine in cotton rats: Effects of low vaccine dose and single vs booster vaccination. Immun. Inflamm. Dis. 2020, 8, 279–291. [Google Scholar] [CrossRef]
- Straight, T.M.; Ottolini, M.G.; Prince, G.A.; Eichelberger, M.C. Evidence of a cross-protective immune response to influenza A in the cotton rat model. Vaccine 2006, 24, 6264–6271. [Google Scholar] [CrossRef]
- Potter, C.W.; Jennings, R.; Rees, R.C.; McLaren, C. Antibody Response of Hamsters to A2/Hong Kong Virus Vaccine after Priming by Heterotypic Virus Infection. Infect. Immun. 1973, 8, 137. [Google Scholar] [CrossRef]
- Smith, G.L.; Murphy, B.R.; Moss, B. Construction and characterization of an infectious vaccinia virus recombinant that expresses the influenza hemagglutinin gene and induces resistance to influenza virus infection in hamsters. Proc. Natl. Acad. Sci. USA 1983, 80, 7155. [Google Scholar] [CrossRef]
- Jennings, R.; Potter, C.W. Enhanced response to influenza A vaccines in hamsters primed by prior heterotype influenza infection. Arch. Gesamte Virusforsch 1973, 42, 197–206. [Google Scholar] [CrossRef]
- Jennings, R.; Brand, C.M.; McLaren, C.; Shepherd, L.; Potter, C.W. The immune response of hamsters to purified haemagglutinins and whole influenza virus vaccines following live influenza virus infection. Med. Microbiol. Immunol. 1974, 160, 295–309. [Google Scholar] [CrossRef]
- Brundage-Anguish, L.J.; Holmes, D.F.; Hosier, N.T.; Murphy, B.R.; Massicott, J.G.; Appleyard, G.; Coggins, L. Live temperature-sensitive equine influenza virus vaccine: Generation of the virus and efficacy in hamsters. Am. J. Vet. Res. 1982, 43, 869–874. [Google Scholar]
- Jennings, R.; Potter, C.W.; McLaren, C.; Brady, M. A new, surface-antigen-adsorbed influenza virus vaccine. I. Studies on immunogenicity in hamsters. J. Hyg. 1975, 75, 341–352. [Google Scholar] [CrossRef] [PubMed]
- Nelson, M.I.; Worobey, M. Origins of the 1918 Pandemic: Revisiting the Swine "Mixing Vessel" Hypothesis. Am. J. Epidemiol. 2018, 187, 2498–2502. [Google Scholar] [CrossRef]
- Mancera Gracia, J.C.; Pearce, D.S.; Masic, A.; Balasch, M. Influenza A Virus in Swine: Epidemiology, Challenges and Vaccination Strategies. Front. Vet. Sci. 2020, 7, 647. [Google Scholar] [CrossRef]
- Vincent, A.L.; Lager, K.M.; Janke, B.H.; Gramer, M.R.; Richt, J.A. Failure of protection and enhanced pneumonia with a US H1N2 swine influenza virus in pigs vaccinated with an inactivated classical swine H1N1 vaccine. Vet. Microbiol. 2008, 126, 310–323. [Google Scholar] [CrossRef] [PubMed]
- Broadbent, A.J.; Santos, C.P.; Paskel, M.; Matsuoka, Y.; Lu, J.; Chen, Z.; Jin, H.; Subbarao, K. Replication of live attenuated cold-adapted H2N2 influenza virus vaccine candidates in non human primates. Vaccine 2015, 33, 193–200. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ma, W.; Kahn, R.E.; Richt, J.A. The pig as a mixing vessel for influenza viruses: Human and veterinary implications. J. Mol. Genet. Med. 2008, 3, 158–166. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Xiao, Y.; Liu, J.; Wang, D.; Li, F.; Wang, C.; Li, C.; Zhu, J.; Song, J.; Sun, H.; et al. Prevalent Eurasian avian-like H1N1 swine influenza virus with 2009 pandemic viral genes facilitating human infection. Proc. Natl. Acad. Sci. USA 2020, 117, 17204–17210. [Google Scholar] [CrossRef] [PubMed]
- Mena, I.; Nelson, M.I.; Quezada-Monroy, F.; Dutta, J.; Cortes-Fernández, R.; Lara-Puente, J.H.; Castro-Peralta, F.; Cunha, L.F.; Trovão, N.S.; Lozano-Dubernard, B.; et al. Origins of the 2009 H1N1 influenza pandemic in swine in Mexico. eLife 2016, 5, e16777. [Google Scholar] [CrossRef] [PubMed]
- Iwatsuki-Horimoto, K.; Nakajima, N.; Ichiko, Y.; Sakai-Tagawa, Y.; Noda, T.; Hasegawa, H.; Kawaoka, Y. Syrian Hamster as an Animal Model for the Study of Human Influenza Virus Infection. J. Virol. 2018, 92, e01693-17. [Google Scholar] [CrossRef]
- Nelson, M.I.; Gramer, M.R.; Vincent, A.L.; Holmes, E.C. Global transmission of influenza viruses from humans to swine. J. Gen. Virol. 2012, 93, 2195–2203. [Google Scholar] [CrossRef]
- Nelson, M.I.; Wentworth, D.E.; Culhane, M.R.; Vincent, A.L.; Viboud, C.; LaPointe, M.P.; Lin, X.; Holmes, E.C.; Detmer, S.E. Introductions and Evolution of Human-Origin Seasonal Influenza A Viruses in Multinational Swine Populations. J. Virol. 2014, 88, 10110. [Google Scholar] [CrossRef]
- Ito, T.; Couceiro, J.N.; Kelm, S.; Baum, L.G.; Krauss, S.; Castrucci, M.R.; Donatelli, I.; Kida, H.; Paulson, J.C.; Webster, R.G.; et al. Molecular basis for the generation in pigs of influenza A viruses with pandemic potential. J. Virol. 1998, 72, 7367–7373. [Google Scholar] [CrossRef] [PubMed]
- Kreijtz, J.H.; Suezer, Y.; de Mutsert, G.; van den Brand, J.M.; van Amerongen, G.; Schnierle, B.S.; Kuiken, T.; Fouchier, R.A.; Löwer, J.; Osterhaus, A.D.; et al. Recombinant modified vaccinia virus Ankara expressing the hemagglutinin gene confers protection against homologous and heterologous H5N1 influenza virus infections in macaques. J. Infect. Dis. 2009, 199, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Reeth, K.V.; Brown, I.; Essen, S.; Pensaert, M. Genetic relationships, serological cross-reaction and cross-protection between H1N2 and other influenza A virus subtypes endemic in European pigs. Virus Res. 2004, 103, 115–124. [Google Scholar] [CrossRef] [PubMed]
- Rajão, D.S.; Chen, H.; Perez, D.R.; Sandbulte, M.R.; Gauger, P.C.; Loving, C.L.; Shanks, G.D.; Vincent, A. Vaccine-associated enhanced respiratory disease is influenced by haemagglutinin and neuraminidase in whole inactivated influenza virus vaccines. J. Gen. Virol. 2016, 97, 1489–1499. [Google Scholar] [CrossRef]
- Carroll, T.D.; Jegaskanda, S.; Matzinger, S.R.; Fritts, L.; McChesney, M.B.; Kent, S.J.; Fairman, J.; Miller, C.J. A Lipid/DNA Adjuvant-Inactivated Influenza Virus Vaccine Protects Rhesus Macaques From Uncontrolled Virus Replication After Heterosubtypic Influenza A Virus Challenge. J. Infect. Dis. 2018, 218, 856–867. [Google Scholar] [CrossRef] [PubMed]
- Jegaskanda, S.; Amarasena, T.H.; Laurie, K.L.; Tan, H.X.; Butler, J.; Parsons, M.S.; Alcantara, S.; Petravic, J.; Davenport, M.P.; Hurt, A.C.; et al. Standard trivalent influenza virus protein vaccination does not prime antibody-dependent cellular cytotoxicity in macaques. J. Virol. 2013, 87, 13706–13718. [Google Scholar] [CrossRef] [PubMed]
- Moncla, L.H.; Ross, T.M.; Dinis, J.M.; Weinfurter, J.T.; Mortimer, T.D.; Schultz-Darken, N.; Brunner, K.; Capuano, S.V., 3rd; Boettcher, C.; Post, J.; et al. A novel nonhuman primate model for influenza transmission. PLoS ONE 2013, 8, e78750. [Google Scholar] [CrossRef]
- Davis, A.S.; Taubenberger, J.K.; Bray, M. The use of nonhuman primates in research on seasonal, pandemic and avian influenza, 1893–2014. Antivir. Res. 2015, 117, 75–98. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.A.; Royer, C.M.; Pinkerton, K.E.; Schelegle, E.S. Nonhuman Primate Models of Respiratory Disease: Past, Present, and Future. ILAR J. 2017, 58, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Reber, A.J.; Chirkova, T.; Kim, J.H.; Cao, W.; Biber, R.; Shay, D.K.; Sambhara, S. Immunosenescence and Challenges of Vaccination against Influenza in the Aging Population. Aging Dis. 2012, 3, 68–90. [Google Scholar] [PubMed]
- Tartaglia, L.A.; Dembski, M.; Weng, X.; Deng, N.; Culpepper, J.; Devos, R.; Richards, G.J.; Campfield, L.A.; Clark, F.T.; Deeds, J.; et al. Identification and expression cloning of a leptin receptor, OB-R. Cell 1995, 83, 1263–1271. [Google Scholar] [CrossRef]
- Jegerlehner, A.; Schmitz, N.; Storni, T.; Bachmann, M.F. Influenza A vaccine based on the extracellular domain of M2: Weak protection mediated via antibody-dependent NK cell activity. J. Immunol. 2004, 172, 5598–5605. [Google Scholar] [CrossRef] [PubMed]
- Wei, C.J.; Boyington, J.C.; McTamney, P.M.; Kong, W.P.; Pearce, M.B.; Xu, L.; Andersen, H.; Rao, S.; Tumpey, T.M.; Yang, Z.Y.; et al. Induction of broadly neutralizing H1N1 influenza antibodies by vaccination. Science 2010, 329, 1060–1064. [Google Scholar] [CrossRef]
- Mooij, P.; Koopman, G.; Mortier, D.; van Heteren, M.; Oostermeijer, H.; Fagrouch, Z.; de Laat, R.; Kobinger, G.; Li, Y.; Remarque, E.J.; et al. Pandemic Swine-Origin H1N1 Influenza Virus Replicates to Higher Levels and Induces More Fever and Acute Inflammatory Cytokines in Cynomolgus versus Rhesus Monkeys and Can Replicate in Common Marmosets. PLoS ONE 2015, 10, e0126132. [Google Scholar] [CrossRef]
- Itoh, Y.; Ozaki, H.; Ishigaki, H.; Sakoda, Y.; Nagata, T.; Soda, K.; Isoda, N.; Miyake, T.; Ishida, H.; Okamoto, K.; et al. Subcutaneous inoculation of a whole virus particle vaccine prepared from a non-pathogenic virus library induces protective immunity against H7N7 highly pathogenic avian influenza virus in cynomolgus macaques. Vaccine 2010, 28, 780–789. [Google Scholar] [CrossRef]
- Koday, M.T.; Leonard, J.A.; Munson, P.; Forero, A.; Koday, M.; Bratt, D.L.; Fuller, J.T.; Murnane, R.; Qin, S.; Reinhart, T.A.; et al. Multigenic DNA vaccine induces protective cross-reactive T cell responses against heterologous influenza virus in nonhuman primates. PLoS ONE 2017, 12, e0189780. [Google Scholar] [CrossRef]
- Koopman, G.; Mortier, D.; Michels, S.; Hofman, S.; Fagrouch, Z.; Remarque, E.J.; Verschoor, E.J.; Mooij, P.; Bogers, W. Influenza virus infection as well as immunization with DNA encoding haemagglutinin protein induces potent antibody-dependent phagocytosis (ADP) and monocyte infection-enhancing responses in macaques. J. Gen. Virol. 2019, 100, 738–751. [Google Scholar] [CrossRef]
- Marcelin, G.; Aldridge, J.R.; Duan, S.; Ghoneim, H.E.; Rehg, J.; Marjuki, H.; Boon, A.C.; McCullers, J.A.; Webby, R.J. Fatal outcome of pandemic H1N1 2009 influenza virus infection is associated with immunopathology and impaired lung repair, not enhanced viral burden, in pregnant mice. J. Virol. 2011, 85, 11208–11219. [Google Scholar] [CrossRef]
- Kim, Y.H.; Kim, J.K.; Kim, D.J.; Nam, J.H.; Shim, S.M.; Choi, Y.K.; Lee, C.H.; Poo, H. Diet-induced obesity dramatically reduces the efficacy of a 2009 pandemic H1N1 vaccine in a mouse model. J. Infect. Dis. 2012, 205, 244–251. [Google Scholar] [CrossRef]
- Ren, Y.Y.; Yin, Y.Y.; Li, W.Q.; Lin, Y.; Liu, T.; Wang, S.; Zhang, S.Y.; Li, Z.; Wang, X.J.; Bi, Z.Q. Risk factors associated with severe manifestations of 2009 pandemic influenza A (H1N1) infection in China: A case-control study. Virol. J. 2013, 10, 149. [Google Scholar] [CrossRef] [PubMed]
- Milner, J.J.; Beck, M.A. The impact of obesity on the immune response to infection. Proc. Nutr. Soc. 2012, 71, 298–306. [Google Scholar] [CrossRef]
- Bradley, S.F. Influenza in the elderly. Prevention is the best strategy in high-risk populations. Postgrad. Med. 1996, 99, 138–139. [Google Scholar] [PubMed]
- Green, W.D.; Beck, M.A. Obesity Impairs the Adaptive Immune Response to Influenza Virus. Ann. Am. Thorac. Soc. 2017, 14, S406–S409. [Google Scholar] [CrossRef]
- Whitley, R.J.; Monto, A.S. Prevention and treatment of influenza in high-risk groups: Children, pregnant women, immunocompromised hosts, and nursing home residents. J. Infect. Dis. 2006, 194 (Suppl. 2), S133–S138. [Google Scholar] [CrossRef] [PubMed]
- van Riel, D.; Mittrucker, H.W.; Engels, G.; Klingel, K.; Markert, U.R.; Gabriel, G. Influenza pathogenicity during pregnancy in women and animal models. Semin. Immunopathol. 2016, 38, 719–726. [Google Scholar] [CrossRef]
- Gottesman, M.E.; Quadro, L.; Blaner, W.S. Studies of vitamin A metabolism in mouse model systems. BioEssays 2001, 23, 409–419. [Google Scholar] [CrossRef]
- Seldeen, K.L.; Pang, M.; Rodríguez-Gonzalez, M.; Hernandez, M.; Sheridan, Z.; Yu, P.; Troen, B.R. A mouse model of vitamin D insufficiency: Is there a relationship between 25(OH) vitamin D levels and obesity? Nutr. Metab. 2017, 14, 26. [Google Scholar] [CrossRef]
- Surman, S.L.; Penkert, R.R.; Jones, B.G.; Sealy, R.E.; Hurwitz, J.L. Vitamin Supplementation at the Time of Immunization with a Cold-Adapted Influenza Virus Vaccine Corrects Poor Mucosal Antibody Responses in Mice Deficient for Vitamins A and D. Clin. Vaccine Immunol. 2016, 23, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Yasui, H.; Kiyoshima, J.; Hori, T. Reduction of influenza virus titer and protection against influenza virus infection in infant mice fed Lactobacillus casei Shirota. Clin. Diagn. Lab. Immunol. 2004, 11, 675–679. [Google Scholar] [CrossRef]
- Pollett, M.; Mackenzie, J.S.; Turner, K.J. The effect of protein-deprivation on the susceptibility to influenza virus infection: A murine model system. Aust. J. Exp. Biol. Med. Sci. 1979, 57, 151–160. [Google Scholar] [CrossRef]
- Penkert, R.R.; Iverson, A.; Rosch, J.W.; Hurwitz, J.L. Prevnar-13 vaccine failure in a mouse model for vitamin A deficiency. Vaccine 2017, 35, 6264–6268. [Google Scholar] [CrossRef] [PubMed]
- Ritz, B.W.; Aktan, I.; Nogusa, S.; Gardner, E.M. Energy restriction impairs natural killer cell function and increases the severity of influenza infection in young adult male C57BL/6 mice. J. Nutr. 2008, 138, 2269–2275. [Google Scholar] [CrossRef] [PubMed]
- Stephensen, C.B.; Blount, S.R.; Schoeb, T.R.; Park, J.Y. Vitamin A deficiency impairs some aspects of the host response to influenza A virus infection in BALB/c mice. J. Nutr. 1993, 123, 823–833. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.K.; Cao, W.; Vora, K.P.; De La Cruz, J.; Shieh, W.J.; Zaki, S.R.; Katz, J.M.; Sambhara, S.; Gangappa, S. Protein energy malnutrition decreases immunity and increases susceptibility to influenza infection in mice. J. Infect. Dis. 2013, 207, 501–510. [Google Scholar] [CrossRef]
- Hernandez-Vargas, E.A.; Wilk, E.; Canini, L.; Toapanta, F.R.; Binder, S.C.; Uvarovskii, A.; Ross, T.M.; Guzmán, C.A.; Perelson, A.S.; Meyer-Hermann, M. Effects of Aging on Influenza Virus Infection Dynamics. J. Virol. 2014, 88, 4123. [Google Scholar] [CrossRef]
- Lu, J.; Duan, X.; Zhao, W.; Wang, J.; Wang, H.; Zhou, K.; Fang, M. Aged Mice are More Resistant to Influenza Virus Infection due to Reduced Inflammation and Lung Pathology. Aging Dis. 2018, 9, 358–373. [Google Scholar] [CrossRef]
- Maue, A.C.; Yager, E.J.; Swain, S.L.; Woodland, D.L.; Blackman, M.A.; Haynes, L. T-cell immunosenescence: Lessons learned from mouse models of aging. Trends Immunol. 2009, 30, 301–305. [Google Scholar] [CrossRef]
- Yoon, S.W.; Wong, S.S.; Zhu, H.; Chen, R.; Li, L.; Zhang, Y.; Guan, Y.; Webby, R.J. Dysregulated T-Helper Type 1 (Th1):Th2 Cytokine Profile and Poor Immune Response in Pregnant Ferrets Infected With 2009 Pandemic Influenza A(H1N1) Virus. J. Infect. Dis. 2018, 217, 438–442. [Google Scholar] [CrossRef] [PubMed]
- Carroll, T.D.; Matzinger, S.R.; Barry, P.A.; McChesney, M.B.; Fairman, J.; Miller, C.J. Efficacy of Influenza Vaccination of Elderly Rhesus Macaques Is Dramatically Improved by Addition of a Cationic Lipid/DNA Adjuvant. J. Infect. Dis. 2013, 209, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Chan, K.H.; Zhang, A.J.; To, K.K.; Chan, C.C.; Poon, V.K.; Guo, K.; Ng, F.; Zhang, Q.W.; Leung, V.H.; Cheung, A.N.; et al. Wild type and mutant 2009 pandemic influenza A (H1N1) viruses cause more severe disease and higher mortality in pregnant BALB/c mice. PLoS ONE 2010, 5, e13757. [Google Scholar] [CrossRef]
- Engels, G.; Hierweger, A.M.; Hoffmann, J.; Thieme, R.; Thiele, S.; Bertram, S.; Dreier, C.; Resa-Infante, P.; Jacobsen, H.; Thiele, K.; et al. Pregnancy-Related Immune Adaptation Promotes the Emergence of Highly Virulent H1N1 Influenza Virus Strains in Allogenically Pregnant Mice. Cell Host Microbe 2017, 21, 321–333. [Google Scholar] [CrossRef] [PubMed]
- Ilievski, V.; Lu, S.J.; Hirsch, E. Activation of toll-like receptors 2 or 3 and preterm delivery in the mouse. Reprod. Sci. 2007, 14, 315–320. [Google Scholar] [CrossRef]
- Kim, J.C.; Kim, H.M.; Kang, Y.M.; Ku, K.B.; Park, E.H.; Yum, J.; Kim, J.A.; Kang, Y.K.; Lee, J.S.; Kim, H.S.; et al. Severe pathogenesis of influenza B virus in pregnant mice. Virology 2014, 448, 74–81. [Google Scholar] [CrossRef]
- Littauer, E.Q.; Esser, E.S.; Antao, O.Q.; Vassilieva, E.V.; Compans, R.W.; Skountzou, I. H1N1 influenza virus infection results in adverse pregnancy outcomes by disrupting tissue-specific hormonal regulation. PLoS Pathog. 2017, 13, e1006757. [Google Scholar] [CrossRef]
- van der Lubbe, J.E.M.; Vreugdenhil, J.; Damman, S.; Vaneman, J.; Klap, J.; Goudsmit, J.; Radošević, K.; Roozendaal, R. Maternal antibodies protect offspring from severe influenza infection and do not lead to detectable interference with subsequent offspring immunization. Virol. J. 2017, 14, 123. [Google Scholar] [CrossRef]
- Vermillion, M.S.; Nelson, A.; Vom Steeg, L.; Loube, J.; Mitzner, W.; Klein, S.L. Pregnancy preserves pulmonary function following influenza virus infection in C57BL/6 mice. Am. J. Physiol. Lung Cell Mol. Physiol. 2018, 315, L517–L525. [Google Scholar] [CrossRef]
- Powers, D.C.; Belshe, R.B. Effect of Age on Cytotoxic T Lymphocyte Memory as well as Serum and Local Antibody Responses Elicited by Inactivated Influenza Virus Vaccine. J. Infect. Dis. 1993, 167, 584–592. [Google Scholar] [CrossRef] [PubMed]
- Bosaeed, M.; Kumar, D. Seasonal influenza vaccine in immunocompromised persons. Hum. Vaccines Immunother. 2018, 14, 1311–1322. [Google Scholar] [CrossRef]
- DiazGranados, C.A.; Dunning, A.J.; Kimmel, M.; Kirby, D.; Treanor, J.; Collins, A.; Pollak, R.; Christoff, J.; Earl, J.; Landolfi, V.; et al. Efficacy of high-dose versus standard-dose influenza vaccine in older adults. N. Engl. J. Med. 2014, 371, 635–645. [Google Scholar] [CrossRef]
- Robertson, C.A.; DiazGranados, C.A.; Decker, M.D.; Chit, A.; Mercer, M.; Greenberg, D.P. Fluzone® High-Dose Influenza Vaccine. Expert Rev. Vaccines 2016, 15, 1495–1505. [Google Scholar] [CrossRef]
- Sun, X.; Yan, Z.; Yi, Y.; Li, Z.; Lei, D.; Rogers, C.S.; Chen, J.; Zhang, Y.; Welsh, M.J.; Leno, G.H.; et al. Adeno-associated virus-targeted disruption of the CFTR gene in cloned ferrets. J. Clin. Investig. 2008, 118, 1578–1583. [Google Scholar] [CrossRef]
- Kou, Z.; Wu, Q.; Kou, X.; Yin, C.; Wang, H.; Zuo, Z.; Zhuo, Y.; Chen, A.; Gao, S.; Wang, X. CRISPR/Cas9-mediated genome engineering of the ferret. Cell Res. 2015, 25, 1372–1375. [Google Scholar] [CrossRef] [PubMed]
- Min, J.-Y.; Vogel, L.; Matsuoka, Y.; Lu, B.; Swayne, D.; Jin, H.; Kemble, G.; Subbarao, K. A Live Attenuated H7N7 Candidate Vaccine Virus Induces Neutralizing Antibody That Confers Protection from Challenge in Mice, Ferrets, and Monkeys. J. Virol. 2010, 84, 11950. [Google Scholar] [CrossRef] [PubMed]
- Paquette, S.G.; Huang, S.S.H.; Banner, D.; Xu, L.; Leόn, A.; Kelvin, A.A.; Kelvin, D.J. Impaired heterologous immunity in aged ferrets during sequential influenza A H1N1 infection. Virology 2014, 464, 177–183. [Google Scholar] [CrossRef] [PubMed]
- Paquette, S.G.; Banner, D.; Huang, S.S.; Almansa, R.; Leon, A.; Xu, L.; Bartoszko, J.; Kelvin, D.J.; Kelvin, A.A. Influenza Transmission in the Mother-Infant Dyad Leads to Severe Disease, Mammary Gland Infection, and Pathogenesis by Regulating Host Responses. PLoS Pathog. 2015, 11, e1005173. [Google Scholar] [CrossRef] [PubMed]
- Fukuyama, S.; Iwatsuki-Horimoto, K.; Kiso, M.; Nakajima, N.; Gregg, R.W.; Katsura, H.; Tomita, Y.; Maemura, T.; da Silva Lopes, T.J.; Watanabe, T.; et al. Pathogenesis of Influenza A(H7N9) Virus in Aged Nonhuman Primates. J. Infect. Dis. 2020, 222, 1155–1164. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, B.C.; Kim, J.R.; Blevins, L.K.; Jorgensen, M.J.; Kock, N.D.; D’Agostino, R.B.; Aycock, S.T.; Hadimani, M.B.; King, S.B.; Parks, G.D.; et al. A Novel R848-Conjugated Inactivated Influenza Virus Vaccine Is Efficacious and Safe in a Neonate Nonhuman Primate Model. J. Immunol. 2016, 197, 555–564. [Google Scholar] [CrossRef]
- Killingley, B.; Enstone, J.; Booy, R.; Hayward, A.; Oxford, J.; Ferguson, N.; Nguyen Van-Tam, J. Potential role of human challenge studies for investigation of influenza transmission. Lancet Infect. Dis. 2011, 11, 879–886. [Google Scholar] [CrossRef]
- Holbrook, B.C.; D’Agostino, R.B.; Tyler Aycock, S.; Jorgensen, M.J.; Hadimani, M.B.; Bruce King, S.; Alexander-Miller, M.A. Adjuvanting an inactivated influenza vaccine with conjugated R848 improves the level of antibody present at 6months in a nonhuman primate neonate model. Vaccine 2017, 35, 6137–6142. [Google Scholar] [CrossRef] [PubMed]
- Holbrook, B.C.; D’Agostino, R.B.; Parks, G.D.; Alexander-Miller, M.A. Adjuvanting an inactivated influenza vaccine with flagellin improves the function and quantity of the long-term antibody response in a nonhuman primate neonate model. Vaccine 2016, 34, 4712–4717. [Google Scholar] [CrossRef]
- Holbrook, B.C.; Hayward, S.L.; Blevins, L.K.; Kock, N.; Aycock, T.; Parks, G.D.; Alexander-Miller, M.A. Nonhuman primate infants have an impaired respiratory but not systemic IgG antibody response following influenza virus infection. Virology 2015, 476, 124–133. [Google Scholar] [CrossRef] [PubMed]
- Bauer, S.A.; Arndt, T.P.; Leslie, K.E.; Pearl, D.L.; Turner, P.V. Obesity in Rhesus and Cynomolgus Macaques: A Comparative Review of the Condition and Its Implications for Research. Comp. Med. 2011, 61, 514–526. [Google Scholar] [PubMed]
- Forero, A.; Fenstermacher, K.; Wohlgemuth, N.; Nishida, A.; Carter, V.; Smith, E.A.; Peng, X.; Hayes, M.; Francis, D.; Treanor, J.; et al. Evaluation of the innate immune responses to influenza and live-attenuated influenza vaccine infection in primary differentiated human nasal epithelial cells. Vaccine 2017, 35, 6112–6121. [Google Scholar] [CrossRef]
- Gerlach, R.L.; Camp, J.V.; Chu, Y.-K.; Jonsson, C.B. Early Host Responses of Seasonal and Pandemic Influenza A Viruses in Primary Well-Differentiated Human Lung Epithelial Cells. PLoS ONE 2013, 8, e78912. [Google Scholar] [CrossRef]
- Hauser, M.J.; Dlugolenski, D.; Culhane, M.R.; Wentworth, D.E.; Tompkins, S.M.; Tripp, R.A. Antiviral Responses by Swine Primary Bronchoepithelial Cells Are Limited Compared to Human Bronchoepithelial Cells Following Influenza Virus Infection. PLoS ONE 2013, 8, e70251. [Google Scholar] [CrossRef]
- Ilyushina, N.A.; Ikizler, M.R.; Kawaoka, Y.; Rudenko, L.G.; Treanor, J.J.; Subbarao, K.; Wright, P.F. Comparative Study of Influenza Virus Replication in MDCK Cells and in Primary Cells Derived from Adenoids and Airway Epithelium. J. Virol. 2012, 86, 11725. [Google Scholar] [CrossRef]
- Jones, J.C.; Baranovich, T.; Marathe, B.M.; Danner, A.F.; Seiler, J.P.; Franks, J.; Govorkova, E.A.; Krauss, S.; Webster, R.G. Risk Assessment of H2N2 Influenza Viruses from the Avian Reservoir. J. Virol. 2014, 88, 1175. [Google Scholar] [CrossRef]
- Martinez-Sobrido, L.; Blanco-Lobo, P.; Rodriguez, L.; Fitzgerald, T.; Zhang, H.; Nguyen, P.; Anderson, C.S.; Holden-Wiltse, J.; Bandyopadhyay, S.; Nogales, A.; et al. Characterizing Emerging Canine H3 Influenza Viruses. PLoS Pathog. 2020, 16, e1008409. [Google Scholar] [CrossRef]
- Schultz-Cherry, S.; Thomas, P.G. Assessing the fitness of distinct clades of influenza A (H9N2) viruses. Emerg. Microbes Infect. 2013, 2, 1–11. [Google Scholar] [CrossRef]
- Bueno de Mesquita, P.J.; Noakes, C.J.; Milton, D.K. Quantitative aerobiologic analysis of an influenza human challenge-transmission trial. Indoor Air 2020, 30, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Han, A.; Czajkowski, L.M.; Donaldson, A.; Baus, H.A.; Reed, S.M.; Athota, R.S.; Bristol, T.; Rosas, L.A.; Cervantes-Medina, A.; Taubenberger, J.K.; et al. A Dose-finding Study of a Wild-type Influenza A(H3N2) Virus in a Healthy Volunteer Human Challenge Model. Clin. Infect. Dis. 2019, 69, 2082–2090. [Google Scholar] [CrossRef]
- Han, A.; Poon, J.L.; Powers, J.H., 3rd; Leidy, N.K.; Yu, R.; Memoli, M.J. Using the Influenza Patient-reported Outcome (FLU-PRO) diary to evaluate symptoms of influenza viral infection in a healthy human challenge model. BMC Infect. Dis. 2018, 18, 353. [Google Scholar] [CrossRef]
- Innis, B.L.; Berlanda Scorza, F.; Blum, J.S.; Jain, V.K.; Older Aguilar, A.; Post, D.J.; Roberts, P.C.; Wairagkar, N.; White, J.; Bresee, J. Meeting report: Convening on the influenza human viral challenge model for universal influenza vaccines, Part 1: Value; challenge virus selection; regulatory, industry and ethical considerations; increasing standardization, access and capacity. Vaccine 2019, 37, 4823–4829. [Google Scholar] [CrossRef]
- Killingley, B.; Enstone, J.E.; Greatorex, J.; Gilbert, A.S.; Lambkin-Williams, R.; Cauchemez, S.; Katz, J.M.; Booy, R.; Hayward, A.; Oxford, J.; et al. Use of a human influenza challenge model to assess person-to-person transmission: Proof-of-concept study. J. Infect. Dis. 2012, 205, 35–43. [Google Scholar] [CrossRef]
- Lambkin-Williams, R.; Noulin, N.; Mann, A.; Catchpole, A.; Gilbert, A.S. The human viral challenge model: Accelerating the evaluation of respiratory antivirals, vaccines and novel diagnostics. Respir. Res. 2018, 19, 123. [Google Scholar] [CrossRef]
- Liebowitz, D.; Gottlieb, K.; Kolhatkar, N.S.; Garg, S.J.; Asher, J.M.; Nazareno, J.; Kim, K.; McIlwain, D.R.; Tucker, S.N. Efficacy, immunogenicity, and safety of an oral influenza vaccine: A placebo-controlled and active-controlled phase 2 human challenge study. Lancet Infect. Dis. 2020, 20, 435–444. [Google Scholar] [CrossRef]
- McBride, J.M.; Lim, J.J.; Burgess, T.; Deng, R.; Derby, M.A.; Maia, M.; Horn, P.; Siddiqui, O.; Sheinson, D.; Chen-Harris, H.; et al. Phase 2 Randomized Trial of the Safety and Efficacy of MHAA4549A, a Broadly Neutralizing Monoclonal Antibody, in a Human Influenza A Virus Challenge Model. Antimicrob. Agents Chemother. 2017, 61. [Google Scholar] [CrossRef]
- Memoli, M.J.; Shaw, P.A.; Han, A.; Czajkowski, L.; Reed, S.; Athota, R.; Bristol, T.; Fargis, S.; Risos, K.; Powers, J.H.; et al. Evaluation of Antihemagglutinin and Antineuraminidase Antibodies as Correlates of Protection in an Influenza A/H1N1 Virus Healthy Human Challenge Model. mBio 2016, 7, e00417-16. [Google Scholar] [CrossRef]
- Nguyen-Van-Tam, J.S.; Killingley, B.; Enstone, J.; Hewitt, M.; Pantelic, J.; Grantham, M.L.; Bueno de Mesquita, P.J.; Lambkin-Williams, R.; Gilbert, A.; Mann, A.; et al. Minimal transmission in an influenza A (H3N2) human challenge-transmission model within a controlled exposure environment. PLoS Pathog. 2020, 16, e1008704. [Google Scholar] [CrossRef] [PubMed]
- Park, J.K.; Xiao, Y.; Ramuta, M.D.; Rosas, L.A.; Fong, S.; Matthews, A.M.; Freeman, A.D.; Gouzoulis, M.A.; Batchenkova, N.A.; Yang, X.; et al. Pre-existing immunity to influenza virus hemagglutinin stalk might drive selection for antibody-escape mutant viruses in a human challenge model. Nat. Med. 2020, 26, 1240–1246. [Google Scholar] [CrossRef]
- Pharo, E.A.; Williams, S.M.; Boyd, V.; Sundaramoorthy, V.; Durr, P.A.; Baker, M.L. Host-Pathogen Responses to Pandemic Influenza H1N1pdm09 in a Human Respiratory Airway Model. Viruses 2020, 12, 679. [Google Scholar] [CrossRef]
- Sherman, A.C.; Mehta, A.; Dickert, N.W.; Anderson, E.J.; Rouphael, N. The Future of Flu: A Review of the Human Challenge Model and Systems Biology for Advancement of Influenza Vaccinology. Front. Cell Infect. Microbiol. 2019, 9, 107. [Google Scholar] [CrossRef]
- Teunis, P.F.; Brienen, N.; Kretzschmar, M.E. High infectivity and pathogenicity of influenza A virus via aerosol and droplet transmission. Epidemics 2010, 2, 215–222. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Trade Name | Manufacturer | Category | Available as | Demographic |
|---|---|---|---|---|
| AFLURIA | Seqirus Pty. Ltd. | IIV | TIV, QIV | Persons > 6 months of age |
| Agriflu | Seqirus Inc. | IIV | TIV | Persons > 18 years of age |
| FLUAD a | Seqirus, Inc. | IIV | TIV | Persons > 65 years of age |
| FluMist | MedImmune | LAIV | QIV | Persons 2–49 years of age |
| Fluarix | GlaxoSmithKline Biologicals | IIV | TIV | Persons > 3 years of age |
| Fluarix Quadrivalent | GlaxoSmithKline Biologicals | IIV | QIV | Persons > 6 months of age |
| Flublok | Protein Sciences Corporation | Recombinant HA | TIV, QIV | Persons > 18 years of age |
| Flucelvax | Seqirus, Inc. | IIV | TIV | Persons > 4 years of age |
| Flucelvax b | Seqirus, Inc. | IIV | QIV | Persons > 4 years of age |
| FluLaval | ID Biomedical Corporation of Quebec | IIV | TIV, QIV | Persons > 6 months of age |
| Fluvirin | Seqirus Vaccines Limited | IIV | TIV | Persons > 4 years of age |
| Fluzone c | Sanofi Pasteur Inc. | IIV | TIV, QIV | Persons > 6 months of age |
| Influenza Virus Vaccine, H5N1 d | Sanofi Pasteur Inc | IIV | Monovalent | Persons 18 through 64 years of age |
| Influenza A (H5N1) Virus Monovalent Vaccine e | ID Biomedical Corporation of Quebec | IIV | Monovalent | Persons > 6 months of age |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roubidoux, E.K.; Schultz-Cherry, S. Animal Models Utilized for the Development of Influenza Virus Vaccines. Vaccines 2021, 9, 787. https://doi.org/10.3390/vaccines9070787
Roubidoux EK, Schultz-Cherry S. Animal Models Utilized for the Development of Influenza Virus Vaccines. Vaccines. 2021; 9(7):787. https://doi.org/10.3390/vaccines9070787
Chicago/Turabian StyleRoubidoux, Ericka Kirkpatrick, and Stacey Schultz-Cherry. 2021. "Animal Models Utilized for the Development of Influenza Virus Vaccines" Vaccines 9, no. 7: 787. https://doi.org/10.3390/vaccines9070787
APA StyleRoubidoux, E. K., & Schultz-Cherry, S. (2021). Animal Models Utilized for the Development of Influenza Virus Vaccines. Vaccines, 9(7), 787. https://doi.org/10.3390/vaccines9070787