
Innovations in the Insect Cell Expression System for Industrial Recombinant Vaccine Antigen Production

Abstract
:1. Introduction
2. Baculovirus AcNPV Backbone and Transfer Vector Development
3. Cell Line Engineering
4. Insect Cell Culture
5. Downstream Processing
6. Conclusions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Van Oers, M.M. Vaccines for viral and parasitic diseases produced with baculovirus vectors. Adv. Virus Res. 2006, 68, 193–253. [Google Scholar]
- Cox, M.M.J. Recombinant protein vaccines produced in insect cells. Vaccine 2012, 30, 1759–1766. [Google Scholar] [CrossRef]
- Miller, L.K. A virus vector for genetic engineering in invertebrates. In Genetic Engineering in the Plant Sciences; Panopaulus, N.J., Ed.; Praeger: New York, NY, USA, 1981; pp. 203–222. [Google Scholar]
- Palomares, L.A.; Srivastava, I.K.; Ramírez, O.T.; Cox, M.M.J. Glycobiotechnology of the insect cell-baculovirus expression system technology. Adv. Biochem. Eng. Biotechnol. 2021, 175, 71–92. [Google Scholar]
- Van Oers, M.M.; Pijlman, G.P.; Vlak, J.M. Thirty years of baculovirus-insect cell protein expression: From dark horse to mainstream technology. J. Gen. Virol. 2015, 96, 6–23. [Google Scholar] [CrossRef] [PubMed]
- Ng, W.H.; Liu, X.; Mahalingam, S. Development of vaccines for SARS-CoV-2. F1000 Res. 2020, 9, 991. [Google Scholar] [CrossRef]
- Fabre, M.L.; Arrías, P.N.; Masson, T.; Pidre, M.L.; Romanowski, V. Baculovirus-derived vectors for immunization and therapeutic applications. Emerg. Reemerg. Viral Pathog. 2020, 197–224. [Google Scholar] [CrossRef]
- Smith, G.E.; Summer, M.D.; Fraser, M.J. Production of human beta interferon in insect cells infected with a baculovirus expression vector. Mol. Cell Biol. 1983, 3, 2156–2165. [Google Scholar]
- Cox, M.M.; Hashimoto, Y. A fast-track influenza virus vaccine produced in insect cells. J. Invertebr. Pathol. 2011, 107, S31–S41. [Google Scholar] [CrossRef] [PubMed]
- Kitts, P.A.; Possee, R.D. A method for producing recombinant baculovirus expression vectors at high frequency. Biotechniques 1993, 14, 810–817. [Google Scholar] [PubMed]
- Kool, M.; Voncken, J.W.; van Lier, F.L.; Tramper, J.; Vlak, J.M. Detection and analysis of Autographa californica nuclear polyhedrosis virus mutants with defective interfering properties. Virology 1991, 183, 739–746. [Google Scholar] [CrossRef]
- Pijlman, G.P.; van den Born, E.; Martens, D.E.; Vlak, J.M. Autographa californica baculoviruses with large genomic deletions are rapidly generated in infected insect cells. Virology 2001, 283, 132–138. [Google Scholar] [CrossRef] [Green Version]
- Pijlman, G.P.; van Schijndel, J.E.; Vlak, J.M. Spontaneous excision of BAC vector sequences from bacmid-derived baculovirus expression vectors upon passage in insect cells. J. Gen. Virol. 2003, 84, 2669–2678. [Google Scholar] [CrossRef]
- Granados, R.R. Trichoplusia ni Cell Line which Supports Replication of Baculoviruses. U.S. Patent 5,300,435, 5 April 1994. [Google Scholar]
- Jehle, J.A.; Blissard, G.W.; Bonning, B.C.; Cory, J.S.; Herniou, E.A.; Rohrmann, G.F.; Theilmann, D.A.; Thiem, S.M.; Vlak, J.M. On the classification and nomenclature of baculoviruses: A proposal for revision. Arch. Virol. 2006, 151, 1257–1266. [Google Scholar] [CrossRef]
- Ikonomou, L.; Schneider, Y.J.; Agathos, S.N. Insect cell culture for industrial production of recombinant proteins. Appl. Microbiol. Biotechnol. 2003, 62, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Roldão, A.; Vicente, T.; Cox, M.; Alves, P.M.; Carrondo, M.J.T. Insect cell culture. In Industrial Scale Suspension Culture of Living Cells; Meyer, H.-P., Schmidhalter, D., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2014; pp. 348–389. [Google Scholar]
- Alldread, R.M.; Birch, J.R.; Metcalfe, H.K.; Farid, S.; Racher, A.J.; Young, R.J.; Kahn, M. Large scale suspension culture in mammalian cells. In Industrial Scale Suspension Culture of Living Cells; Meyer, H.-P., Schmidhalter, D., Eds.; John Wiley and Sons: Hoboken, NJ, USA, 2014; pp. 410–462. [Google Scholar]
- Huang, Y.M.; Hu, W.; Rustandi, E.; Chang, K.; Yusuf-Makagiansar, H.; Ryll, T. Maximizing productivity of CHO cell-based fed-batch culture using chemically defined media conditions and typical manufacturing equipment. Biotechnol. Prog. 2010, 26, 1400–1410. [Google Scholar] [CrossRef] [PubMed]
- Chavez-Pena, C.; Kamen, A.A. RNA interference technology to improve the baculovirus-insect cell expression system. Biotechnol. Adv. 2018, 36, 443–451. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, B.; Vidigal, J.; Correia, R.; Carrondo, M.J.T.; Alves, P.M.; Teixeira, A.P.; Roldão, A. Adaptive laboratory evolution of stable insect cell lines for improved HIV-Gag VLPs production. J. Biotechnol. 2020, 307, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Correia, R.; Fernandes, B.; Alves, P.M.; Carrondo, M.J.T.; Roldão, A. Improving influenza HA-Vlps production in insect high five cells via adaptive laboratory evolution. Vaccines 2020, 8, 589. [Google Scholar] [CrossRef] [PubMed]
- Heffner, K.M.; Wang, Q.; Deniz Baycin Hizal, D.B.; Özge Can, Ö.; Betenbaugh, M.J. Glycoengineering of mammalian expression systems on a cellular level. Adv. Biochem. Eng. Biotechnol. 2021, 175, 37–70. [Google Scholar]
- Possee, R.D.; Hitchman, R.B.; Richards, K.S.; Mann, S.G.; Siaterli, E.; Nixon, C.P.; Irving, H.; Assenberg, R.; Alderton, D.; Owens, R.J.; et al. Generation of baculovirus vectors for the high-throughput production of proteins in insect cells. Biotechnol. Bioeng. 2008, 101, 1115–1122. [Google Scholar] [CrossRef]
- Hitchman, R.B.; Possee, R.D.; Crombie, A.T.; Chambers, A.; Ho, K.; Siaterli, E.; Lissina, O.; Sternard, H.; Novy, R.; Loomis, K.; et al. Genetic modification of a baculovirus vector for increased expression in insect cells. Cell Biol. Toxicol. 2010, 26, 57–68. [Google Scholar] [CrossRef] [PubMed]
- Stolt-Bergner, P.; Benda, C.; Bergbrede, T.; Besir, H.; Celie, P.H.N.; Chang, C.; Drechsel, D.; Fischer, A.; Geerlof, A.; Giabbai, B.; et al. Baculovirus-driven protein expression in insect cells: A benchmarking study. J. Struct. Biol. 2018, 203, 71–80. [Google Scholar] [CrossRef] [Green Version]
- Kim, E.J.; Kramer, S.F.; Hebert, C.G.; Valdes, J.J.; Bentley, W.E. Metabolic engineering of the baculovirus-expression system via inverse “shotgun” genomic analysis and RNA interference (dsRNA) increases product yield and cell longevity. Biotechnol. Bioeng. 2007, 98, 645–654. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.S.; Lee, H.Y.; Kim, Y.J.; Jung, H.D.; Choi, K.J.; Yang, J.M.; Kim, S.S.; Kim, K. Small interfering (Si) RNA mediated baculovirus replication reduction without affecting target gene expression. Virus Res. 2015, 199, 68–76. [Google Scholar] [CrossRef]
- Schultz, K.L.; Friesen, P.D. Baculovirus DNA replication-specific expression factors trigger apoptosis and shutoff of host protein synthesis during infection. J. Virol. 2009, 83, 11123–11132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salem, T.Z.; Zhang, F.; Thiem, S.M. Reduced expression of Autographa californica nucleopolyhedrovirus ORF34, an essential gene, enhances heterologous gene expression. Virology 2013, 435, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Zhao, K.; Lan, L.; Shi, N.; Nan, H.; Shi, Y.; Xu, X.; Chen, H. Improvement of protein production by engineering a novel antiapoptotic baculovirus vector to suppress the expression of Sf-caspase-1 and Tn-caspase-1. Biotechnol. Bioeng. 2021, 118, 2977–2989. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Sebastián, S.; López-Vidal, J.; Escribano, J.M. Significant productivity improvement of the baculovirus expression vector system by engineering a novel expression cassette. PLoS ONE 2014, 9, e96562. [Google Scholar] [CrossRef]
- López-Vidal, J.; Gómez-Sebastián, S.; Bárcena, J.; Nuñez Mdel, C.; Martínez-Alonso, D.; Dudognon, B.; Guijarro, E.; Escribano, J.M. Improved production efficiency of virus-like particles by the baculovirus expression vector system. PLoS ONE 2015, 10, e0140039. [Google Scholar] [CrossRef]
- Steele, K.H.; Stone, B.J.; Franklin, K.M.; Fath-Goodin, A.; Zhang, X.; Jiang, H.; Webb, B.A.; Geisler, C. Improving the baculovirus expression vector system with vankyrin-enhanced technology. Biotechnol. Prog. 2017, 33, 1496–1507. [Google Scholar] [CrossRef]
- Grose, C.; Putman, Z.; Esposito, D. A review of alternative promoters for optimal recombinant protein expression in baculovirus-infected insect cells. Protein Expr. Purif. 2021, 186, 105924. [Google Scholar] [CrossRef]
- Manohar, S.L.; Kanamasa, S.; Nishina, T.; Kato, T.; Park, E.Y. Enhanced gene expression in insect cells and silkworm larva by modified polyhedrin promoter using repeated Burst sequence and very late transcriptional factor-1. Biotechnol. Bioeng. 2010, 107, 909–916. [Google Scholar] [CrossRef]
- Habib, S.; Pandey, S.; Chatterji, U.; Burma, S.; Ahmad, R.; Jain, A.; Hasnain, S.E. Bifunctionality of the AcMNPV homologous region sequence (hr1): Enhancer and ORI functions have different sequence requirements. DNA Cell Biol. 1996, 15, 737–747. [Google Scholar] [CrossRef] [PubMed]
- Gwak, W.-S.; Kim, H.-S.; Bae, J.-S.; Kim, T.-H.; Bae, S.M.; Woo, S.-D. Development of a novel enhanced baculovirus expression vector via promoter combination. J. Asia-Pacific Entomol. 2020, 23, 909–914. [Google Scholar] [CrossRef]
- Lou, Y.; Ji, G.; Liu, Q.; Wang, P.; Zhang, R.; Zhang, Y.; Liu, X. Secretory expression and scale-up production of recombinant human thyroid peroxidase via baculovirus/insect cell system in a wave-type bioreactor. Protein Expr. Purif. 2018, 149, 7–12. [Google Scholar] [CrossRef]
- Teng, C.Y.; van Oers, M.M.; Wu, T.Y. Additive effect of calreticulin and translation initiation factor eIF4E on secreted protein production in the baculovirus expression system. Appl. Microbiol. Biotechnol. 2013, 97, 8505–8516. [Google Scholar] [CrossRef] [PubMed]
- Mundhada, H.; Seoane, J.M.; Schneider, K.; Koza, A.; Christensen, H.B.; Klein, T.; Phaneuf, P.V.; Herrgard, M.; Feist, A.M.; Nielsen, A.T. Increased production of L-Serine in Escherichia coli through adaptive laboratory evolution. Metab. Eng. 2017, 39, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sunley, K.; Tharmalingam, T.; Butler, M. CHO cells adapted to hypothermic growth produce high yields of recombinant beta-interferon. Biotechnol. Prog. 2008, 24, 898–906. [Google Scholar] [CrossRef]
- Wagner, J.M.; Pajerowski, J.D.; Daniels, C.L.; McHugh, P.M.; Flynn, J.A.; Balliet, J.W.; Casimiro, D.R.; Subramanian, S. Enhanced production of Chikungunya virus-like particles using a high-pH adapted Spodoptera frugiperda insect cell line. PLoS ONE 2014, 9, e94401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hebert, C.G.; Valdes, J.J.; Bentley, W.E. In vitro and in vivo RNA interference mediated suppression of Tn-caspase-1 for improved recombinant protein production in High Five cell culture with the baculovirus expression vector system. Biotechnol. Bioeng. 2009, 104, 390–399. [Google Scholar] [CrossRef]
- Hebert, C.G.; Valdes, J.J.; Bentley, W.E. Investigating apoptosis: Characterization and analysis of Trichoplusia ni-caspase-1 through overexpression and RNAi mediated silencing. Insect Biochem. Mol. Biol. 2009, 39, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Lai, Y.K.; Hsu, J.T.; Chu, C.C.; Chang, T.Y.; Pan, K.L.; Lin, C.C. Enhanced recombinant protein production and differential expression of molecular chaperones in sf-caspase-1-repressed stable cells after baculovirus infection. BMC Biotechnol. 2012, 12, 83. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Zhou, Y.; Chen, K.; Ju, X. Suppression of Bm-Caspase-1 expression in BmN cells enhances recombinant protein production in a baculovirus expression vector system. Mol. Biotechnol. 2016, 58, 319–327. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.C.; Hebert, C.G.; Hung, C.W.; Quan, D.N.; Carter, K.K.; Bentley, W.E. Tuning cell cycle of insect cells for enhanced protein production. J. Biotechnol. 2013, 168, 55–61. [Google Scholar] [CrossRef]
- Pastor, A.R.; González-Domínguez, G.; Díaz-Salinas, M.A.; Ramírez, O.T.; Palomares, L.A. Defining the multiplicity and time of infection for the production of Zaire Ebola virus-like particles in the insect cell-baculovirus expression system. Vaccine 2019, 37, 6962–6969. [Google Scholar] [CrossRef]
- Reuveny, S.; Kim, Y.J.; Kemp, C.W.; Shiloach, J. Production of recombinant proteins in high-density insect cell cultures. Biotechnol. Bioeng. 1993, 42, 235–239. [Google Scholar] [CrossRef]
- Drugmand, J.C.; Schneider, Y.J.; Agathos, S.N. Insect cells as factories for biomanufacturing. Biotechnol. Adv. 2012, 30, 1140–1157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Konz, J.O.; King, J.; Cooney, C.L. Effects of oxygen on recombinant protein expression. Biotechnol. Prog. 1998, 14, 393–409. [Google Scholar] [CrossRef] [PubMed]
- Meghrous, J.; Mahmoud, W.; Jacob, D.; Chubet, R.; Cox, M.M.J.; Kamen, A. Development of a simple and high-yielding fed-batch process for the production of influenza vaccines. Vaccine 2009, 28, 309–316. [Google Scholar] [CrossRef] [PubMed]
- Birch, J.R.; Racher, A.J. Antibody production. Adv. Drug Deliv. Rev. 2006, 58, 671–685. [Google Scholar] [CrossRef]
- Mitchell-Logean, C.; Murhammer, D.W. Bioreactor headspace purging reduces dissolved carbon dioxide accumulation in insect cell cultures and enhances cell growth. Biotechnol. Prog. 1997, 13, 875–877. [Google Scholar] [CrossRef]
- Meghrous, J.; Khramtsov, N.; Buckland, B.C.; Cox, M.M.; Palomares, L.A.; Srivastava, I.K. Dissolved carbon dioxide determines the productivity of a recombinant hemagglutinin component of an influenza vaccine produced by insect cells. Biotechnol. Bioeng. 2015, 112, 2267–2275. [Google Scholar] [CrossRef] [PubMed]
- Butler, M. Serum and protein free media. In Animal Cell Culture, Cell Engineering; Al-Rubeai, M., Ed.; Springer: Dordrecht, The Netherlands, 2015; Volume 9, pp. 223–236. [Google Scholar]
- Bedard, C.; Tom, R.; Kamen, A. Growth, nutrient consumption, and endproduct accumulation in Sf-9 and Bti-Eaa insect-cell cultures—Insights into growth limitation and metabolism. Biotechnol. Prog. 1993, 9, 615–624. [Google Scholar] [CrossRef] [PubMed]
- Reuveny, S.; Kemp, C.W.; Eppstein, L.; Shiloach, J. Carbohydrate metabolism in insect cell cultures during cell growth and recombinant protein production. Ann. N. Y. Acad. Sci. 1992, 665, 230–237. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, R.; Jesionowski, G.; Ferrance, J.; Ataai, M.M. Insect cell physiology. Cytotechnology 1997, 24, 1–9. [Google Scholar] [CrossRef]
- Ohman, L.; Ljunggren, J.; Häggström, L. Induction of a metabolic switch in insect cells by substrate-limited fed batch cultures. Appl. Microbiol. Biotechnol. 1995, 43, 1006–1013. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, R.H. Replacement of vertebrate serum with lipids and other factors in the culture of invertebrate cells, tissues, parasites, and pathogens. Vitr. Cell. Dev. Biol. 1991, 27, 470–478. [Google Scholar] [CrossRef]
- Mitsuhashi, J. (Ed.) Nutritional Requirements of Insect Cells In Vitro, in Invertebrate Cell System Applications; CRC Press: Boca Raton, FL, USA, 1989; pp. 3–20. [Google Scholar]
- Papoutsakis, E.T. Media additives for protecting freely suspended animal cells against agitation and aeration damage. Trends Biotechnol. 1991, 9, 316–324. [Google Scholar] [CrossRef]
- Burky, J.E.; Wesson, M.C.; Young, A.; Farnsworth, S.; Dionne, B.; Zhu, Y.; Hartman, T.E.; Qu, L.; Zhou, W.; Sauer, P.W. Protein-free fed-batch culture of non-GS NS0 cell lines for production of recombinant antibodies. Biotechnol. Bioeng. 2007, 96, 281–293. [Google Scholar] [CrossRef]
- Spens, E.; Häggström, L. Defined protein and animal component-free NS0 fed-batch culture. Biotechnol. Bioeng. 2007, 98, 1183–1194. [Google Scholar] [CrossRef]
- Kurasawa, J.H.; Park, A.; Sowers, C.R.; Halpin, R.A.; Tovchigrechko, A.; Dobson, C.L.; Schmelzer, A.E.; Gao, C.; Wilson, S.D.; Ikeda, Y. Chemically defined, high-density insect cell-based expression system for scalable AAV vector production. Mol. Ther. Methods Clin. Dev. 2020, 19, 330–340. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Fan, Z.; Lin, Y.; Wang, T.Y. Serum-free medium for recombinant protein expression in Chinese hamster ovary cells. Front. Bioeng. Biotechnol. 2021, 9, 646363. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Z.; Sharfstein, S.T. Sodium butyrate stimulates monoclonal antibody over-expression in CHO cells by improving gene accessibility. Biotechnol. Bioeng. 2008, 100, 189–194. [Google Scholar] [CrossRef]
- Sung, Y.H.; Lee, G.M. Enhanced human thrombopoietin production by sodium butyrate addition to serum-free suspension culture of bcl-2-overexpressing CHO cells. Biotechnol. Prog. 2005, 21, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Yun, Z.; Takagi, M.; Yoshida, T. Effect of antioxidants on the apoptosis of CHO cells and production of tissue plasminogen activator in suspension culture. J. Biosci. Bioeng. 2001, 91, 581–585. [Google Scholar] [CrossRef]
- Spenger, A.; Ernst, W.; Condreay, J.P.; Kost, T.A.; Grabherr, R. Influence of promoter choice and trichostatin. A treatment on expression of baculovirus delivered genes in mammalian cells. Protein Expr. Purif. 2004, 38, 17–23. [Google Scholar] [CrossRef]
- Marek, M.; van Oers, M.M.; Devaraj, F.F.; Vlak, J.M.; Merten, O.W. Engineering of baculovirus vectors for the manufacture of virion-free biopharmaceuticals. Biotechnol. Bioeng. 2011, 108, 1056–1067. [Google Scholar] [CrossRef] [PubMed]
- Buckland, B.; Boulanger, R.; Fino, M.; Srivastava, I.; Holtz, K.; Khramtsov, N.; McPherson, C.; Meghrous, J.; Kubera, P.; Cox, M.M. Technology transfer and scale-up of the Flublok recombinant hemagglutinin (HA) influenza vaccine manufacturingprocess. Vaccine 2014, 42, 5496–5502. [Google Scholar] [CrossRef] [PubMed]
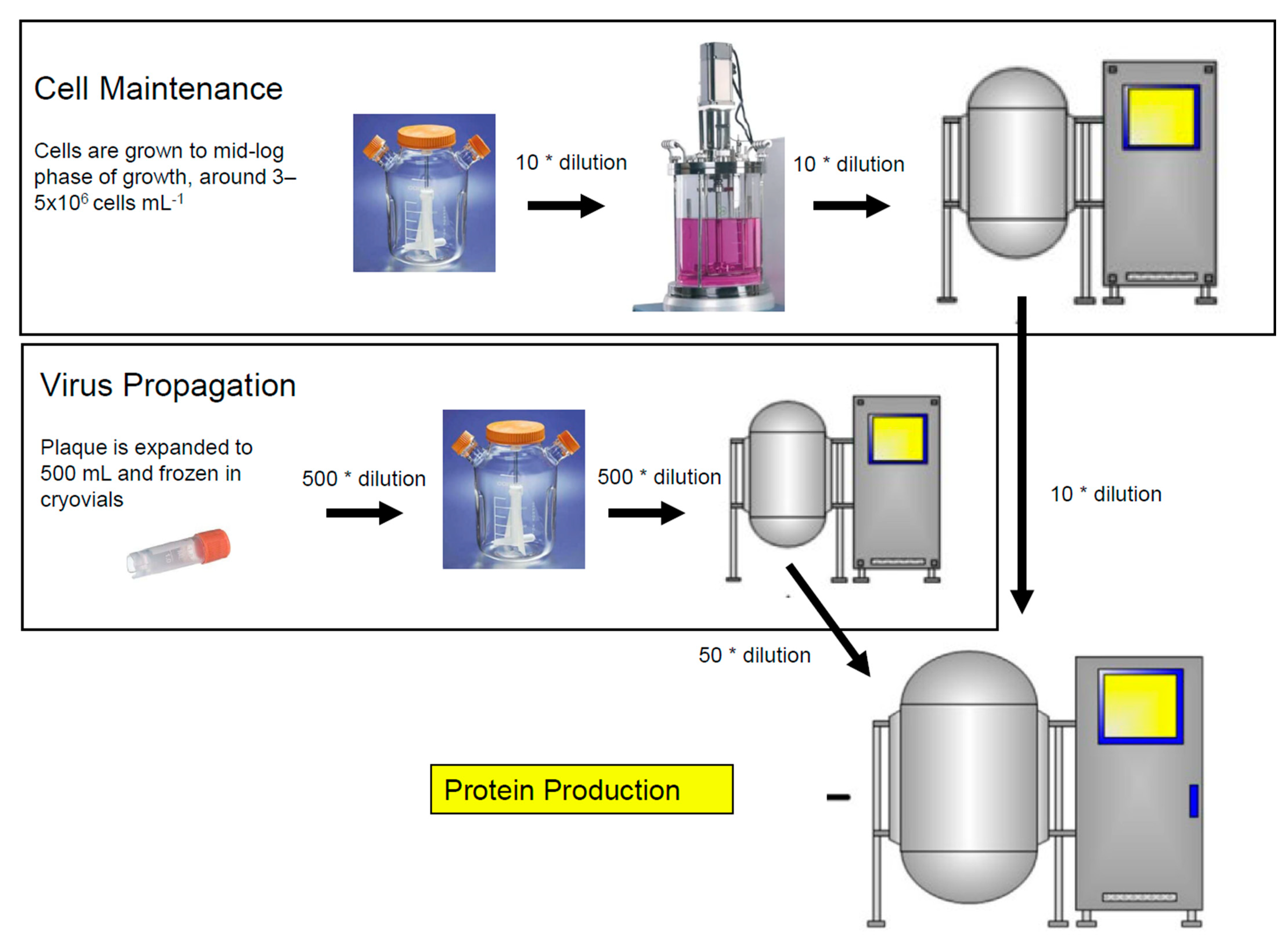

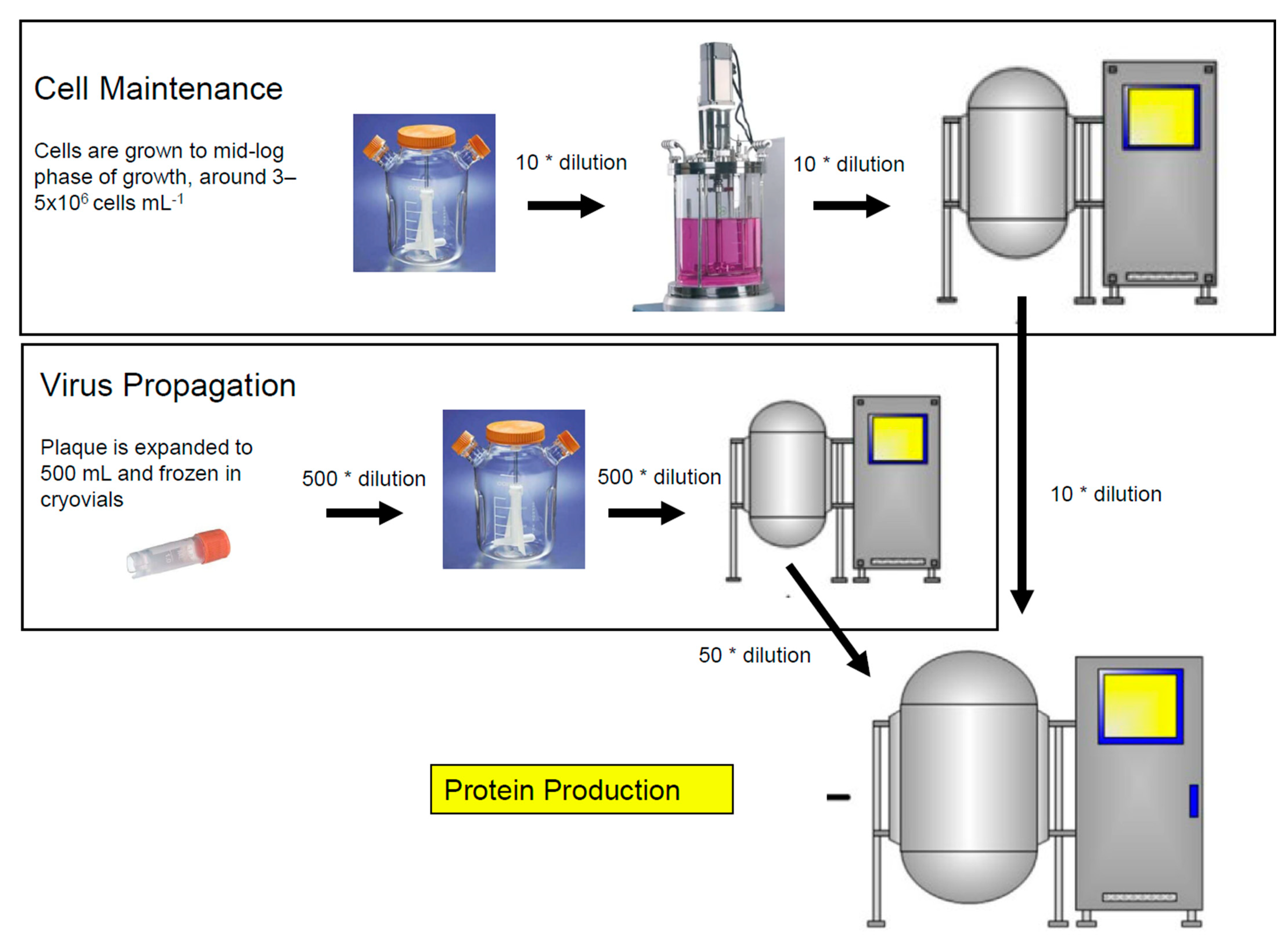{kind=link}
| Features | Description | Supplier (Assessed on 19 December 2021) |
|---|---|---|
| Promoter | Polyhedrin, P10, or basic promoter (late promoter) (single or double copy) | BD Biosciences (www.bdbiosciences.com) |
| Promoter + transactivation factors | Polyhedrin promoter plus transactivation factors IE1 and IE0 + HR linked to p10 chimeric promoter | Algenex (www.algenex.com) |
| Signal sequences | Acidic glycoprotein gp67 (also known as gp64) | BD Biosciences (www.bdbiosciences.com) |
| Honeybee | ThermoFisher (Waltham, MA, USA) (www.thermofisher.com) | |
| Delay cell death | Baculovirus vectors encoding P-vank-1 gene. | ParaTechs (www.paratechs.com) |
| Purification tags | His-tag or GST tag | BD Biosciences |
| Chaperones | Hsp40 and Hsc70 major ER molecular chaperones/disulfide isomerase | AB Vector (www.abvector.com) |
| Supplier Website (Assessed on 15 October 2021) | Medium Name | Chemically Defined | Animal-Component Free | Hydrolysate-Free | Serum-Free | Protein-free | Reported Max Cell Density Cells/mL |
|---|---|---|---|---|---|---|---|
| www.thermofisher.com | Sf-900 III (SFM) | N | Y | N | Y | N | 1–1.4 × 107 |
| ExpiSf CD | Y | Y | Y* | Y | Y | 2 × 107 | |
| www.sartorius.com | 4Cell® Insect CD | Y | Y | Y | Y | Y | 1 × 107 |
| www.expressionsystems.com | ESF AF contains L-Glutamine and Pluronic® F-68 | N | Y | ND | Y | Y | 2 × 107 |
| www.fishersci.com | Insectagro™ with L-Glutamine | N | ND | ND | Y | Y | ND |
| www.cytivalifesciences.com | SFM4Insect™ contains L-Glutamine and poloxamer 188 | N | Y | ND | Y | Y | ND |
| www.bdbiosciences.com | BaculoGold Max-XP | N | ND | N | Y | N | ND |
| www.labchem-wako.fujifilm.com | WakoVAC PSFM-J1 | N | ND | N | Y | N | ND |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cox, M.M.J. Innovations in the Insect Cell Expression System for Industrial Recombinant Vaccine Antigen Production. Vaccines 2021, 9, 1504. https://doi.org/10.3390/vaccines9121504
Cox MMJ. Innovations in the Insect Cell Expression System for Industrial Recombinant Vaccine Antigen Production. Vaccines. 2021; 9(12):1504. https://doi.org/10.3390/vaccines9121504
Chicago/Turabian StyleCox, Manon M. J. 2021. "Innovations in the Insect Cell Expression System for Industrial Recombinant Vaccine Antigen Production" Vaccines 9, no. 12: 1504. https://doi.org/10.3390/vaccines9121504
APA StyleCox, M. M. J. (2021). Innovations in the Insect Cell Expression System for Industrial Recombinant Vaccine Antigen Production. Vaccines, 9(12), 1504. https://doi.org/10.3390/vaccines9121504