
A Review of UK-Registered and Candidate Vaccines for Bovine Respiratory Disease

Abstract
1. Introduction
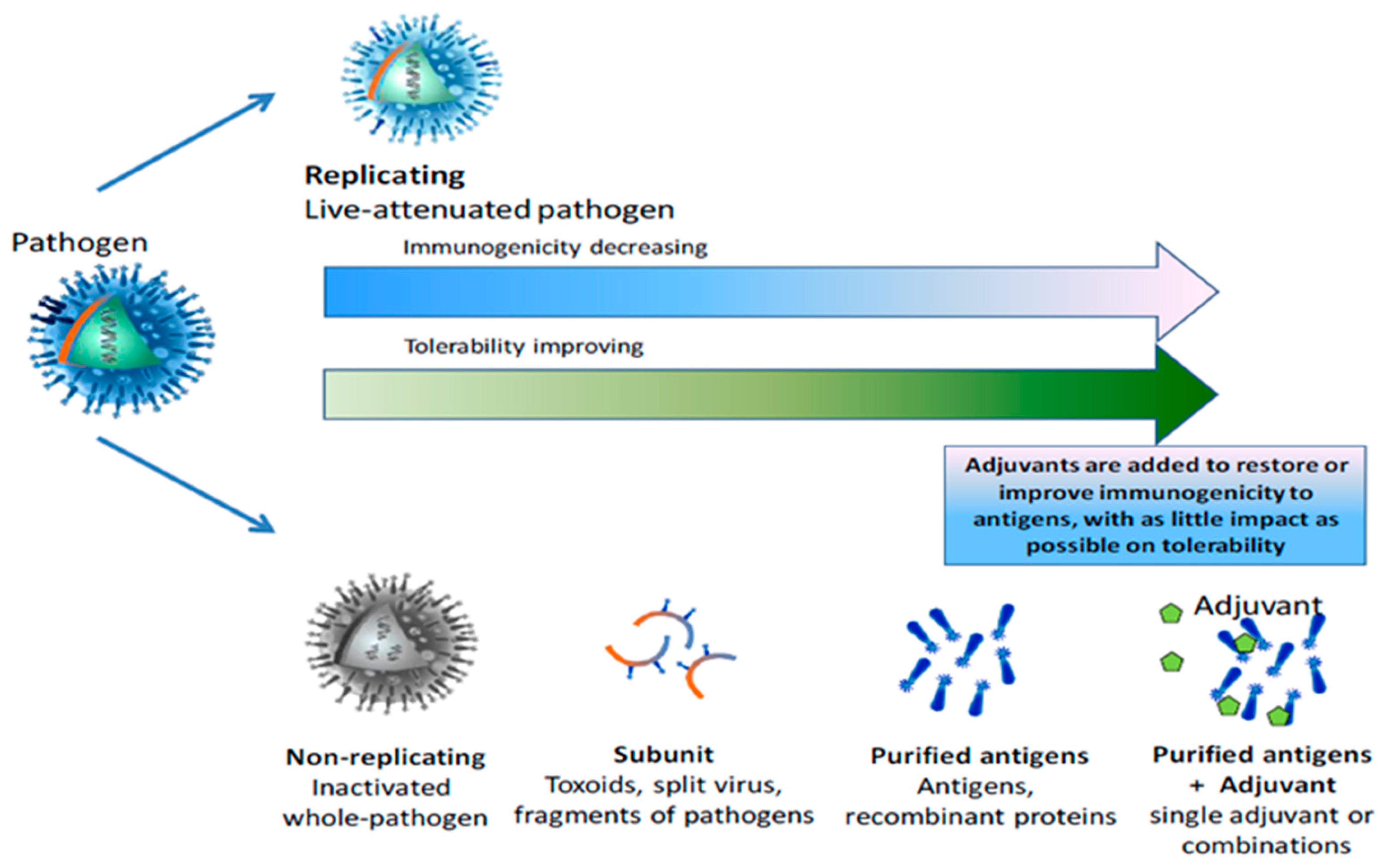
2. Currently Available Vaccines against BRD
3. Limitations of Currently Available BRD Vaccines
3.1. Age of Administration
3.2. Route of Administration
3.3. Type of Vaccines Available:
3.3.1. Modified-Live (MLV) Vaccines
3.3.2. Inactivated Vaccines
3.3.3. Immunogenicity of Modified-Live and Inactivated Vaccines
3.3.4. Vaccine-Enhanced Disease
3.4. Storage Conditions
4. Practical Considerations for Vaccine Development
4.1. Antimicrobial Medicines
4.2. EU Regulations
4.3. Veterinary Adjuvants
4.4. Vaccination Regimes
4.5. Pre-Partum Vaccination
5. Candidate BRD Vaccines
5.1. Peptide Vaccines
5.2. Immune Stimulatory Complexes (ISCOMs)
5.3. Virus-Like Particles (VLP)
5.4. Nanoparticle Vaccines
5.5. DNA Vaccines
5.6. Messenger RNA (mRNA) Vaccines
5.7. Viral Vectors
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
| Bovalto Pastobov (2007; Vm 08327/4173) |
| Multivalent: | N—Mannheimia haemolytica (formerly Pasteurella haemolytica) |
| Antigen: | Type A1 |
| Recommended age: | Minimum 4 weeks, booster 28 days later |
| Route of administration: | Sub-cutaneous or intramuscular |
| Adjuvant/excipients: | Aluminium hydroxide (4.2 mg), thiomersal, salts, water |
| Bovela lyophilisate (EU/2/14/176/001-016) |
| Multivalent: | N—BVDV (1—non cyto KE-9; 2—non cyto NY-93) |
| Antigen: | Modified live (both strains) |
| Recommended age: | 3 months + |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Not detailed |
| Bovilis BVDV suspension (1999; Vm06376/4025) |
| Multivalent: | N—BVDV 1 (non cyto C-86) |
| Antigen: | Inactivated |
| Recommended age: | 8 months |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Aluminium 3+ (Alum hydroxide) 6–9 mg |
| BOVILIS IBR marker inac (2006; vm06376/4053) |
| Multivalent: | N—BHV-1 (GK/D gE negative strain) |
| Antigen: | Inactivated |
| Recommended age: | 3 months onwards |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Alum 3+ (6.0–8.8 mg) formaldehyde 0.6–1 mg, trometamol |
| BOVILIS IBR MARKER LIVE (2006, vm06376/4032) |
| Multivalent: | N—BHV-1 (GK/D strain, gE negative) |
| Antigen: | Live |
| Recommended age: | 2 wk (Intranasal) 12 wk (Intramuscular) |
| Route of administration: | Intranasal or Intramuscular |
| Adjuvant/excipients: | Sorbitol, monosodium glutamate; glycine |
| HIPRABOVIS IBR marker live (2011; eu/2/10/114/001) |
| Multivalent: | N—BHV-1 (gE deleted; Ceddel strain) |
| Antigen: | Live |
| Recommended age: | 12 wk + |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Not disclosed |
| NASYM (EU/2/19/241/001-004 |
| Multivalent: | N—BRSV (LYM-56) |
| Antigen: | Live |
| Recommended age: | 9d (Intranasal) 10w (intramuscular) |
| Route of administration: | Intranasal OR Intramuscular |
| Adjuvant/excipients: | Not disclosed |
| RISPOVAL IBR MARKER (VM42058/4127, 1999) |
| Multivalent: | N—BHV-1 (gE deleted, Difivac Strain) |
| Antigen: | Inactivated |
| Recommended age: | 2w–3 m (if later than use 3rd booster) |
| Route of administration: | Sub cut Sub-cutaneous |
| Adjuvant/excipients: | QUIL A (0.25 MG) ALUM HYDROX (14–24 MG) |
| Rispoval Pateurella (VM42058/4128, 1999) |
| Multivalent: | N—Man.H type A1 (NL 1009 strain) |
| Antigen: | Inactivated |
| Recommended age: | 12 wk + |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Liquid paraffin and alum hydrox |
| Rispoval RS (vm 42058/4129, 2005) |
| Multivalent: | N—BRSV (RB94) |
| Antigen: | Live attenuated |
| Recommended age: | 7d–4m (If 7d then 3rd booster to counteract MDA) |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Not detailed |
| Tracherine (VM42058/4156; 2010) |
| Multivalent: | N—IBR (RBL106) |
| Antigen: | Live attenuated |
| Recommended age: | 3w (10 wk ideally for reduced MDA interference) |
| Route of administration: | Intranasal |
| Adjuvant/excipients: | Not detailed |
| Bovalto Respi 3 (2016) (Vm 08327/4273) |
| Multivalent: | Y—BRSV (BiO-24) and BPIV-3 (BiO-23) Man.H (A1) |
| Antigen: | All inactivated |
| Recommended age: | In cattle devoid of MDA (~6 months) |
| Route of administration: | Sub-cutaneous |
| Adjuvant/excipients: | AH (8.0 mg) Quil A (0.4 mg) Thiomersal (0.2 mg) Formaldehyde (1.0 mg) |
| Bovalto Respi 4 (2016; Vm 08327/4274) |
| Multivalent: | Y—BRSV (BiO24) BBPIV-3V (Bio23) Man H (A1) BVDVV (Bio25) |
| Antigen: | All inactivated |
| Recommended age: | 2 wk+ |
| Route of administration: | Sub-cutaneous |
| Adjuvant/excipients: | AH (8.0 mg) Quil A (0.4 mg) Thiomersal (0.2 mg) Formaldehyde (1.0 mg) |
| Bovalto Respi Intranasal (2018; Vm 08327/4289) |
| Multivalent: | Y—BPIV-3 (Bio23/A) BRSV (Bio24/A) |
| Antigen: | Modified live (both) |
| Recommended age: | 10 day + |
| Route of administration: | Intranasal |
| Adjuvant/excipients: | Not detailed |
| Bovilis bovipast rsp (1999; Vm 01708/4458) |
| Multivalent: | Y—BRSV (ev908) BPIV-3 (sf-4 reisinger) Man.H A1 (cross reactive to A6) |
| Antigen: | All inactivated |
| Recommended age: | ~2 wks of age, prior to housing |
| Route of administration: | Sub-cutaneous |
| Adjuvant/excipients: | Alum hydr (37.5 mg) Quil A (0.625 mg) Thiomersal (0.032–0.58 mg) |
| IMURESP-RP (vm 42058/4072 2005) |
| Multivalent: | Y—BPIV-3 (ts RLB103) IBR (ts RLB106) |
| Antigen: | Live attenuated |
| Recommended age: | 3–10 wk but 10+ wk preferred |
| Route of administration: | Intranasal |
| Adjuvant/excipients: | Not disclosed |
| Rispoval 3 (vm 42058/4124; 2005) |
| Multivalent: | Y—BPIV-3 (RLB103) BRSV (375) BVDVV (type 1 5960 c + 6309 n-c) |
| Antigen: | Modified live (BPIV-3, BRSV) inactivated BVDV |
| Recommended age: | 12 wk+ |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Alhydrogel 2% 0.8 mL (equiv. of 24.36 mg alum hydroxide) |
| Rispoval 4 (VM42058/4125, 2001) |
| Multivalent: | Y—As above (17) including IBR/BHV-1 |
| Antigen: | Inactivated |
| Recommended age: | 12 wk + |
| Route of administration: | Intramuscular |
| Adjuvant/excipients: | Not detailed |
| Rispoval RS + BPIV-3 (VM42058/4130) |
| Multivalent: | Y—BRSV (375) and BPIV-3 (ts RLB103) |
| Antigen: | Modified Live |
| Recommended age: | 9 day + |
| Route of administration: | Intranasal |
| Adjuvant/excipient: | Not disclosed |
References
- Callan, R.J.; Garry, F.B. Biosecurity and bovine respiratory disease. Veter. Clin Food Anim. Pract. 2002, 18, 57–77. [Google Scholar] [CrossRef]
- Scott, P. Respiratory Disease in Dairy and Beef Rearer Units. 2016. Available online: www.nadis.org.uk/4804 (accessed on 17 November 2018).
- Johnson, K.K.; Pendell, D.L. Market Impacts of Reducing the Prevalence of Bovine Respiratory Disease in United States Beef Cattle Feedlots. Front. Veter. Sci. 2017, 4, 189. [Google Scholar] [CrossRef] [PubMed]
- Blakebrough-Hall, C.; Mcmeniman, J.P.; González, L.A. Economics of Animal Production: An evaluation of the economic effects of bovine respiratory disease on animal performance, carcass traits and economic outcomes in feedlot cattle defined using four BRD diagnosis methods. J. Anim. Sci. 2020, 98, 1–11. [Google Scholar] [CrossRef]
- Kamdi, B.; Singh, R.; Singh, V.; Singh, S.; Kumar, P.; Singh, K.P.; George, N.; Dhama, K. Immunofluorescence and molecular diagnosis of bovine respiratory syncytial virus and bovine parainfluenza virus in the naturally infected young cattle and buffaloes from India. Microb. Pathog. 2020, 145, 104165. [Google Scholar] [CrossRef]
- Timsit, E.; Workentine, M.; van der Meer, F.; Alexander, T. Distinct bacterial metacommunities inhabit the upper and lower respiratory tracts of healthy feedlot cattle and those diagnosed with bronchopneumonia. Veter. Microbiol. 2018, 221, 105–113. [Google Scholar] [CrossRef] [PubMed]
- Kurćubić, V.; Đoković, R.; Ilić, Z.; Petrović, M. Etiopathogenesis and economic significance of bovine respiratory disease complex (BRDC). Acta Agric. Serbica 2018, 23, 85–100. [Google Scholar] [CrossRef]
- Delabouglise, A.; James, A.; Valarcher, J.F.; Hägglund, S.; Raboisson, D.; Rushton, J. Linking disease epidemiology and livestock productivity: The case of bovine respiratory disease in France. PLoS ONE 2017, 12, e0189090. [Google Scholar] [CrossRef] [PubMed]
- Kudirkiene, E.; Aagaard, A.K.; Schmidt, L.M.; Pansri, P.; Krogh, K.M.; Olsen, J.E. Occurrence of major and minor pathogens in calves diagnosed with bovine respiratory disease. Veter. Microbiol. 2021, 259, 109135. [Google Scholar] [CrossRef] [PubMed]
- Steiner, A.; Meylan, M.; Lava, M.; Schüpbach, G. Antimicrobial drug use and risk factors associated with treatment incidence and mortality in Swiss veal calves reared under improved welfare conditions. Prev. Vet. Med. 2016, 126, 121–130. [Google Scholar] [CrossRef]
- Makoschey, B.; Berge, A.C. Review on bovine respiratory syncytial virus and bovine parainfluenza – usual suspects in bovine respiratory disease—A narrative review. BMC Veter. Res. 2021, 17, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Bell, C.J.; Blackburn, P.; Elliott, M.; Patterson, T.I.A.P.; Ellison, S.; Lahuerta-Marin, A.; Ball, H.J. Investigation of polymerase chain reaction assays to improve detection of bacterial involvement in bovine respiratory disease. J. Veter. Diagn. Investig. 2014, 26, 631–634. [Google Scholar] [CrossRef] [PubMed]
- Baptista, A.L.; Rezende, A.L.; Fonseca, P.D.A.; Massi, R.P.; Nogueira, G.M.; Magalhães, L.Q.; Headley, S.A.; Menezes, G.L.; Alfieri, A.A.; Saut, J.P.E. Bovine respiratory disease complex associated mortality and morbidity rates in feedlot cattle from southeastern Brazil. J. Infect. Dev. Ctries. 2017, 11, 791–799. [Google Scholar] [CrossRef] [PubMed]
- Glover, I.D. Improving calf-side diagnosis of respiratory disease on UK dairy farms. Livestock 2017, 22, 230–238. [Google Scholar] [CrossRef][Green Version]
- Cavirani, S. Immunization of calves and herd immunity to Bovine respiratory Disease Complex (BRDC). Large Anim. Rev. 2019, 25, 17–24. [Google Scholar]
- Saade, G.; Deblanc, C.; Bougon, J.; Marois-Créhan, C.; Fablet, C.; Auray, G.; Belloc, C.; Leblanc-Maridor, M.; Gagnon, C.A.; Zhu, J.; et al. Coinfections and their molecular consequences in the porcine respiratory tract. Veter. Res. 2020, 51, 1–19. [Google Scholar] [CrossRef]
- Tibbo, M.; Woldemeskel, M.; Gopilo, A. An outbreak of respiratory disease complex in sheep in Central Ethiopia. Trop. Anim. Health Prod. 2001, 33, 355–365. [Google Scholar] [CrossRef] [PubMed]
- Scott, P.R. Treatment and Control of Respiratory Disease in Sheep. Veter. Food Anim. Pract. 2011, 27, 175–186. [Google Scholar] [CrossRef]
- Reagan, K.L.; Sykes, J.E. Canine Infectious Respiratory Disease. Veter. Food Anim. Pract. 2020, 50, 405–418. [Google Scholar] [CrossRef] [PubMed]
- Cohn, L.A. Feline Respiratory Disease Complex. Veter. Clin. N. Am. Anim. Pract. 2011, 41, 1273–1289. [Google Scholar] [CrossRef]
- Bryson, D. Calf Pneumonia. Veter. Clin. N. Am. Food Anim. Pract. 1985, 1, 237–257. [Google Scholar] [CrossRef]
- Larsen, L.E.; Tegtmeier, C.; Pedersen, E. Bovine respiratory syncytial virus (BRSV) pneumonia in beef herds despite vaccination. Acta Veter. Scand. 2001, 42, 113–121. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, L.J.; Berghaus, L.J.; Arnold, K.; Anderson, M.L.; Corbeil, L.B. Immune mechanisms of pathogenetic synergy in concurrent bovine pulmonary infection with Haemophilus somnus and bovine respiratory syncytial virus. Veter. Immunol. Immunopathol. 2005, 107, 119–130. [Google Scholar] [CrossRef] [PubMed]
- Beadling, C.; Slifka, M.K. How do viral infections predispose patients to bacterial infections? Curr. Opin. Infect. Dis. 2004, 17, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Manohar, P.; Loh, B.; Athira, S.; Nachimuthu, R.; Hua, X.; Welburn, S.C.; Leptihn, S. Secondary Bacterial Infections During Pulmonary Viral Disease: Phage Therapeutics as Alternatives to Antibiotics? Front. Microbiol. 2020, 11, 1434. [Google Scholar] [CrossRef]
- Bell, R.; Turkington, H.; Cosby, S. The Bacterial and Viral Agents of BRDC: Immune Evasion and Vaccine Developments. Vaccines 2021, 9, 337. [Google Scholar] [CrossRef] [PubMed]
- Hulbert, L.E.; Moisá, S.J. Stress, immunity, and the management of calves. J. Dairy Sci. 2016, 99, 3199–3216. [Google Scholar] [CrossRef]
- Van der Fels-Klerx, H.; Horst, H.; Dijkhuizen, A. Risk factors for bovine respiratory disease in dairy youngstock in The Netherlands: The perception of experts. Livest. Prod. Sci. 2000, 66, 35–46. [Google Scholar] [CrossRef]
- Pinchak, W.E.; Tolleson, D.R.; McCloy, M.; Hunt, L.J.; Gill, R.J.; Ansley, R.J.; Bevers, S.J. Morbidity effects on productivity and profitability of stocker cattle grazing in the Southern Plains. J. Anim. Sci. 2004, 82, 2773–2779. [Google Scholar] [CrossRef]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The epidemiology of bovine respiratory disease: What is the evidence for preventive measures? Can. Veter. J. 2010, 51, 1351–1359. [Google Scholar] [PubMed Central]
- Ribble, C.S.; Meek, A.H.; Janzen, E.D.; Guichon, P.T.; Jim, G.K. Effect of time of year, weather, and the pattern of auction market sales on fatal fibrinous pneumonia (shipping fever) in calves in a large feedlot in Alberta (1985–1988). Can. J. Veter. Res. 1995, 59, 167–172. [Google Scholar] [PubMed Central]
- MacVean, D.W.; Franzen, D.K.; Keefe, T.J.; Bennett, B.W. Airborne particle concentration and meteorologic conditions associated with pneumonia incidence in feedlot cattle. Am. J. Veter. Res. 1986, 47, 2676–2682. [Google Scholar] [PubMed]
- Alexander, B.H.; MacVean, D.W.; Salman, M.D. Risk factors for lower respiratory tract disease in a cohort of feedlot cattle. J. Am. Veter. Med Assoc. 1989, 195, 207–211. [Google Scholar] [PubMed]
- Härtel, H.; Nikunen, S.; Neuvonen, E.; Tanskanen, R.; Kivelä, S.-L.; Aho, P.; Soveri, T.; Saloniemi, H. Viral and Bacterial Pathogens in Bovine Respiratory Disease in Finland. Acta Veter. Scand. 2004, 45, 193–200. [Google Scholar] [CrossRef]
- Hay, K.; Morton, J.; Schibrowski, M.; Clements, A.; Mahony, T.; Barnes, T. Associations between prior management of cattle and risk of bovine respiratory disease in feedlot cattle. Prev. Veter. Med. 2016, 127, 37–43. [Google Scholar] [CrossRef]
- Hägglund, S.; Hjort, M.; Graham, D.; Öhagen, P.; Törnquist, M.; Alenius, S. A six-year study on respiratory viral infections in a bull testing facility. Veter. J. 2007, 173, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Hotchkiss, E.J.; Dagleish, M.P.; Willoughby, K.; McKendrick, I.J.; Finlayson, J.; Zadoks, R.N.; Newsome, E.; Brulisauer, F.; Gunn, G.J.; Hodgson, J.C. Prevalence of Pasteurella multocidaand other respiratory pathogens in the nasal tract of Scottish calves. Veter. Rec. 2010, 167, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Neibergs, H.L.; Seabury, C.M.; Wojtowicz, A.J.; Wang, Z.; Scraggs, E.; Kiser, J.N.; Neupane, M.; E Womack, J.; Van Eenennaam, A.; Hagevoort, G.R.; et al. Susceptibility loci revealed for bovine respiratory disease complex in pre-weaned holstein calves. BMC Genom. 2014, 15, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.; Earley, B.; McCabe, M.S.; Lemon, K.; Duffy, C.; McMenamy, M.; Cosby, S.L.; Kim, J.; Blackshields, G.; Taylor, J.; et al. Experimental challenge with bovine respiratory syncytial virus in dairy calves: Bronchial lymph node transcriptome response. Sci. Rep. 2019, 9, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Johnston, D.; Earley, B.; McCabe, M.S.; Kim, J.; Taylor, J.F.; Lemon, K.; Duffy, C.; McMenamy, M.; Cosby, S.L.; Waters, S.M. Messenger RNA biomarkers of Bovine Respiratory Syncytial Virus infection in the whole blood of dairy calves. Sci. Rep. 2021, 11, 1–7. [Google Scholar] [CrossRef]
- Mitra, N.; Cernicchiaro, N.; Torres, S.; Li, F.; Hause, B. Metagenomic characterisation of the virome associated with bovine respiratory disease in feedlot cattle identified novel viruses and suggests an etiologic role for influenza D virus. J. Gen. Virol. 2016, 97, 1771–1784. [Google Scholar] [CrossRef] [PubMed]
- Dane, H.; Duffy, C.; Guelbenzu, M.; Hause, B.; Fee, S.; Forster, F.; McMenamy, M.J.; Lemon, K. Detection of influenza D virus in bovine respiratory disease samples, UK. Transbound. Emerg. Dis. 2019, 66, 2184–2187. [Google Scholar] [CrossRef]
- Ellis, J.A. Update on viral pathogenesis in BRD. Anim. Health. Res. Rev. 2009, 10, 149–153. [Google Scholar] [CrossRef] [PubMed]
- Prysliak, T.; van der Merwe, J.; Lawman, Z.; Wilson, D.; Townsend, H. Respiratory disease caused by Mycoplasma bovis is enhanced by exposure to bovine herpes virus 1 (BHV-1) but not to bovine viral diarrhea virus (BVDV) type 2. Can. Veter. J. 2011, 52, 1195, ISSN: 0008-5286. [Google Scholar]
- Fahkrajang, W.; Sudaryatma, P.E.; Mekata, H.; Hamabe, S.; Saito, A.; Okabayashi, T. Bovine respiratory coronavirus enhances bacterial adherence by upregulating expression of cellular receptors on bovine respiratory epithelial cells. Veter. Microbiol. 2021, 255, 109017. [Google Scholar] [CrossRef] [PubMed]
- Ohlson, A.; Alenius, S.; Tråvén, M.; Emanuelson, U. A longitudinal study of the dynamics of bovine corona virus and respiratory syncytial virus infections in dairy herds. Veter. J. 2013, 197, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Uttenthal, Å.; Larsen, L.E.; Philipsen, J.S.; Tjørnehøj, K.; Viuff, B.; Nielsen, K.H.; Nielsen, T.K. Antibody dynamics in BRSV-infected Danish dairy herds as determined by isotype-specific immunoglobulins. Veter. Microbiol. 2000, 76, 329–341. [Google Scholar] [CrossRef]
- Collins, J.; Teegarden, R.; MacVean, D.; Salman Smith, G.; Frank, G. Prevalence and specificity of antibodies to bovine respiratory syncytial virus in sera from feedlot and range cattle. Am. J. Veter. Res. 1988, 49, 1316–1319. [Google Scholar] [PubMed]
- Obando, R.; Hidalgo, M.; Merza, M.; Montoya, A.; Klingeborn, B.; Moreno-López, J. Seroprevalence to bovine virus diarrhoea virus and other viruses of the bovine respiratory complex in Venezuela (Apure State). Prev. Veter. Med. 1999, 41, 271–278. [Google Scholar] [CrossRef]
- Solís-Calderón, J.; Segura-Correa, J.; Aguilar-Romero, F.; Segura-Correa, V. Detection of antibodies and risk factors for infection with bovine respiratory syncytial virus and parainfluenza virus-3 in beef cattle of Yucatan, Mexico. Prev. Veter. Med. 2007, 82, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Kimman, T.G.; Westenbrink, F.; Schreuder, B.E.; Straver, P.J. Local and systemic antibody response to bovine respiratory syncytial virus infection and reinfection in calves with and without maternal antibodies. J. Clin. Microbiol. 1987, 25, 1097–1106. [Google Scholar] [CrossRef] [PubMed]
- National Office of Animal Health. Compendium of Data Sheets for Animal Medicines 2021; National Office of Animal Health: Enfield, UK, 2021; 1584p. [Google Scholar]
- Chase, C.C.; Hurley, D.J.; Reber, A.J. Neonatal Immune Development in the Calf and Its Impact on Vaccine Response. Veter. Clin. N. Am. Food Anim. Pract. 2008, 24, 87–104. [Google Scholar] [CrossRef]
- Niewiesk, S. Maternal Antibodies: Clinical Significance, Mechanism of Interference with Immune Responses, and Possible Vaccination Strategies. Front. Immunol. 2014, 5, 446. [Google Scholar] [CrossRef]
- Richeson, J.T.; Falkner, T.R. Bovine Respiratory Disease Vaccination: What Is the Effect of Timing? Veter. Clin. N. Am. Food Anim. Pract. 2020, 36, 473–485. [Google Scholar] [CrossRef] [PubMed]
- Cortese, V.S. Neonatal Immunology. Veter. Clin. N. Am. Food Anim. Pract. 2009, 25, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Tregoning, J.S.; Wang, B.L.; McDonald, J.U.; Yamaguchi, Y.; Harker, J.A.; Goritzka, M.; Openshaw, P.J. Neonatal antibody responses are attenuated by interferon-γ produced by NK and T cells during RSV infection. Proc. Natl. Acad. Sci. USA 2013, 110, 5576–5581. [Google Scholar] [CrossRef] [PubMed Central]
- Ellis, J.; Gow, S.; Bolton, M.; Burdett, W.; Nordstrom, S. Inhibition of priming for bovine respiratory syncytial virus-specific protective immune responses following parenteral vaccination of passively immune calves. Can. Veter. J. 2014, 55, 1180–1185. [Google Scholar]
- Gershwin, L.J.; Behrens, N.E.; McEligot, H.A.; Carvallo-Chaigneau, F.R.; Crum, L.T.; Gunnarson, B.M.; Corbeil, L.B. A recombinant subunit vaccine for bovine RSV and Histophilus somni protects calves against dual pathogen challenge. Vaccine 2017, 35, 1954–1963. [Google Scholar] [CrossRef] [PubMed]
- Van der Sluijs, M.; Kuhn, E.; Makoschey, B. A single vaccination with an inactivated bovine respiratory syncytial virus vaccine primes the cellular immune response in calves with maternal antibody. BMC Veter. Res. 2010, 6, 2. [Google Scholar] [CrossRef]
- Blodörn, K.; Hägglund, S.; Fix, J.; Dubuquoy, C.; Makabi-Panzu, B.; Thom, M.; Karlsson, P.; Roque, J.-L.; Karlstam, E.; Pringle, J.; et al. Vaccine Safety and Efficacy Evaluation of a Recombinant Bovine Respiratory Syncytial Virus (BRSV) with Deletion of the SH Gene and Subunit Vaccines Based On Recombinant Human RSV Proteins: N-nanorings, P and M2-1, in Calves with Maternal Antibodies. PLoS ONE 2014, 9, e100392. [Google Scholar] [CrossRef] [PubMed]
- Fulton, R.W.; E Briggs, R.; E Payton, M.; Confer, A.W.; Saliki, J.; Ridpath, J.F.; Burge, L.J.; Duff, G.C. Maternally derived humoral immunity to bovine viral diarrhea virus (BVDV) 1a, BVDV1b, BVDV2, bovine herpesvirus-1, parainfluenza-3 virus bovine respiratory syncytial virus, Mannheimia haemolytica and Pasteurella multocida in beef calves, antibody decline by half-life studies and effect on response to vaccination. Vaccine 2004, 22, 643–649. [Google Scholar] [CrossRef] [PubMed]
- Richeson, J.T.; Hughes, H.D.; Broadway, P.R.; Carroll, J.A. Vaccination management of beef cattle: Delayed vaccination and Endotoxin stacking. Veter. Clin. Food Anim. 2019, 35, 575–592. [Google Scholar] [CrossRef]
- Poulet, H.; Minke, J.; Pardo, M.C.; Juillard, V.; Nordgren, B.; Audonnet, J.-C. Development and registration of recombinant veterinary vaccines: The example of the canarypox vector platform. Vaccine 2007, 25, 5606–5612. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J.; Gow, S.; West, K.; Waldner, C.; Rhodes, C.; Mutwiri, G.; Rosenberg, H. Response of calves to challenge exposure with virulent bovine respiratory syncytial virus following intranasal administration of vaccines formulated for parenteral administration. J. Am. Veter. Med Assoc. 2007, 230, 233–243. [Google Scholar] [CrossRef]
- Rossi, P.S.; Mattei, R.I.; Schllemer, N.R.; Thomaz, G.R.; Antunes, A.V.; Virmond, M.P.; Taube, M.J.; Bertagnon, H.G. The effect of bovine vaccines against respiratory viruses administered either intranasal or intramuscular on broncho-alveolar fluid cells of heifers. Veter. Q. 2021, 41, 97–106. [Google Scholar] [CrossRef] [PubMed]
- Wright, P.F.; Karron, R.A.; Belshe, R.B.; Thompson, J.; Crowe, J.; Boyce, T.G.; Halburnt, L.L.; Reed, G.W.; Whitehead, S.S.; Anderson, E.L.; et al. Evaluation of a Live, Cold-Passaged, Temperature-Sensitive, Respiratory Syncytial Virus Vaccine Candidate in Infancy. J. Infect. Dis. 2000, 182, 1331–1342. [Google Scholar] [CrossRef]
- Henderson, D.A. The eradication of smallpox—An overview of the past, present, and future. Vaccine 2011, 29 (Suppl. 4), D7–D9. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.; Baron, M.; Cameron, A.; Kock, R.; Jones, B.; Pfeiffer, D.; Mariner, J.; McKeever, D.; Oura, C.; Roeder, P.; et al. Rinderpest eradicated; what next? Veter. Rec. 2011, 169, 10–11. [Google Scholar] [CrossRef] [PubMed]
- Roeder, P.L. Rinderpest: The end of cattle plague. Prev. Veter. Med. 2011, 102, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Sanders, B.; Koldijk, M.; Schuitemaker, H. Inactivated Viral Vaccines. In Infectious Disease and Vaccines Therapeutic Area; Springer: Berlin/Heidelberg, Germany, 2015; pp. 45–80. [Google Scholar]
- Clem, A.S. Fundamentals of vaccine immunology. J. Glob. Infect. Dis. 2011, 3, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Ficht, T.A.; Kahl-McDonagh, M.M.; Arenas-Gamboa, A.M.; Rice-Ficht, A.C. Brucellosis: The case for live, attenuated vaccines. Vaccine 2009, 27, D40–D43. [Google Scholar] [CrossRef] [PubMed]
- Goldsby, R.A.; Kindt, T.J.; Osborne, B.A.; Kuby, J. Immunology, 5th ed; Freeman: New York, NY, USA, 2003. [Google Scholar]
- National Institute of Allergy and Infectious Disease. Vaccine Types. Available online: https://www.niaid.nih.gov/research/vaccine-types (accessed on 10 October 2021).
- Stevens, E.T.; Brown, M.S.; Burdett, W.W.; Bolton, M.W.; Nordstrom, S.T.; Chase, C.C.L. Efficacy of a Non-adjuvanted, Modified-live Virus Vaccine in Calves with Maternal Antibodies against a Virulent Bovine Viral Diarrhea Virus Type 2a Challenge Seven Months following Vaccination. Am. Assoc. Bov. Pract. 2011, 45, 23–31. [Google Scholar] [CrossRef]
- Salerno-Gonçalves, R.; Sztein, M.B. Cell-mediated immunity and the challenges for vaccine development. Trends Microbiol. 2006, 14, 536–542. [Google Scholar] [CrossRef] [PubMed]
- West, K.; Petrie, L.; Haines, D.M.; Konoby, C.; Clark, E.G.; Martin, K.; A Ellis, J. The effect of formalin-inactivated vaccine on respiratory disease associated with bovine respiratory syncytial virus infection in calves. Vaccine 1999, 17, 809–820. [Google Scholar] [CrossRef]
- Vangeel, I.; Antonis, A.F.; Fluess, M.; Riegler, L.; Peters, A.R.; Harmeyer, S.S. Efficacy of a modified live intranasal bovine respiratory syncytial virus vaccine in 3-week-old calves experimentally challenged with BRSV. Veter. J. 2007, 174, 627–635. [Google Scholar] [CrossRef]
- Xue, W.; Ellis, J.; Mattick, D.; Smith, L.; Brady, R.; Trigo, E. Immunogenicity of a modified-live virus vaccine against bovine viral diarrhea virus types 1 and 2, infectious bovine rhinotracheitis virus, bovine parainfluenza-3 virus, and bovine respiratory syncytial virus when administered intranasally in young calves. Vaccine 2010, 28, 3784–3792. [Google Scholar] [CrossRef] [PubMed]
- Chamorro, M.F.; Palomares, R.A. Bovine Respiratory Disease Vaccination Against Viral Pathogens: Modified-Live Versus Inactivated Antigen Vaccines, Intranasal Versus Parenteral, What Is the Evidence? Veter. Clin. Food Anim. 2020, 36, 461–472. [Google Scholar] [CrossRef] [PubMed]
- Kerkhofs, P.; Tignon, M.; Petry, H.; Mawhinney, I.; Sustronck, B. Immune responses to bovine respiratory syncytial virus (BRSV) following use of an inactivated BRSV-PI3-Mannheimia haemolytica vaccine and a modified live BRSV–BVDV vaccine. Veter. J. 2004, 167, 208–210. [Google Scholar] [CrossRef]
- Knudson, C.J.; Hartwig, S.M.; Meyerholz, D.; Varga, S.M. RSV Vaccine-Enhanced Disease Is Orchestrated by the Combined Actions of Distinct CD4 T Cell Subsets. PLoS Pathog. 2015, 11, e1004757. [Google Scholar] [CrossRef] [PubMed]
- Woolums, A.R.; Singer, R.S.; Boyle, G.A.; Gershwin, L.J. Interferon gamma production during bovine respiratory syncytial virus (BRSV) infection is diminished in calves vaccinated with formalin-inactivated BRSV. Vaccine 1999, 17, 1293–1297. [Google Scholar] [CrossRef]
- Demasius, W.; Weikard, R.; Hadlich, F.; Müller, K.E.; Kühn, C. Monitoring the immune response to vaccination with an inactivated vaccine associated to bovine neonatal pancytopenia by deep sequencing transcriptome analysis in cattle. Veter. Res. 2013, 44, 93. [Google Scholar] [CrossRef]
- Ellis, J.A.; Gow, S.P.; Mahan, S.; Leyh, R. Duration of immunity to experimental infection with bovine respiratory syncytial virus following intranasal vaccination of young passively immune calves. J. Am. Veter. Med Assoc. 2013, 243, 1602–1608. [Google Scholar] [CrossRef]
- Patel, J.R.; Didlick, S.A. Evaluation of efficacy of an inactivated vaccine against bovine respiratory syncytial virus in calves with maternal antibodies. Am. J. Veter. Res. 2004, 65, 417–421. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Canchola, J.G.; Brandt, C.D.; Pyles, G.; Chanock, R.M.; Jensen, K.; Parrott, R.H. Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 1968, 89, 422–434, ISSN: 00029262. [Google Scholar] [CrossRef] [PubMed]
- Antonis, A.F.G.; Schrijver, R.S.; Daus, F.; Steverink, P.J.G.M.; Stockhofe, N.; Hensen, E.J.; Langedijk, J.P.M.; van der Most, R.G. Vaccine-Induced Immunopathology during Bovine Respiratory Syncytial Virus Infection: Exploring the Parameters of Pathogenesis. J. Virol. 2003, 77, 12067–12073. [Google Scholar] [CrossRef] [PubMed]
- Gershwin, L.J.; Schelegle, E.S.; Gunther, R.A.; Anderson, M.L.; Woolums, A.R.; Larochelle, D.R.; Boyle, G.A.; Friebertshauser, K.E.; Singer, R.S. A bovine model of vaccine enhanced respiratory syncytial virus pathophysiology. Vaccine 1998, 16, 1225–1236. [Google Scholar] [CrossRef]
- Kalina, W.V.; Woolums, A.R.; Gershwin, L.J. Formalin-inactivated bovine RSV vaccine influences antibody levels in bronchoalveolar lavage fluid and disease outcome in experimentally infected calves. Vaccine 2005, 23, 4625–4630. [Google Scholar] [CrossRef]
- Delgado, M.F.; Coviello, S.; Monsalvo, A.C.; A Melendi, G.; Hernandez, J.Z.; Batalle, J.P.; Diaz, L.; Trento, A.; Chang, H.-Y.; Mitzner, W.; et al. Lack of antibody affinity maturation due to poor Toll-like receptor stimulation leads to enhanced respiratory syncytial virus disease. Nat. Med. 2009, 15, 34–41. [Google Scholar] [CrossRef]
- Williams, P.D.; Paixão, G. On-farm storage of livestock vaccines may be a risk to vaccine efficacy: A study of the performance of on-farm refrigerators to maintain the correct storage temperature. BMC Veter. Res. 2018, 14, 1–7. [Google Scholar] [CrossRef]
- Saylor, K.; Gillam, F.; Lohneis, T.; Zhang, C. Designs of Antigen Structure and Composition for Improved Protein-Based Vaccine Efficacy. Front. Immunol. 2020, 11, 283. [Google Scholar] [CrossRef]
- Ren, Q.; Xiong, H.; Li, Y.; Xu, R.; Zhu, C. Evaluation of an outside-the-Cold-Chain Vaccine Delivery Strategy in Remote Regions of Western China. Public Health Rep. 2009, 124, 745–750. [Google Scholar] [CrossRef]
- Glenn, G.M.; Smith, G.; Fries, L.; Raghunandan, R.; Lu, H.; Zhou, B.; Thomas, D.N.; Hickman, S.P.; Kpamegan, E.; Boddapati, S.; et al. Safety and immunogenicity of a Sf9 insect cell-derived respiratory syncytial virus fusion protein nanoparticle vaccine. Vaccine 2013, 31, 524–532. [Google Scholar] [CrossRef] [PubMed]
- Atuhaire, D.K.; Lieberman, D.; Marcotty, T.; Musoke, A.J.; Madan, D. An alternative cold chain for storing and transporting East Coast fever vaccine. Veter. Parasitol. 2020, 288, 109304. [Google Scholar] [CrossRef]
- Gomez, M.; McCollum, J.; Wang, H.; Bachchhav, S.; Tetreau, I.; Gerhardt, A.; Press, C.; Kramer, R.M.; Fox, C.B.; Vehring, R. Evaluation of the stability of a spray-dried tuberculosis vaccine candidate designed for dry powder respiratory delivery. Vaccine 2021, 39, 5025–5036. [Google Scholar] [CrossRef]
- Cameron, A.; McAllister, T.A. Antimicrobial usage and resistance in beef production. J. Anim. Sci. Biotechnol. 2016, 7, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Johnson, J.C.; Bryson, W.L.; Barringer, S.; Hunsaker, B.D. Evaluation of on-arrival versus prompted metaphylaxis regimes using ceftiofur crystalline free acid for feedlot heifers at risk of developing bovine respiratory disease. Veter. Ther. 2008, 9, 53–62. [Google Scholar]
- Van Boeckel, T.P.; Brower, C.; Gilbert, M.; Grenfell, B.T.; Levin, S.A.; Robinson, T.P.; Teillant, A.; Laxminarayan, R. Global trends in antimicrobial use in food animals. Proc. Natl. Acad. Sci. USA 2015, 112, 5649–5654. [Google Scholar] [CrossRef]
- Department of Health; Department for Environment Food and Rural Affairs. UK Five Year Antimicrobial Resistance Strategy: 2013 to 2018. 2013. Available online: https://www.gov.uk/government/uploads/system/uploads/attachment_data/file/244058/20130902_UK_5_year_AMR_strategy.pdf (accessed on 31 July 2016).
- Singh, K.B.; Mehta, S. The clinical development process for a novel preventive vaccine: An overview. J. Postgrad. Med. 2016, 62, 4–11. [Google Scholar] [CrossRef] [PubMed]
- Knight-Jones, T.J.D.; Edmond, K.; Gubbins, S.; Paton, D.J. Veterinary and human vaccine evaluation methods. Proc. R. Soc. B 2014, 281, 20132839. [Google Scholar] [CrossRef]
- Draayer, H. Overview of currently approved veterinary vaccine potency testing methods and methods in development that do not require animal use. Procedia Vaccinol. 2011, 5, 171–174. [Google Scholar] [CrossRef]
- Six, A.; Bellier, B.; Thomas-Vaslin, V.; Klatzmann, D. Systems biology in vaccine design. Microb. Biotechnol. 2012, 5, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Mackay, D.; Jimenez, C.M.; Ioannou, F.; Ilott, M. The contribution of field efficacy studies to the evaluation of applications for veterinary vaccines evaluated through the European Union centralised authorisation procedure. Biologicals 2021, 70, 7–16. [Google Scholar] [CrossRef]
- The European Agency for the Evaluation of Veterinary Medicines. Guidance for Veterinary Medicinal Products; EMEA/CVMP/852/99-FINAL; European Medicines Agency: Amsterdam, The Netherlands, 2001.
- Levings, R.L.; Roth, J.A. Immunity to Bovine Herpesvirus 1: II. Adaptive Immunity and Vaccinology. Anim. Health Res. Rev. 2013, 14, 103–123. [Google Scholar] [CrossRef] [PubMed]
- Guzman, E.; Taylor, G. Immunology of bovine respiratory syncytial virus in calves. Mol. Immunol. 2015, 66, 48–56. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, S.; Safhi, M.M.; Kannadasan, M.; Sukumaran, N. Vaccine adjuvants – Current status and prospects on controlled release adjuvancity. Saudi Pharm. J. 2011, 19, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Di Pasquale, A.; Preiss, S.; Silva, F.M.D.O.E.; Garçon, N. Vaccine Adjuvants: From 1920 to 2015 and Beyond. Vaccines 2015, 3, 320–343. [Google Scholar] [CrossRef]
- Mogil, J.S.; Crager, S.E. What should we be measuring in behavioral studies of chronic pain in animals? Pain 2004, 112, 12–15. [Google Scholar] [CrossRef]
- Burakova, Y.; Madera, R.; McVey, S.; Schlup, J.R.; Shi, J. Adjuvants for Animal Vaccines. Viral Immunol. 2018, 31, 11–22. [Google Scholar] [CrossRef]
- Parker, R.; Deville, S.; Dupuis, L.; Bertrand, F.; Aucouturier, J. Adjuvant formulation for veterinary vaccines: Montanide™ Gel safety profile. Procedia Vaccinol. 2009, 1, 140–147. [Google Scholar] [CrossRef]
- Dar, P.; Kalaivanan, R.; Sied, N.; Mamo, B.; Kishore, S.; Suryanarayana, V.; Kondabattula, G. Montanide ISA™ 201 adjuvanted FMD vaccine induces improved immune responses and protection in cattle. Vaccine 2013, 31, 3327–3332. [Google Scholar] [CrossRef]
- Kolta, S.; Vishanath, B.S.; Dechamma, H.J.; Ganesh, K.; Suryanarayana, V.V.S.; Reddy, G.R. DNA vaccine (P1-2A-3C-pCDNA) co-administered with Bovine IL-18 gives protective immune response against Foot and Mouth Disease in cattle. Veter. Microbiol. 2016, 193, 106–115. [Google Scholar] [CrossRef]
- Wilson-Welder, J.H.; Boggiatto, P.; Nally, J.E.; Wafa, E.; Alt, D.P.; Hornsby, R.L.; Frank, A.; Jones, D.E.; Olsen, S.C.; Bowden, N.B.; et al. Bovine immune response to leptospira antigen in different novel adjuvants and vaccine delivery platforms. Vaccine 2020, 38, 3464–3473. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, V.S.; Santos, F.D.S.; Junior, A.G.D.S.; Piraine, R.E.A.; Rodrigues, P.R.C.; Brasil, C.L.; Conrad, N.L.; Leite, F.P.L. Recombinant bovine IL17A acts as an adjuvant for bovine herpesvirus vaccine. Res. Veter. Sci. 2021, 136, 185–191. [Google Scholar] [CrossRef] [PubMed]
- Carine, L.; Mathieu, B.; Laurent, R.; Jean-François, T.; Karl, W.; Stefan, R.; Gilles, M.; Jean-Jacques, L.; Pierre, K.; Letellier, C.; et al. Vaccination of calves using the BRSV nucleocapsid protein in a DNA prime–protein boost strategy stimulates cell-mediated immunity and protects the lungs against BRSV replication and pathology. Vaccine 2008, 26, 4840–4848. [Google Scholar] [CrossRef] [PubMed]
- Hill, K.; Arsic, N.; Nordstrom, S.; Griebel, P.J. Immune memory induced by intranasal vaccination with a modified-live viral vaccine delivered to colostrum fed neonatal calves. Vaccine 2019, 37, 7455–7462. [Google Scholar] [CrossRef]
- Lu, S. Heterologous prime-boost vaccination. Curr. Opin. Immun. 2009, 21, 346–351. [Google Scholar] [CrossRef]
- Kardani, K.; Bolhassani, A.; Shahbazi, S. Prime-boost vaccine strategy against viral infections: Mechanisms and benefits. Vaccine 2016, 34, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Mapletoft, J.W.; Latimer, L.; Babiuk, L.A.; Hurk, S.V.D.L.-V.D. Intranasal Immunization of Mice with a Bovine Respiratory Syncytial Virus Vaccine Induces Superior Immunity and Protection Compared to Those by Subcutaneous Delivery or Combinations of Intranasal and Subcutaneous Prime-Boost Strategies. Clin. Vaccine Immunol. 2010, 17, 23–35. [Google Scholar] [CrossRef]
- Miao, C.; Woolums, A.R.; Zarlenga, D.S.; Brown, C.C.; Brown, J.C.; Williams, S.M.; Scott, M.A. Effects of a single intranasal dose of modified-live bovine respiratory syncytial virus vaccine on cytokine messenger RNA expression following viral challenge in calves. Am. J. Veter. Res. 2004, 65, 725–733. [Google Scholar] [CrossRef]
- Riffault, S.; Hägglund, S.; Guzman, E.; Näslund, K.; Jouneau, L.; Dubuquoy, C.; Valarcher, J.F. A Single Shot Pre-fusion-Stabilized Bovine RSV F Vaccine is Safe and E ff ective in Newborn Calves with Maternally Derived Antibodies. Vaccines 2020, 8, 231. [Google Scholar] [CrossRef] [PubMed]
- Mawhinney, I.C.; Burrows, M.R. Protection against bovine respiratory syncytial virus challenge following a single dose of vaccine in young calves with maternal antibody. Veter. Rec. 2005, 156, 139–143. [Google Scholar] [CrossRef] [PubMed]
- Loucks, M.E.; Morrill, J.L.; Dayton, A.D. Effect of prepartum vaccination with K99 Esherichia coli vaccine on maternal and calf blood antibody concentration and calf health. J. Dairy Sci. 1985, 68, 1841–1847. [Google Scholar] [CrossRef]
- Kohara, J.; Hirai, T.; Mori, K.; Ishizaki, H.; Tsunemitsu, H. Enhancement of Passive Immunity with Maternal Vaccine against Newborn Calf Diarrhea. J. Veter. Med Sci. 1997, 59, 1023–1025. [Google Scholar] [CrossRef] [PubMed]
- Dudek, K.; Bednarek, D.; Ayling, R.D.; Szacawa, E. Stimulation and analysis of the immune response in calves from vaccinated pregnant cows. Res. Veter.Sci. 2014, 97, 32–37. [Google Scholar] [CrossRef] [PubMed]
- Baccili, C.C.; Silva, C.C.; Baldacim, P.; Greghi, G.F.; Vasconcellos, F.M.; Cacciacarro, B.S.; Gomes, V. Influence of maternal vaccination for the passive immune transfer against bovine respiratory viruses. Arq. Bras. Med. Veterinária Zootec. 2018, 70, 391–400. [Google Scholar] [CrossRef]
- Tardon, M.C.; Allard, M.; Dutoit, V.; Dietrich, P.; Walker, P.R. Peptides as cancer vaccines. Curr. Opin. Pharmacol. 2019, 47, 20–26. [Google Scholar] [CrossRef]
- Combadière, B.; Beaujean, M.; Chaudesaigues, C.; Vieillard, V. Peptide-Based Vaccination for Antibody Responses Against HIV. Vaccines 2019, 7, 105. [Google Scholar] [CrossRef] [PubMed]
- Nakagami, H.; Koriyama, H.; Morishita, R. Peptide Vaccines for Hypertension and Diabetes Mellitus. Vaccines 2014, 2, 832–840. [Google Scholar] [CrossRef] [PubMed]
- Belsham, G.J. Towards improvements in foot-and-mouth disease vaccine performance. Acta Veter. Scand. 2020, 62, 1–12. [Google Scholar] [CrossRef]
- Paarlberg, P.L.; Lee, J.G.; Seitzinger, A.H. Potential revenue impact of an outbreak of foot-and-mouth disease in the United States. J. Am. Veter. Med Assoc. 2002, 220, 988–992. [Google Scholar] [CrossRef] [PubMed]
- Greenwood, D.L.; Dynon, K.; Kalkanidis, M.; Xiang, S.; Plebanski, M.; Scheerlinck, J.-P.Y. Vaccination against foot-and-mouth disease virus using peptides conjugated to nano-beads. Vaccine 2008, 26, 2706–2713. [Google Scholar] [CrossRef]
- Wang, C.Y.; Chang, T.Y.; Walfield, A.M.; Ye, J.; Shen, M.; Chen, S.P.; Li, M.C.; Lin, Y.L.; Jong, M.H.; Yang, P.C.; et al. Effective synthetic peptide vaccine for foot-and-mouth disease in swine. Vaccine 2002, 20, 2603–2610. [Google Scholar] [CrossRef]
- Zhang, Z.; Pan, L.; Ding, Y.; Zhou, P.; Lv, J.; Chen, H.; Fang, Y.; Liu, X.; Chang, H.; Zhang, J.; et al. Efficacy of synthetic peptide candidate vaccines against serotype-A foot-and-mouth disease virus in cattle. Appl. Microbiol. Biotechnol. 2015, 99, 1389–1398. [Google Scholar] [CrossRef] [PubMed]
- Bastien, N.; Taylor, G.; Thomas, L.H.; Wyld, S.G.; Simard, C.; Trudel, M. Immunization with a peptide derived from the G glycoprotein of bovine respiratory syncytial virus (BRSV) reduces the incidence of BRSV-associated pneumonia in the natural host. Vaccine 1997, 15, 1385–1390. [Google Scholar] [CrossRef]
- Trudel, M.; Nadon, F.; Séguin, C.; Binz, H. Protection of BALE/c mice from respiratory syncytial virus infection by immunization with a synthetic peptide derived from the G glycoprotein. Virology 1991, 185, 749–757. [Google Scholar] [CrossRef]
- Power, U.; Plotnicky-Gilquin, H.; Huss, T.; Robert, A.; Trudel, M.; Ståhl, S.; Uhlén, M.; Nguyen, T.N.; Binz, H. Induction of Protective Immunity in Rodents by Vaccination with a Prokaryotically Expressed Recombinant Fusion Protein Containing a Respiratory Syncytial Virus G Protein Fragment. Virology 1997, 230, 155–166. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Jaberolansar, N.; Chappell, K.J.; Watterson, D.; Bermingham, I.M.; Toth, I.; Young, P.R.; Skwarczynski, M. Induction of high titred, non-neutralising antibodies by self-adjuvanting peptide epitopes derived from the respiratory syncytial virus fusion protein. Sci. Rep. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Trudel, M.; Nadon, F.; Séguin, C.; Simard, C.; Lussier, G. Experimental polyvalent ISCOMs subunit vaccine induces antibodies that neutralize human and bovine respiratory syncytial virus. Vaccine 1989, 7, 12–16. [Google Scholar] [CrossRef]
- Hägglund, S.; Hu, K.; Vargmar, K.; Poré, L.; Olofson, A.-S.; Blodörn, K.; Anderson, J.; Ahooghalandari, P.; Pringle, J.; Taylor, G.; et al. Bovine respiratory syncytial virus ISCOMs—Immunity, protection and safety in young conventional calves. Vaccine 2011, 29, 8719–8730. [Google Scholar] [CrossRef]
- Franz, J.; Hampl, J.; Stepanek, J.; Ŝmid, B. Preparation of Immunostimulating Complexes (ISCOM) Containing Bovine Herpesvirus 1 Proteins. Acta Veter. Brno 1992, 61, 37–41. [Google Scholar] [CrossRef]
- Kamstrup, S.; Roensholt, L.; Jensen, M.; Dalsgaard, K. Production of a highly immunogenic subunit ISCOM vaccine against Bovine Viral Diarrhea Virus. Vaccine 1999, 17, 1057–1064. [Google Scholar] [CrossRef]
- Crisci, E.; Bárcena, J.; Montoya, M. Virus-like particles: The new frontier of vaccines for animal viral infections. Veter. Immunol. Immunopathol. 2012, 148, 211–225. [Google Scholar] [CrossRef] [PubMed]
- Loy, J.D.; Gander, J.; Mogler, M.; Veen, R.V.; Ridpath, J.; Harris, D.H.; Kamrud, K. Development and evaluation of a replicon particle vaccine expressing the E2 glycoprotein of bovine viral diarrhea virus (BVDV) in cattle. Virol. J. 2013, 10, 35. [Google Scholar] [CrossRef] [PubMed]
- Zolnik, B.S.; González-Fernández, Á.; Sadrieh, N.; Dobrovolskaia, M.A. Minireview: Nanoparticles and the Immune System. Endocrinology 2010, 151, 458–465. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, F.; Earley, B.; Cassidy, J.P.; Markey, B.; Doherty, S.; Welsh, M.D. Comparing the immune response to a novel intranasal nanoparticle PLGA vaccine and a commercial BPI3V vaccine in dairy calves. BMC Veter. Res. 2015, 11, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, O.V.; Adair, B.M.; Welsh, M.; Earley, B. Immunogenetic responses in calves to intranasal delivery of bovine respiratory syncytial virus (BRSV) epitopes encapsulated in poly (DL-lactide-co-glycolide) microparticles. Res. Veter. Sci. 2013, 95, 786–793. [Google Scholar] [CrossRef] [PubMed]
- Cerbu, C.; Kah, M.; White, J.C.; Astete, C.E. Fate of Biodegradable Engineered Nanoparticles Used in Health Perspective. Molecules 2021, 26, 523. [Google Scholar] [CrossRef] [PubMed]
- Dhama, K.; Mahendran, M.; Gupta, P.K.; Rai, A. DNA vaccines and their applications in veterinary practice: Current perspectives. Veter. Res. Commun. 2008, 32, 341–356. [Google Scholar] [CrossRef]
- Oshop, G.; Elankumaran, S.; Heckert, R. DNA vaccination in the avian. Veter. Immunol. Immunopathol. 2002, 89, 1–12. [Google Scholar] [CrossRef]
- Dunham, S.P. The application of nucleic acid vaccines in veterinary medicine. Res. Veter. Sci. 2002, 73, 9–16. [Google Scholar] [CrossRef]
- Patial, S.; Chaturvedi, V.; Rai, A.; Saini, M.; Chandra, R.; Saini, Y.; Gupta, P.K. Virus neutralizing antibody response in mice and dogs with a bicistronic DNA vaccine encoding rabies virus glycoprotein and canine parvovirus VP2. Vaccine 2007, 25, 4020–4028. [Google Scholar] [CrossRef]
- Babiuk, S.; Babiuk, L.A. DNA vaccines. In Encyclopedia of Virology, 3rd ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 51–55. [Google Scholar]
- Nobiron, I.; Thompson, I.; Brownlie, J.; Collins, M.E. DNA vaccination against bovine viral diarrhoea virus induces humoral and cellular responses in cattle with evidence for protection against viral challenge. Vaccine 2003, 21, 2082–2092. [Google Scholar] [CrossRef]
- Ulmer, J.B.; Donnelly, J.J.; Parker, S.E.; Rhodes, G.H.; Felgner, P.L.; Dwarki, V.J.; Liu, M.A. Heterolgous protection against influenza by injection of DNA encoding a viral protein. Science 1993, 259, 1745–1749. [Google Scholar] [CrossRef] [PubMed]
- Hamers, C.; Juillard, V.; Fischer, L. DNA Vaccination against Pseudorabies Virus and Bovine Respiratory Syncytial Virus Infections of Young Animals in the Face of Maternally Derived Immunity. J. Comp. Pathol. 2007, 137, S35–S41. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.; Bruce, C.; Barbet, A.F.; Wyld, S.G.; Thomas, L.H. DNA vaccination against respiratory syncytial virus in young calves. Vaccine 2005, 23, 1242–1250. [Google Scholar] [CrossRef] [PubMed]
- Schrijver, R.S.; Langedijk, J.P.; Keil, G.M.; Middel, W.G.; Maris-Veldhuis, M.; Van Oirschot, J.T.; A Rijsewijk, F. Immunization of cattle with a BHV1 vector vaccine or a DNA vaccine both coding for the G protein of BRSV. Vaccine 1997, 15, 1908–1916. [Google Scholar] [CrossRef]
- Kornuta, C.A.; Langellotti, C.A.; Bidart, J.E.; Soria, I.; Quattrocchi, V.; Gammella, M.; Valenzuela, F.C.; Mignaqui, A.C.; Ferraris, S.; Charleston, B.; et al. A plasmid encoding the extracellular domain of CD40 ligand and Montanide™ GEL01 as adjuvants enhance the immunogenicity and the protection induced by a DNA vaccine against BoHV-1. Vaccine 2021, 39, 1007–1017. [Google Scholar] [CrossRef]
- Redding, L.; Weiner, D.B. DNA vaccines in veterinary use. Expert Rev. Vaccines 2009, 8, 1251–1276. [Google Scholar] [CrossRef]
- Jackson, N.A.C.; Kester, K.E.; Casimiro, D.; Gurunathan, S.; DeRosa, F. The promise of mRNA vaccines: A biotech and industrial perspective. npj Vaccines 2020, 5, 1–6. [Google Scholar] [CrossRef]
- Dolgin, E. The tangled history of mRNA vaccines. Nature 2021, 597, 318–324. [Google Scholar] [CrossRef] [PubMed]
- Pardi, N.; Hogan, M.; Pelc, R.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Petsch, B.; Schnee, M.; Vogel, A.; Lange, E.; Hoffmann, B.; Voss, D.; Schlake, T.; Thess, A.; Kallen, K.-J.; Stitz, L.; et al. Protective efficacy of in vitro synthesized, specific mRNA vaccines against influenza A virus infection. Nat. Biotechnol. 2012, 30, 1210–1216. [Google Scholar] [CrossRef] [PubMed]
- Alberer, M.; Gnad-Vogt, U.; Hong, H.S.; Mehr, K.T.; Backert, L.; Finak, G.; Gottardo, R.; Bica, M.A.; Garofano, A.; Koch, S.D.; et al. Safety and immunogenicity of a mRNA rabies vaccine in healthy adults: An open-label, non-randomised, prospective, first-in-human phase 1 clinical trial. Lancet 2017, 390, 1511–1520. [Google Scholar] [CrossRef]
- Batra, S.A.; Shanthalingam, S.; Donofrio, G.; Haldorson, G.J.; Chowdhury, S.; White, S.N.; Srikumaran, S. Immunization of bighorn sheep against Mannheimia haemolytica with a bovine herpesvirus 1-vectored vaccine. Vaccine 2017, 35, 1630–1636. [Google Scholar] [CrossRef]
- Bilge, S.; Touraj, D.; Farzani, A.; Dogan, F.; Akkutay, Z.; Aykut, Y. Development of a BoHV—4 viral vector expressing tgD of BoHV—1 and evaluation of its immunogenicity in mouse model. Braz. J. Microbiol. 2021, 52, 1119–1133. [Google Scholar] [CrossRef]
- Chowdhury, S.I.; Pannhorst, K.; Sangewar, N.; Pavulraj, S.; Wen, X.; Stout, R.W.; Mwangi, W.; Paulsen, D.B. BoHV-1-Vectored BVDV-2 Subunit Vaccine Induces BVDV Cross-Reactive Cellular Immune Responses and Protects against BVDV-2 Challenge. Vaccines 2021, 9, 46. [Google Scholar] [CrossRef] [PubMed]
- Taylor, G.; Thomas, L.H.; Wyld, S.G.; Wertz, G.W.; Hardy, R.; Cook, R.S.; Furze, J.M.; Lerch, R. Recombinant vaccinia viruses expressing the F, G or N, but not the M2, protein of bovine respiratory syncytial virus (BRSV) induce resistance to BRSV challenge in the calf and protect against the development of pneumonic lesions. J. Gen. Virol. 1997, 78, 3195–3206. [Google Scholar] [CrossRef]
- Antonis, A.F.; van der Most, R.G.; Suezer, Y.; Stockhofe-Zurwieden, N.; Daus, F.; Sutter, G.; Schrijver, R.S. Vaccination with recombinant modified vaccinia virus Ankara expressing bovine respiratory syncytial virus (bRSV) proteins protects calves against RSV challenge. Vaccine 2007, 25, 4818–4827. [Google Scholar] [CrossRef]
- Nelson, G.; Marconi, P.; Periolo, O.; La Torre, J.; Alvarez, M.A. Immunocompetent truncated E2 glycoprotein of bovine viral diarrhea virus (BVDV) expressed in Nicotiana tabacum plants: A candidate antigen for new generation of veterinary vaccines. Vaccine 2012, 30, 4499–4504. [Google Scholar] [CrossRef]
- Faye, L.; Gomord, V. Editorial Success stories in molecular farming—A brief overview. Plant Biotechnol. J. 2010, 8, 525–528. [Google Scholar] [CrossRef]
- Rybicki, E.P. Plant-made vaccines for humans and animals. Plant Biotechnol. J. 2010, 8, 620–637. [Google Scholar] [CrossRef]
- Taylor, G.; Wyld, S.G.; Keil, G.; I Morrison, W.; Hensen, E.; Cook, R.S.; A Rijsewijk, F.; Gaddum, R.M.; Thomas, L.H.; Van Oirschot, J.T. Resistance to bovine respiratory syncytial virus (BRSV) induced in calves by a recombinant bovine herpesvirus-1 expressing the attachment glycoprotein of BRSV. J. Gen. Virol. 1998, 79, 1759–1767. [Google Scholar] [CrossRef]
- Briggs, R.E.; Tabatabai, L.B.; Tatum, F.M. Microbial Pathogenesis Mucosal and parenteral vaccination against pneumonic pasteurellosis in cattle with a modi fi ed-live in-frame lktA deletion mutant of Mannheimia haemolytica. Microb. Pathog. 2012, 52, 302–309. [Google Scholar] [CrossRef] [PubMed]
- AFBI/DAERA. All-Island Disease Surveillance Report: A Joint AFBI/DAFM Veterinary Laboratories Report; AFBI: Belfast, UK, 2016. [Google Scholar]
- Pretty, J.; Sutherland, W.J.; Ashby, J.; Auburn, J.; Baulcombe, D.; Bell, M.; Bentley, J.; Bickersteth, S.; Brown, K.; Burke, J.; et al. The top 100 questions of importance to the future of global agriculture. Int. J. Agric. Sustain. 2010, 8, 219–236. [Google Scholar] [CrossRef]
- Daly, P.A. Agricultural employment: Has the decline ended? Mon. Labor Rev. 1981, 104, 11–17. [Google Scholar]
- Roser, M. Employment in agriculture. Our World in Data. 2013. Available online: https://ourworldindata.org/employment-in-agriculture (accessed on 27 September 2021).
- Schneider, U.; Havlík, P.; Schmid, E.; Valin, H.; Mosnier, A.; Obersteiner, M.; Böttcher, H.; Skalský, R.; Balkovič, J.; Sauer, T.; et al. Impacts of population growth, economic development, and technical change on global food production and consumption. Agric. Syst. 2011, 104, 204–215. [Google Scholar] [CrossRef]
- Department for Environment Food and Rural Affairs. Total Income from Farming in the United Kingdom, First Estimate for 2020; Department for Environment Food and Rural Affairs: Bristol, UK, 2021.
- Allen, M. Northern Ireland’s Agri-Food Sector—Background and possible ‘Brexit’ Considerations. 2016. Available online: http://www.niassembly.gov.uk/globalassets/documents/raise/publications/2016-2021/2016/aera/6616.pdf (accessed on 16 July 2017).
- Playfair, D. Northern Ireland Agri-Food Sector—Agriculture, Fishing and Forestry. 2018. Available online: https://www.daera-ni.gov.uk/sites/default/files/publications/daera/Key%20Statistics%20for%202018.pdf (accessed on 22 March 2019).
- Smith, R.A.; Step, D.L.; Woolums, A.R. Bovine Respiratory Disease Looking Back and Looking Forward, What Do We See? Veter. Clin. Food Anim. 2020, 36, 239–251. [Google Scholar] [CrossRef]

| Viruses | Bacteria | Mycoplasma spp. |
|---|---|---|
| Bovine herpesvirus-1 (BHV-1) | Mannheimia (Pasteurella) haemolytica | Mycoplasma bovis |
| Bovine respiratory syncytial virus (BRSV) | Pasteurella multocida | Ureaplasma spp. |
| Bovine viral diarrhoea virus (BVDV) | Histophilus (Haemophilus) somni | |
| Bovine parainfluenza virus type-3 (BPIV-3) | ||
| Bovine adenovirus (BAV) | ||
| Bovine coronavirus (BCoV) |
| Advantages |
| More neutral pH and lower levels of enzymatic activity than digestive tract |
| Prime neonatal calf in the presence of MDA |
| Needle-free/non-invasive |
| Induction of systemic and mucosal immunity |
| User-friendly (potential use in herds/developing world/remote farms) |
| Disadvantages |
| Rapid clearance of low affinity antigens |
| Potential antigen loss during inoculation (impact on cost) |
| Inefficient uptake |
| Lack of compatible adjuvants for mucosal vaccines |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lemon, J.L.; McMenamy, M.J. A Review of UK-Registered and Candidate Vaccines for Bovine Respiratory Disease. Vaccines 2021, 9, 1403. https://doi.org/10.3390/vaccines9121403
Lemon JL, McMenamy MJ. A Review of UK-Registered and Candidate Vaccines for Bovine Respiratory Disease. Vaccines. 2021; 9(12):1403. https://doi.org/10.3390/vaccines9121403
Chicago/Turabian StyleLemon, Joanne L., and Michael J. McMenamy. 2021. "A Review of UK-Registered and Candidate Vaccines for Bovine Respiratory Disease" Vaccines 9, no. 12: 1403. https://doi.org/10.3390/vaccines9121403
APA StyleLemon, J. L., & McMenamy, M. J. (2021). A Review of UK-Registered and Candidate Vaccines for Bovine Respiratory Disease. Vaccines, 9(12), 1403. https://doi.org/10.3390/vaccines9121403