
Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus

 , and
, and
Abstract
1. Introduction
2. Materials and Methods
2.1. Porvac® Subunit Vaccine E2-CD154
2.2. Experimental Animals
2.3. Immunization Schedule and Experimental Design
2.4. Neutralizing Antibodies Detection
2.5. Viral Isolation
2.6. Elispot Assay for Detection of CSFV-Specific IFN-γ Producing Cells
2.7. Statistical Analysis
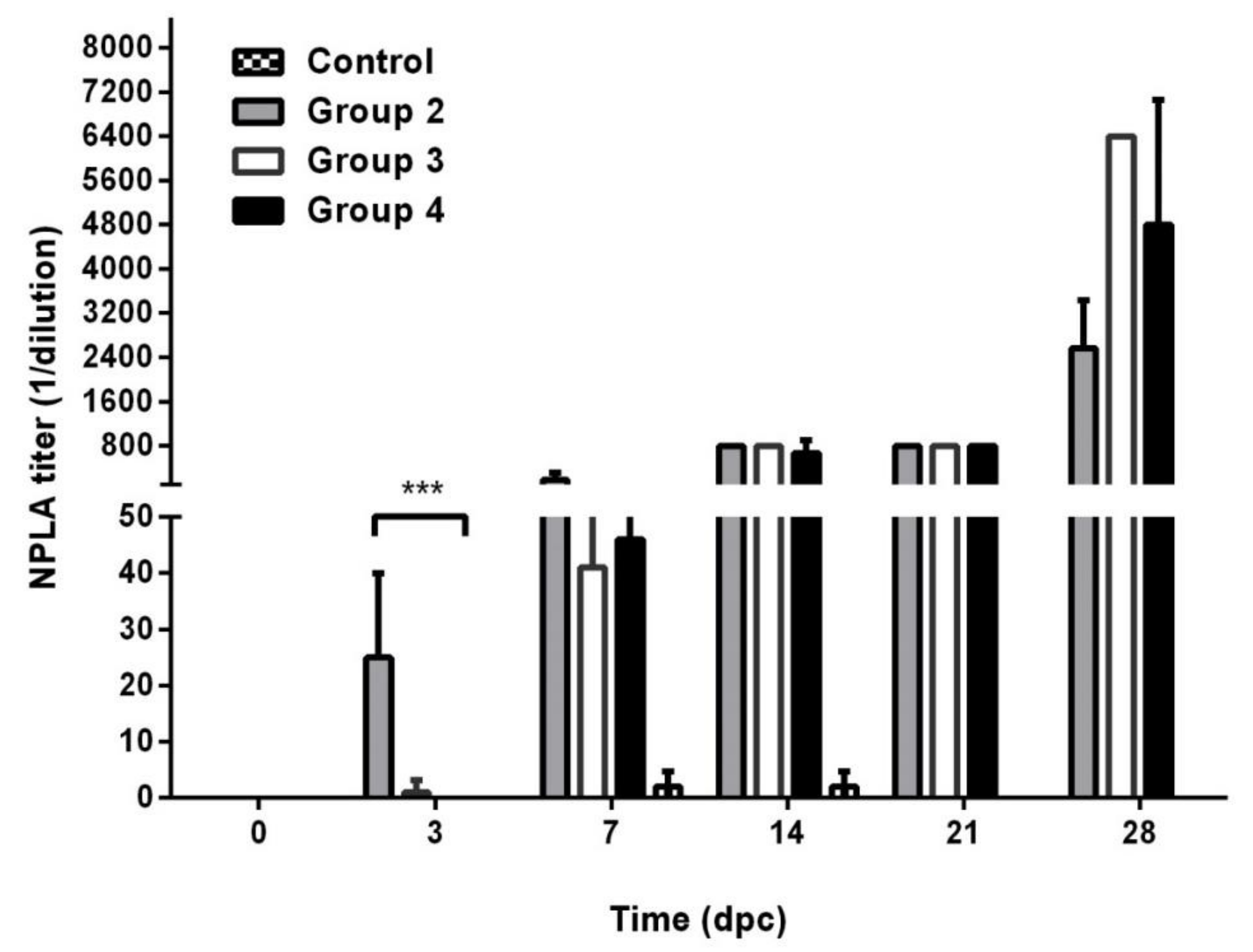
3. Results
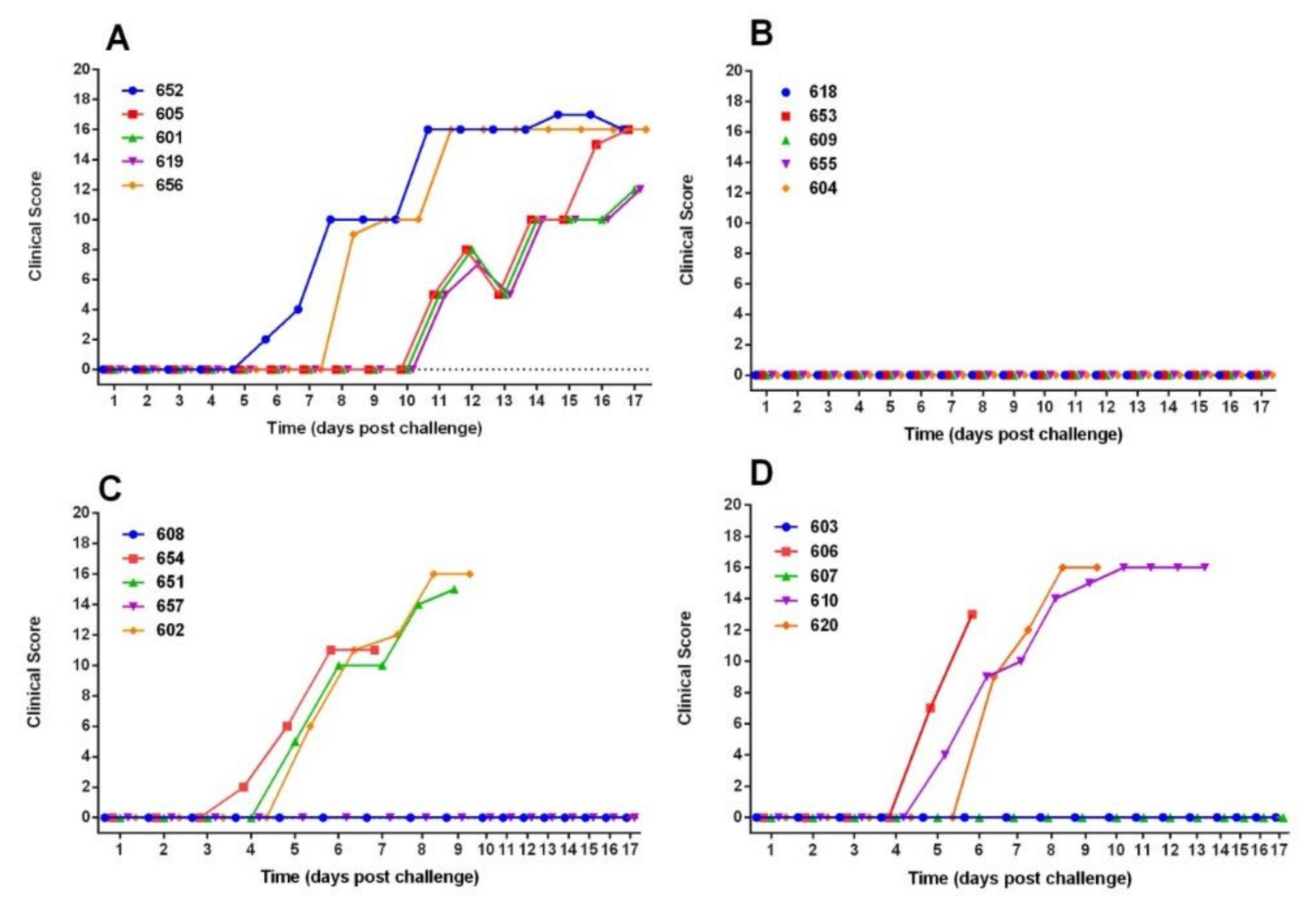
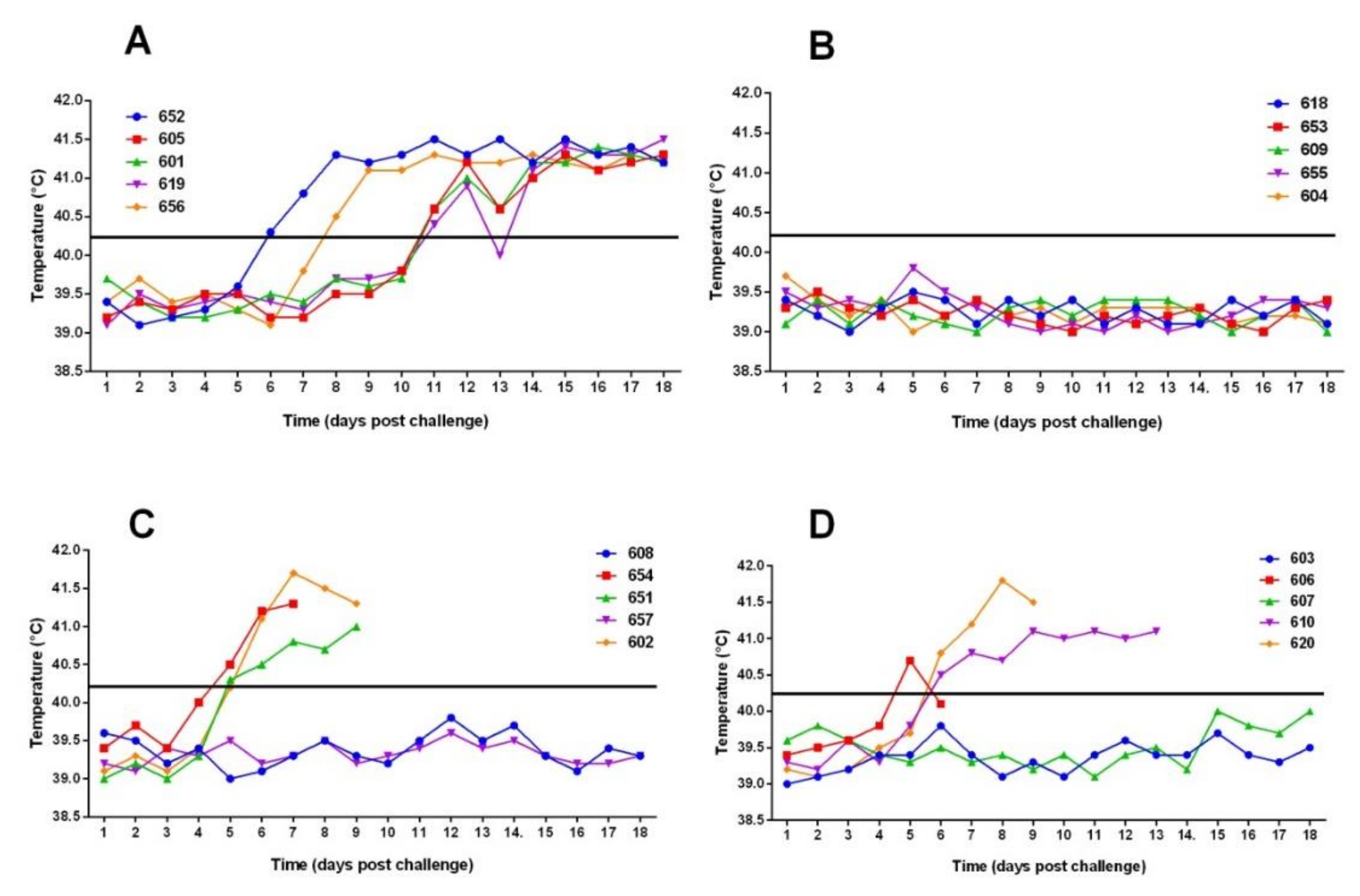
3.1. Vaccination Five Days Prior to Challenge Protected Animals Against Clinical Disease and Prevented Infection
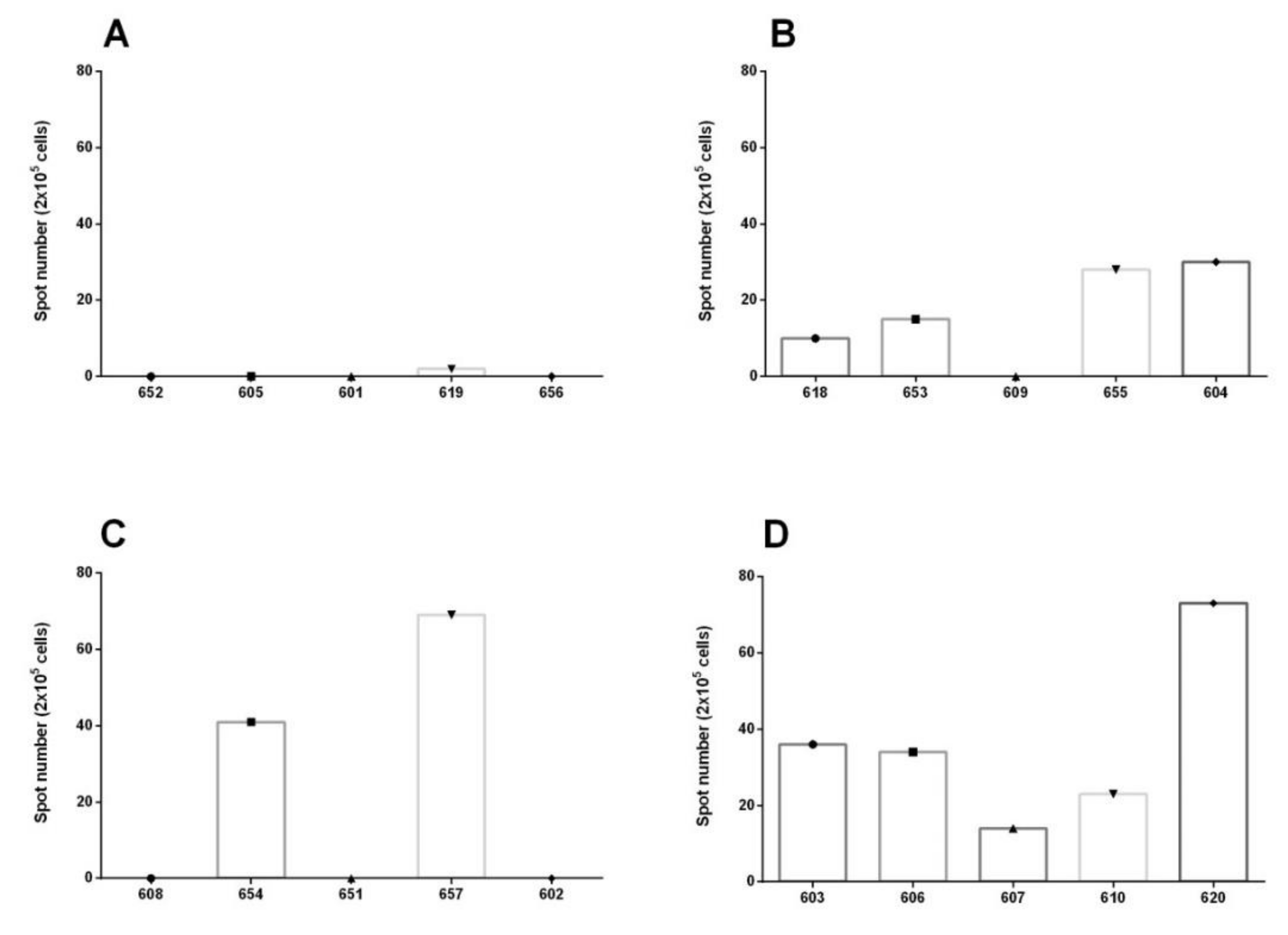
3.2. IFN Gamma Determination
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Edwards, S.; Fukusho, A.; Lefèvre, P.-C.; Lipowski, A.; Pejsak, Z.; Roehe, P.; Westergaard, J. Classical swine fever: The global situation. Vet. Microbiol. 2000, 73, 103–119. [Google Scholar] [CrossRef]
- Moennig, V.; Floegel-Niesmann, G.; Greiser-Wilke, I. Clinical Signs and Epidemiology of Classical Swine Fever: A Review of New Knowledge. Vet. J. 2003, 165, 11–20. [Google Scholar] [CrossRef]
- Dong, X.-N.; Chen, Y.-H. Marker vaccine strategies and candidate CSFV marker vaccines. Vaccine 2007, 25, 205–230. [Google Scholar] [CrossRef]
- Moennig, V. Introduction to classical swine fever: Virus, disease and control policy. Vet. Microbiol. 2000, 73, 93–102. [Google Scholar] [CrossRef]
- Ganges, L.; Crooke, H.R.; Bohórquez, J.A.; Postel, A.; Sakoda, Y.; Becher, P.; Ruggli, N. Classical swine fever virus: The past, present and future. Virus Res. 2020, 289, 198151. [Google Scholar] [CrossRef]
- Wang, F.-I.; Chang, C.-Y. Classical Swine Fever: A Truly Classical Swine Disease. Pathogens 2020, 9, 745. [Google Scholar] [CrossRef]
- Suradhat, S.; Intrakamhaeng, M.; Damrongwatanapokin, S. The correlation of virus-specific interferon-gamma production and protection against classical swine fever virus infection. Vet. Immunol. Immunopathol. 2001, 83, 177–189. [Google Scholar] [CrossRef]
- Graham, S.P.; Everett, H.E.; Johns, H.L.; Haines, F.J.; La Rocca, S.A.; Khatri, M.; Wright, I.K.; Drew, T.; Crooke, H.R. Characterisation of virus-specific peripheral blood cell cytokine responses following vaccination or infection with classical swine fever viruses. Vet. Microbiol. 2010, 142, 34–40. [Google Scholar] [CrossRef]
- Graham, S.P.; Everett, H.E.; Haines, F.J.; Johns, H.L.; Sosan, O.A.; Salguero, F.J.; Clifford, D.J.; Steinbach, F.; Drew, T.W.; Crooke, H.R. Challenge of Pigs with Classical Swine Fever Viruses after C-Strain Vaccination Reveals Remarkably Rapid Protection and Insights into Early Immunity. PLoS ONE 2012, 7, e29310. [Google Scholar] [CrossRef]
- Van Oirschot, J. Vaccinology of classical swine fever: From lab to field. Vet. Microbiol. 2003, 96, 367–384. [Google Scholar] [CrossRef] [PubMed]
- Blome, S.; Staubach, C.; Henke, J.; Carlson, J.; Beer, M. Classical swine fever—An updated review. Viruses 2017, 9, 86. [Google Scholar] [CrossRef] [PubMed]
- Suárez, M.; Sordo, Y.; Prieto, Y.; Rodríguez, M.P.; Méndez, L.; Rodríguez, E.M.; Rodríguez-Mallon, A.; Lorenzo, E.; Santana, E.; González, N.; et al. A single dose of the novel chimeric subunit vaccine E2-CD154 confers early full protection against classical swine fever virus. Vaccine 2017, 35, 4437–4443. [Google Scholar] [CrossRef]
- Muñoz-González, S.; Sordo, Y.; Pérez-Simó, M.; Suárez, M.; Canturri, A.; Rodriguez, M.P.; Frías-Lepoureau, M.T.; Domingo, M.; Estrada, M.P.; Ganges, L. Efficacy of E2 glycoprotein fused to porcine CD154 as a novel chimeric subunit vaccine to prevent classical swine fever virus vertical transmission in pregnant sows. Vet. Microbiol. 2017, 205, 110–116. [Google Scholar] [CrossRef]
- Postel, A.; Schmeiser, S.; Perera, C.L.; Rodríguez, L.J.P.; Frias-Lepoureau, M.T.; Becher, P. Classical swine fever virus isolates from Cuba form a new subgenotype 1.4. Vet. Microbiol. 2013, 161, 334–338. [Google Scholar] [CrossRef]
- Mittelholzer, C.; Moser, C.; Tratschin, J.-D.; Hofmann, M.A. Analysis of classical swine fever virus replication kinetics allows differentiation of highly virulent from avirulent strains. Vet. Microbiol. 2000, 74, 293–308. [Google Scholar] [CrossRef]
- Terpstra, C.; Bloemraad, M.; Gielkens, A. The neutralizing peroxidase-linked assay for detection of antibody against swine fever virus. Vet. Microbiol. 1984, 9, 113–120. [Google Scholar] [CrossRef]
- OIE. OIE Terrestrial Manual. Chapter 2.8.3. Classical Swine Fever (Hog Cholera) Infection with Classical Swine Fever Virus; OIE, Ed.; World Organisation for Animal Health (OIE): Paris, France, 2014. [Google Scholar]
- Dewulf, J.; Laevens, H.; Koenen, F.; Mintiens, K.; De Kruif, A. An experimental infection with classical swine fever virus in pregnant sows: Transmission of the virus, course of the disease, antibody response and effect on gestation. J. Vet. Med. Ser. B 2001, 48, 583–591. [Google Scholar] [CrossRef] [PubMed]
- Ganges, L.; Núñez, J.I.; Sobrino, F.; Borrego, B.; Fernández-Borges, N.; Frías-Lepoureau, M.T.; Rodríguez, F. Recent advances in the development of recombinant vaccines against classical swine fever virus: Cellular responses also play a role in protection. Vet. J. 2008, 177, 169–177. [Google Scholar] [CrossRef]
- Biront, P.; Leunen, J.; Vandeputte, J. Inhibition of virus replication in the tonsils of pigs previously vaccinated with a Chinese strain vaccine and challenged oronasally with a virulent strain of classical swine fever virus. Vet. Microbiol. 1987, 14, 105–113. [Google Scholar] [CrossRef]
- De Smit, A.J.; Bouma, A.; De Kluijver, E.P.; Terpstra, C.; Moormann, R. Duration of the protection of an E2 subunit marker vaccine against classical swine fever after a single vaccination. Vet. Microbiol. 2001, 78, 307–317. [Google Scholar] [CrossRef]
- Leifer, I.; Lange, E.; Reimann, I.; Blome, S.; Juanola, S.; Duran, J.P.; Beer, M. Modified live marker vaccine candidate CP7_E2alf provides early onset of protection against lethal challenge infection with classical swine fever virus after both intramuscular and oral immunization. Vaccine 2009, 27, 6522–6529. [Google Scholar] [CrossRef]
- Frese, M.; Schwärzle, V.; Barth, K.; Krieger, N.; Lohmann, V.; Mihm, S.; Haller, O.; Bartenschlager, R. Interferon-γ inhibits replication of subgenomic and genomic hepatitis C virus RNAs. Hepatology 2002, 35, 694–703. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Chen, X.C.; Konduri, M.; Fomina, N.; Lu, J.; Jin, L.; Kolykhalov, A.; Tan, S.-L. Mechanistic link between the anti-HCV effect of interferon gamma and control of viral replication by a ras-MAPK signaling cascade. Hepatology 2006, 43, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Kanda, T.; Steele, R.; Ray, R.; Ray, R.B. Inhibition of Intrahepatic Gamma Interferon Production by Hepatitis C Virus Nonstructural Protein 5A in Transgenic Mice. J. Virol. 2009, 83, 8463–8469. [Google Scholar] [CrossRef]
- Segundo, F.D.-S.; Moraes, M.P.; Santos, T.D.L.; Dias, C.C.A.; Grubman, M.J. Interferon-Induced Protection against Foot-and-Mouth Disease Virus Infection Correlates with Enhanced Tissue-Specific Innate Immune Cell Infiltration and Interferon-Stimulated Gene Expression. J. Virol. 2010, 84, 2063–2077. [Google Scholar] [CrossRef]
- Fernandez-Sainz, I.; Ramanathan, P.; O’Donnell, V.; Segundo, F.D.-S.; Velazquez-Salinas, L.; Sturza, D.; Zhu, J.; Santos, T.D.L.; Borca, M. Treatment with interferon-alpha delays disease in swine infected with a highly virulent CSFV strain. Virology 2015, 483, 284–290. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bouma, A.; De Smit, A.; De Jong, M.; De Kluijver, E.; Moormann, R. Determination of the onset of the herd-immunity induced by the E2 sub-unit vaccine against classical swine fever virus. Vaccine 2000, 18, 1374–1381. [Google Scholar] [CrossRef]
- Graham, S.P.; Haines, F.J.; Johns, H.L.; Sosan, O.; La Rocca, S.A.; Lamp, B.; Rümenapf, T.; Everett, H.E.; Crooke, H.R. Characterisation of vaccine-induced, broadly cross-reactive IFN-γ secreting T cell responses that correlate with rapid protection against classical swine fever virus. Vaccine 2012, 30, 2742–2748. [Google Scholar] [CrossRef]
- Lieberman, L.A.; Hunter, C.A. Regulatory pathways involved in the infection-induced production of IFN-γ by NK cells. Microbes Infect. 2002, 4, 1531–1538. [Google Scholar] [CrossRef]
- Neves, P.C.C.; Santos, J.R.; Tubarão, L.N.; Bonaldo, M.C.; Galler, R. Early IFN-Gamma Production after YF 17D Vaccine Virus Immunization in Mice and Its Association with Adaptive Immune Responses. PLoS ONE 2013, 8, e81953. [Google Scholar] [CrossRef]
- Gares, S.L.; Fischer, K.P.; Congly, S.E.; Lacoste, S.; Addison, W.R.; Tyrrell, D.L.; Gutfreund, K.S. Immunotargeting with CD154 (CD40 Ligand) Enhances DNA Vaccine Responses in Ducks. Clin. Vaccine Immunol. 2006, 13, 958–965. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Ramos, O.S.; Pose, A.G.; Gómez-Puerta, S.; Gomez, J.N.; Redondo, A.V.; Benites, J.C.Á.; Amarán, L.S.; Parra, N.C.; Alonso, J.R.T. Avian CD154 enhances humoral and cellular immune responses induced by an adenovirus vector-based vaccine in chickens. Comp. Immunol. Microbiol. Infect. Dis. 2011, 34, 259–265. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Song, G. The role of CD40-CD154 interaction in cell immunoregulation. J. Biomed. Sci. 2004, 11, 426–438. [Google Scholar] [CrossRef] [PubMed]
- Tregaskes, C.A.; Glansbeek, H.L.; Gill, A.C.; Hunt, L.G.; Burnside, J.; Young, J.R. Conservation of biological properties of the CD40 ligand, CD154 in a non-mammalian vertebrate. Dev. Comp. Immunol. 2005, 29, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, X.; Du, Y.; Li, Y.; Wang, X.; Jiang, P. CD40 ligand expressed in adenovirus can improve the immunogenicity of the GP3 and GP5 of porcine reproductive and respiratory syndrome virus in swine. Vaccine 2010, 28, 7514–7522. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Blood | Spleenen | Tonsil | Ileum | Rectal Swab | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Group | Animals | 3 dpc | 7 dips | 14 dpc | 21 dpc | 28 dpc | At sacrifice | 21 dpc | 28 dpc | ||
| 1 | 652 | - | + | - | N/S | N/S | + | + | + | N/S | N/S |
| 605 | - | - | + | N/S | N/S | + | + | + | N/S | N/S | |
| 601 | - | - | + | N/S | N/S | + | + | + | N/S | N/S | |
| 619 | - | - | + | N/S | N/S | + | + | + | N/S | N/S | |
| 656 | - | - | + | N/S | N/S | N/S | N/S | N/S | N/S | N/S | |
| 2 | 618 | - | - | - | - | - | - | - | - | - | - |
| 653 | - | - | - | - | - | - | - | - | - | - | |
| 609 | - | - | - | - | - | - | - | - | - | - | |
| 655 | + | - | - | - | - | - | - | - | - | - | |
| 604 | - | - | - | - | - | - | - | - | - | - | |
| 3 | 608 | - | - | - | - | - | - | - | - | - | - |
| 654 | + | + | N/S | N/S | N/S | + | + | - | N/S | N/S | |
| 651 | - | - | N/S | N/S | N/S | + | + | - | N/S | N/S | |
| 657 | - | - | - | - | - | - | - | - | - | - | |
| 602 | - | - | N/S | N/S | N/S | + | + | - | N/S | N/S | |
| 4 | 603 | - | - | - | - | - | - | - | - | - | - |
| 606 | + | + | N/S | N/S | N/S | + | + | - | N/S | N/S | |
| 607 | - | - | - | - | - | - | - | + | - | - | |
| 610 | - | + | + | N/S | N/S | + | + | - | N/S | N/S | |
| 620 | - | - | N/S | N/S | N/S | + | + | - | N/S | N/S | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sordo-Puga, Y.; Suárez-Pedroso, M.; Naranjo-Valdéz, P.; Pérez-Pérez, D.; Santana-Rodríguez, E.; Sardinas-Gonzalez, T.; Mendez-Orta, M.K.; Duarte-Cano, C.A.; Estrada-Garcia, M.P.; Rodríguez-Moltó, M.P. Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus. Vaccines 2021, 9, 167. https://doi.org/10.3390/vaccines9020167
Sordo-Puga Y, Suárez-Pedroso M, Naranjo-Valdéz P, Pérez-Pérez D, Santana-Rodríguez E, Sardinas-Gonzalez T, Mendez-Orta MK, Duarte-Cano CA, Estrada-Garcia MP, Rodríguez-Moltó MP. Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus. Vaccines. 2021; 9(2):167. https://doi.org/10.3390/vaccines9020167
Chicago/Turabian StyleSordo-Puga, Yusmel, Marisela Suárez-Pedroso, Paula Naranjo-Valdéz, Danny Pérez-Pérez, Elaine Santana-Rodríguez, Talia Sardinas-Gonzalez, Mary Karla Mendez-Orta, Carlos A. Duarte-Cano, Mario Pablo Estrada-Garcia, and María Pilar Rodríguez-Moltó. 2021. "Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus" Vaccines 9, no. 2: 167. https://doi.org/10.3390/vaccines9020167
APA StyleSordo-Puga, Y., Suárez-Pedroso, M., Naranjo-Valdéz, P., Pérez-Pérez, D., Santana-Rodríguez, E., Sardinas-Gonzalez, T., Mendez-Orta, M. K., Duarte-Cano, C. A., Estrada-Garcia, M. P., & Rodríguez-Moltó, M. P. (2021). Porvac® Subunit Vaccine E2-CD154 Induces Remarkable Rapid Protection against Classical Swine Fever Virus. Vaccines, 9(2), 167. https://doi.org/10.3390/vaccines9020167