
The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis




Abstract
1. Introduction
1.1. Invasive Candidiasis
1.2. Innate Immunity in Invasive Candidiasis
1.3. Adaptive Immunity in Invasive Candidiasis: Cellular Responses
1.4. Adaptive Immunity in Invasive Candidiasis: Humoral Responses
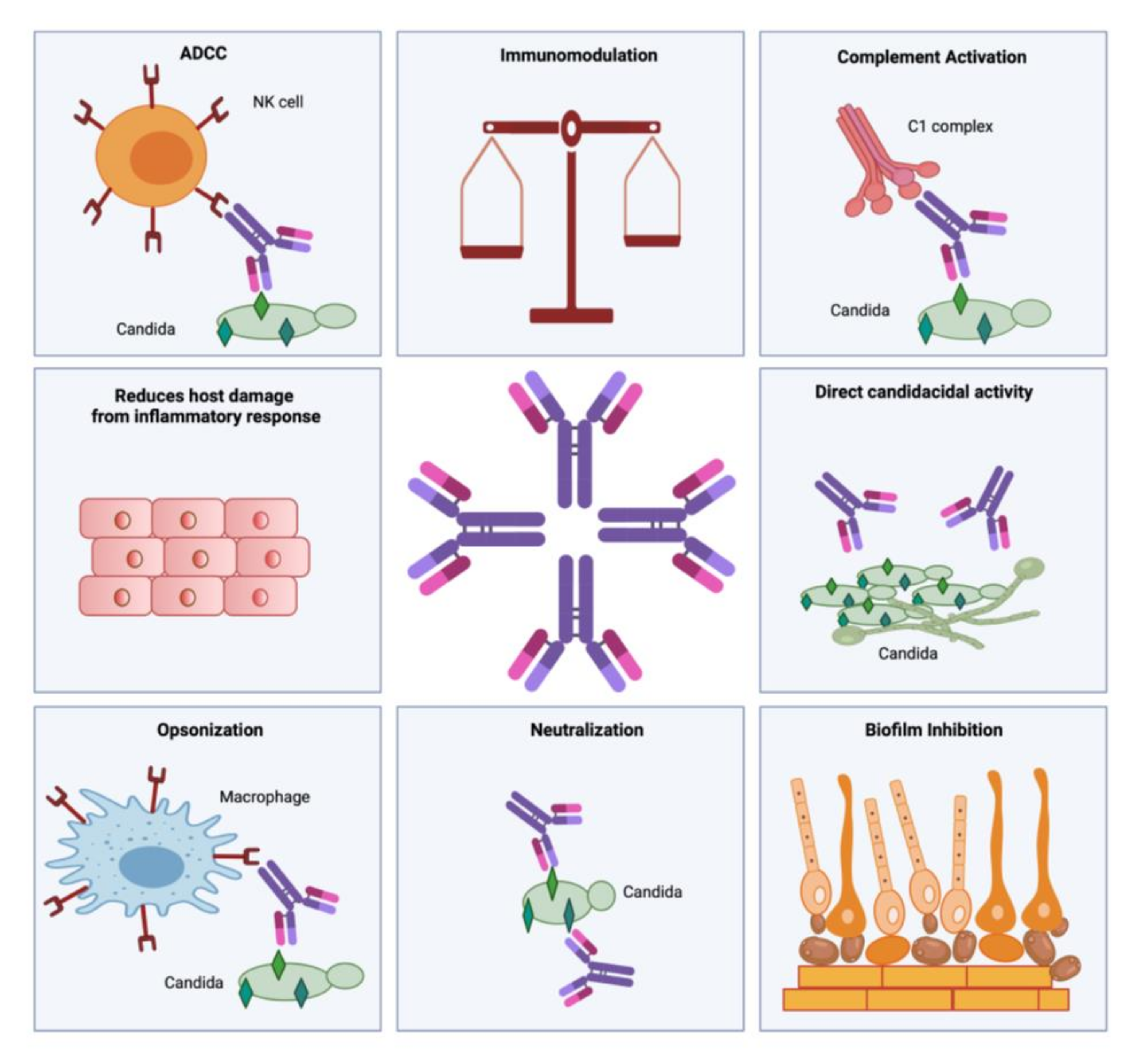
1.4.1. B-Cell Responses in Invasive Candidiasis:
1.4.2. Antibody Responses in Invasive Candidiasis
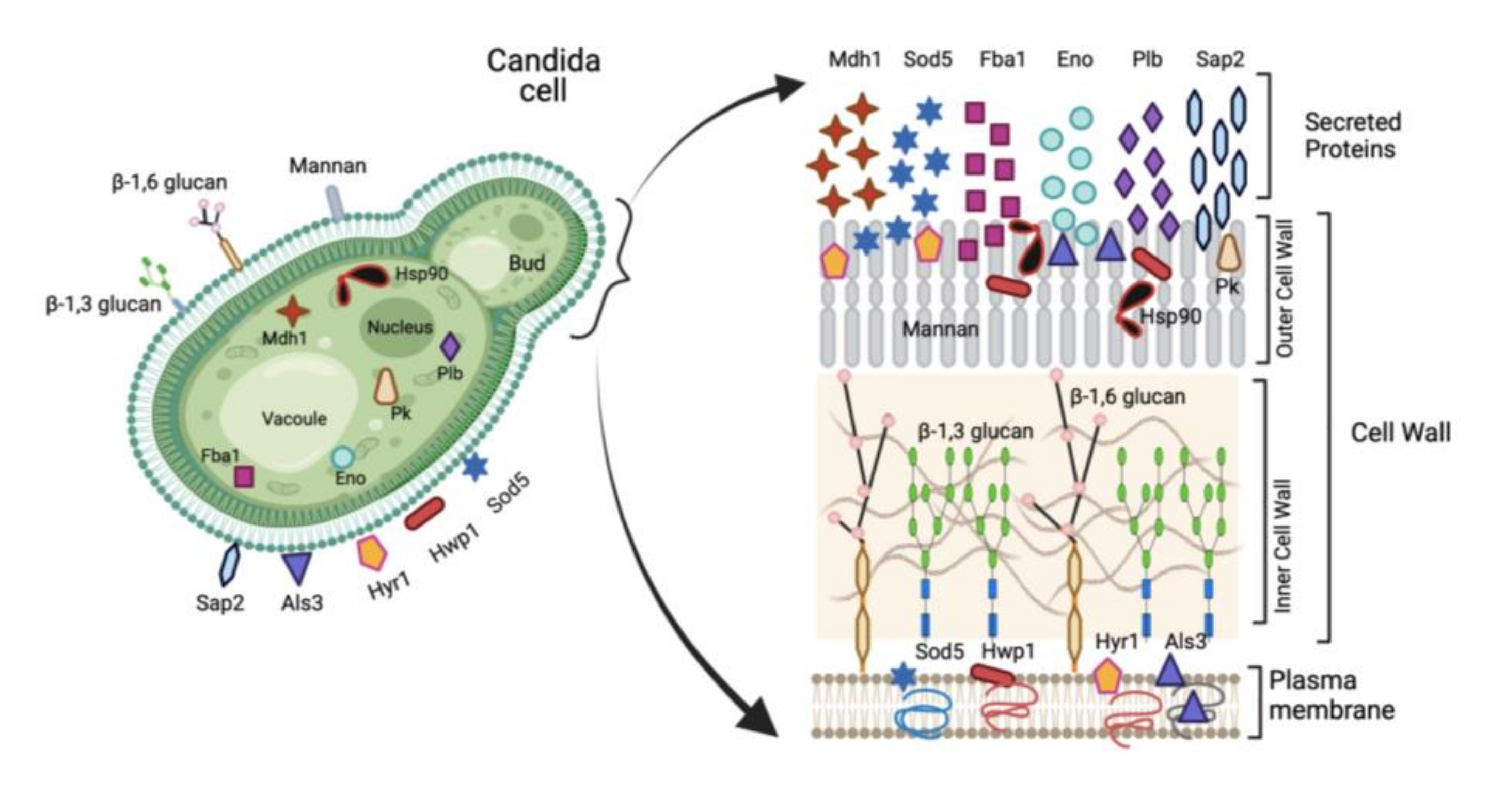
2. Vaccine Candidates in Invasive Candidiasis
2.1. Humoral Immune Responses to Mannan Polysaccharide
2.2. Humoral Immune Responses to β-Glucan Polysaccharide
2.3. Humoral Immune Responses to Laminarin
2.4. Humoral Immune Responses to Hsp90
2.5. Humoral Immune Responses to Agglutinin-Like Sequence 3 (Als3)
2.6. Humoral Immune Responses to Secreted Aspartyl Proteinase 2 (Sap2)
2.7. Humoral Immune Responses to Hyphally Regulated Protein 1 (Hyr1)
2.8. Humoral Immune Responses to Hyphal Wall Protein 1 (Hwp1)
2.9. Humoral Immune Responses to Enolase (Eno)
2.10. Humoral Immune Responses to Phospholipase B (PLB)
2.11. Humoral Immune Responses to Fructose-Bisphosphate Aldolase (Fba1)
2.12. Humoral Immune Responses to Pyruvate Kinase (Pk)
2.13. Humoral Immune Responses to Superoxide Dismutase (Sod5)
2.14. Humoral Immune Responses to Malate Dehydrogenase (Mdh1)
3. Conclusions
4. Future Perspective
- The serious threat posed by invasive fungal infections is a persisting problem and new therapeutic options are urgently required including vaccine/s and/or antibody-based therapies. Challenges associated with the extensive use of antifungal drugs like antifungal drug-associated toxicity and emergence of antifungal drug resistance, along with increased incidences of infections due to multi-drug resistant non-albicans Candida species merits the need for developing alternate immunotherapies against invasive candidiasis.
- Due to the promising potential of humoral immunity in conferring antifungal protection, identification of fungal antigens which elicit protective antibodies will be crucial for designing effective multi-valent and/or multi-epitope-based anti-Candida vaccine/s.
- There is a need to synthesize novel peptides and oligosaccharides using a combination of protective antigens that could be used to make multi-epitope conjugate vaccines including B-cell and T-cell epitopes for generating memory. Although few experimental vaccines have been designed, extensive animal-based studies are lacking.
- Additionally, identification, isolation and characterization of protective anti-Candida monoclonal antibodies along with elucidation of their protection mechanisms, will be beneficial in generating more precise antibody-based therapies. More clinical trials are needed to validate passive antibody therapy against invasive candidiasis.
- Novel cytokine-based adjunctive immunotherapies and immunomodulators should be explored for the treatment of invasive candidiasis. More studies are needed to demonstrate the feasibility of such immunotherapies for improving the prognosis of invasive candidiasis.
- Synergistic use of monoclonal antibodies in combination with anti-fungal drugs has been seen to impart cooperative protection against Candida. More studies are needed to assess the safety and efficacy of combination therapies.
- Susceptibility of immunocompromised individuals having deficiencies in an immune cell repertoire to invasive candidiasis justifies future research for characterization of the protective B-cell and T-cell repertoires, aiding the development of novel vaccination strategies.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- François, L.M.; Duncan, W.; Bernhard, H. Candida albicans pathogenicity mechanisms. Virulence 2013, 4, 119–128. [Google Scholar] [CrossRef]
- Warnock, D.W. Trends in the epidemiology of invasive fungal infections. Jpn. J. Med. Mycol. 2007, 48, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Calandra, T.; Roberts, J.A.; Antonelli, M.; Bassetti, M.; Vincent, J.L. Diagnosis and management of invasive candidiasis in the ICU: An updated approach to an old enemy. Crit. Care 2016, 20, 125. [Google Scholar] [CrossRef]
- Pappas, P.G.; Lionakis, M.S.; Arendrup, M.C.; Ostrosky-Zeichner, L.; Kullberg, B.J. Invasive candidiasis. Nat. Rev. Dis. Prim. 2018, 4, 1–20. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, B.J.; Arendrup, M.C. Invasive Candidiasis. N. Engl. J. Med. 2015, 373, 1445–1456. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.; Alfouzan, W. Candida auris: Epidemiology, diagnosis, pathogenesis, antifungal susceptibility, and infection control measures to combat the spread of infections in healthcare facilities. Microorganisms 2021, 9, 807. [Google Scholar] [CrossRef]
- Gintjee, T.J.; Donnelley, M.A.; Thompson, G.R. Aspiring Antifungals: Review of Current Antifungal Pipeline Developments. J. Fungi 2020, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, S.; Sae-Tia, S.; Fries, B.C. Candidiasis and mechanisms of antifungal resistance. Antibiotics 2020, 9, 312. [Google Scholar] [CrossRef]
- Papon, N.; Courdavault, V.; Clastre, M.; Bennett, R.J.; Nac, N.-C. Emerging and Emerged Pathogenic Candida Species: Beyond the Candida albicans Paradigm. PLoS Pathog. 2013, 9, e1003550. [Google Scholar] [CrossRef]
- Whibley, N.; Gaffen, S.L. Beyond Candida albicans: Mechanisms of immunity to non-albicans Candida species. Cytokine 2015, 76, 42–52. [Google Scholar] [CrossRef]
- Nami, S.; Aghebati-Maleki, A.; Morovati, H.; Aghebati-Maleki, L. Current antifungal drugs and immunotherapeutic approaches as promising strategies to treatment of fungal diseases. Biomed. Pharmacother. 2019, 110, 857–868. [Google Scholar] [CrossRef] [PubMed]
- Lionakis, M.S. New insights into innate immune control of systemic candidiasis. Med. Mycol. 2014, 52, 555–564. [Google Scholar] [CrossRef]
- Medici, N.P.; Poeta, M. Del New insights on the development of fungal vaccines: From immunity to recent challenges. Mem. Inst. Oswaldo Cruz 2015, 110, 966–973. [Google Scholar] [CrossRef] [PubMed]
- Vonk, A.G.; Netea, M.G.; Van Der Meer, J.W.M.; Kullberg, B.J. Host defence against disseminated Candida albicans infection and implications for antifungal immunotherapy. Expert Opin. Biol. Ther. 2006, 6, 891–903. [Google Scholar] [CrossRef] [PubMed]
- Harpf, V.; Rambach, G.; Würzner, R.; Lass-Flörl, C.; Speth, C. Candida and Complement: New Aspects in an Old Battle. Front. Immunol. 2020, 11, 1–13. [Google Scholar] [CrossRef]
- Tsoni, S.V.; Kerrigan, A.M.; Marakalala, M.J.; Srinivasan, N.; Duffield, M.; Taylor, P.R.; Botto, M.; Steele, C.; Brown, G.D. Complement C3 plays an essential role in the control of opportunistic fungal infections. Infect. Immun. 2009, 77, 3679–3685. [Google Scholar] [CrossRef]
- Brown, G.D.; Denning, D.W.; Gow, N.A.R.; Levitz, S.M.; Netea, M.G.; White, T.C. Hidden killers: Human fungal infections. Sci. Transl. Med. 2012, 4, 165rv13. [Google Scholar] [CrossRef]
- Jones-Carson, J.; Vazquez-Torres, F.A.; Balish, E. B cell-independent selection of memory T cells after mucosal immunization with Candida albicans. J. Immunol. 1997, 158, 4328–4335. [Google Scholar]
- Qian, Q.; Jutila, M.A.; Van Rooijen, N.; Cutler, J.E. Elimination of mouse splenic macrophages correlates with increased susceptibility to experimental disseminated candidiasis. J. Immunol. 1994, 152, 5000–5008. [Google Scholar]
- Netea, M.G.; Gijzen, K.; Coolen, N.; Verschueren, I.; Figdor, C.; Van Der Meer, J.W.M.; Torensma, R.; Kullberg, B.J. Human dendritic cells are less potent at killing Candida albicans than both monocytes and macrophages. Microbes Infect. 2004, 6, 985–989. [Google Scholar] [CrossRef]
- Ngo, L.Y.; Kasahara, S.; Kumasaka, D.K.; Knoblaugh, S.E.; Jhingran, A.; Hohl, T.M. Inflammatory monocytes mediate early and organ-specific innate defense during systemic candidiasis. J. Infect. Dis. 2014, 209, 109–119. [Google Scholar] [CrossRef]
- Netea, M.G.; van Tits, L.J.; Curfs, J.H.; Amiot, F.; Meis, J.F.; van der Meer, J.W.; Kullberg, B.J. Increased susceptibility of TNF-alpha lymphotoxin-alpha double knockout mice to systemic candidiasis through impaired recruitment of neutrophils and phagocytosis of Candida albicans. J Immunol. 1999, 163, 1498–1505. [Google Scholar]
- Netea, M.G.; Joosten, L.A.B.; Van Der Meer, J.W.M.; Kullberg, B.J.; Van De Veerdonk, F.L. Immune defence against Candida fungal infections. Nat. Rev. Immunol. 2015, 15, 630–642. [Google Scholar] [CrossRef]
- Newman, S.L.; Holly, A. Candida albicans is phagocytosed, killed, and processed for antigen presentation by human dendritic cells. Infect. Immun. 2001, 69, 6813–6822. [Google Scholar] [CrossRef]
- Romagnoli, G.; Nisini, R.; Chiani, P.; Mariotti, S.; Teloni, R.; Cassone, A.; Torosantucci, A. The interaction of human dendritic cells with yeast and germ-tube forms of. J. Leukoc. Biol. 2004, 75, 117–126. [Google Scholar] [CrossRef]
- Lionakis, M.S.; Netea, M.G. Candida and Host Determinants of Susceptibility to Invasive Candidiasis. PLoS Pathog. 2013, 9, e1003079. [Google Scholar] [CrossRef]
- Gardner, A. Diagnosing fungal infections in neutropenic patients. Clin. J. Oncol. Nurs. 2007, 11, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Uppuluri, P.; Singh, S.; Alqarihi, A.; Schmidt, C.S.; Hennessey, J.P.; Yeaman, M.R.; Filler, S.G.; Edwards, J.E.; Ibrahim, A.S. Human Anti-Als3p antibodies are surrogate markers of NDV-3A vaccine efficacy against recurrent vulvovaginal candidiasis. Front. Immunol. 2018, 9, 1349. [Google Scholar] [CrossRef] [PubMed]
- Shukla, M.; Rohatgi, S. Vaccination with secreted aspartyl proteinase 2 protein from candida parapsilosis can enhance survival of mice during c. tropicalis-mediated systemic candidiasis. Infect. Immun. 2020, 88, e00312–e00320. [Google Scholar] [CrossRef] [PubMed]
- Quintin, J.; Voigt, J.; van der Voort, R.; Jacobsen, I.D.; Verschueren, I.; Hube, B.; Giamarellos-Bourboulis, E.J.; van der Meer, J.W.M.; Joosten, L.A.B.; Kurzai, O.; et al. Differential role of NK cells against Candida albicans infection in immunocompetent or immunocompromised mice. Eur. J. Immunol. 2014, 44, 2405–2414. [Google Scholar] [CrossRef]
- Voigt, J.; Malone, D.F.G.; Dias, J.; Leeansyah, E.; Björkström, N.K.; Ljunggren, H.G.; Gröbe, L.; Klawonn, F.; Heyner, M.; Sandberg, J.K.; et al. Proteome analysis of human CD56 neg NK cells reveals a homogeneous phenotype surprisingly similar to CD56 dim NK cells. Eur. J. Immunol. 2018, 48, 1456–1469. [Google Scholar] [CrossRef]
- Schmidt, S.; Zimmermann, S.Y.; Tramsen, L.; Koehl, U.; Lehrnbecher, T. Natural killer cells and antifungal host response. Clin. Vaccine Immunol. 2013, 20, 452–458. [Google Scholar] [CrossRef][Green Version]
- Conti, H.R.; Gaffen, S.L. Host responses to Candida albicans: Th17 cells and mucosal candidiasis. Microbes Infect. 2010, 12, 518–527. [Google Scholar] [CrossRef] [PubMed]
- Van De Veerdonk, F.L.; Netea, M.G. T-cell subsets and antifungal host defenses. Curr. Fungal Infect. Rep. 2010, 4, 238–243. [Google Scholar] [CrossRef] [PubMed]
- Richardson, J.P.; Moyes, D.L. Adaptive immune responses to Candida albicans infection. Virulence 2015, 6, 327–337. [Google Scholar] [CrossRef]
- Hernández-Santos, N.; Gaffen, S.L. Th17 cells in immunity to Candida albicans. Cell Host Microbe 2012, 11, 425–435. [Google Scholar] [CrossRef]
- Romani, L.; Zelante, T.; Palmieri, M.; Napolioni, V.; Picciolini, M.; Velardi, A.; Aversa, F.; Puccetti, P. The cross-talk between opportunistic fungi and the mammalian host via microbiota’s metabolism. Semin. Immunopathol. 2015, 37, 163–171. [Google Scholar] [CrossRef]
- Huang, W.; Na, L.; Fidel, P.L.; Schwarzenberger, P. Requirement of interleukin-17A for systemic anti-Candida albicans host defense in mice. J. Infect. Dis. 2004, 190, 624–631. [Google Scholar] [CrossRef]
- Van De Veerdonk, F.L.; Kullberg, B.J.; Verschueren, I.C.; Hendriks, T.; Van Der Meer, J.W.M.; Joosten, L.A.B.; Netea, M.G. Differential effects of IL-17 pathway in disseminated candidiasis and zymosan-induced multiple organ failure. Shock 2010, 34, 407–411. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Ibrahim, A.S.; Xu, X.; Farber, J.M.; Avanesian, V.; Baquir, B.; French, S.W.; Edwards, J.E., Jr.; Spellberg, B. Th1-Th17 Cells Mediate Protective Adaptive Immunity against Staphylococcus aureus and Candida albicans Infection in Mice. PLoS Pathog. 2009, 5, e1000703. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Spellberg, B.J.; Avanesian, V.; Fu, Y.; Edwards, J.E. The anti-candida vaccine based on the recombinant N-terminal domain of Als1p is broadly active against disseminated candidiasis. Infect. Immun. 2006, 74, 3039–3041. [Google Scholar] [CrossRef]
- Coker, L.A.; Mercadal, C.M.; Rouse, B.T.; Moore, R.N. Differential effects of CD4+ and CD8+ cells in acute, systemic murine candidosis. J. Leukoc. Biol. 1992, 51, 305–306. [Google Scholar] [CrossRef]
- Ashman, R.B.; Fulurija, A.; Papadimitriou, J.M. Both CD4+ and CD8+ lymphocytes reduce the severity of tissue lesions in murine systemic candidiasis, and CD4+ cells also demonstrate strain-specific immunopathological effects. Microbiology 1999, 145, 1631–1640. [Google Scholar] [CrossRef]
- Mody, C.H.; Oykhman, P. Direct microbicidal activity of cytotoxic T-lymphocytes. J. Biomed. Biotechnol. 2010, 2010, 249482. [Google Scholar] [CrossRef]
- Daniel Gozalbo, M.L.G. IFN-g in Candida albicans infections. Front. Biosci. 2009, 14, 1970–1978. [Google Scholar] [CrossRef] [PubMed]
- Hamad, M. Innate and adaptive antifungal immune responses: Partners on an equal footing. Mycoses 2012, 55, 205–217. [Google Scholar] [CrossRef] [PubMed]
- LeibundGut-Landmann, S.; Wüthrich, M.; Hohl, T.M. Immunity to fungi. Curr. Opin. Immunol. 2012, 24, 449–458. [Google Scholar] [CrossRef]
- Antachopoulos, C.; Roilides, E. Cytokines and fungal infections. Br. J. Haematol. 2005, 129, 583–596. [Google Scholar] [CrossRef]
- Van de Veerdonk, F.L.; Kullberg, B.J.; Netea, M.G. Adjunctive immunotherapy with recombinant cytokines for the treatment of disseminated candidiasis. Clin. Microbiol. Infect. 2012, 18, 112–119. [Google Scholar] [CrossRef] [PubMed]
- Pathakumari, B.; Liang, G.; Liu, W. Immune defence to invasive fungal infections: A comprehensive review. Biomed. Pharmacother. 2020, 130, 110550. [Google Scholar] [CrossRef]
- Balish, E.; Wagner, R.D.; Pierson, C.; Warner, T. Candidiasis in interferon-gamma knockout (IFN-gamma-/-) mice. J. Infect. Dis. 1998, 178, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Roilides, E.; Holmes, A.; Blake, C.; Pizzo, P.A.; Walsh, T.J. Effects of granulocyte colony-stimulating factor and interferon-γ on antifungal activity of human polymorphonuclear neutrophils against pseudohyphae of different medically important Candida species. J. Leukoc. Biol. 1995, 57, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Londono, L.P.; Jones, H.B.; Vie, A.T.J.; McPheat, W.L.; Booth, G.; Gao, X.M.; Dougan, G. Characterisation of Candida albicans infections of haematogenous and mucosal origin in mice lacking the interferon γ receptor protein. FEMS Immunol. Med. Microbiol. 2000, 27, 117–125. [Google Scholar] [CrossRef]
- Romani, L.; Cenci, E.; Mencacci, A.; Spaccapelo, R.; Grohmann, U.; Puccetti, P.; Bistoni, F. Gamma interferon modifies CD4+ subset expression in murine candidiasis. Infect. Immun. 1992, 60, 4950–4952. [Google Scholar] [CrossRef] [PubMed]
- Delsing, C.E.; Gresnigt, M.S.; Leentjens, J.; Preijers, F.; Frager, F.A.; Kox, M.; Monneret, G.; Venet, F.; Bleeker-Rovers, C.P.; van de Veerdonk, F.L.; et al. Interferon-gamma as adjunctive immunotherapy for invasive fungal infections: A case series. BMC Infect. Dis. 2014, 14, 1–12. [Google Scholar] [CrossRef]
- Netea, M.G.; Vonk, A.G.; van den Hoven, M.; Verschueren, I.; Joosten, L.A.; van Krieken, J.H.; van den Berg, W.B.; Van der Meer, J.W.K.B. Differential role of IL-18 and IL-12 in the host defense against disseminated Candida albicans infection. Eur. J. Immunol. 2003, 33, 3409–3417. [Google Scholar] [CrossRef]
- Káposzta, R.; Maródi, L.; Da Silva, R.P.; Gordon, S. Cellular mechanisms of phagocytosis of Candida by murine macrophages. Adv. Cell. Mol. Biol. Membr. Organelles 1999, 6, 317–331. [Google Scholar] [CrossRef]
- Mencacci, A.; Cenci, E.; Bacci, A.; Montagnoli, C.; Bistoni, F.; Romani, L. Cytokines in candidiasis and aspergillosis. Curr. Pharm. Biotechnol. 2000, 1, 235–251. [Google Scholar] [CrossRef] [PubMed]
- Whibley, N.; Jaycox, J.R.; Reid, D.; Garg, A.V.; Taylor, J.A.; Clancy, C.J.; Nguyen, M.H.; Biswas, P.S.; McGeachy, M.J.; Brown, G.D.; et al. Delinking CARD9 and IL-17: CARD9 Protects against Candida tropicalis Infection through a TNF-α–Dependent, IL-17–Independent Mechanism. J. Immunol. 2015, 195, 3781–3792. [Google Scholar] [CrossRef]
- Vonk, A.G.; Netea, M.G.; Van Krieken, J.H.; Iwakura, Y.; Van Der Meer, J.W.M.; Kullberg, B.J. Endogenous Interleukin (IL)–1 a and IL-1 b Are Crucial for Host Defense against Disseminated Candidiasis. J. Infect. Dis. 2006, 193, 1419–1426. [Google Scholar] [CrossRef] [PubMed]
- Chin, V.K.; Foong, K.J.; Maha, A.; Rusliza, B.; Norhafizah, M.; Chong, P.P. Early expression of local cytokines during systemic Candida albicans infection in a murine intravenous challenge model. Biomed. Rep. 2014, 2, 869–874. [Google Scholar] [CrossRef] [PubMed]
- Beno, D.W.A.; Stover, A.G.; Mathews, H.L. Growth inhibition of Candida albicans hyphae by CD8+ lymphocytes. J. Immunol. 1995, 154, 5273–5281. [Google Scholar] [PubMed]
- Romani, L. Immunity to Candida albicans: Th1, Th2 cells and beyond. Curr. Opin. Microbiol. 1999, 2, 363–367. [Google Scholar] [CrossRef]
- Van Enckevort, F.H.; Netea, M.G.; Hermus, A.R.; Sweep, C.G.; Meis, J.F.; Van der Meer, J.W.; Kullberg, B.J. Increased susceptibility to systemic candidiasis in interleukin-6 deficient mice. Med. Mycol. 1999, 37, 419–426. [Google Scholar] [CrossRef]
- Unsinger, J.; Burnham, C.D.; Mcdonough, J.; Morre, M.; Prakash, P.S.; Caldwell, C.C.; Dunne, W.M.; Hotchkiss, R.S. Interleukin-7 Ameliorates Immune Dysfunction and Improves Survival in a 2-Hit Model of Fungal Sepsis. J. Infect. Dis. 2012, 206, 606–616. [Google Scholar] [CrossRef]
- Romani, L. Immunity to fungi. New Insights Med. Mycol. 2007, 24, 1–18. [Google Scholar] [CrossRef]
- Romani, L.; Mencacci, A.; Tonnetti, L.; Spaccapelo, R.; Cenci, E.; Wolf, S.; Puccetti, P.; Bistoni, F. Interleukin-12 but not interferon-gamma production correlates with induction of T helper type-1 phenotype in murine candidiasis. Eur. J. Immunol. 1994, 24, 909–915. [Google Scholar] [CrossRef]
- Stuyt, R.J.L.; Netea, M.G.; Verschueren, I.; Fantuzzi, G.; Dinarello, C.A.; Van Der Meer, J.W.M.; Kullberg, B.J. Role of Interleukin-18 in Host Defense against Disseminated Candida albicans Infection. Infect. Immun. 2002, 70, 3284–3286. [Google Scholar] [CrossRef]
- Whitney, P.G.; Bär, E.; Osorio, F.; Rogers, N.C.; Schraml, B.U.; Deddouche, S.; LeibundGut-Landmann, S.; Reis e Sousa, C. Syk Signaling in Dendritic Cells Orchestrates Innate Resistance to Systemic Fungal Infection. PLoS Pathog. 2014, 10, e1004276. [Google Scholar] [CrossRef]
- Nur, S.; Sparber, F.; Lemberg, C.; Guiducci, E.; Schweizer, T.A.; Zwicky, P.; Becher, B.; LeibundGut-Landmann, S. IL-23 supports host defense against systemic Candida albicans infection by ensuring myeloid cell survival. PLoS Pathog. 2019, 15, e1008115. [Google Scholar] [CrossRef]
- Li, M.; Li, C.; Wu, X.; Chen, T.; Ren, L.; Xu, B.; Cao, J. Microbiota-driven interleukin-17 production provides immune protection against invasive candidiasis. Crit. Care 2020, 24, 1–14. [Google Scholar] [CrossRef]
- Huppler, A.R.; Bishu, S.; Gaffen, S.L. Mucocutaneous candidiasis: The IL-17 pathway and implications for targeted immunotherapy. Arthritis Res. Ther. 2012, 14, 217. [Google Scholar] [CrossRef]
- Conti, H.R.; Gaffen, S.L. IL-17–Mediated Immunity to the Opportunistic Fungal Pathogen Candida albicans. J. Immunol. 2015, 195, 780–788. [Google Scholar] [CrossRef]
- Haraguchi, N.; Ishii, Y.; Morishima, Y.; Yoh, K.; Matsuno, Y.; Kikuchi, N.; Sakamoto, T.; Takahashi, S.; Hizawa, N. Impairment of host defense against disseminated candidiasis in mice overexpressing GATA-3. Infect. Immun. 2010, 78, 2302–2311. [Google Scholar] [CrossRef]
- Mencacci, A.; Del Sero, G.; Cenci, E.; D’Ostiani, C.F.; Bacci, A.; Montagnoli, C.; Kopf, M.; Romani, L. Endogenous interleukin 4 is required for development of protective CD4+ T helper type 1 cell responses to Candida albicans. J. Exp. Med. 1998, 187, 307–317. [Google Scholar] [CrossRef] [PubMed]
- Spaccapelo, R.; Romani, L.; Tonnetti, L.; Cenci, E.; Mencacci, A.; Del Sero, G.; Tognellini, R.; Reed, S.G.; Puccetti, P.; Bistoni, F. TGF-beta is important in determining the in vivo patterns of susceptibility or resistance in mice infected with Candida albicans. J. Immunol. 1995, 155, 1349–1360. [Google Scholar] [PubMed]
- Sinha, B.K.; Prasad, S.; Monga, D.P. Studies of the role of B-cells in the resistance of mice to experimental candidiasis. Zentralbl Bakteriol. Mikrobiol. Hyg. A 1987, 266, 316–322. [Google Scholar] [CrossRef]
- Carrow, E.W.; Hector, R.F.; Domer, J.E. Immunodeficient CBA/N mice respond effectively to Candida albicans. Clin. Immunol. Immunopathol. 1984, 33, 371–380. [Google Scholar] [CrossRef]
- Bistoni, F.; Verducci, G.; Perito, S.; Vecchiarelli, A.; Puccetti, P.; Marconi, P.; Cassone, A. Immunomodulation by a low-virulence, agerminative variant of candida albicans. Further evidence for macrophage activation as one of the effector mechanisms of nonspecific anti-infectious protection. Med. Mycol. 1988, 26, 285–299. [Google Scholar] [CrossRef]
- Balish, E.; Jensen, J.; Warner, T. Resistance of SCID Mice to Candida albicans Administered Intravenously or Colonizing the Gut: Role of Polymorphonuclear Leukocytes and Macrophages. J. Infect. Dis. 1993, 167, 912–919. [Google Scholar] [CrossRef]
- Jensen, J.; Warner, T.B.E. The role of phagocytic cells in resistance to disseminated candidiasis in granulocytopenic mice. J. Infect. Dis. 1994, 170, 900–905. [Google Scholar] [CrossRef]
- Wagner, R.D.; Vazquez-Torres, A.; Jones-Carson, J.; Warner, T.; Balish, E. B cell knockout mice are resistant to mucosal and systemic candidiasis of endogenous origin but susceptible to experimental systemic candidiasis. J. Infect. Dis. 1996, 174, 589–597. [Google Scholar] [CrossRef]
- Santoni, G.; Boccanera, M.; Adriani, D.; Lucciarini, R.; Amantini, C.; Morrone, S.; Cassone, A.; De Bernardis, F. Immune cell-mediated protection against vaginal candidiasis: Evidence for a major role of vaginal CD4+ T cells and possible participation of other local lymphocyte effectors. Infect. Immun. 2002, 70, 4791–4797. [Google Scholar] [CrossRef] [PubMed]
- De Bernardis, F.; Santoni, G.; Boccanera, M.; Lucciarini, R.; Arancia, S.; Sandini, S.; Amantini, C.; Cassone, A. Protection against rat vaginal candidiasis by adoptive transfer of vaginal B lymphocytes. FEMS Yeast Res. 2010, 10, 432–440. [Google Scholar] [CrossRef] [PubMed]
- Van De Veerdonk, F.L.; Lauwerys, B.; Marijnissen, R.J.; Timmermans, K.; Di Padova, F.; Koenders, M.I.; Gutierrez-Roelens, I.; Durez, P.; Netea, M.G.; Van Der Meer, J.W.M.; et al. The anti-CD20 antibody rituximab reduces the Th17 cell response. Arthritis Rheum. 2011, 63, 1507–1516. [Google Scholar] [CrossRef]
- Lilly, E.A.; Ikeh, M.; Nash, E.E.; Fidel, P.L., Jr.; Noverr, M.C. Immune Protection against Lethal Fungal-Bacterial Intra-Abdominal Infections. mBio 2018, 9, 1–14. [Google Scholar] [CrossRef]
- Rapaka, R.R.; Ricks, D.M.; Alcorn, J.F.; Chen, K.; Khader, S.A.; Zheng, M.; Plevy, S.; Bengtén, E.; Kolls, J.K. Conserved natural IgM antibodies mediate innate and adaptive immunity against the opportunistic fungus Pneumocystis murina. J. Exp. Med. 2010, 207, 2907–2919. [Google Scholar] [CrossRef]
- Li, W.; Fu, M.; An, J.G.; Xing, Y.; Zhang, P.; Zhang, X.; Wang, Y.C.; Li, C.X.; Tian, R.; Su, W.J.; et al. Host defence against C. albicans infections in IgH transgenic mice with VH derived from a natural anti-keratin antibody. Cell. Microbiol. 2007, 9, 306–315. [Google Scholar] [CrossRef] [PubMed]
- Tian, R.; Fu, M.; Zhang, Z.; Ren, J.; An, J.; Liu, Y.; Li, W. In Situ IgM Production and Clonal Expansion of B-1 Cells in Peritoneal Cavity Promote Elimination of C. albicans Infection in IgH Transgenic Mice with VH Derived from a Natural Antibody. PLoS ONE 2013, 8, e60779. [Google Scholar] [CrossRef]
- Rudkin, F.M.; Raziunaite, I.; Workman, H.; Essono, S.; Belmonte, R.; MacCallum, D.M.; Johnson, E.M.; Silva, L.M.; Palma, A.S.; Feizi, T.; et al. Single human B cell-derived monoclonal anti-Candida antibodies enhance phagocytosis and protect against disseminated candidiasis. Nat. Commun. 2018, 9, 5288. [Google Scholar] [CrossRef] [PubMed]
- Li, R.; Rezk, A.; Li, H.; Gommerman, J.L.; Prat, A.; Bar-Or, A. Antibody-Independent Function of Human B Cells Contributes to Antifungal T Cell Responses. J. Immunol. 2017, 198, 3245–3254. [Google Scholar] [CrossRef]
- Casadevall, A.; Feldmesser, M.; Pirofski, L.A. Induced humoral immunity and vaccination against major human fungal pathogens. Curr. Opin. Microbiol. 2002, 5, 386–391. [Google Scholar] [CrossRef]
- Vudhichamnong, K.; Walker, D.M.; Ryley, H.C. The effect of secretory immunoglobulin A on the in-vitro adherence of the yeast Candida albicans to human oral epithelial cells. Arch. Oral. Biol. 1982, 27, 617–621. [Google Scholar] [CrossRef]
- Maiti, P.K.; Kumar, A.; Kumar, R.; Mohapatra, L.N. Role of antibodies and effect of BCG vaccination in experimental candidiasis in mice. Mycopathologia 1985, 91, 79–85. [Google Scholar] [CrossRef]
- Cassone, A.; Bernardis, F.; Torososantucci, A. An Outline of the Role of Anti-Candida Antibodies Within the Context of Passive Immunization and Protection from Candidiasis. Curr. Mol. Med. 2005, 5, 377–382. [Google Scholar] [CrossRef]
- Cassone, A.; De Bernardis, F.; Santoni, G. Anticandidal immunity and vaginitis: Novel opportunities for immune intervention. Infect. Immun. 2007, 75, 4675–4686. [Google Scholar] [CrossRef][Green Version]
- Bohler, K.; Klade, H.; Poitschek, C.; Reinthaller, A. Immunohistochemical study of in vivo and in vitro IgA coating of candida species in vulvovaginal candidiasis. Genitourin. Med. 1994, 70, 182–186. [Google Scholar] [CrossRef] [PubMed]
- De Carvalho, R.J.; Cunha, C.M.; Silva, D.A.; Sopelete, M.C.; Urzedo, J.E.; Moreira, T.A.; Moraes Pde, S.; Taketomi, E.A. IgA, IgE and IgG subclasses to Candida albicans in serum and vaginal fluid from patients with vulvovaginal candidiasis. Rev. Assoc. Med. Bras. 2003, 49, 434–438. [Google Scholar] [CrossRef] [PubMed][Green Version]
- De Bernardis, F.; Liu, H.; O’Mahony, R.; La Valle, R.; Bartollino, S.; Sandini, S.; Grant, S.; Brewis, N.; Tomlinson, I.; Basset, R.C.; et al. Human domain antibodies against virulence traits of Candida albicans inhibit fungus adherence to vaginal epithelium and protect against experimental vaginal candidiasis. J. Infect. Dis. 2007, 195, 149–157. [Google Scholar] [CrossRef]
- Han, Y.; Cutler, J.E. Antibody response that protects against disseminated candidiasis. Infect. Immun. 1995, 63, 2714–2719. [Google Scholar] [CrossRef]
- Lopez-Ribot Jose, L.; Casanova, M.; Murgui, A.; Martinez, J. Antibody response to Candida albicans cell wall antigens. FEMS Immunol. Med. Microbiol. 2004, 41, 187–196. [Google Scholar] [CrossRef]
- Carneiro, C.; Correia, A.; Lima, T.; Vilanova, M.; Pais, C.; Gomes, A.C.; Real Oliveira, M.E.C.D.; Sampaio, P. Protective effect of antigen delivery using monoolein-based liposomes in experimental hematogenously disseminated candidiasis. Acta Biomater. 2016, 39, 133–145. [Google Scholar] [CrossRef]
- Xin, H.; Cutler, J.E. Vaccine and monoclonal antibody that enhance mouse resistance to candidiasis. Clin. Vaccine Immunol. 2011, 18, 1656–1667. [Google Scholar] [CrossRef] [PubMed]
- Wan Tso, G.H.; Reales-Calderon, J.A.; Pavelka, N. The Elusive Anti-Candida Vaccine: Lessons from the past and opportunities for the future. Front. Immunol. 2018, 9, 897. [Google Scholar] [CrossRef]
- Wang, X.J.; Sui, X.; Yan, L.; Wang, Y.; Cao, Y.B.; Jiang, Y.Y. Vaccines in the treatment of invasive candidiasis. Virulence 2015, 6, 309–315. [Google Scholar] [CrossRef] [PubMed]
- Ken-Ichi, I.; Masaharu, Y.; Iwao, N.; Hiroyasu, S.; Noriko, N.M.; Yoshiyuki Adachi, N.O. Role of anti-beta-glucan antibody in host defense against fungi. FEMS Immunol. Med. Microbiol. 2005, 44, 99–109. [Google Scholar]
- Mikulska, M.; Calandra, T.; Sanguinetti, M.; Poulain, D.; Viscoli, C. The use of mannan antigen and anti-mannan antibodies in the diagnosis of invasive candidiasis: Recommendations from the Third European Conference on Infections in Leukemia. Crit. Care 2010, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Kanbe, T.; Cherniak, R.; Cutler, J.E. Biochemical characterization of Candida albicans epitopes that can elicit protective and nonprotective antibodies. Infect. Immun. 1997, 65, 4100–4107. [Google Scholar] [CrossRef]
- Caesar-TonThat, T.C.; Cutler, J.E. A monoclonal antibody to Candida albicans enhances mouse neutrophil candidacidal activity. Infect. Immun. 1997, 65, 5354–5357. [Google Scholar] [CrossRef]
- Cassone, A.; Boccanera, M.; Adriani, D.; Santoni, G.; De Bernardis, F. Rats clearing a vaginal infection by Candida albicans acquire specific, antibody-mediated resistance to vaginal reinfection. Infect. Immun. 1995, 63, 2619–2624. [Google Scholar] [CrossRef]
- De Bernardis, F.; Boccanera, M.; Adriani, D.; Spreghini, E.; Santoni, G.; Cassone, A. Protective role of antimannan and anti-aspartyl proteinase antibodies in an experimental model of Candida albicans vaginitis in rats. Infect. Immun. 1997, 65, 3399–3405. [Google Scholar] [CrossRef] [PubMed]
- Mangeney, M.; Fischer, A.; Le Deist, F.; Latgé, J.P.; Durandy, A. Direct activation of human B lymphocytes by Candida albicans-derived mannan antigen. Cell. Immunol. 1989, 122, 329–337. [Google Scholar] [CrossRef]
- Han, Y.; Morrison, R.P.; Cutler, J.E. A vaccine and monoclonal antibodies that enhance mouse resistance to Candida albicans vaginal infection. Infect. Immun. 1998, 66, 5771–5776. [Google Scholar] [CrossRef]
- Zhang, M.X.; Kozel, T.R. Mannan-Specific Immunoglobulin G Antibodies in Normal Human Serum Mediate Classical Pathway Initiation of C3 Binding to Candida albicans. Infect. Immun. 1997, 65, 3822–3827. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.X.; Cutler, J.E.; Han, Y.; Kozel, T.R. Contrasting roles of mannan-specific monoclonal immunoglobulin M antibodies in the activation of classical and alternative pathways by Candida albicans. Infect. Immun. 1998, 66, 6027–6029. [Google Scholar] [CrossRef] [PubMed]
- Jouault, T.; Delaunoy, C.; Sendid, B.; Ajana, F.; Poulain, D. Differential humoral response against α- and β-linked mannose residues associated with tissue invasion by Candida albicans. Clin. Diagn. Lab. Immunol. 1997, 4, 328–333. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Ulrich, M.A.; Cutler, J.E. Candida albicans mannan extract-protein conjugates induce a protective immune response against experimental candidiasis. J. Infect. Dis. 1999, 179, 1477–1484. [Google Scholar] [CrossRef]
- Han, Y.; Riesselman, M.H.; Cutler, J.E. Protection against candidiasis by an immunoglobulin G3 (IgG3) monoclonal antibody specific for the same mannotriose as an IgM protective antibody. Infect. Immun. 2000, 68, 1649–1654. [Google Scholar] [CrossRef]
- Han, Y.; Kozel, T.R.; Zhang, M.X.; MacGill, R.S.; Carroll, M.C.; Cutler, J.E. Complement Is Essential for Protection by an IgM and an IgG3 Monoclonal Antibody Against Experimental, Hematogenously Disseminated Candidiasis. J. Immunol. 2001, 167, 1550–1557. [Google Scholar] [CrossRef]
- Cutler, J. Defining Criteria for Anti-Mannan Antibodies to Protect Against Candidiasis. Curr. Mol. Med. 2005, 5, 383–392. [Google Scholar] [CrossRef]
- Bundle, D.R.; Nycholat, C.; Costello, C.; Rennie, R.; Lipinski, T. Design of a Candida albicans disaccharide conjugate vaccine by reverse engineering a protective monoclonal antibody. ACS Chem. Biol. 2012, 7, 1754–1763. [Google Scholar] [CrossRef]
- De Bernardis, F.; Santoni, G.; Boccanera, M.; Spreghini, E.; Adriani, D.; Morelli, L.; Cassone, A. Local anticandidal immune responses in a rat model of vaginal infection by and protection against Candida albicans. Infect. Immun. 2000, 68, 3297–3304. [Google Scholar] [CrossRef]
- Bystrický, S.; Paulovičová, E.; Machová, E. Candida albicans mannan-protein conjugate as vaccine candidate. Immunol. Lett. 2003, 85, 251–255. [Google Scholar] [CrossRef]
- Kozel, T.R.; MacGill, R.S.; Percival, A.; Zhou, Q. Biological Activities of Naturally Occurring Antibodies Reactive with Candida albicans Mannan. Infect. Immun. 2004, 72, 209–218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.X.; Bohlman, M.C.; Itatani, C.; Burton, D.R.; Parren, P.W.H.I.; St. Jeor, S.C.; Kozel, T.R. Human recombinant antimannan immunoglobulin G1 antibody confers resistance to hematogenously disseminated candidiasis in mice. Infect. Immun. 2006, 74, 362–369. [Google Scholar] [CrossRef] [PubMed]
- Boxx, G.M.; Nishiya, C.T.; Kozel, T.R.; Zhang, M.X. Characteristics of Fc-independent human antimannan antibody-mediated alternative pathway initiation of C3 deposition to Candida albicans. Mol. Immunol. 2009, 46, 473–480. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Nishiya, C.T.; Boxx, G.M.; Robison, K.; Itatani, C.; Kozel, T.R.; Zhang, M.X. Influence of IgG subclass on human antimannan antibody-mediated resistance to hematogenously disseminated candidiasis in mice. Infect. Immun. 2016, 84, 386–394. [Google Scholar] [CrossRef]
- Wu, X.; Lipinski, T.; Carrel, F.R.; Bailey, J.J.; Bundle, D.R. Synthesis and immunochemical studies on a Candida albicans cluster glycoconjugate vaccine. Org. Biomol. Chem. 2007, 5, 3477–3485. [Google Scholar] [CrossRef]
- Wu, X.; Lipinski, T.; Paszkiewicz, E.; Bundle, D.R. Synthesis and immunochemical characterization of 5-linked glycoconjugate vaccines against Candida albicans. Chem. A Eur. J. 2008, 14, 6474–6482. [Google Scholar] [CrossRef]
- Paulovičová, E.; Machová, E.; Tulinská, J.; Bystrický, S. Cell and antibody mediated immunity induced by vaccination with novel Candida dubliniensis mannan immunogenic conjugate. Int. Immunopharmacol. 2007, 7, 1325–1333. [Google Scholar] [CrossRef]
- Machová, E.; Bystrický, S. Growth inhibition of Candida albicans in the presence of antiserum elicited in rabbits by mannan-protein conjugate. Z. Naturforsch. C J. Biosci. 2008, 63, 909–912. [Google Scholar] [CrossRef]
- Xin, H.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Synthetic glycopeptide vaccines combining β-mannan and peptide epitopes induce protection against candidiasis. Proc. Natl. Acad. Sci. USA 2008, 105, 13526–13531. [Google Scholar] [CrossRef]
- Xin, H.; Cartmell, J.; Bailey, J.J.; Dziadek, S.; Bundle, D.R.; Cutler, J.E. Self-adjuvanting glycopeptide conjugate vaccine against disseminated candidiasis. PLoS ONE 2012, 7, e35106. [Google Scholar] [CrossRef] [PubMed]
- Paulovičová, L.; Bystrický, S.; Paulovičová, E.; Karelin, A.A.; Tsvetkov, Y.E.; Nifantiev, N.E. Model α-mannoside conjugates: Immunogenicity and induction of candidacidal activity. FEMS Immunol. Med. Microbiol. 2010, 58, 307–313. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, T.; Kitov, P.I.; Szpacenko, A.; Paszkiewicz, E.; Bundle, D.R. Synthesis and immunogenicity of a glycopolymer conjugate. Bioconjug. Chem. 2011, 22, 274–281. [Google Scholar] [CrossRef] [PubMed]
- Lipinski, T.; Wu, X.; Sadowska, J.; Kreiter, E.; Yasui, Y.; Cheriaparambil, S.; Rennie, R.; Bundle, D.R. A β-mannan trisaccharide conjugate vaccine aids clearance of Candida albicans in immunocompromised rabbits. Vaccine 2012, 30, 6263–6269. [Google Scholar] [CrossRef] [PubMed]
- Paulovičová, L.; Paulovičová, E.; Karelin, A.A.; Tsvetkov, Y.E.; Nifantiev, N.E.; Bystrický, S. Humoral and cell-mediated immunity following vaccination with synthetic Candida cell wall mannan derived heptamannoside-protein conjugate: Immunomodulatory properties of heptamannoside-BSA conjugate. Int. Immunopharmacol. 2012, 14, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Rhew, K.Y. Comparison of two Candida mannan vaccines: The role of complement in protection against disseminated candidiasis. Arch. Pharm. Res. 2012, 35, 2021–2027. [Google Scholar] [CrossRef]
- Paulovičová, L.; Paulovičová, E.; Karelin, A.A.; Tsvetkov, Y.E.; Nifantiev, N.E.; Bystrický, S. Effect of branched α-oligomannoside structures on induction of anti-candida humoral immune response. Scand. J. Immunol. 2013, 77, 431–441. [Google Scholar] [CrossRef]
- Paulovičová, L.; Paulovičová, E.; Karelin, A.A.; Tsvetkov, Y.E.; Nifantiev, N.E.; Bystrický, S. Immune cell response to Candida cell wall mannan derived branched α-oligomannoside conjugates in mice. J. Microbiol. Immunol. Infect. 2015, 48, 9–19. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Machová, E.; Korcová, J.; Čížová, A.; Bystrický, S. Inhibition of yeast growth by broadly cross-reactive antisera elicited by heterologous mannan-protein conjugate. J. Microbiol. Biotechnol. 2015, 25, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Liao, J.; Pan, B.; Liao, G.; Zhao, Q.; Gao, Y.; Chai, X.; Zhuo, X.; Wu, Q.; Jiao, B.; Pan, W.; et al. Synthesis and immunological studies of β-1,2-mannan-peptide conjugates as antifungal vaccines. Eur. J. Med. Chem. 2019, 173, 250–260. [Google Scholar] [CrossRef] [PubMed]
- Paulovičová, L.; Paulovičová, E.; Farkaš, P.; Cízová, A.; Bystrický, P.; Jančinová, V.; Turánek, J.; Pericolini, E.; Gabrielli, E.; Vecchiarelli, A.; et al. Bioimmunological activities of Candida glabrata cellular mannan. FEMS Yeast Res. 2019, 19, foz009. [Google Scholar] [CrossRef]
- Sendid, B.; Lecointe, K.; Collot, M.; Danzé, P.M.; Damiens, S.; Drucbert, A.S.; Fradin, C.; Vilcot, J.P.; Grenouillet, F.; Dubar, F.; et al. Dissection of the anti-Candida albicans mannan immune response using synthetic oligomannosides reveals unique properties of β-1,2 mannotriose protective epitopes. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Bromuro, C.; Torosantucci, A.; Chiani, P.; Conti, S.; Polonelli, L.; Cassone, A. Interplay between protective and inhibitory antibodies dictates the outcome of experimentally disseminated candidiasis in recipients of a Candida albicans vaccine. Infect. Immun. 2002, 70, 5462–5470. [Google Scholar] [CrossRef] [PubMed]
- Torosantucci, A.; Bromuro, C.; Chiani, P.; De Bernardis, F.; Berti, F.; Galli, C.; Norelli, F.; Bellucci, C.; Polonelli, L.; Costantino, P.; et al. A novel glyco-conjugate vaccine against fungal pathogens. J. Exp. Med. 2005, 202, 597–606. [Google Scholar] [CrossRef]
- Rachini, A.; Pietrella, D.; Lupo, P.; Torosantucci, A.; Chiani, P.; Bromuro, C.; Proietti, C.; Bistoni, F.; Cassone, A.; Vecchiarelli, A. An anti-β-glucan monoclonal antibody inhibits growth and capsule formation of Cryptococcus neoformans in vitro and exerts therapeutic, anticryptococcal activity in vivo. Infect. Immun. 2007, 75, 5085–5094. [Google Scholar] [CrossRef]
- Torosantucci, A.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Palma, A.S.; Liu, Y.; Minogna, G.; Maras, B.; Colone, M.; Stringaro, A.; et al. Protection by anti-β-glucan antibodies is associated with restricted β-1,3 glucan binding specificity and inhibition of fungal growth and adherence. PLoS ONE 2009, 4, e5392. [Google Scholar] [CrossRef]
- Cassone, A.; Bromuro, C.; Chiani, P.; Torosantucci, A. Hyr1 protein and β-glucan conjugates as anti-candida vaccines. J. Infect. Dis. 2010, 202, 1930. [Google Scholar] [CrossRef]
- Capodicasa, C.; Chiani, P.; Bromuro, C.; De Bernardis, F.; Catellani, M.; Palma, A.S.; Liu, Y.; Feizi, T.; Cassone, A.; Benvenuto, E.; et al. Plant production of anti-β-glucan antibodies for immunotherapy of fungal infections in humans. Plant Biotechnol. J. 2011, 9, 776–787. [Google Scholar] [CrossRef]
- Pietrella, D.; Rachini, A.; Torosantucci, A.; Chiani, P.; Brown, A.J.P.; Bistoni, F.; Costantino, P.; Mosci, P.; d’Enfert, C.; Rappuoli, R.; et al. A β-glucan-conjugate vaccine and anti-β-glucan antibodies are effective against murine vaginal candidiasis as assessed by a novel in vivo imaging technique. Vaccine 2010, 28, 1717–1725. [Google Scholar] [CrossRef]
- Bromuro, C.; Romano, M.; Chiani, P.; Berti, F.; Tontini, M.; Proietti, D.; Mori, E.; Torosantucci, A.; Costantino, P.; Rappuoli, R.; et al. Beta-glucan-CRM197 conjugates as candidates antifungal vaccines. Vaccine 2010, 28, 2615–2623. [Google Scholar] [CrossRef]
- Magliani, W.; Conti, S.; Giovati, L.; Maffei, D.L.; Polonelli, L. Anti-beta-glucan-like immunoprotective candidacidal antiidiotypic antibodies. Front. Biosci. 2008, 13, 6920–6937. [Google Scholar] [CrossRef]
- Ishibashi, K.I.; Dogasaki, C.; Motoi, M.; Miura, N.; Adachi, Y.; Ohno, N. Anti-fungal cell wall β-glucan antibody in animal sera. Jpn. J. Med. Mycol. 2010, 51, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Chiani, P.; Bromuro, C.; Cassone, A.; Torosantucci, A. Anti-β-glucan antibodies in healthy human subjects. Vaccine 2009, 27, 513–519. [Google Scholar] [CrossRef] [PubMed]
- Torosantucci, A.; Tumbarello, M.; Bromuro, C.; Chiani, P.; Posteraro, B.; Sanguinetti, M.; Cauda, R.; Cassone, A. Antibodies against a β-glucan-protein complex of Candida albicans and its potential as indicator of protective immunity in candidemic patients. Sci. Rep. 2017, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Zito, A.; Bromuro, C.; Mandili, G.; Chiani, P.; Horenstein, A.L.; Malavasi, F.; Cauda, R.; Cassone, A.; Torosantucci, A. A murine, bispecific monoclonal antibody simultaneously recognizing β-Glucan and MP65 determinants in Candida species. PLoS ONE 2016, 11, e0148714. [Google Scholar] [CrossRef] [PubMed]
- Polonelli, L.; Magliani, W.; Conti, S.; Bracci, L.; Lozzi, L.; Neri, P.; Adriani, D.; De Bernardis, F.; Cassone, A. Therapeutic Activity of an Engineered Synthetic Killer Antiidiotypic Antibody Fragment against Experimental Mucosal and Systemic Candidiasis. Infect. Immun. 2003, 71, 6205–6212. [Google Scholar] [CrossRef]
- Polonelli, L.; Beninati, C.; Teti, G.; Felici, F.; Ciociola, T.; Giovati, L.; Sperindè, M.; Lo Passo, C.; Pernice, I.; Domina, M.; et al. Yeast killer toxin-like candidacidal Ab6 antibodies elicited through the manipulation of the idiotypic cascade. PLoS ONE 2014, 9, e105727. [Google Scholar] [CrossRef] [PubMed]
- Selvakumar, D.; Karim, N.; Miyamoto, M.; Furuichi, Y.; Komiyama, T. Recombinant single-chain anti-idiotypic antibody: An effective fungal β-1,3-glucan synthase inhibitor. Biol. Pharm. Bull. 2006, 29, 1848–1853. [Google Scholar] [CrossRef] [PubMed]
- Kabir, M.E.; Karim, N.; Krishnaswamy, S.; Selvakumar, D.; Miyamoto, M.; Furuichi, Y.; Komiyama, T. Peptide derived from anti-idiotypic single-chain antibody is a potent antifungal agent compared to its parent fungicide HM-1 killer toxin peptide. Appl. Microbiol. Biotechnol. 2011, 92, 1151–1160. [Google Scholar] [CrossRef]
- Huang, H.; Ostroff, G.R.; Lee, C.K.; Specht, C.A.; Levitz, S.M. Robust stimulation of humoral and cellular immune responses following vaccination with antigen-loaded β-Glucan particles. mBio 2010, 1, e00164-e10. [Google Scholar] [CrossRef]
- Mowlds, P.; Coates, C.; Renwick, J.; Kavanagh, K. Dose-dependent cellular and humoral responses in Galleria mellonella larvae following β-glucan inoculation. Microbes Infect. 2010, 12, 146–153. [Google Scholar] [CrossRef]
- Sheehan, G.; Kavanagh, K. Analysis of the early cellular and humoral responses of galleria mellonella larvae to infection by Candida albicans. Virulence 2018, 9, 163–172. [Google Scholar] [CrossRef]
- Lipinski, T.; Fitieh, A.; St. Pierre, J.; Ostergaard, H.L.; Bundle, D.R.; Touret, N. Enhanced Immunogenicity of a Tricomponent Mannan Tetanus Toxoid Conjugate Vaccine Targeted to Dendritic Cells via Dectin-1 by Incorporating β-Glucan. J. Immunol. 2013, 190, 4116–4128. [Google Scholar] [CrossRef]
- Bundle, D.R.; Paszkiewicz, E.; Elsaidi, H.R.H.; Mandal, S.S.; Sarkar, S. A three component synthetic vaccine containing a β-mannan T-cell peptide epitope and a β-glucan dendritic cell ligand. Molecules 2018, 23, 1961. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhou, Z.; Burgula, S.; Liao, J.; Yuan, C.; Wu, Q.; Guo, Z. Synthesis and Immunological Studies of Linear Oligosaccharides of β-Glucan As Antigens for Antifungal Vaccine Development. Bioconjug. Chem. 2015, 26, 466–476. [Google Scholar] [CrossRef] [PubMed]
- Liao, G.; Zhou, Z.; Liao, J.; Zu, L.; Wu, Q.; Guo, Z. 6-O-Branched Oligo-β-glucan-Based Antifungal Glycoconjugate Vaccines. ACS Infect. Dis. 2016, 2, 123–131. [Google Scholar] [CrossRef]
- Paulovičová, E.; Paulovičová, L.; Pilišiová, R.; Jančinová, V.; Yashunsky, D.V.; Karelin, A.A.; Tsvetkov, Y.E.; Nifantiev, N.E. The evaluation of β-(1→3)-nonaglucoside as an anti-Candida albicans immune response inducer. Cell. Microbiol. 2016, 18, 1294–1307. [Google Scholar] [CrossRef] [PubMed]
- Matveev, A.L.; Krylov, V.B.; Khlusevich, Y.A.; Baykov, I.K.; Yashunsky, V.; Emelyanova, L.A.; Tsvetkov, Y.E.; Karelin, A.A.; Bardashova, A.V.; Wong, S.S.W.; et al. Novel mouse monoclonal antibodies specifically recognizing B-(1–3)-D-glucan antigen. PLoS ONE 2019, 14, e0215535. [Google Scholar] [CrossRef]
- Shen, H.; Yu, Y.; Chen, S.M.; Sun, J.J.; Fang, W.; Guo, S.Y.; Hou, W.T.; Qiu, X.R.; Zhang, Y.; Chen, Y.L.; et al. Dectin-1 Facilitates IL-18 Production for the Generation of Protective Antibodies Against Candida albicans. Front. Microbiol. 2020, 11, 1–16. [Google Scholar] [CrossRef]
- Kumar, H.; Kumagai, Y.; Tsuchida, T.; Koenig, P.A.; Satoh, T.; Guo, Z.; Jang, M.H.; Saitoh, T.; Akira, S.; Kawai, T. Involvement of the NLRP3 Inflammasome in Innate and Humoral Adaptive Immune Responses to Fungal β-Glucan. J. Immunol. 2009, 183, 8061–8067. [Google Scholar] [CrossRef]
- Seo, B.-S.; Lee, S.-H.; Lee, J.-E.; Yoo, Y.-C.; Lee, J.; Park, S.-R. Dectin-1 Stimulation Selectively Reinforces LPS-driven IgG1 Production by Mouse B Cells. Immune Netw. 2013, 13, 205. [Google Scholar] [CrossRef] [PubMed]
- Hoogeboom, R.; van Kesse, K.P.M.; Hochstenbach, F.; Wormhoudt, T.A.; Reinten, R.J.A.; Wagner, K.; Kater, A.P.; Guikema, J.E.J.; Bende, R.J.; van Noesel, C.J.M. A mutated B cell chronic lymphocytic leukemia subset that recognizes and responds to fungi. J. Exp. Med. 2013, 210, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Adamo, R.; Tontini, M.; Brogioni, G.; Romano, M.R.; Costantini, G.; Danieli, E.; Proietti, D.; Berti, F.; Costantino, P. Synthesis of laminarin fragments and evaluation of a β-(1,3) glucan hexasaccaride-CRM 197 conjugate as vaccine candidate against Candida albicans. J. Carbohydr. Chem. 2011, 30, 249–280. [Google Scholar] [CrossRef]
- Li, W.J.; Long, K.; Dong, H.L.; Gao, X.M. Adjuvanticity of a recombinant calreticulin fragment in assisting anti-β-glucan igG responses in T cell-deficient mice. Clin. Vaccine Immunol. 2013, 20, 582–589. [Google Scholar] [CrossRef] [PubMed]
- Matthews, R.; Burnie, J. The role of hsp90 in fungal infection. Immunol. Today 1992, 13, 345–348. [Google Scholar] [CrossRef]
- Matthews, R.C.; Burnie, J.P.; Howat, D.; Rowland, T.; Walton, F. Autoantibody to heat-shock protein 90 can mediate protection against systemic candidosis. Immunology 1991, 74, 20–24. [Google Scholar]
- Matthews, R.; Burnie, J.P.; Lee, W. The application of epitope mapping in the development of a new serological test for systemic candidosis. J. Immunol. Methods 1991, 143, 73–79. [Google Scholar] [CrossRef]
- Matthews, R.; Hodgetts, S.; Burnie, J. Preliminary Assessment of a Human Recombinant Antibody Fragment to hsp90 in Murine Invasive Candidiasis. J. Infect. Dis. 1995, 171, 1668–1671. [Google Scholar] [CrossRef]
- Matthews, R.C.; Rigg, G.; Hodgetts, S.; Carter, T.; Chapman, C.; Gregory, C.; Illidge, C.; Burnie, J. Preclinical assessment of the efficacy of mycograb, a human recombinant antibody against fungal HSP90. Antimicrob. Agents Chemother. 2003, 47, 2208–2216. [Google Scholar] [CrossRef]
- Pachl, J.; Svododa, P.; Jacobs, F.; Vandewoude, K.; Van Der Hoven, B.; Spronk, P.; Masterson, G.; Malbrain, M.; Aoun, M.; Garbino, J.; et al. A randomized, blinded, multicenter trial of lipid-associated amphotericin B alone versus in combination with an antibody-based inhibitor of heat shock protein 90 in patients with invasive candidiasis. Clin. Infect. Dis. 2006, 42, 1404–1413. [Google Scholar] [CrossRef]
- Karwa, R.; Wargo, K.A. Efungumab: A novel agent in the treatment of invasive candidiasis. Ann. Pharmacother. 2009, 43, 1818–1823. [Google Scholar] [CrossRef]
- Louie, A.; Stein, D.S.; Zack, J.Z.; Liu, W.; Conde, H.; Fregeau, C.; VanScoy, B.D.; Drusano, G.L. Dose range evaluation of mycograb C28Y variant, a human recombinant antibody fragment to heat shock protein 90, in combination with amphotericin B-desoxycholate for treatment of murine systemic candidiasis. Antimicrob. Agents Chemother. 2011, 55, 3295–3304. [Google Scholar] [CrossRef] [PubMed]
- Richie, D.L.; Ghannoum, M.A.; Isham, N.; Thompson, K.V.; Ryder, N.S. Nonspecific effect of mycograb on amphotericin B MIC. Antimicrob. Agents Chemother. 2012, 56, 3963–3964. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bugli, F.; Cacaci, M.; Martini, C.; Torelli, R.; Posteraro, B.; Sanguinetti, M.; Paroni Sterbini, F. Human monoclonal antibody-based therapy in the treatment of invasive candidiasis. Clin. Dev. Immunol. 2013, 2013, 403121. [Google Scholar] [CrossRef] [PubMed]
- Raška, M.; Běláková, J.; Wudattu, N.K.; Kafková, L.; Růžičková, K.; Šebestová, M.; Kolář, Z.; Weigl, E. Comparison of protective effect of protein and DNA vaccines hsp90 in murine model of systemic candidiasis. Folia Microbiol. 2005, 50, 77–82. [Google Scholar] [CrossRef]
- Yang, Q.; Wang, L.; Lu, D.N.; Gao, R.J.; Song, J.N.; Hua, P.Y.; Yuan, D.W. Prophylactic vaccination with phage-displayed epitope of C. albicans elicits protective immune responses against systemic candidiasis in C57BL/6 mice. Vaccine 2005, 23, 4088–4096. [Google Scholar] [CrossRef]
- Wang, G.; Sun, M.; Fang, J.; Yang, Q.; Tong, H.; Wang, L. Protective immune responses against systemic candidiasis mediated by phage-displayed specific epitope of Candida albicans heat shock protein 90 in C57BL/6J mice. Vaccine 2006, 24, 6065–6073. [Google Scholar] [CrossRef] [PubMed]
- Raska, M.; Belakova, J.; Horynova, M.; Krupka, M.; Novotny, J.; Sebestova, M.; Weigl, E. Systemic and mucosal immunization with Candida albicans hsp90 elicits hsp90-specific humoral response in vaginal mucosa which is further enhanced during experimental vaginal candidiasis. Med. Mycol. 2008, 46, 411–420. [Google Scholar] [CrossRef]
- Mašek, J.; Bartheldyová, E.; Turánek-Knotigová, P.; Škrabalová, M.; Korvasová, Z.; Plocková, J.; Koudelka, Š.; Škodová, P.; Kulich, P.; Křupka, M.; et al. Metallochelating liposomes with associated lipophilised norAbuMDP as biocompatible platform for construction of vaccines with recombinant His-tagged antigens: Preparation, structural study and immune response towards rHsp90. J. Control. Release 2011, 151, 193–201. [Google Scholar] [CrossRef]
- Knotigová, P.T.; Zyka, D.; Mašek, J.; Kovalová, A.; Křupka, M.; Bartheldyová, E.; Kulich, P.; Koudelka, Š.; Lukáč, R.; Kauerová, Z.; et al. Molecular adjuvants based on nonpyrogenic lipophilic derivatives of norAbuMDP/gmdp formulated in nanoliposomes: Stimulation of innate and adaptive immunity. Pharm. Res. 2015, 32, 1186–1199. [Google Scholar] [CrossRef]
- Yang, F.; Xiao, C.; Qu, J.; Wang, G. Structural characterization of low molecular weight polysaccharide from Astragalus membranaceus and its immunologic enhancement in recombinant protein vaccine against systemic candidiasis. Carbohydr. Polym. 2016, 145, 48–55. [Google Scholar] [CrossRef]
- Li, X.; Yang, Y.; Yang, F.; Wang, F.; Li, H.; Tian, H.; Wang, G. Chitosan hydrogel loaded with recombinant protein containing epitope C from HSP90 of Candida albicans induces protective immune responses against systemic candidiasis. Int. J. Biol. Macromol. 2021, 173, 327–340. [Google Scholar] [CrossRef]
- Hoyer, L.L.; Cota, E. Candida albicans Agglutinin-Like Sequence (Als) Family Vignettes: A Review of Als Protein Structure and Function. Front Microbiol. 2016, 7, 280. [Google Scholar] [CrossRef]
- Phan, Q.T.; Myers, C.L.; Fu, Y.; Sheppard, D.C.; Yeaman, M.R.; Welch, W.H.; Ibrahim, A.S.; Edwards, J.E.; Filler, S.G. Als3 is a Candida albicans invasin that binds to cadherins and induces endocytosis by host cells. PLoS Biol. 2007, 5, 0543–0557. [Google Scholar] [CrossRef]
- David, A. Coleman, a Soon-Hwan Oh, a Xiaomin Zhao, a Hongyuan Zhao, b Jeff, T. Hutchins, b John, H. Vernachio, b Joseph, M. Patti, b and L.L.H. Monoclonal antibodies specific for Candida albicans Als3 that immunolabel fungal cells in vitro and in vivo and block adhesion to host surfaces. J. Biotechnol. Methods 2009, 78, 71–78. [Google Scholar] [CrossRef]
- Cleary, I.A.; Reinhard, S.M.; Lindsay Miller, C.; Murdoch, C.; Thornhill, M.H.; Lazzell, A.L.; Monteagudo, C.; Thomas, D.P.; Saville, S.P. Candida albicans adhesin Als3p is dispensable for virulence in the mouse model of disseminated candidiasis. Microbiology 2011, 157, 1806–1815. [Google Scholar] [CrossRef] [PubMed]
- Murciano, C.; Moyes, D.L.; Runglall, M.; Tobouti, P.; Islam, A.; Hoyer, L.L.; Naglik, J.R. Evaluation of the role of Candida albicans agglutinin-like sequence (ALS) proteins in human oral epithelial cell interactions. PLoS ONE 2012, 7, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Liang, G.; Wang, Q.; She, X.; Shi, D.; Shen, Y.; Su, X.; Wang, X.; Wang, W.; Li, D.; et al. Different Host Immunological Response to C. albicans by Human Oral and Vaginal Epithelial Cells. Mycopathologia 2019, 184, 1–12. [Google Scholar] [CrossRef]
- Spellberg, B.; Ibrahim, A.S.; Edwards, J.E.; Filler, S.G. Mice with disseminated candidiasis die of progressive sepsis. J. Infect. Dis. 2005, 192, 336–343. [Google Scholar] [CrossRef]
- Ibrahim, A.S.; Spellberg, B.J.; Avenissian, V.; Fu, Y.; Filler, S.G.; Edwards, J.E.; Edwards, J.E., Jr. Vaccination with recombinant N-terminal domain of Als1p improves survival during murine disseminated candidiasis by enhancing cell-mediated, not humoral, immunity. Infect. Immun. 2005, 73, 999–1005. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.J.; Ibrahim, A.S.; Avenissian, V.; Filler, S.G.; Myers, C.L.; Fu, Y.; Edwards, J.E. The anti-Candida albicans vaccine composed of the recombinant N terminus of Als1p reduces fungal burden and improves survival in both immunocompetent and immunocompromised mice. Infect. Immun. 2005, 73, 6191–6193. [Google Scholar] [CrossRef]
- Spellberg, B.J.; Ibrahim, A.S.; Avanesian, V.; Fu, Y.; Myers, C.; Phan, Q.T.; Filler, S.G.; Yeaman, M.R.; Edwards, J.E. Efficacy of the anti-Candida rAls3p-N or rAls1p-N vaccines against disseminated and mucosal candidiasis. J. Infect. Dis. 2006, 194, 256–260. [Google Scholar] [CrossRef]
- Lin, L.; Ibrahim, A.S.; Avanesian, V.; Edwards, J.E.; Fu, Y.; Baquir, B.; Taub, R.; Spellberg, B. Considerable differences in vaccine immunogenicities and efficacies related to the diluent used for aluminum hydroxide adjuvant. Clin. Vaccine Immunol. 2008, 15, 582–584. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, B.; Ibrahim, A.S.; Yeaman, M.R.; Lin, L.; Fu, Y.; Avanesian, V.; Bayer, A.S.; Filler, S.G.; Lipke, P.; Otoo, H.; et al. The antifungal vaccine derived from the recombinant N terminus of Als3p protects mice against the bacterium Staphylococcus aureus. Infect. Immun. 2008, 76, 4574–4580. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Ibrahim, A.S.; Baquir, B.; Avanesian, V.; Fu, Y.; Spellberg, B. Immunological surrogate marker of rAls3p-N vaccine-induced protection against Staphylococcus aureus. FEMS Immunol. Med. Microbiol. 2009, 55, 293–295. [Google Scholar] [CrossRef]
- Spellberg, B.; Ibrahim, A.S.; Lin, L.; Avanesian, V.; Fu, Y.; Lipke, P.; Otoo, H.; Ho, T.; Edwards, J.E. Antibody titer threshold predicts anti-candidal vaccine efficacy even though the mechanism of protection is induction of cell-mediated immunity. J. Infect. Dis. 2008, 197, 967–971. [Google Scholar] [CrossRef]
- Baquir, B.; Lin, L.; Ibrahim, A.S.; Fu, Y.; Avanesian, V.; Tu, A.; Edwards, J., Jr.; Spellberg, B. Immunological Reactivity of Blood from Healthy Humans to the rAls3p-N Vaccine Protein. J. Infect. Dis. 2010, 201, 473–477. [Google Scholar] [CrossRef]
- Brena, S.; Omaetxebarría, M.J.; Elguezabal, N.; Cabezas, J.; Moragues, M.D.; Pontón, J. Fungicidal monoclonal antibody C7 binds to Candida albicans Als3. Infect. Immun. 2007, 75, 3680–3682. [Google Scholar] [CrossRef]
- Beucher, B.; Marot-Leblond, A.; Billaud-Nail, S.; Oh, S.H.; Hoyer, L.L.; Robert, R. Recognition of Candida albicans Als3 by the germ tube-specific monoclonal antibody 3D9.3. FEMS Immunol. Med. Microbiol. 2009, 55, 314–323. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.S.; White, C.J.; Ibrahim, A.S.; Filler, S.G.; Fu, Y.; Yeaman, M.R.; Edwards, J.E.; Hennessey, J.P. NDV-3, a recombinant alum-adjuvanted vaccine for Candida and Staphylococcus aureus, is safe and immunogenic in healthy adults. Vaccine 2012, 30, 7594–7600. [Google Scholar] [CrossRef] [PubMed]
- Ibrahim, A.S.; Luo, G.; Gebremariam, T.; Lee, H.; Schmidt, C.S.; Hennessey, J.P.; French, S.W.; Yeaman, M.R.; Filler, S.G.; Edwards, J.E. NDV-3 protects mice from vulvovaginal candidiasis through T- and B-cell immune response. Vaccine 2013, 31, 5549–5556. [Google Scholar] [CrossRef] [PubMed]
- Yeaman, M.R.; Filler, S.G.; Chaili, S.; Barr, K.; Wang, H.; Kupferwasser, D.; Hennessey, J.P.; Fu, Y.; Schmidt, C.S.; Edwards, J.E.; et al. Mechanisms of NDV-3 vaccine efficacy in MRSA skin versus invasive infection. Proc. Natl. Acad. Sci. USA 2014, 111, E5555–E5563. [Google Scholar] [CrossRef]
- Edwards, J.E.; Schwartz, M.M.; Schmidt, C.S.; Sobel, J.D.; Nyirjesy, P.; Schodel, F.; Marchus, E.; Lizakowski, M.; Demontigny, E.A.; Hoeg, J.; et al. A Fungal Immunotherapeutic Vaccine (NDV-3A) for Treatment of Recurrent Vulvovaginal Candidiasis-A Phase 2 Randomized, Double-Blind, Placebo-Controlled Trial. Clin. Infect. Dis. 2018, 66, 1928–1936. [Google Scholar] [CrossRef]
- Alqarihi, A.; Singh, S.; Edwards, J.E.; Ibrahim, A.S.; Uppuluri, P. NDV-3A vaccination prevents C. albicans colonization of jugular vein catheters in mice. Sci. Rep. 2019, 9, 1–6. [Google Scholar] [CrossRef]
- Singh, S.; Uppuluri, P.; Alqarihi, A.; Elhassan, H.; French, S.; Lockhart, S.R.; Chiller, T.; Edwards, J.E.; Ibrahim, A.S. The NDV-3A vaccine protects mice from multidrug resistant Candida auris infection. PLoS Pathog. 2019, 15, e1007460. [Google Scholar] [CrossRef]
- Naglik, J.R.; Challacombe, S.J.; Hube, B. Candida albicans Secreted Aspartyl Proteinases in Virulence and Pathogenesis. Microbiol. Mol. Biol. Rev. 2003, 67, 400–428. [Google Scholar] [CrossRef]
- Hube, B.; Sanglard, D.; Odds, F.C.; Hess, D.; Brown, A.J.P.; Gow, N.A.R.; Monod, M.; Scha, W. Disruption of Each of the Secreted Aspartyl Proteinase Genes Attenuates Virulence. Infect. Immun. 1997, 65, 3529–3538. [Google Scholar] [CrossRef] [PubMed]
- Sanglard, D.; Hube, B.; Monod, M.; Odds, F.C.; Gow, N.A.R. A triple deletion of the secreted aspartyl proteinase genes SAP4, SAP5, and SAP6 of Candida albicans causes attenuated virulence. Infect. Immun. 1997, 65, 3539–3546. [Google Scholar] [CrossRef] [PubMed]
- De Bernardis, F.; Arancia, S.; Morelli, L.; Hube, B.; Sanglard, D.; Schafer, W. Evidence that Members of the Secretory Aspartyl Proteinase Gene Family, in Particular SAP2, Are Virulence Factors for Candida Vaginitis. J. Infect. Dis. 1999, 179, 201–208. [Google Scholar] [CrossRef]
- Borg, M.; Ruchel, R. Expression of Extracellular Acid Proteinase by Proteolytic Candida spp. during Experimental Infection of Oral Mucosa. Infect. Immun. 1988, 56, 626–631. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, F.; Odds, F.C. Inducible proteinase of Candida albicans in diagnostic serology and in the pathogenesis of systemic candidosis. J. Med. Microbiol. 1980, 13, 423–435. [Google Scholar] [CrossRef]
- Smolenski, G.; Sullivan, P.A.; Cutfield, S.M.; Cutfield, J.F. Analysis of secreted aspartic proteinases from Candida albicans: Purification and characterization of individual Sapl, Sap2 and Sap3 isoenzymes. Microbiology 1996, 143, 349–356. [Google Scholar] [CrossRef] [PubMed][Green Version]
- White, T.C.; Miyasaki, S.H.; Agabian, N. Three distinct secreted aspartyl proteinases in Candida albicans. J. Bacteriol. 1993, 175, 6126–6133. [Google Scholar] [CrossRef] [PubMed][Green Version]
- White, T.C.; Agabian, N. Candida albicans secreted aspartyl proteinases: Isoenzyme pattern is determined by cell type, and levels are determined by environmental factors. J. Bacteriol. 1995, 177, 5215–5221. [Google Scholar] [CrossRef]
- Drobacheff, C.; Millon, L.; Monod, M.; Piarroux, R.; Robinet, E.; Laurent, R.; Meillet, D. Increased Serum and Salivary Immunoglobulins against Candida albicans in HIV-Infected Patients with Oral Candidiasis. Clin. Chem. Lab. Med. 2001, 39, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Millon, L.; Drobacheff, C.; Piarroux, R.; Monod, M.; Reboux, G.; Laurent, R.; Meillet, D. Longitudinal study of anti-Candida albicans mucosal immunity against aspartic proteinases in HIV-infected patients. J. Acquir. Immune Defic. Syndr. 2001, 26, 137–144. [Google Scholar] [CrossRef]
- Ghadjari, A.; Matthews, R.C.; Burnie, J.P. Epitope Mapping Candida Albicans Proteinase (SAP 2). FEMS Immunol. Med. Microbiol. 1997, 19, 115–123. [Google Scholar]
- De Bernardis, F.; Boccanera, M.; Adriani, D.; Girolamo, A.; Cassone, A. Intravaginal and intranasal immunizations are equally effective in inducing vaginal antibodies and conferring protection against vaginal candidiasis. Infect. Immun. 2002, 70, 2725–2729. [Google Scholar] [CrossRef]
- Suenobu, N.; Kweon, M.N.; Kiyono, H. Nasal vaccination induces the ability to eliminate Candida colonization without influencing the pre-existing antigen-specific IgE Abs: A possibility for the control of Candida-related atopic dermatitis. Vaccine 2002, 20, 2972–2980. [Google Scholar] [CrossRef]
- Vilanova, M.; Teixeira, L.; Caramalho, Í.; Torrado, E.; Marques, A.; Madureira, P.; Ribeiro, A.; Ferreira, P.; Gama, M.; Demengeot, J. Protection against systemic candidiasis in mice immunized with secreted aspartic proteinase 2. Immunology 2004, 111, 334–342. [Google Scholar] [CrossRef] [PubMed]
- Rahman, D.; Mistry, M.; Thavaraj, S.; Challacombe, S.J.; Naglik, J.R. Murine model of concurrent oral and vaginal Candida albicans colonization to study epithelial host-pathogen interactions. Microbes Infect. 2007, 9, 615–622. [Google Scholar] [CrossRef] [PubMed]
- Staib, P.; Lermann, U.; Blaß-Warmuth, J.; Degel, B.; Würzner, R.; Monod, M.; Schirmeister, T.; Morschhäuser, J. Tetracycline-inducible expression of individual secreted aspartic proteases in Candida albicans allows isoenzyme-specific inhibitor screening. Antimicrob. Agents Chemother. 2008, 52, 146–156. [Google Scholar] [CrossRef]
- Sandini, S.; La Valle, R.; Deaglio, S.; Malavasi, F.; Cassone, A.; De Bernardis, F. A highly immunogenic recombinant and truncated protein of the secreted aspartic proteases family (rSap2t) of Candida albicans as a mucosal anticandidal vaccine. FEMS Immunol. Med. Microbiol. 2011, 62, 215–224. [Google Scholar] [CrossRef] [PubMed]
- De Bernardis, F.; Amacker, M.; Arancia, S.; Sandini, S.; Gremion, C.; Zurbriggen, R.; Moser, C.; Cassone, A. A virosomal vaccine against candidal vaginitis: Immunogenicity, efficacy and safety profile in animal models. Vaccine 2012, 30, 4490–4498. [Google Scholar] [CrossRef]
- De Bernardis, F.; Arancia, S.; Sandini, S.; Graziani, S.; Norelli, S. Studies of immune responses in Candida vaginitis. Pathogens 2015, 4, 697–707. [Google Scholar] [CrossRef]
- Wang, Y.; Su, Q.; Dong, S.; Shi, H.; Gao, X.; Wang, L. Hybrid phage displaying SLAQVKYTSASSI induces protection against Candida albicans challenge in BALB/c mice. Hum. Vaccines Immunother. 2014, 10, 1057–1063. [Google Scholar] [CrossRef]
- Xu, M.; Liu, J. Immunoprotection by Sap2 and dendritic cells against systemic Candida albicans infection in mice. J. South. Med. Univ. 2014, 34, 1030–1034. [Google Scholar]
- Pericolini, E.; Gabrielli, E.; Amacker, M.; Kasper, L.; Roselletti, E.; Luciano, E.; Sabbatini, S.; Kaeser, M.; Moser, C.; Hube, B.; et al. Secretory aspartyl proteinases cause vaginitis and can mediate vaginitis caused by Candida albicans in mice. mBio 2015, 6, e00724-15. [Google Scholar] [CrossRef]
- Huai, Y.; Dong, S.; Zhu, Y.; Li, X.; Cao, B.; Gao, X.; Yang, M.; Wang, L.; Mao, C. Genetically Engineered Virus Nanofibers as an Efficient Vaccine for Preventing Fungal Infection. Adv. Healthc. Mater. 2016, 5, 786–794. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Shi, H.; Cao, D.; Wang, Y.; Zhang, X.; Li, Y.; Gao, X.; Wang, L. Novel nanoscale bacteriophage-based single-domain antibodies for the therapy of systemic infection caused by Candida albicans. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Bailey, D.A.; Feldmann, P.J.F.; Bovey, M.; Gow, N.A.R.; Brown, A.J.P. The Candida albicans HYR1 gene, which is activated in response to hyphal development, belongs to a gene family encoding yeast cell wall proteins. J. Bacteriol. 1996, 178, 5353–5360. [Google Scholar] [CrossRef] [PubMed]
- Luo, G.; Ibrahim, A.S.; Spellberg, B.; Nobile, C.J.; Mitchell, A.P.; Fu, Y. Candida albicans Hyr1p Confers Resistance to Neutrophil Killing and Is a Potential Vaccine Target. J. Infect. Dis. 2010, 201, 1718–1728. [Google Scholar] [CrossRef]
- Luo, G.; Ibrahim, A.S.; French, S.W.; Edwards, J.E.; Fu, Y. Active and passive immunization with rHyr1p-N protects mice against hematogenously disseminated candidiasis. PLoS ONE 2011, 6, e25909. [Google Scholar] [CrossRef]
- Uppuluri, P.; Lin, L.; Alqarihi, A.; Luo, G.; Youssef, E.G.; Alkhazraji, S.; Yount, N.Y.; Ibrahim, B.A.; Bolaris, M.A.; Edwards, J.E.; et al. The Hyr1 protein from the fungus Candida albicans is a cross kingdom immunotherapeutic target for Acinetobacter bacterial infection. PLoS Pathog. 2018, 14, e1007056. [Google Scholar] [CrossRef]
- Youssef, E.G.; Zhang, L.; Alkhazraji, S.; Gebremariam, T.; Singh, S.; Yount, N.Y.; Yeaman, M.R.; Uppuluri, P.; Ibrahim, A.S. Monoclonal IgM Antibodies Targeting Candida albicans Hyr1 Provide Cross-Kingdom Protection Against Gram-Negative Bacteria. Front. Immunol. 2020, 11, 6–8. [Google Scholar] [CrossRef]
- Tsuchimori, N.; Sharkey, L.L.; Fonzi, W.A.; French, S.W.; Edwards, J.E.; Filler, S.G. Reduced virulence of HWP1-deficient mutants of Candida albicans and their interactions with host cells. Infect. Immun. 2000, 68, 1997–2002. [Google Scholar] [CrossRef]
- Nobile, C.J.; Andes, D.R.; Nett, J.E.; Smith, F.J.; Yue, F.; Phan, Q.T.; Edwards, J.E.; Filler, S.G.; Mitchell, A.P. Critical role of Bcr1-dependent adhesins in C. albicans biofilm formation in vitro and in vivo. PLoS Pathog. 2006, 2, e63. [Google Scholar] [CrossRef]
- Dwivedi, P.; Thompson, A.; Xie, Z.; Kashleva, H.; Ganguly, S.; Mitchell, A.P.; Dongari-Bagtzoglou, A. Role of Bcr1-activated genes Hwp1 and Hyr1 in Candida albicans oral mucosal biofilms and neutrophil evasion. PLoS ONE 2011, 6, e16218. [Google Scholar] [CrossRef]
- Naglik, J.R.; Fostira, F.; Ruprai, J.; Staab, J.F.; Challacombe, S.J.; Sundstrom, P. Candida albicans HWP1 gene expression and host antibody responses in colonization and disease. J. Med. Microbiol. 2006, 55, 1323–1327. [Google Scholar] [CrossRef] [PubMed]
- Corouge, M.; Loridant, S.; Fradin, C.; Salleron, J.; Damiens, S.; Moragues, M.D.; Souplet, V.; Jouault, T.; Robert, R.; Dubucquoi, S.; et al. Humoral immunity links Candida albicans infection and celiac disease. PLoS ONE 2015, 10, e0121776. [Google Scholar] [CrossRef] [PubMed]
- Rosario-colon, J.; Eberle, K.; Adams, A.; Courville, E.; Xin, H. Candida cell-surface-specific monoclonal antibodies protect mice against candida auris invasive infection. Int. J. Mol. Sci. 2021, 22, 6162. [Google Scholar] [CrossRef] [PubMed]
- Silva, R.C.; Padovan, A.C.B.; Pimenta, D.C.; Ferreira, R.C.; da Silva, C.V.; Briones, M.R.S. Extracellular enolase of Candida albicans is involved in colonization of mammalian intestinal epithelium. Front. Cell. Infect. Microbiol. 2014, 4, 1–9. [Google Scholar] [CrossRef]
- Satala, D.; Karkowska-Kuleta, J.; Zelazna, A.; Rapala-Kozik, M.; Kozik, A. Moonlighting proteins at the candidal cell surface. Microorganisms 2020, 8, 1046. [Google Scholar] [CrossRef]
- Sundstrom, P.; Jensen, J.; Balish, E. Humoral and cellular immune responses to enolase after alimentary tract colonization or intravenous immunization with candida albicans. J. Infect. Dis. 1994, 170, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Van Deventer, H.J.; Goessens, W.H.; Van Vliet, A.J.; Verbrugh, H.A. Anti-enolase antibodies partially protective against systemic candidiasis in mice. Clin. Microbiol. Infect. 1996, 2, 36–43. [Google Scholar] [CrossRef]
- Montagnoli, C.; Sandini, S.; Bacci, A.; Romani, L.; La Valle, R. Immunogenicity and protective effect of recombinant enolase of Candida albicans in a murine model of systemic candidiasis. Med. Mycol. 2004, 42, 319–324. [Google Scholar] [CrossRef] [PubMed]
- Li, W.Q.; Hu, X.C.; Zhang, X.; Ge, Y.; Zhao, S.; Hu, Y.; Ashman, R.B. Immunisation with the glycolytic enzyme enolase confers effective protection against Candida albicans infection in mice. Vaccine 2011, 29, 5526–5533. [Google Scholar] [CrossRef]
- Laín, A.; Moragues, M.D.; García Ruiz, J.C.; Mendoza, J.; Camacho, A.; Del Palacio, A.; Pontón, J. Evaluation of a novel enzyme-linked immunosorbent assay to detect immunoglobulin G antibody to enolase for serodiagnosis of invasive candidiasis. Clin. Vaccine Immunol. 2007, 14, 318–319. [Google Scholar] [CrossRef]
- Li, F.Q.; Ma, C.F.; Shi, L.N.; Lu, J.F.; Wang, Y.; Huang, M.; Kong, Q. Diagnostic value of immunoglobulin G antibodies against Candida enolase and fructose-bisphosphate aldolase for candidemia. BMC Infect. Dis. 2013, 13, 1. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Aoki, W.; Nomura, T.; Miyoshi, A.; Tafuku, S.; Sewaki, T.U.M. An oral vaccine against candidiasis generated by a yeast molecular display system. Pathog. Dis. 2013, 69, 262–268. [Google Scholar] [CrossRef] [PubMed]
- He, Z.X.; Chen, J.; Li, W.; Cheng, Y.; Zhang, H.P.; Zhang, L.N.; Hou, T.W. Serological response and diagnostic value of recombinant candida cell wall protein enolase, phosphoglycerate kinase, and β-glucosidase. Front. Microbiol. 2015, 6, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Leu, S.J.; Lee, Y.C.; Lee, C.H.; Liao, P.Y.; Chiang, C.W.; Yang, C.M.; Su, C.H.; Ou, T.Y.; Liu, K.J.; Lo, H.J.; et al. Generation and Characterization of Single Chain Variable Fragment against Alpha-Enolase of Candida albicans. Int. J. Mol. Sci. 2020, 21, 2903. [Google Scholar] [CrossRef] [PubMed]
- Djordjevic, J.T. Role of phospholipases in fungal fitness, pathogenicity, and drug development-lessons from Cryptococcus neoformans. Front. Microbiol. 2010, 1, 25. [Google Scholar] [CrossRef]
- Theiss, S.; Ishdorj, G.; Brenot, A.; Kretschmar, M.; Lan, C.Y.; Nichterlein, T.; Hacker, J.; Nigam, S.; Agabian, N.; Köhler, G.A. Inactivation of the phospholipase B gene PLB5 in wild-type Candida albicans reduces cell-associated phospholipase A2 activity and attenuates virulence. Int. J. Med. Microbiol. 2006, 296, 405–420. [Google Scholar] [CrossRef] [PubMed]
- Ghannoum, M.A. Potential role of phospholipases in virulence and fungal pathogenesis. Clin. Microbiol. Rev. 2000, 13, 122–143. [Google Scholar] [CrossRef]
- Leidich, S.D.; Ibrahim, A.S.; Fu, Y.; Koul, A.; Jessup, C.; Vitullo, J.; Fonzi, W.; Mirbod, F.; Nakashima, S.; Nozawa, Y.; et al. Cloning and disruption of caPLB1, a phospholipase B gene involved in the pathogenicity of Candida albicans. J. Biol. Chem. 1998, 273, 26078–26086. [Google Scholar] [CrossRef]
- Mukherjee, P.K.; Seshan, K.R.; Leidich, S.D.; Chandra, J.; Cole, G.T.; Ghannoum, M.A. Reintroduction of the PLB1 gene into Candida albicans restores virulence in vivo. Microbiology 2001, 147, 2585–2597. [Google Scholar] [CrossRef]
- Heilmann, C.J.; Sorgo, A.G.; Siliakus, A.R.; Dekker, H.L.; Brul, S.; Koster, C.G.; de Koning, L.J.; Klis, F.M. Hyphal induction in the human fungal pathogen Candida albicans reveals a characteristic wall protein profile. Microbiology 2011, 157, 2297–2307. [Google Scholar] [CrossRef]
- Elamin Elhasan, L.M.; Hassan, M.B.; Elhassan, R.M.; Abdelrhman, F.A.; Salih, E.A.; Ibrahim, H.A.; Mohamed, A.A.; Osman, H.S.; Khalil, M.S.M.; Alsafi, A.A.; et al. Epitope-Based Peptide Vaccine Design against Fructose Bisphosphate Aldolase of Candida glabrata: An Immunoinformatics Approach. J. Immunol. Res. 2021, 2021, 1–19. [Google Scholar] [CrossRef]
- Xin, H. Active Immunizations with Peptide-DC Vaccines and Passive Transfer with Antibodies Protect Neutropenic Mice against Disseminated Candidiasis. Vaccine 2016, 34, 245–251. [Google Scholar] [CrossRef]
- Shi, H.; Dong, S.; Zhang, X.; Chen, X.; Gao, X.; Wang, L. Phage vaccines displaying YGKDVKDLFDYAQE epitope induce protection against systemic candidiasis in mouse model. Vaccine 2018, 36, 5717–5724. [Google Scholar] [CrossRef]
- Medrano-Díaz, C.L.; Vega-González, A.; Ruiz-Baca, E.; Moreno, A.; Cuéllar-Cruz, M. Moonlighting proteins induce protection in a mouse model against Candida species. Microb. Pathog. 2018, 124, 21–29. [Google Scholar] [CrossRef] [PubMed]
- Xin, H.; Glee, P.; Adams, A.; Mohiuddin, F.; Eberle, K. Design of a mimotope-peptide based double epitope vaccine against disseminated candidiasis. Vaccine 2019, 37, 2430–2438. [Google Scholar] [CrossRef] [PubMed]
- Adams, A.L.; Eberle, K.; Colón, J.R.; Courville, E.; Xin, H. Synthetic conjugate peptide Fba-Met6 (MP12) induces complement-mediated resistance against disseminated Candida albicans. Vaccine 2021, 39, 4099–4107. [Google Scholar] [CrossRef] [PubMed]
- Swoboda, R.K.; Bertram, G.; Hollander, H.; Greenspan, D.; Greenspan, J.S.; Gow, N.A.R.; Gooday, G.W.; Brown, A.J.P. Glycolytic enzymes of Candida albicans are nonubiquitous immunogens during candidiasis. Infect. Immun. 1993, 61, 4263–4271. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Arenas, E.; Molero, G.; Nombela, C.; Diez-Orejas, R.; Gil, C. Low virulent strains of Candida albicans: Unravelling the antigens for a future vaccine. Proteomics 2004, 4, 3007–3020. [Google Scholar] [CrossRef] [PubMed]
- Pitarch, A.; Díez-Orejas, R.; Molero, G.; Pardo, M.; Sánchez, M.; Gil, C.; Nombela, C. Analysis of the serologic response to systemic Candida albicans infection in a murine model. Proteomics 2001, 1, 550–559. [Google Scholar] [CrossRef]
- Schatzman, S.S.; Peterson, R.L.; Teka, M.; He, B.; Cabelli, D.E.; Cormack, B.P.; Culotta, V.C. Copper-only superoxide dismutase enzymes and iron starvation stress in Candida fungal pathogens. J. Biol. Chem. 2020, 295, 570–583. [Google Scholar] [CrossRef]
- Martchenko, M.; Alarco, A.M.; Harcus, D.; Whiteway, M. Superoxide dismutases in Candida albicans: Transcriptional regulation and functional characterization of the hyphal-induced SOD5 gene. Mol. Biol. Cell. 2004, 15, 456–467. [Google Scholar] [CrossRef] [PubMed]
- König, A.; Hube, B.; Kasper, L. The Dual Function of the Fungal Toxin Candidalysin during Candida albicans-Macrophage Interaction and Virulence. Toxins 2020, 12, 469. [Google Scholar] [CrossRef]
- Gil-Bona, A.; Llama-Palacios, A.; Parra, C.M.; Vivanco, F.; Nombela, C.; Monteoliva, L.; Gil, C. Proteomics unravels extracellular vesicles as carriers of classical cytoplasmic proteins in Candida albicans. J. Proteome Res. 2015, 14, 142–153. [Google Scholar] [CrossRef] [PubMed]
- Frohner, I.E.; Bourgeois, C.; Yatsyk, K.; Majer, O.; Kuchler, K. Candida albicans cell surface superoxide dismutases degrade host-derived reactive oxygen species to escape innate immune surveillance. Mol. Microbiol. 2009, 71, 240–252. [Google Scholar] [CrossRef] [PubMed]
- Robinett, N.G.; Culbertson, E.M.; Peterson, R.L.; Sanchez, H.; Andes, D.R.; Nett, J.E.; Culotta, V.C. Exploiting the vulnerable active site of a copper-only superoxide dismutase to disrupt fungal pathogenesis. J. Biol. Chem. 2019, 294, 2700–2713. [Google Scholar] [CrossRef] [PubMed]
- Aoki, W.; Ueda, T.; Tatsukami, Y.; Kitahara, N.; Morisaka, H.; Kuroda, K.; Ueda, M. Time-course proteomic profile of Candida albicans during adaptation to a fetal serum. Pathog. Dis. 2013, 67, 67–75. [Google Scholar] [CrossRef] [PubMed]
- Shibasaki, S.; Karasaki, M.; Aoki, W.; Ueda, M. Molecular and physiological study of Candida albicans by quantitative proteome analysis. Proteomes 2018, 6, 34. [Google Scholar] [CrossRef]
- Shibasaki, S.; Aoki, W.; Nomura, T.; Karasaki, M.; Sewaki, T.; Ueda, M. Evaluation of Mdh1 protein as an antigenic candidate for a vaccine against candidiasis. Biocontrol Sci. 2014, 19, 51–55. [Google Scholar] [CrossRef]
- Sui, X.; Yan, L.; Jiang, Y. ying The vaccines and antibodies associated with Als3p for treatment of Candida albicans infections. Vaccine 2017, 35, 5786–5793. [Google Scholar] [CrossRef]
- Chen, S.M.; Zou, Z.; Guo, S.Y.; Hou, W.T.; Qiu, X.R.; Zhang, Y.; Song, L.J.; Hu, X.Y.; Jiang, Y.Y.; Shen, H.; et al. Preventing Candida albicans from subverting host plasminogen for invasive infection treatment. Emerg. Microbes Infect. 2020, 9, 2417–2432. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| S.N | Candida Antigen | Experimental Vaccine | Implicated Humoral Mechanisms of Protection | References |
|---|---|---|---|---|
| 1. | Mannan | liposome-encapsulated mannan | agglutination of Candida cells, in vitro neutrophil candidacidal activity, complement aided protection | [100,113,138] |
| mannan extracts | anti-adhesion or anti-germ tube formation effects | [110] | ||
| mannan–BSA conjugate | antibody isotype switching | [117,138] | ||
| mannan-HSA conjugate | inhibition of C. albicans growth, B-cell immune-enhancement and antifungal activity | [123,130,131,141] | ||
| mannan oligosaccharide conjugates | complement fixation and complement mediated clearance of C. albicans, enhanced candidacidal activity, enhanced phagocytic activity, induction of respiratory burst of granulocytes, Immunomodulation and antibody isotype switching | [121,128,129,134,135,136,137,139,142] | ||
| 2. | β-glucan | β-glucan-conjugate vaccine | opsonization by enhanced phagocyte mediated extracellular hyphal killing, in vitro growth-inhibition of C. albicans, inhibition of hyphal growth, invasion and adherence. | [146,148,151,152,175] |
| β-glucan oligosaccharides plus keyhole limpet hemocyanin (KLH) | antibody class switching | [167,168] | ||
| linear β-(1→3)-nonaglucoside plus BSA (G9 | immunomodulation, candidacidal activity, inhibition of fungal growth adherence and dissemination, enhanced conidial phagocytosis. | [169,170] | ||
| OVA plus Curdlan | B-cell activation and enhancement of antibody response. | [172] | ||
| β-glucan | Induction of fungicidal activity of hemocytes. | [163,164] | ||
| 3. | Laminarin | Laminarin-CRM197 conjugate | enhanced opsonisation and phagocyte mediated hyphal killing, in vitro growth-inhibition of C. albicans, inhibition of hyphal growth, fungal invasion and adherence, biofilm inhibition, direct antifungal properties. | [146,148,152,154,175] |
| Laminarin conjugated with recombinant calreticulin fragment (LAM-CRT conjugate) | Inhibition of C. albicans growth in vitro. | [177] | ||
| β-mannan trisaccharide-(tetanus toxoid)-Laminarin tricomponent conjugate | immunomodulation of the immune response, isotype switching of IgG subclasses. | [165] | ||
| 4. | Hsp90 | recombinant Hsp90 protein | antibody mediated neutralization | [180,187,190,193,194] |
| Hybrid-phage displaying Hsp90 epitope | neutralization | [188,189] | ||
| proteoliposomal formulation of Hsp90 | immunomodulation and neutralization | [191,192] | ||
| Hsp90 expressing DNA vaccine | neutralization | [187] | ||
| 5. | Als3 | Recombinant Als3 protein | enhanced phagocyte recruitment and inflammatory cytokine production | [40,204,205,206,207,208,209] |
| Als3 with alum formulation (NDV-3A) | recruitment of phagocytes, enhanced neutrophil-mediated killing of C. albicans, enhanced opsonophagocytic killing by macrophages, interference with fungal adherence and invasion, inhibition of biofilm formation, reduction in hyphal elongation, inhibition of yeast dispersal from hyphal layers of biofilms. | [28,212,213,214,215,216,217] | ||
| 6. | Sap2 | recombinant Sap2 protein | Sap2-specific antibodies induced enzyme neutralization, inhibited inflammatory response, exhibited increased Candida biofilm inhibition ability in vitro, and enhanced neutrophil-mediated fungal killing | [29,111,230,231,232,233,235,238,239,240,241] |
| virosomal formulation of Sap2 (PEV-7) | Sap2 neutralization, inhibition of fungal adherence. | [236,237] | ||
| hybrid phage displaying Sap2 epitope SLAQVKYTSASSI | Induction of Sap2 specific antibody production, immunomodulation | [238] | ||
| epitope peptide of Sap2 displayed on virus nanofibers | Sap2 antibodies prevented fungal adhesion and colonization of C. albicans in the host. | [241] | ||
| 7. | Hyr1 | recombinant Hyr1 protein | opsonophagocytosis, enhanced neutrophil-mediated killing activity, inhibition of biofilm formation. | [90,149,244,245,246] |
| 8. | Hwp1 | Hwp1 glycopeptide conjugate | inhibition of biofilm formation, antibody-mediated growth inhibition of C. albicans | [103,132] |
| 9. | Enolase | recombinant enolase protein | enolase-specific IgG1 and IgG2a antibodies enhanced neutrophil-mediated killing of C. albicans, opsonophagocytosis and Induced complement activation. | [257,258,259,262] |
| Saccharomyces cerevisiae cells displaying enolase 1 | not determined | [262] | ||
| β-(Mannan trisaccharide)-Eno1 peptide conjugate vaccine | not determined | [103,132] | ||
| 10. | Fba1 | recombinant Fba1 protein | not determined | [273,274] |
| Fba peptide conjugated with β-1,2-mannotriose | complement activation and interference with Candida hyphal growth | [103,132,133] | ||
| Fba peptide pulsed DC formulation | not determined | [272] | ||
| Phage displaying Fba epitope | not determined | [273] | ||
| Fba peptide and mimotope conjugate | not determined | [275] | ||
| Fba-methionine synthase peptide construct (MP12) | phagocytosis, antibody-dependent complement activation | [276] | ||
| 11. | Pk | recombinant Pk protein | not determined | [274] |
| 12. | Mdh1 | recombinant Mdh1p protein | not determined | [288] |
| S.N. | Candida Antigen | Monoclonal Antibodies | Mechanisms of Protection | References |
|---|---|---|---|---|
| 1 | Mannan | B6.1 (IgM) | neutrophil mediated ingestion and killing of Candida | [108,109] |
| C3.1 (IgG) | complement fixation, enhanced phagocytosis and fungal killing | [118,119,120] | ||
| M1g1 (IgG) | complement activation and phagocytic killing of C. albicans | [125,126] | ||
| AF1 (IgM) | anticandidal protection in vaginitis | [111] | ||
| 2. | β-glucan | 3G11 (IgG1) | enhanced Candida phagocytosis, inhibition of fungal growth | [170] |
| 5H5 (IgG3) | cooperative Candida growth inhibitory activity with fluconazole | [170] | ||
| 2G8 (IgG2b) | inhibited Candida adherence and exhibited direct fungal growth inhibitory activity | [146,148,149] | ||
| 1E12 (IgM) | non-protective | [148] | ||
| 3. | Hsp90 | Mycograb (Efungumab) | exerted antifungal activity by inhibiting Hsp90 | [181,182,183] |
| 4. | Als3 | C7 (IgM) | inhibition of adherence, interference with fungal filamentation and exhibited direct candidacidal activity | [210] |
| 3D9.3 (IgM) | decreases adhesion of C. albicans germ tubes to epithelial and endothelial cells | [211] | ||
| 3-A5 (IgG1) | blocked adhesion of C. albicans to endothelial and epithelial cells. | [197,289] | ||
| 113 (IgG1 | blocked adhesion of C. albicans to endothelial and epithelial cells. | [197,289] | ||
| 5. | Sap2 | GF1 (IgG1) | protected against C. albicans vaginal challenge | [111] |
| NL2/2A8 (IgM) | rapid clearance of fungal cells from mucosal sites | [235] | ||
| NL2/9B9 (IgM) | rapid clearance of fungal cells from mucosal sites | [235] | ||
| 6. | Hwp1 | 6H1 (IgG2b) | induced significant protection against C. auris infection in vivo | [253] |
| 7. | Hyr1 | 17 human anti-Hyr mAbs (IgG1) | enhanced opsonophagocytosis of C. albicans and C. auris | [90] |
| H3 (IgM) | cross kingdom immune-protection against gram negative bacteria | [247] | ||
| H4 (IgM) | cross kingdom immune-protection against gram negative bacteria | [247] | ||
| 8. | Enolase | CaS1 (Single chain variable fragment) | inhibited the growth as well as plasminogen binding activity of C. albicans | [264] |
| 12D9 (IgG) | conferred antifungal activity by neutralizing Eno1, enhanced opsonophagocytosis and neutrophil mediated killing | [290] | ||
| 9. | Fba1 | E2-9 (IgM) | complement activation and interference with Candida hyphal growth | [103] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shukla, M.; Chandley, P.; Rohatgi, S. The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis. Vaccines 2021, 9, 1159. https://doi.org/10.3390/vaccines9101159
Shukla M, Chandley P, Rohatgi S. The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis. Vaccines. 2021; 9(10):1159. https://doi.org/10.3390/vaccines9101159
Chicago/Turabian StyleShukla, Manisha, Pankaj Chandley, and Soma Rohatgi. 2021. "The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis" Vaccines 9, no. 10: 1159. https://doi.org/10.3390/vaccines9101159
APA StyleShukla, M., Chandley, P., & Rohatgi, S. (2021). The Role of B-Cells and Antibodies against Candida Vaccine Antigens in Invasive Candidiasis. Vaccines, 9(10), 1159. https://doi.org/10.3390/vaccines9101159