
VLP-Based Vaccines as a Suitable Technology to Target Trypanosomatid Diseases

 ,
,  ,
,  , and
, and 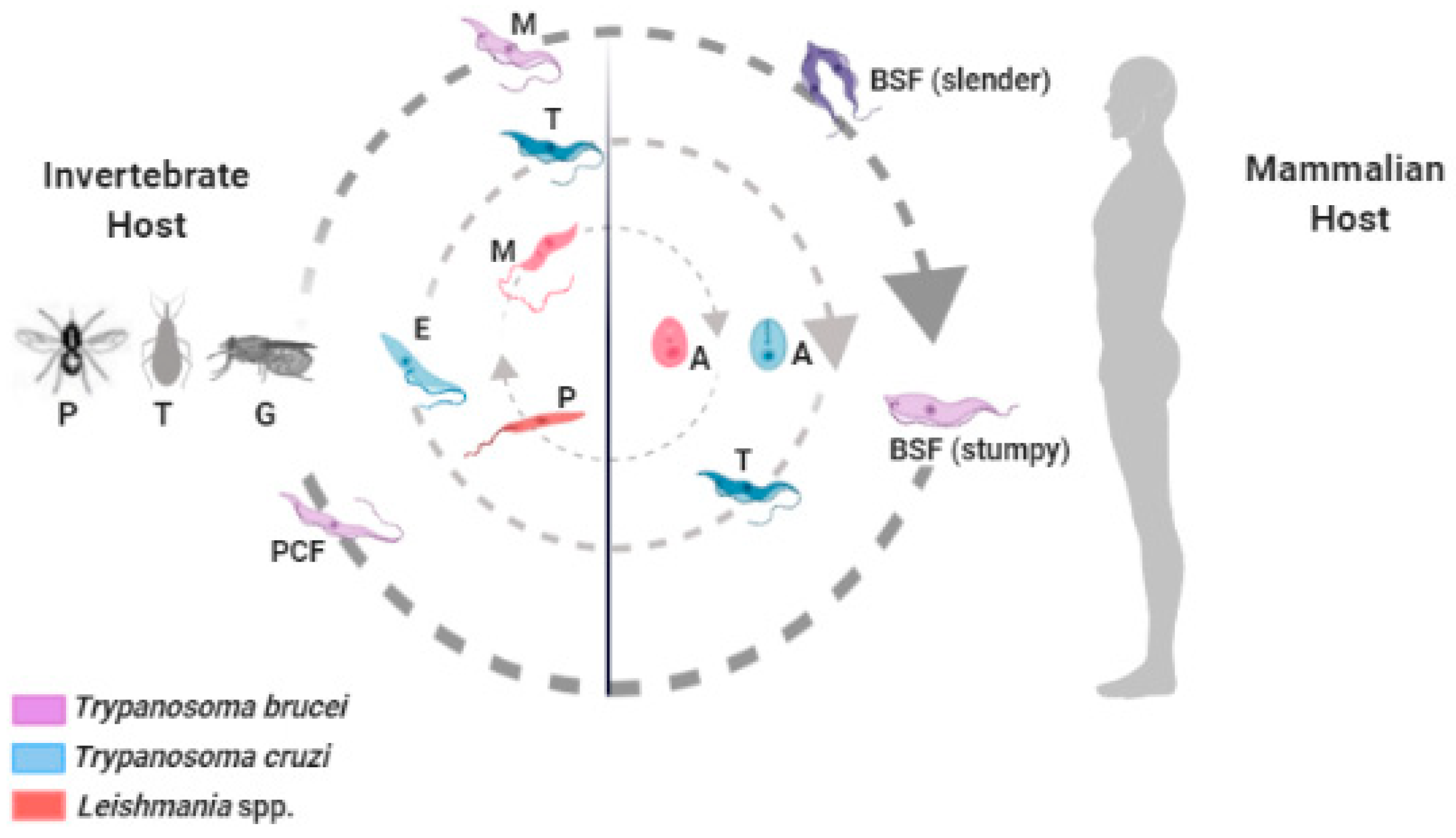{kind=link}
{kind=link}
Abstract
1. Introduction
2. Application of the Use of VLPs in the Development of New Vaccines
3. Vaccines to Control Diseases Caused by Trypanosomatids: What Do We Need and Where Are We?
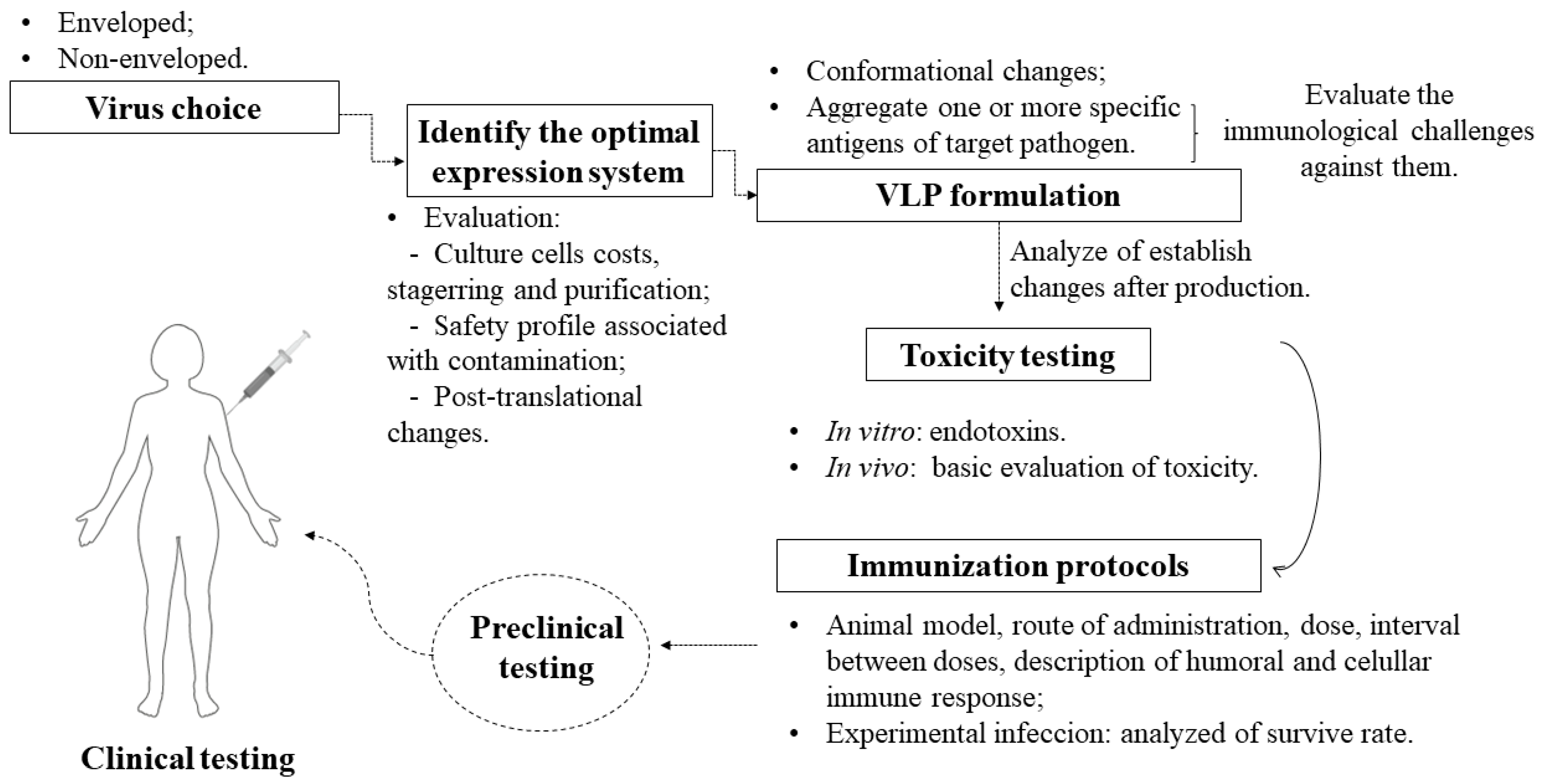
4. Development of Vaccines Based on VLPs: A Checklist
- Should the chosen virus be enveloped or not? Enveloped VLPs are inherited from structured viral proteins, thus generating a particle with target antigens integrated on the external surface, whereas non-enveloped VLPs are based on the expression of one or more necessary viral proteins without inheriting them from a host [56]. Both have the additional possibility to integrate antigens [57] and additional adjuvants [7] in vaccine formulations.
- Which expression system is ideal? It should be noted that, depending on the system, there will be limitations such as cost and difficulty in scheduling [58], as well as a higher level of purification when faced with contamination from cells [59]. Some systems differ from mammalian cells mainly in the protein glycosylation phase [60] and do not give rise to a eukaryotic expression environment. The insect cell system has come to solve some problems in the production of particulates, as it has characteristics such as a eukaryotic environment, which is required for the glycosylation of proteins: this post-translational characteristic is also found in human cells, but the cost of their application is higher [61]. In addition, there is malleability in large-scale cultivation and the possibility of using it for the simultaneous expression of many proteins that facilitate the assembly of VLPs, which requires a structure with a high level of safety and a special culture system [58].
- The structural modification of the conformational and immunological characteristics of the VLP can adjust the relative degree of the immune response for a balanced induction of humoral and cellular immunity [62], thereby generating an improved candidate capable of redirecting the immune response against specific targets when combined with antigens [63]. Thus, this modulates the balance of Th1 and Th2 immune responses to increase the specificity and affinity towards the parasite [48]. Therefore, conformational changes and the aggregation of antigens are necessary to analyze changes in stability that can compromise the effectiveness of VLP vaccines [64].
- Before choosing the antigens, it is necessary to assess the immunological challenges found in the parasites that cause Chagas disease, leishmaniasis, and sleeping sickness. They have specific mechanisms of escape from the immune system that end up reducing the specific response of the immune system. Based on this assessment, one can then envision a single vaccine for three diseases or specific vaccines for each one, as VLPs will act as transporters of antigens, with greater security than using soluble antigens [6]. The applicability of antigens presented in various evolutionary forms of the protozoan, as well as the attempt to administer immunizations with more than one antigen to cover all forms of life and increase the antigenic repertoire, must be considered in studies on T. cruzi, given the genetic variability of this parasite [65]. In addition, the use of the same antigens for both prophylactic and therapeutic strategies should be considered, taking into account that prophylactic and therapeutic vaccines can prevent infection and interfere with the progression of infection, respectively. For an infected person, a therapeutic vaccine with or without an association with available drugs can have a significant impact on preventing complications, as has already been reported for Chagas disease [66].
- After the production and necessary testing of these particles in isolation and in association with antigens, in vitro and in vivo tests need to be performed, for example, endotoxin evaluation [67]. In addition, a basic toxicity assessment must be performed, according to the WHO guidelines for non-clinical evaluation of vaccines [68]. This step can also assist in understanding the humoral and cellular immunogenicity of the isolated particles without antigen association. The choice of adjuvant must include consideration of the cost–benefit trade-off of co-stimulation, as VLPs can assume the role of self-adjuvant due to their particulate and multivalent nature, causing efficient incorporation by antigen-presenting cells (APCs) [69,70].
- In vivo tests on immunization strategies with these specific antigens and/or with a chimeric VLP should be performed by firstly establishing the animal model, time, and doses, characterizing the humoral and cellular immune responses, and importantly, considering different routes of administration to understand whether this influences the stability of VLPs [16].
- Viral challenges in animals (experimental infection) used in experiments are necessary to understand, for example, whether the formulation of specific antigens with VLP technology helps or not in the survival of those infected. After pre-clinical trials proceed as expected, clinical trials have a significant step in producing results that confirm a specific, lasting, and harmless response in humans.
5. Final Considerations and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- DNDi—Drugs for Neglected Diseases Initiative. Diseases. 2019. Available online: https://dndi.org/diseases/ (accessed on 10 December 2020).
- Pérez-Molina, J.; Molina, I. Chagas disease. Lancet 2018, 391, 82–94. [Google Scholar] [CrossRef]
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Kennedy, P.G.E. Update on human African trypanosomiasis (sleeping sickness). J. Neurol. 2019, 266, 2334–2337. [Google Scholar] [CrossRef] [PubMed]
- Chroboczek, J.; Szurgot, I.; Szolajska, E. Virus-like particles as vaccine. Acta Biochim. Pol. 2014, 61, 531–539. [Google Scholar] [CrossRef]
- Crisci, E.; Bárcena, J.; Montoya, M. Virus-like particles: The new frontier of vaccines for animal viral infections. Veter Immunol. Immunopathol. 2012, 148, 211–225. [Google Scholar] [CrossRef]
- Gomes, A.C.; Flace, A.; Saudan, P.; Zabel, F.; Cabral-Miranda, G.; El Turabi, A.; Manolova, V.; Bachmann, M.F. Adjusted Particle Size Eliminates the Need of Linkage of Antigen and Adjuvants for Appropriated T Cell Responses in Virus-Like Particle-Based Vaccines. Front. Immunol. 2017, 8, 226. [Google Scholar] [CrossRef]
- Grgacic, E.V.; Anderson, D.A. Virus-like particles: Passport to immune recognition. Methods 2006, 40, 60–65. [Google Scholar] [CrossRef]
- Zeltins, A. Construction and Characterization of Virus-Like Particles: A Review. Mol. Biotechnol. 2012, 53, 92–107. [Google Scholar] [CrossRef]
- Pijlman, G.P. Enveloped virus-like particles as vaccines against pathogenic arboviruses. Biotechnol. J. 2015, 10, 659–670. [Google Scholar] [CrossRef] [PubMed]
- Venters, C.; Graham, W.; Cassidy, W. Recombivax-HB: Perspectives past, present and future. Expert Rev. Vaccines 2004, 3, 119–129. [Google Scholar] [CrossRef] [PubMed]
- Hieu, N.T.; Kim, K.H.; Janowicz, Z.; Timmermans, I. Comparative efficacy, safety and immunogenicity of Hepavax-Gene and Engerix-B, recombinant hepatitis B vaccines, in infants born to HBsAg and HBeAg positive mothers in Vietnam: An assessment at 2 years. Vaccine 2002, 20, 1803–1808. [Google Scholar] [CrossRef]
- McLemore, M.R. Gardasil®: Introducing the New Human Papillomavirus Vaccine. Clin. J. Oncol. Nurs. 2006, 10, 559–560. [Google Scholar] [CrossRef] [PubMed]
- Paavonen, J.; Jenkins, D.; Bosch, F.X.; Naud, P.; Salmerón, J.; Wheeler, C.M.; Chow, S.-N.; Apter, D.L.; Kitchener, H.C.; Castellsague, X.; et al. Efficacy of a prophylactic adjuvanted bivalent L1 virus-like-particle vaccine against infection with human papillomavirus types 16 and 18 in young women: An interim analysis of a phase III double-blind, randomised controlled trial. Lancet 2007, 369, 2161–2170. [Google Scholar] [CrossRef]
- Herrero, R.; Hildesheim, A.; Rodríguez, A.C.; Wacholder, S.; Bratti, C.; Solomon, D.; González, P.; Porras, C.; Jiménez, S.; Guillen, D.; et al. Rationale and design of a community-based double-blind randomized clinical trial of an HPV 16 and 18 vaccine in Guanacaste, Costa Rica. Vaccine 2008, 26, 4795–4808. [Google Scholar] [CrossRef]
- Qian, C.; Liu, X.; Xu, Q.; Wang, Z.; Chen, J.; Li, T.; Zheng, Q.; Yu, H.; Gu, Y.; Li, S.; et al. Recent Progress on the Versatility of Virus-Like Particles. Vaccines 2020, 8, 139. [Google Scholar] [CrossRef]
- Singh, K.; Marasini, B.; Chen, X.; Ding, L.; Wang, J.-J.; Xiao, P.; Villinger, F.; Spearman, P. A Bivalent, Spherical Virus-Like Particle Vaccine Enhances Breadth of Immune Responses against Pathogenic Ebola Viruses in Rhesus Macaques. J. Virol. 2020, 94. [Google Scholar] [CrossRef] [PubMed]
- Metz, S.W.; Pijlman, G.P. Production of Chikungunya Virus-Like Particles and Subunit Vaccines in Insect Cells. In Chikungunya Virus: Methods and Protocols; Humana Press: Totowa, NJ, USA, 2016; Volume 1426, pp. 297–309. [Google Scholar]
- Metz, S.W.; Thomas, A.; White, L.; Stoops, M.; Corten, M.; Hannemann, H.; De Silva, A.M. Dengue virus-like particles mimic the antigenic properties of the infectious dengue virus envelope. Virol. J. 2018, 15, 1–10. [Google Scholar] [CrossRef]
- Pillet, S.; Couillard, J.; Trépanier, S.; Poulin, J.-F.; Yassine-Diab, B.; Guy, B.; Ward, B.J.; Landry, N. Immunogenicity and safety of a quadrivalent plant-derived virus like particle influenza vaccine candidate—Two randomized Phase II clinical trials in 18 to 49 and ≥50 years old adults. PLoS ONE 2019, 14, e0216533. [Google Scholar] [CrossRef]
- Huertas-Díaz, M.C.; Phan, S.; Elson, A.; Nuñez, I.; Wei, H.; Sakamoto, K.; He, B. Parainfluenza virus 5 (PIV5) amplifying virus-like particles expressing respiratory syncytial virus (RSV) antigens protect mice against RSV infection. Vaccine 2019, 37, 2925–2934. [Google Scholar] [CrossRef]
- Boigard, H.; Alimova, A.; Martin, G.R.; Katz, A.; Gottlieb, P.; Galarza, J.M. Zika virus-like particle (VLP) based vaccine. PLoS Negl. Trop. Dis. 2017, 11, e0005608. [Google Scholar] [CrossRef]
- Jiménez-Chávez, Á.d.J.; Moreno-Fierros, L.; Bustos-Jaimes, I. Therapy with multi-epitope virus-like particles of B19 parvovirus reduce tumor growth and lung metastasis in an aggressive breast cancer mouse model. Vaccine 2019, 37, 7256–7268. [Google Scholar] [CrossRef]
- World Health Organization. DRAFT Landscape of COVID-19 Candidate Vaccines. 2021. Available online: https://www.who.int/publications/m/item/draft-landscape-of-covid-19-candidate-vaccines (accessed on 5 January 2021).
- Lee, S.-H.; Kang, H.-J.; Lee, D.-H.; Kang, S.-M.; Quan, F.-S. Virus-like particle vaccines expressing Toxoplasma gondii rhoptry protein 18 and microneme protein 8 provide enhanced protection. Vaccine 2018, 36, 5692–5700. [Google Scholar] [CrossRef]
- Choi, W.H.; Park, J.S. Immunogenicity and Protective Effect of a Virus-Like Particle Containing the SAG1 Antigen of Toxoplasma gondii as a Potential Vaccine Candidate for Toxoplasmosis. Biomedicines 2020, 8, 91. [Google Scholar] [CrossRef]
- Chichester, J.A.; Green, B.J.; Jones, R.M.; Shoji, Y.; Miura, K.; Long, C.A.; Lee, C.K.; Ockenhouse, C.F.; Morin, M.J.; Streatfield, S.J.; et al. Safety and immunogenicity of a plant-produced Pfs25 virus-like particle as a transmission blocking vaccine against malaria: A Phase 1 dose-escalation study in healthy adults. Vaccine 2018, 36, 5865–5871. [Google Scholar] [CrossRef]
- Janitzek, C.M.; Peabody, J.; Thrane, S.; Carlsen, P.H.R.; Theander, T.G.; Salanti, A.; Chackerian, B.; Nielsen, M.A.; Sander, A.F. A proof-of-concept study for the design of a VLP-based combinatorial HPV and placental malaria vaccine. Sci. Rep. 2019, 9, 5260. [Google Scholar] [CrossRef]
- Yenkoidiok-Douti, L.; Williams, A.E.; Canepa, G.E.; Molina-Cruz, A.; Barillas-Mury, C. Engineering a Virus-Like Particle as an Antigenic Platform for a Pfs47-Targeted Malaria Transmission-Blocking Vaccine. Sci. Rep. 2019, 9, 16833–16839. [Google Scholar] [CrossRef]
- De Castro, M.C.A.B.; de Albuquerque, S.D.C.G.; Tavares, D.C.; De Paiva-Cavalcanti, M.; Pereira, V.A. Vaccines Against Trypanosomatids. In Tropical Diseases: An Overview of Major Diseases Occurring in the Americas; Bentham eBooks; Betham Science Publishers: Sharjah, United Arab Emirates, 2017; pp. 88–113. [Google Scholar]
- Michel-Todó, L.; Bigey, P.; Reche, P.; Pinazo, M.-J.; Gascón, J.; Alonso-Padilla, J. Design of an Epitope-Based Vaccine Ensemble for Animal Trypanosomiasis by Computational Methods. Vaccines 2020, 8, 130. [Google Scholar] [CrossRef]
- Oli, A.N.; Obialor, W.O.; Ifeanyichukwu, M.O.; Odimegwu, D.C.; Okoyeh, J.N.; Emechebe, G.O.; Adejumo, S.A.; Ibeanu, G.C. Immunoinformatics and Vaccine Development: An Overview. ImmunoTargets Ther. 2020, 9, 13–30. [Google Scholar] [CrossRef]
- Rios, L.E.; Vázquez-Chagoyán, J.C.; Pacheco, A.O.; Zago, M.P.; Garg, N.J. Immunity and vaccine development efforts against Trypanosoma cruzi. Acta Trop. 2019, 200, 105168. [Google Scholar] [CrossRef] [PubMed]
- Dumonteil, E.; Herrera, C.; Tu, W.; Goff, K.; Fahlberg, M.; Haupt, E.; Kaur, A.; Marx, P.A.; Ortega-Lopez, J.; Hotez, P.J.; et al. Safety and immunogenicity of a recombinant vaccine against Trypanosoma cruzi in Rhesus macaques. Vaccine 2020, 38, 4584–4591. [Google Scholar] [CrossRef] [PubMed]
- Bivona, A.E.; Alberti, A.S.; Cerny, N.; Trinitario, S.N.; Malchiodi, E.L. Chagas disease vaccine design: The search for an efficient Trypanosoma cruzi immune-mediated control. Biochim. Biophys. Acta Mol. Basis Dis. 2020, 1866, 165658. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in humans: Drug or vaccine therapy? Drug Des. Dev. Ther. 2017, 12, 25–40. [Google Scholar] [CrossRef] [PubMed]
- Thomaz-Soccol, V.; Da Costa, E.S.F.; Karp, S.G.; Letti, L.A.J.; Soccol, C.R.; Soccol, F.T. Recent Advances in Vaccines Against Leishmania Based on Patent Applications. Recent Patents Biotechnol. 2017, 11, 21–32. [Google Scholar] [CrossRef]
- Black, S.J.; Mansfield, J.M. Prospects for vaccination against pathogenic African trypanosomes. Parasite Immunol. 2016, 38, 735–743. [Google Scholar] [CrossRef] [PubMed]
- Jackson, A.P. The Evolution of Amastin Surface Glycoproteins in Trypanosomatid Parasites. Mol. Biol. Evol. 2009, 27, 33–45. [Google Scholar] [CrossRef]
- Jackson, A.P. Gene family phylogeny and the evolution of parasite cell surfaces. Mol. Biochem. Parasitol. 2016, 209, 64–75. [Google Scholar] [CrossRef]
- Pech-Canul, Á.D.L.C.; Monteón, V.; Solís-Oviedo, R.-L. A Brief View of the Surface Membrane Proteins fromTrypanosoma cruzi. J. Parasitol. Res. 2017, 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Rafati, S.; Fasel, N.; Masina, S. Leishmania Cysteine Proteinases: From Gene to Subunit Vaccine. Curr. Genom. 2003, 4, 253–261. [Google Scholar] [CrossRef]
- Abaza, S.Z. Expression of cysteine proteinases and cystatins in parasites and use of cysteine proteinase inhibitors in parasitic diseases. Part III: Protozoa (3): Kinetoplastids. Parasitol. United J. 2019, 12, 163–186. [Google Scholar] [CrossRef]
- Soares, R.P.; Altoé, E.C.F.; Ennes-Vidal, V.; Da Costa, S.M.; Rangel, E.F.; De Souza, N.A.; Da Silva, V.C.; Volf, P.; D’Avila-Levy, C.M. In Vitro Inhibition of Leishmania Attachment to Sandfly Midguts and LL-5 Cells by Divalent Metal Chelators, Anti-gp63 and Phosphoglycans. Protist 2017, 168, 326–334. [Google Scholar] [CrossRef]
- Moreno, C.J.G.; Torres, T.; Silva, M.S. Variable Surface Glycoprotein from Trypanosoma brucei Undergoes Cleavage by Matrix Metalloproteinases: An in silico Approach. Pathogens 2019, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Parthasarathy, A.; Kalesh, K. Defeating the trypanosomatid trio: Proteomics of the protozoan parasites causing neglected tropical diseases. RSC Med. Chem. 2020, 11, 625–645. [Google Scholar] [CrossRef]
- Sarkar, B.; Islam, S.S.; Zohora, U.S.; Ullah, M.A. Virus like particles-A recent advancement in vaccine development. Korean J. Microbiol. 2019, 55, 327–343. [Google Scholar] [CrossRef]
- Silva-Barrios, S.; Charpentier, T.; Stäger, S. The Deadly Dance of B Cells with Trypanosomatids. Trends Parasitol. 2018, 34, 155–171. [Google Scholar] [CrossRef]
- Silva-Barrios, S.; Stäger, S. Hypergammaglobulinemia sustains the development of regulatory responses during chronic Leishmania donovani infection in mice. Eur. J. Immunol. 2019, 49, 1082–1091. [Google Scholar] [CrossRef]
- Fernández-Presas, A.M.; Padilla-Noriega, L.; Robert, L.; Jiménez, J.A.; Solano, S.; Delgado, J.; Tato, P.; Molinari, J.L.; Becker, I. Enveloped and non-enveloped viral-like particles in Trypanosoma cruzi epimastigotes. Rev. Inst. Med. Trop. São Paulo 2017, 59. [Google Scholar] [CrossRef]
- Moura, A.P.V.; Santos, L.C.B.; Brito, C.R.N.; Valencia, E.; Junqueira, C.; Filho, A.A.P.; Sant’Anna, M.R.V.; Gontijo, N.F.; Bartholomeu, D.C.; Fujiwara, R.T.; et al. Virus-like Particle Display of the α-Gal Carbohydrate for Vaccination againstLeishmaniaInfection. ACS Cent. Sci. 2017, 3, 1026–1031. [Google Scholar] [CrossRef] [PubMed]
- Cecilio, P.; Pérez-Cabezas, B.; Fernandez, L.; Moreno, J.; Carrillo, E.; Requena, J.M.; Fichera, E.; Reed, S.G.; Coler, R.N.; Kamhawi, S.; et al. Pre-clinical antigenicity studies of an innovative multivalent vaccine for human visceral leishmaniasis. PLoS Negl. Trop. Dis. 2017, 11, e0005951. [Google Scholar] [CrossRef]
- Chackerian, B. Virus-like particles: Flexible platforms for vaccine development. Expert Rev. Vaccines 2007, 6, 381–390. [Google Scholar] [CrossRef]
- Deml, L.; Speth, C.; Dierich, M.P.; Wolf, H.; Wagner, R. Recombinant HIV-1 Pr55gag virus-like particles: Potent stimulators of innate and acquired immune responses. Mol. Immunol. 2005, 42, 259–277. [Google Scholar] [CrossRef]
- Pavanelli, W.R. Citocinas e óxido nítrico na Doença de Chagas. Biosaúde 2016, 10, 147–156. [Google Scholar]
- Zlotnick, A.; Mukhopadhyay, S. Virus assembly, allostery and antivirals. Trends Microbiol. 2011, 19, 14–23. [Google Scholar] [CrossRef] [PubMed]
- Visciano, M.; Diomede, L.; Tagliamonte, M.; Tornesello, M.L.; Asti, V.; Bomsel, M.; Buonaguro, F.; Lopalco, L.; Buonaguro, L. Generation of HIV-1 Virus-Like Particles expressing different HIV-1 glycoproteins. Vaccine 2011, 29, 4903–4912. [Google Scholar] [CrossRef] [PubMed]
- Naskalska, A.; Pyrć, K. Virus like particles as immunogens and universal nanocarriers. Pol. J. Microbiol. 2015, 64, 3–13. [Google Scholar] [CrossRef]
- Hervas-Stubbs, S.; Rueda, P.; Lopez, L.; Leclerc, C. Insect Baculoviruses Strongly Potentiate Adaptive Immune Responses by Inducing Type I IFN. J. Immunol. 2007, 178, 2361–2369. [Google Scholar] [CrossRef]
- Balke, I.; Zeltins, A. Recent Advances in the Use of Plant Virus-Like Particles as Vaccines. Viruses 2020, 12, 270. [Google Scholar] [CrossRef] [PubMed]
- Roy, P.; Noad, R. Virus-Like Particles as a Vaccine Delivery System: Myths and Facts. In Advances in Experimental Medicine and Biology; Springer: Berlin, Germany, 2009; Volume 655, pp. 145–158. [Google Scholar]
- Jeong, H.; Seong, B.L. Exploiting virus-like particles as innovative vaccines against emerging viral infections. J. Microbiol. 2017, 55, 220–230. [Google Scholar] [CrossRef] [PubMed]
- Perotti, M.; Perez, L. Virus-Like Particles and Nanoparticles for Vaccine Development against HCMV. Viruses 2019, 12, 35. [Google Scholar] [CrossRef]
- Jain, N.K.; Sahni, N.; Kumru, O.S.; Joshi, S.B.; Volkin, D.B.; Middaugh, C.R. Formulation and stabilization of recombinant protein based virus-like particle vaccines. Adv. Drug Deliv. Rev. 2015, 93, 42–55. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Morales, O.; Monteón-Padilla, V.; Carrillo-Sánchez, S.C.; Rios-Castro, M.; Martínez-Cruz, M.; Carabarin-Lima, A.; Arce-Fonseca, M. Experimental Vaccines against Chagas Disease: A Journey through History. J. Immunol. Res. 2015, 2015, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Beaumier, C.M.; Gillespie, P.M.; Strych, U.; Hayward, T.; Hotez, P.J.; Bottazzi, M.E. Status of vaccine research and development of vaccines for Chagas disease. Vaccine 2016, 34, 2996–3000. [Google Scholar] [CrossRef] [PubMed]
- Brito, L.A.; Singh, M. COMMENTARY: Acceptable Levels of Endotoxin in Vaccine Formulations During Preclinical Research. J. Pharm. Sci. 2011, 100, 34–37. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. WHO guidelines on nonclinical evaluation of vaccines. World Health Organ. Tech. Rep. Ser. 2005, 927, 55. [Google Scholar]
- Yan, D.; Wei, Y.-Q.; Guo, H.-C.; Sun, S.-Q. The application of virus-like particles as vaccines and biological vehicles. Appl. Microbiol. Biotechnol. 2015, 99, 10415–10432. [Google Scholar] [CrossRef] [PubMed]
- Frietze, K.M.; Peabody, D.S.; Chackerian, B. Engineering virus-like particles as vaccine platforms. Curr. Opin. Virol. 2016, 18, 44–49. [Google Scholar] [CrossRef]
- Serradell, M.C.; Rupil, L.L.; Martino, R.A.; Prucca, C.G.; Carranza, P.G.; Saura, A.; Fernández, E.A.; Gargantini, P.R.; Tenaglia, A.H.; Petiti, J.P.; et al. Efficient oral vaccination by bioengineering virus-like particles with protozoan surface proteins. Nat. Commun. 2019, 10, 361. [Google Scholar] [CrossRef]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Queiroz, A.M.V.; Oliveira, J.W.d.F.; Moreno, C.J.; Guérin, D.M.A.; Silva, M.S. VLP-Based Vaccines as a Suitable Technology to Target Trypanosomatid Diseases. Vaccines 2021, 9, 220. https://doi.org/10.3390/vaccines9030220
Queiroz AMV, Oliveira JWdF, Moreno CJ, Guérin DMA, Silva MS. VLP-Based Vaccines as a Suitable Technology to Target Trypanosomatid Diseases. Vaccines. 2021; 9(3):220. https://doi.org/10.3390/vaccines9030220
Chicago/Turabian StyleQueiroz, Aline Maria Vasconcelos, Johny Wysllas de Freitas Oliveira, Cláudia Jassica Moreno, Diego M. A. Guérin, and Marcelo Sousa Silva. 2021. "VLP-Based Vaccines as a Suitable Technology to Target Trypanosomatid Diseases" Vaccines 9, no. 3: 220. https://doi.org/10.3390/vaccines9030220
APA StyleQueiroz, A. M. V., Oliveira, J. W. d. F., Moreno, C. J., Guérin, D. M. A., & Silva, M. S. (2021). VLP-Based Vaccines as a Suitable Technology to Target Trypanosomatid Diseases. Vaccines, 9(3), 220. https://doi.org/10.3390/vaccines9030220